
Effects of Clipping Intensity on the Physiology of Dicranopteris pedata and Its Interroot Soil in the Rare-Earth-Mining Area in Southern China
Abstract
:1. Introduction
2. Research Methodology
2.1. Study Area
2.2. Sampling and Pretreatment
2.3. Determination of D. pedata and Soil Physical and Chemical Indices
2.4. Soil Genomic DNA Extraction, PCR Amplification, and Sequencing
2.5. Data Processing and Statistical Analysis
3. Results
3.1. Effect of Clipping Intensity on the Growth and Physiology of D. pedata
3.1.1. Growth and Physiology Indices of D. pedata after Clipping
3.1.2. Correlation Analysis of the Physiological Indices of D. pedata
3.2. Effect of Clipping Intensity on Interroot Soil
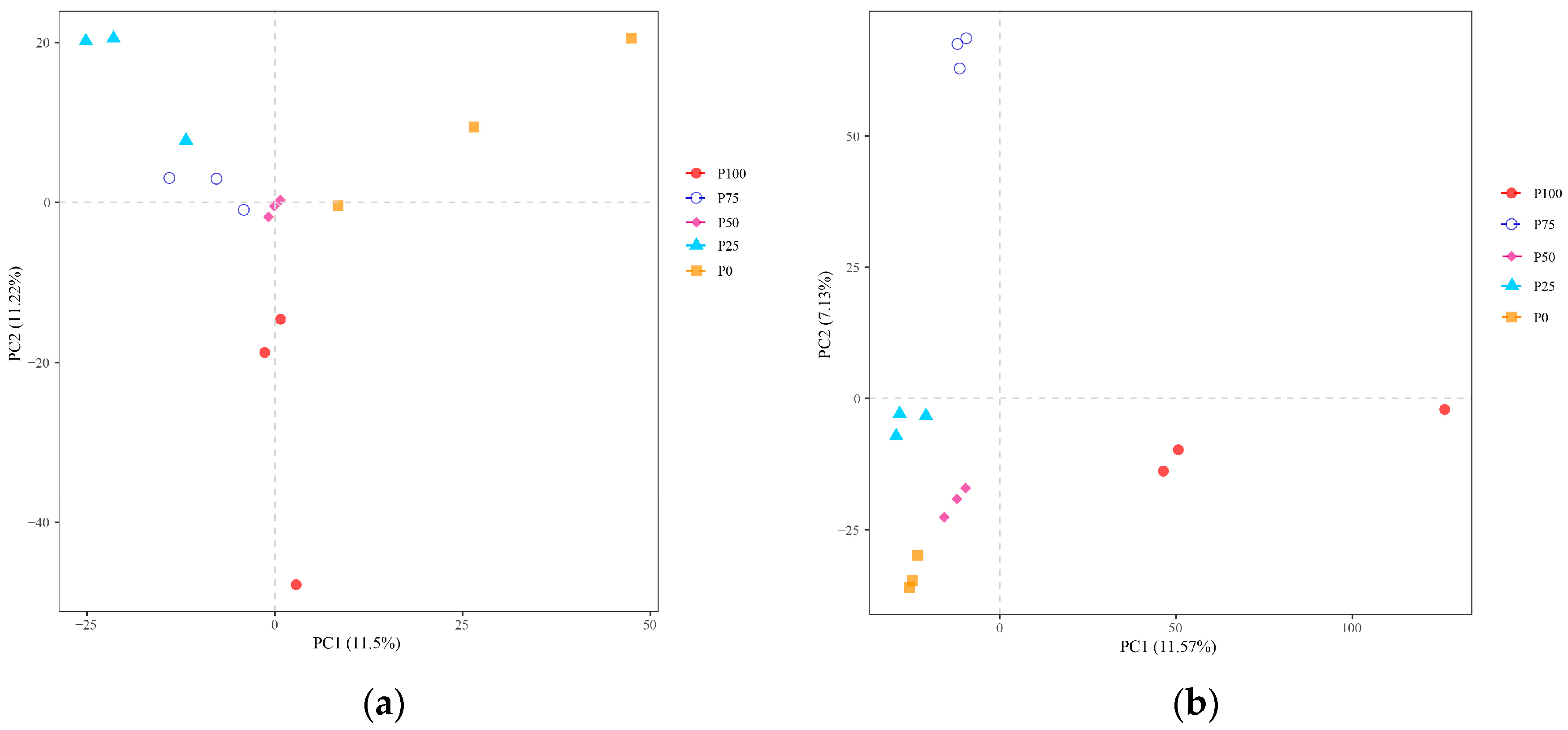
3.2.1. Interroot Soil Microbial Diversity Analysis of Different Clipping Intensities
3.2.2. Interroot Soil Enzyme Activities at Different Clipping Intensities
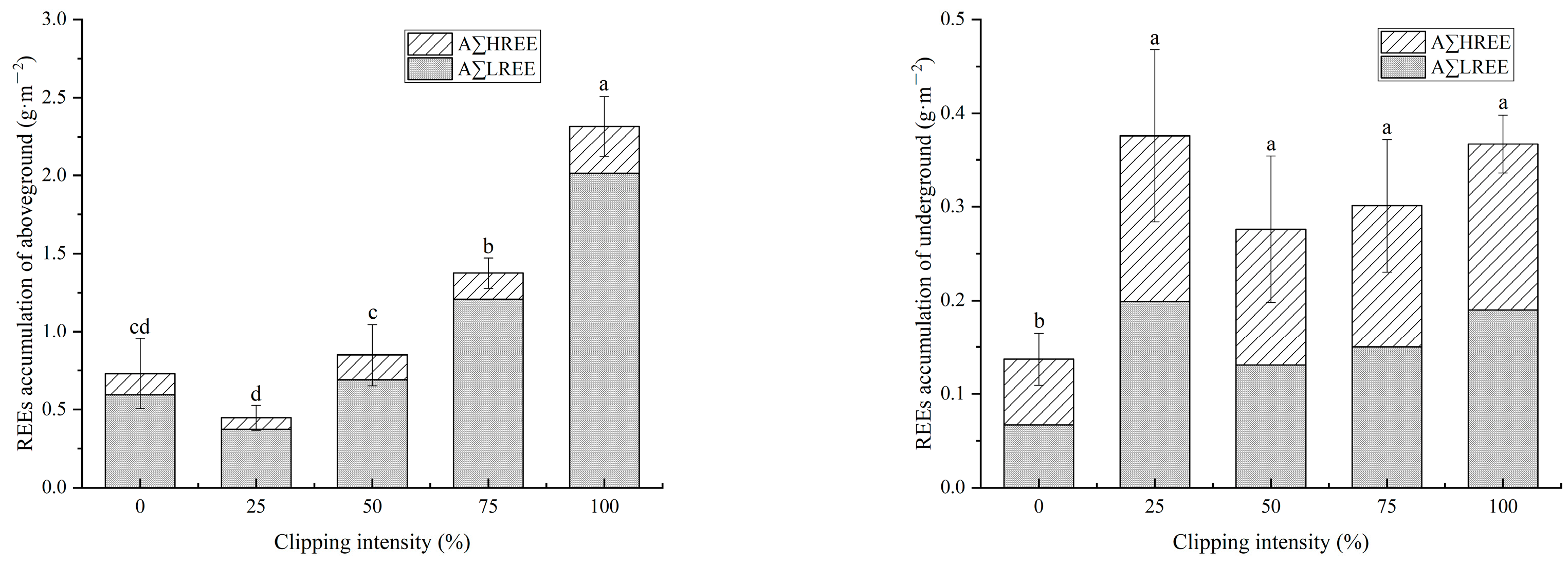
3.3. Phytoremediation of D. pedata after Clipping
4. Discussion
4.1. Effect of Clipping Intensity on the Growth and Physiology of D. pedata
4.2. Effects of Clipping on the Interroot Soil of D. pedata
4.3. Efficiency of Phytoremediation after Clipping D. pedata
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gupta, C.K.; Krishnamurthy, N. Extractive metallurgy of rare earths. Int. Mater. Rev. 1992, 37, 197–248. [Google Scholar] [CrossRef]
- Jordens, A.; Cheng, Y.P.; Waters, K.E. A review of the beneficiation of rare earth element bearing minerals. Miner. Eng. 2013, 41, 97–114. [Google Scholar] [CrossRef]
- Chakhmouradian, A.R.; Wall, F. Rare earth elements: Minerals, Mines, Magnets (and More). Elements 2012, 8, 333–340. [Google Scholar] [CrossRef]
- Chen, H.; Chen, Z.; Chen, Z.; Ou, X.; Chen, J. Calculation of toxicity coefficient of potential ecological risk assessment of rare earth elements. Bull. Environ. Contam. Toxicol. 2020, 104, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Celis, R.; Real, M.; Hermosin, M.C.; Cornejo, J. Sorption and leaching behaviour of polar aromatic acids in agricultural soils by batch and column leaching tests. Eur. J. Soil Sci. 2005, 56, 287–297. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, Z.; Bai, L. Rare earth element migration in gullies with different Dicranopteris dichotoma covers in the Huangnikeng gully group, Changting County, Southeast China. Chemosphere 2016, 164, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Gwenzi, W.; Mangori, L.; Danha, C.; Chaukura, N.; Dunjana, N.; Sanganyado, E. Sources, behaviour, and environmental and human health risks of high-technology rare earth elements as emerging contaminants. Sci. Total Environ. 2018, 636, 299–313. [Google Scholar] [CrossRef]
