
A Lipidomic Profile of a Sustainable Source of Omega-3 Long-Chain Polyunsaturated Fatty Acids, Greenshell Mussels™, Perna canaliculus



Abstract
:1. Introduction
2. Materials and Methods
2.1. Greenshell Mussels (GSM) Sampling
2.2. Traditional Lipid Chemistry Analysis
2.3. Lipidomic Analysis
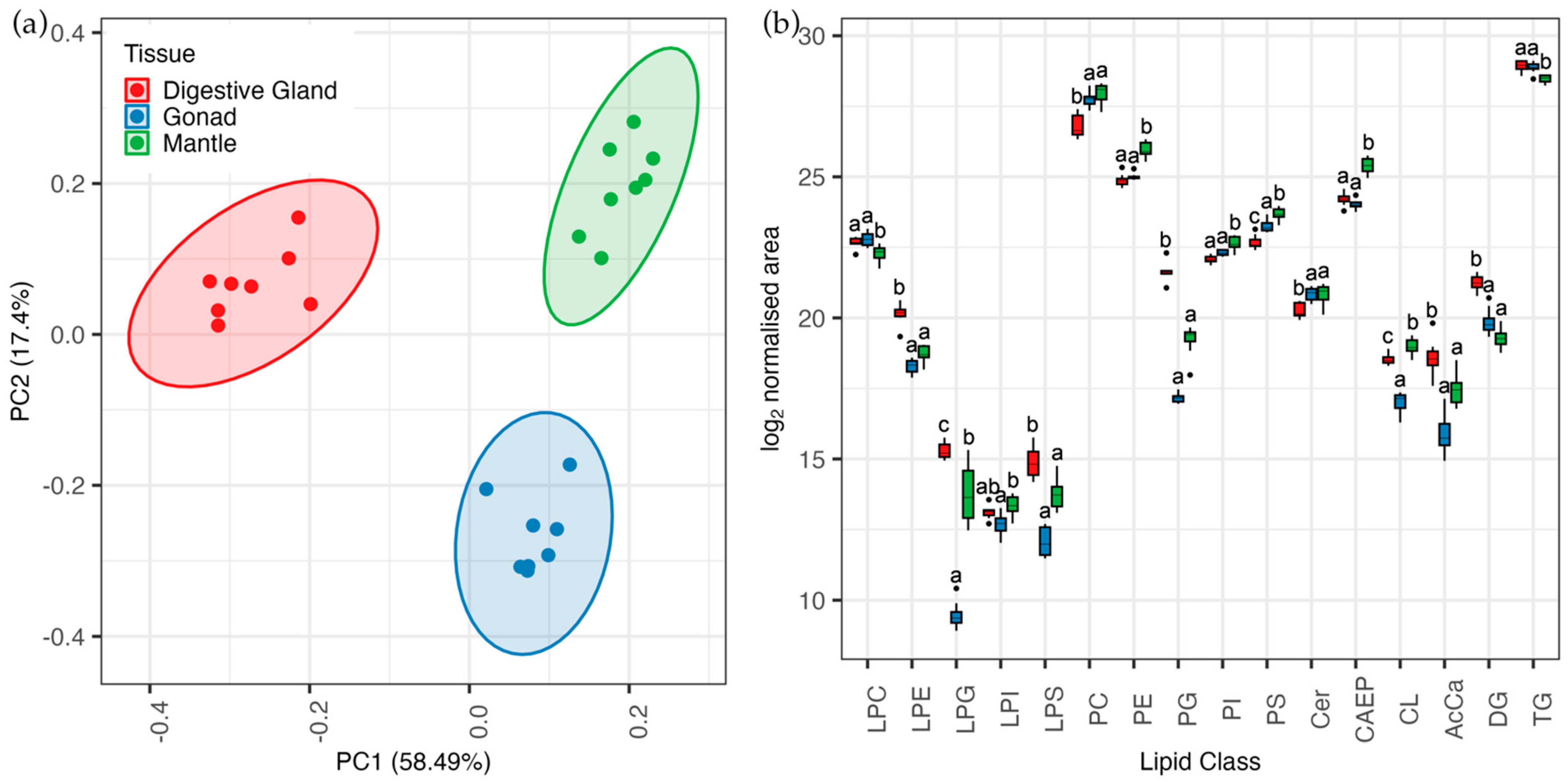
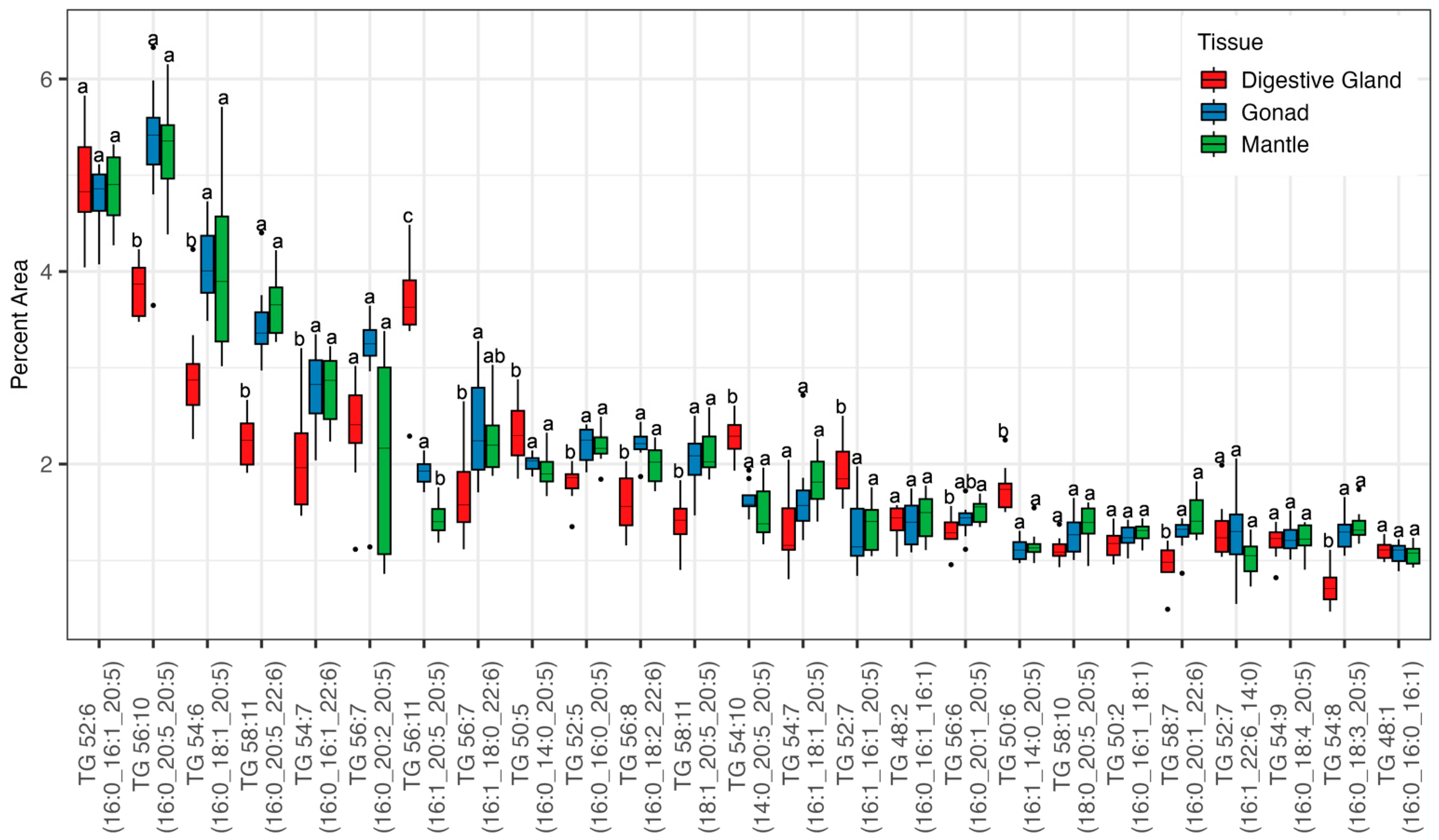