- Raskin, I.; Smith, S. Phytoremediation of metals: Using plants to remove pollutants from the environment. Curr. Opin. Biotechnol. 1997, 8, 221–226. [Google Scholar] [CrossRef]
- Mani, D.; Kumar, C. Biotechnological advances in bioremediation of heavy metals contaminated ecosystems: An overview with special reference to phytoremediation. Int. J. Environ. Sci. Technol. 2013, 11, 843–872. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals-Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Yang, Y.; Liang, Y.; Ghosh, A.; Song, Y.; Chen, H.; Tang, M. Assessment of arbuscular mycorrhizal fungi status and heavy metal accumulation characteristics of tree species in a lead-zinc mine area: Potential applications for phytoremediation. Environ. Sci. Pollut. Res. 2015, 22, 13179–13193. [Google Scholar] [CrossRef] [PubMed]
- Sekara, A.; Poniedzialek, M.; Ciura, J.; Jedrszczyk, E. Cadmium and lead accumulation and distribution in the organs of nine crops: Implications for phytoremediation. Pol. J. Environ. Stud. 2005, 14, 509–516. [Google Scholar]
- Tiodar, E.D.; Văcar, C.L.; Podar, D. Phytoremediation and microorganisms-Assisted phytoremediation of mercury-Contaminated soils: Challenges and Perspectives. Int. J. Environ. Res. Public Health 2021, 18, 2435. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, Z.; Liu, W.; Liu, S.; Zhang, L.; Zhong, L.; Luo, X.; Liang, H. Restoration of rare earth mine areas: Organic amendments and phytoremediation. Environ. Sci. Pollut. Res. 2015, 22, 17151–17160. [Google Scholar] [CrossRef]
- Massari, S.; Ruberti, M. Rare earth elements as critical raw materials: Focus on international markets and future strategies. Resour. Policy 2013, 38, 36–43. [Google Scholar] [CrossRef]
- Ali, H.; Naseer, M.; Sajad, M.A. Phytoremediation of heavy metals by Trifolium alexandrinum. Int. J. Environ. Sci. 2012, 2, 1459–1469. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, Z.; Yan, X.; Bai, L. Stoichiometric mechanisms of Dicranopteris dichotoma growth and resistance to nutrient limitation in the Zhuxi watershed in the red soil hilly region of China. Plant Soil 2015, 398, 367–379. [Google Scholar] [CrossRef]
- Wang, X.; Li, H.; Liu, Y.; Cui, T.; Liao, M.A.; Lin, L.; Wang, Z.; Deng, H.; Liang, D.; Xia, H.; et al. Mowing enhances the phytoremediation ability of cadmium-contaminated soil in the post-grafting generations of potential cadmium-hyperaccumulator Solanum photeinocarpum. Int. J. Phytoremediation 2021, 23, 652–657. [Google Scholar] [CrossRef]
- Sairu, M.; Xusheng, J.; Jie, L.; Zhifan, D.; Demin, D.; Shiqi, C.; Guo, Y. Mowing improves chromium phytoremediation in Leersia hexandra Swartz. Sustainability 2023, 15, 6244. [Google Scholar] [CrossRef]
- Ji, P.; Sun, T.; Song, Y.; Ackland, M.L.; Liu, Y. Strategies for enhancing the phytoremediation of cadmium-contaminated agricultural soils by Solanum nigrum L. Environ. Pollut. 2010, 159, 762–768. [Google Scholar] [CrossRef]
- Schrama, M.J.J.; Cordlandwehr, V.; Visser, E.J.W.; Elzenga, T.M.; de Vries, Y.; Bakker, J.P. Grassland cutting regimes affect soil properties, and consequently vegetation composition and belowground plant traits. Plant Soil 2012, 366, 401–413. [Google Scholar] [CrossRef]
- Ilmarinen, K.; Mikola, J. Soil feedback does not explain mowing effects on vegetation structure in a semi-natural grassland. Acta Oecologica 2009, 35, 838–848. [Google Scholar] [CrossRef]
- Yang, W.-J.; Gu, J.-F.; Zhou, H.; Huang, F.; Yuan, T.-Y.; Zhang, J.-Y.; Wang, S.-L.; Sun, Z.-G.; Yi, H.-W.; Liao, B.-H. Effect of three Napier grass varieties on phytoextraction of Cd- and Zn-contaminated cultivated soil under mowing and their safe utilization. Environ. Sci. Pollut. Res. 2020, 27, 16134–16144. [Google Scholar] [CrossRef] [PubMed]
- Kuzyakov, Y.; Xu, X. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef]
- Mencel, J.; Mocek-Płóciniak, A.; Kryszak, A. Soil microbial community and enzymatic activity of grasslands under different use practices: A review. Agronomy 2022, 12, 1136. [Google Scholar] [CrossRef]
- Fu, L.; Yan, Y.; Li, X.; Liu, Y.; Lu, X. Rhizosphere soil microbial community and its response to different utilization patterns in the semi-arid alpine grassland of northern Tibet. Front. Microbiol. 2022, 13, 931795. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Mi, X.; Shao, H.; Ma, K. Strong plant-soil associations in a heterogeneous subtropical broad-leaved forest. Plant Soil 2011, 347, 211–220. [Google Scholar] [CrossRef]