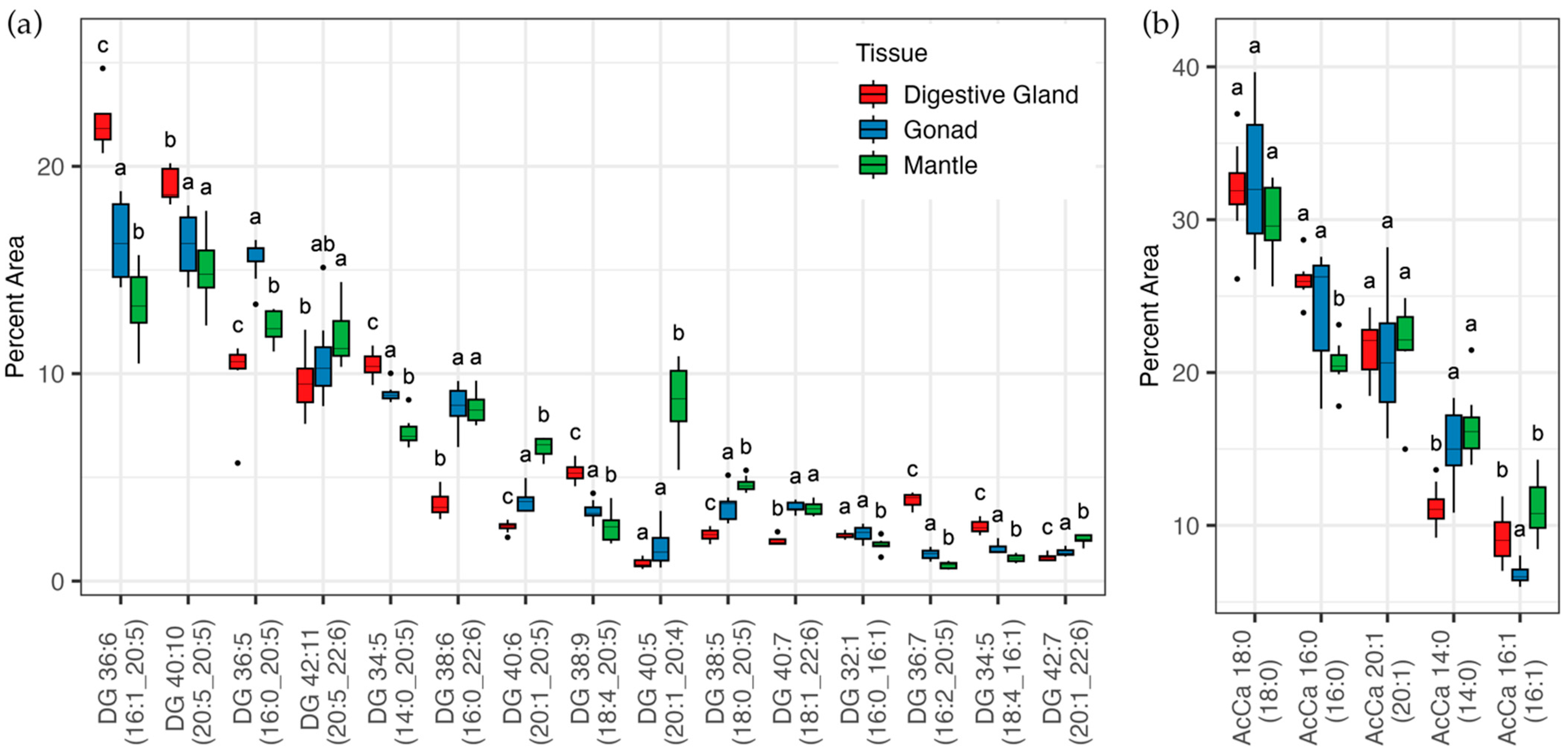
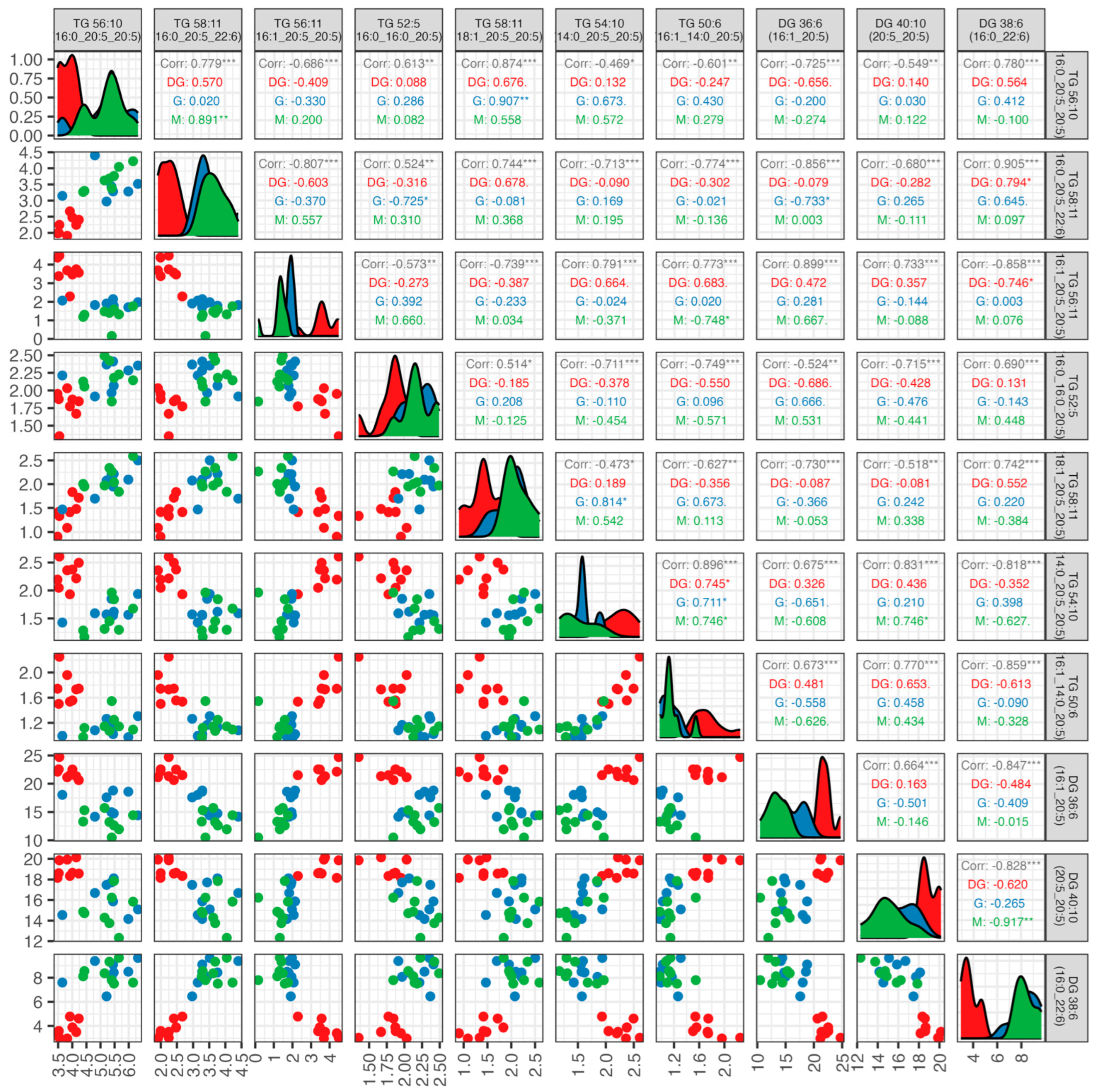