- Wang, X.K.; Huang, J.L. Principles and Techniques of Plant Physiology and Biochemistry Experiments; Higher Education Press: Beijing, China, 2015. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Renella, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasier, F.; Moscatelli, M.C.; Marinari, S. Soil enzymology: Classical and molecular approaches. Biol. Fertil. Soils 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2013, 30, 614–620. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 9, 13057. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Belsky, A.J.; Carson, W.P.; Jensen, C.L.; Fox, G.A. Overcompensation by plants: Herbivore optimization or red herring? Evol. Ecol. 1993, 7, 109–121. [Google Scholar] [CrossRef]
- Chen, Z.Q.; Chen, Z.B. Clipping strategy to assist phytoremediation by hyperaccumulator Dicranopteris dichotoma at rare earth mines. Int. J. Phytoremediation 2020, 22, 1038–1047. [Google Scholar] [CrossRef]
- Yang, L.; Huang, Y.; Lima, L.V.; Sun, Z.; Liu, M.; Wang, J.; Liu, N.; Ren, H. Rethinking the ecosystem functions of Dicranopteris, a widespread genus of ferns. Front. Plant Sci. 2020, 11, 581513. [Google Scholar] [CrossRef]
- Martin, F.M.; Dommanget, F.; Lavallée, F.; Evette, A. Clonal growth strategies of Reynoutria japonica in response to light, shade, and mowing, and perspectives for management. NeoBiota 2020, 56, 89–110. [Google Scholar] [CrossRef]
- Klimešová, J.; de Bello, F. CLO-PLA: The database of clonal and bud bank traits of Central European flora. J. Veg. Sci. 2009, 20, 511–516. [Google Scholar] [CrossRef]
- Wan, J.Z.; Wang, C.J.; Yu, F.H. Large-scale environmental niche variation between clonal and non-clonal plant species: Roles of clonal growth organs and ecoregions. Sci. Total Environ. 2018, 652, 1071–1076. [Google Scholar] [CrossRef]
- Suzuki, J.-I.; Stuefer, J. On the ecological and evolutionary significance of storage in clonal plants. Plant Species Biol. 1999, 14, 11–17. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Jiang, Y.X.; Zhu, X.D.; Chen, Z.Q.; Chen, Z.B.; Jiao, Y.J. Effect of short term high temperature stress on the physiological characteristics of Dicranoteris dichotoma. Subtrop. Soil Water Conserv. 2022, 34, 26–30. [Google Scholar]
- Song, K.L.; Zhu, X.D.; Chen, Z.Q.; Chen, Z.B.; Jiao, Y.J. Effects of short-term high temperature stress on photosynthetic characteristics of Dicranopteris dichotoma leaves. Subtrop. Soil Water Conserv. 2022, 34, 38–42. [Google Scholar]
- d’Aquino, L.; de Pinto, M.C.; Nardi, L.; Morgana, M.; Tommasi, F. Effect of some light rare earth elements on seed germination, seedling growth and antioxidant metabolism in Triticum durum. Chemosphere 2009, 75, 900–905. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.S.; Guo, C.R.; Cheng, Y.X. Mechanism of cerium ion clearing superoxide radical. J. Chin. Rare Earth Soc. 1997, 2, 55–58. [Google Scholar]
- Zhang, L.; Zeng, F.; Xiao, R. Effect of lanthanum ions (La3+) on the reactive oxygen species scavenging enzymes in wheat leaves. Biol. Trace Elem. Res. 2003, 91, 243–252. [Google Scholar] [CrossRef]
- Pang, X.; Wang, D.H.; Xing, X.Y.; Peng, A.; Zhang, F.S.; Li, C.J. Effect of La3+ on the activities of antioxidant enzymes in wheat seedlings under lead stress in solution culture. Chemosphere 2002, 47, 1033–1039. [Google Scholar] [CrossRef]
- Li, X.; Qiu, L.; Li, J.; Wang, H.; Wang, Y.; Xi, D. Role of jasmonate in Lolium perenne compensatory growth and photosynthesis: Uncoupling with photosynthesis and differential effects on growth. Acta Physiol. Plantarum. 2020, 42, 1–10. [Google Scholar] [CrossRef]
- Wei, Z.; Hong, F.; Yin, M.; Li, H.; Hu, F.; Zhao, G.; WoonchungWong, J. Subcellular and molecular localization of rare earth elements and structural characterization of yttrium bound chlorophyll a in naturally grown fern Dicranopteris dichotoma. Microchem. J. 2005, 80, 1–8. [Google Scholar] [CrossRef]
- Wang, X.P.; Shan, X.Q.; Zhang, S.Z.; Wen, B. Distribution of rare earth elements among chloroplast components of hyperaccumulator Dicranopteris dichotoma. Anal. Bioanal. Chem. 2003, 376, 913–917. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ji, B.; Hu, Y.; Liu, R.; Sun, W. A review on in situ phytoremediation of mine tailings. Chemosphere 2017, 184, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Schmalenberger, A.; O’Sullivan, O.; Gahan, J.; Cotter, P.D.; Courtney, R. Bacterial communities established in bauxite residues with different restoration histories. Environ. Sci. Technol. 2013, 47, 7110–7119. [Google Scholar] [CrossRef] [PubMed]
- Mesa, V.; Navazas, A.; González-Gil, R.; González, A.; Weyens, N.; Lauga, B.; Gallego, J.L.R.; Sánchez, J.; Peláez, A.I. Use of endophytic and rhizosphere bacteria to improve phytoremediation of arsenic-contaminated industrial soils by autochthonous Betula celtiberica. Appl. Environ. Microbiol. 2017, 83, e03411–e03416. [Google Scholar] [CrossRef]
- Bai, R.; Zhou, M.; Guo, Y.; Sheng, J.; Yuan, Y.; Li, Q.; Hou, L.; Bai, W. Long-term mowing reinforces connections between soil microbial and plant communities in a temperate steppe. Plant Soil 2022, 491, 177–190. [Google Scholar] [CrossRef]
- Chen, L.; Baoyin, T.; Minggagud, H. Effects of mowing regimes on above- and belowground biota in semi-arid grassland of northern China. J. Environ. Manag. 2021, 277, 111441. [Google Scholar] [CrossRef]