3. Results
3.1. Traditional Lipid Chemistry Analysis
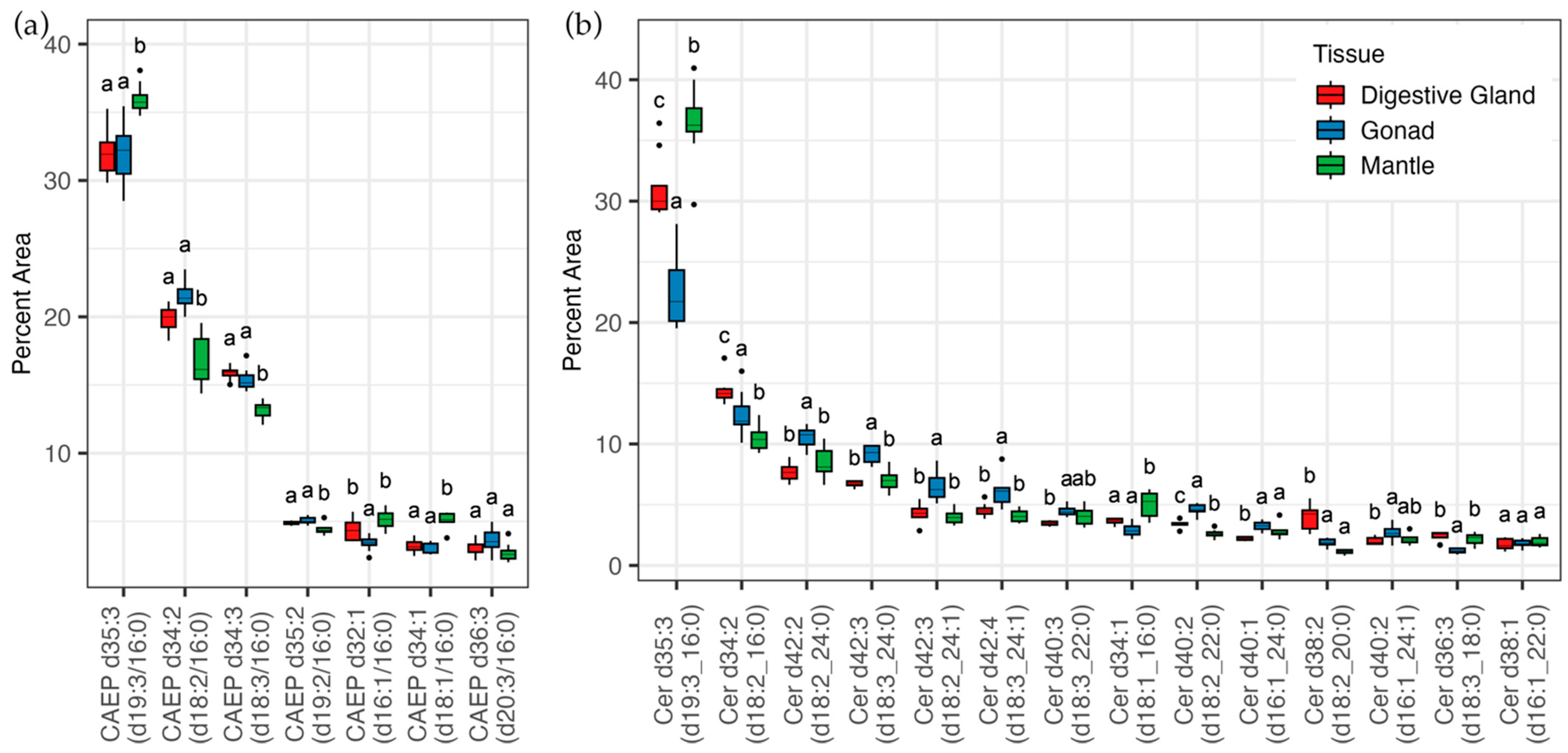