- Francioli, D.; van Rijssel, S.Q.; van Ruijven, J.; Termorshuizen, A.J.; Cotton, T.E.A.; Dumbrell, A.J.; Raaijmakers, J.M.; Weigelt, A.; Mommer, L. Plant functional group drives the community structure of saprophytic fungi in a grassland biodiversity experiment. Plant Soil 2020, 461, 91–105. [Google Scholar] [CrossRef]
- Maltz, M.R.; Treseder, K.K.; McGuire, K.L. Links between plant and fungal diversity in habitat fragments of coastal shrubland. PLoS ONE 2017, 12, e0184991. [Google Scholar] [CrossRef]
- Borowik, A.; Wyszkowska, J.; Kucharski, J.; Bacmaga, M.; Tomkiel, M. Response of microorganisms and enzymes to soil contamination with a mixture of terbuthylazine, mesotrione, and S-metolachlor. Environ. Sci. Pollut. Res. 2017, 24, 1910–1925. [Google Scholar] [CrossRef]
- Han, K.N.; Kellar, J.J.; Cross, W.M.; Safarzadeh, S. Opportunities and challenges for treating rare-earth elements. Geosystem Eng. 2014, 17, 178–194. [Google Scholar] [CrossRef]
- Qin, B.; Liu, W.; He, E.; Li, Y.; Liu, C.; Ruan, J.; Qiu, R.; Tang, Y. Vacuum pyrolysis method for reclamation of rare earth elements from hyperaccumulator Dicranopteris dichotoma grown in contaminated soil. J. Clean. Prod. 2019, 229, 480–488. [Google Scholar] [CrossRef]
- Chour, Z.; Laubie, B.; Morel, J.L.; Tang, Y.; Qiu, R.; Simonnot, M.-O.; Muhr, L. Recovery of rare earth elements from Dicranopteris dichotoma by an enhanced ion exchange leaching process. Chem. Eng. Process. Process Intensif. 2018, 130, 208–213. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Clipping Intensity | Control | ||||
|---|---|---|---|---|---|---|
| Sorts | 100 | 75 | 50 | 25 | 0 | |
| VC in 2022 (%) | 89 | 82 | 88 | 92 | 100 | |
| Compensation index | 1.82 ± 0.13 a | 1.58 ± 0.11 b | 1.53 ± 0.10 b | 1.53 ± 0.07 b | 0 c | |
| Physiological | CAT(U·mL−1) | 4.51 ± 0.83 a | 3.08 ± 0.67 b | 3.04 ± 0.33 b | 2.79 ± 0.23 b | 3.11 ± 0.57 b |
| SOD(U·L−1) | 1395.48 ± 106.82 b | 1445.36 ± 217.17 b | 1705.46 ± 345.88 ab | 2043.57 ± 517.62 a | 2151.00 ± 183.74 a | |
| POD(U·L−1) | 6.72 ± 0.99 b | 8.66 ± 0.69 b | 12.05 ± 2.71 a | 12.41 ± 1.68 a | 12.81 ± 1.42 a | |
| Chl(U·L−1) | 1.99 ± 0.32 a | 1.83 ± 0.13 a | 1.83 ± 0.06 a | 1.91 ± 0.09 a | 1.40 ± 0.25 b | |
| REEs (mg·kg−1) | Light | 1759.07 ± 126.64 a | 1617.97 ± 85.48 a | 1433.36 ± 339.09 a | 1542.07 ± 285.58 a | 935.65 ± 253.93 b |
| Heavy | 263.89 ± 23.17 ab | 228.22 ± 76.62 b | 328.60 ± 73.38 a | 307.42 ± 12.85 ab | 217.97 ± 33.53 b | |
| Total | 2022.96 ± 149.74 a | 1846.19 ± 122.16 a | 1761.96 ± 330.61 a | 1849.50 ± 272.75 a | 1153.63 ± 286.26 b | |
| Treatment | Clipping Intensity | Control | ||||
|---|---|---|---|---|---|---|
| Sorts | 100 | 75 | 50 | 25 | 0 | |
| Bacteria α diversity | Chao1 | 7721.46 ± 467.10 | 7814.18 ± 294.07 | 7315.51 ± 819.40 | 7504.08 ± 452.93 | 7152.80 ± 378.29 |
| Shannon | 9.00 ± 0.27 | 9.20 ± 0.26 | 8.81 ± 0.51 | 8.90 ± 0.33 | 8.42 ± 1.17 | |
| Fungi α diversity | Chao1 | 1027.43 ± 131.91 | 1067.73 ± 184.03 | 1062.44 ± 222.99 | 1092.75 ± 130.60 | 1060.84 ± 276.06 |
| Shannon | 4.36 ± 2.20 | 4.20 ± 1.02 | 3.75 ± 0.65 | 4.61 ± 0.71 | 3.47 ± 2.11 | |
| Treatment | Clipping Intensity | Control | ||||
|---|---|---|---|---|---|---|
| Sorts | 100 | 75 | 50 | 25 | 0 | |
| Soil enzyme activity | CAT(U·L−1) | 7193.64 ± 1094.37 | 6707.03 ± 839.43 | 8242.65 ± 1943.55 | 6867.15 ± 1317.66 | 8140.87 ± 440.15 |
| Urease(U·L−1) | 456.27 ± 102.34 b | 556.07 ± 55.62 ab | 563.08 ± 25.02 a | 633.36 ± 23.31 a | 648.42 ± 37.66 a | |
| Surase(U·L−1) | 921.25 ± 121.64 | 983.28 ± 33.60 | 966.40 ± 131.66 | 986.14 ± 126.04 | 1094.39 ± 134.64 | |
| Treatment | Clipping Intensity | Control | ||||
|---|---|---|---|---|---|---|
| Sorts | 100 | 75 | 50 | 25 | 0 | |
| Phytoextraction (g·m−2) | Light | 1.80 ± 0.21 a | 0.98 ± 0.08 b | 0.60 ± 0.15 c | 0.34 ± 0.08 d | 0 e |
| Heavy | 0.27 ± 0.03 a | 0.14 ± 0.04 b | 0.14 ± 0.04 b | 0.07 ± 0.01 c | 0 d | |
| Total | 2.07 ± 0.24 a | 1.12 ± 0.06 b | 0.74 ± 0.15 c | 0.41 ± 0.08 d | 0 e | |
| Phytoremediation time (years) | Light | 19.56 ± 1.69 c | 32.69 ± 3.19 bc | 59.52 ± 15.06 b | 108.74 ± 25.18 a | − |
| Heavy | 65.43 ± 5.95 c | 124.48 ± 38.65 b | 131.03 ± 41.25 b | 265.87 ± 8.75 a | − | |
| Total | 25.54 ± 2.22 c | 42.96 ± 3.16 bc | 71.62 ± 14.73 b | 134.79 ± 25.80 a | − | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.; Chen, Z.; Li, W.; Chen, Z. Effects of Clipping Intensity on the Physiology of Dicranopteris pedata and Its Interroot Soil in the Rare-Earth-Mining Area in Southern China. Sustainability 2024, 16, 664. https://doi.org/10.3390/su16020664
Lin Y, Chen Z, Li W, Chen Z. Effects of Clipping Intensity on the Physiology of Dicranopteris pedata and Its Interroot Soil in the Rare-Earth-Mining Area in Southern China. Sustainability. 2024; 16(2):664. https://doi.org/10.3390/su16020664
Chicago/Turabian StyleLin, Yu, Zhiqiang Chen, Weiye Li, and Zhibiao Chen. 2024. "Effects of Clipping Intensity on the Physiology of Dicranopteris pedata and Its Interroot Soil in the Rare-Earth-Mining Area in Southern China" Sustainability 16, no. 2: 664. https://doi.org/10.3390/su16020664