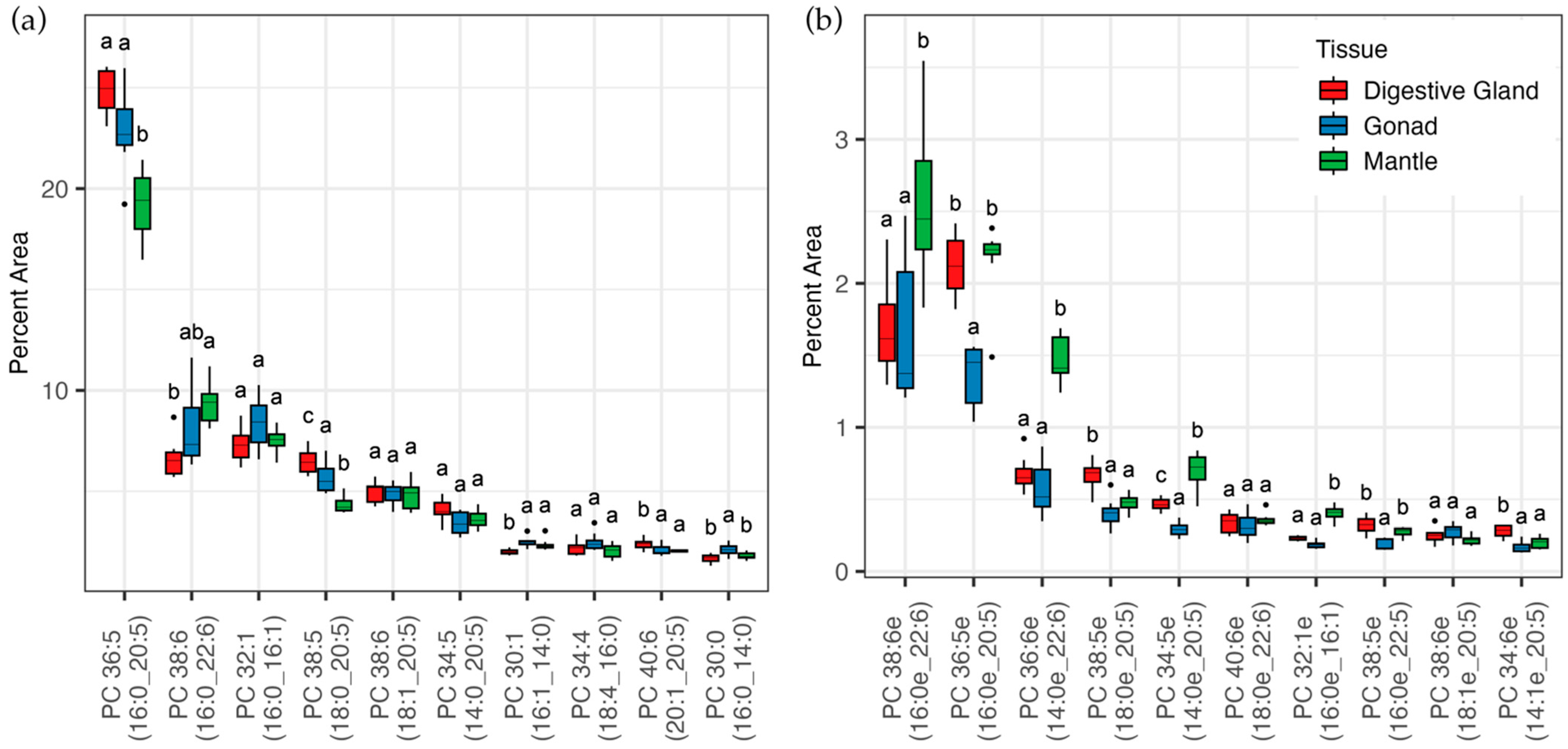
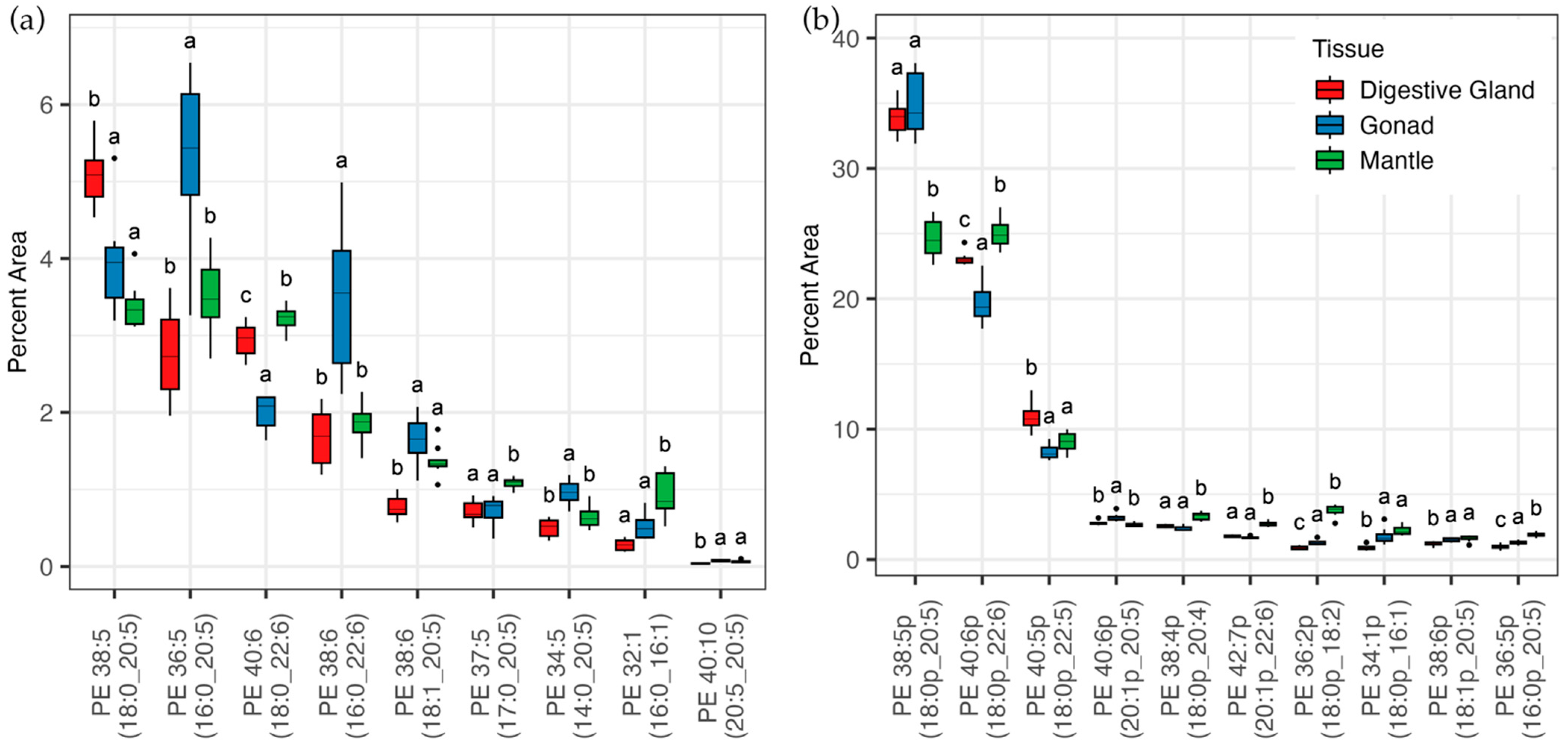
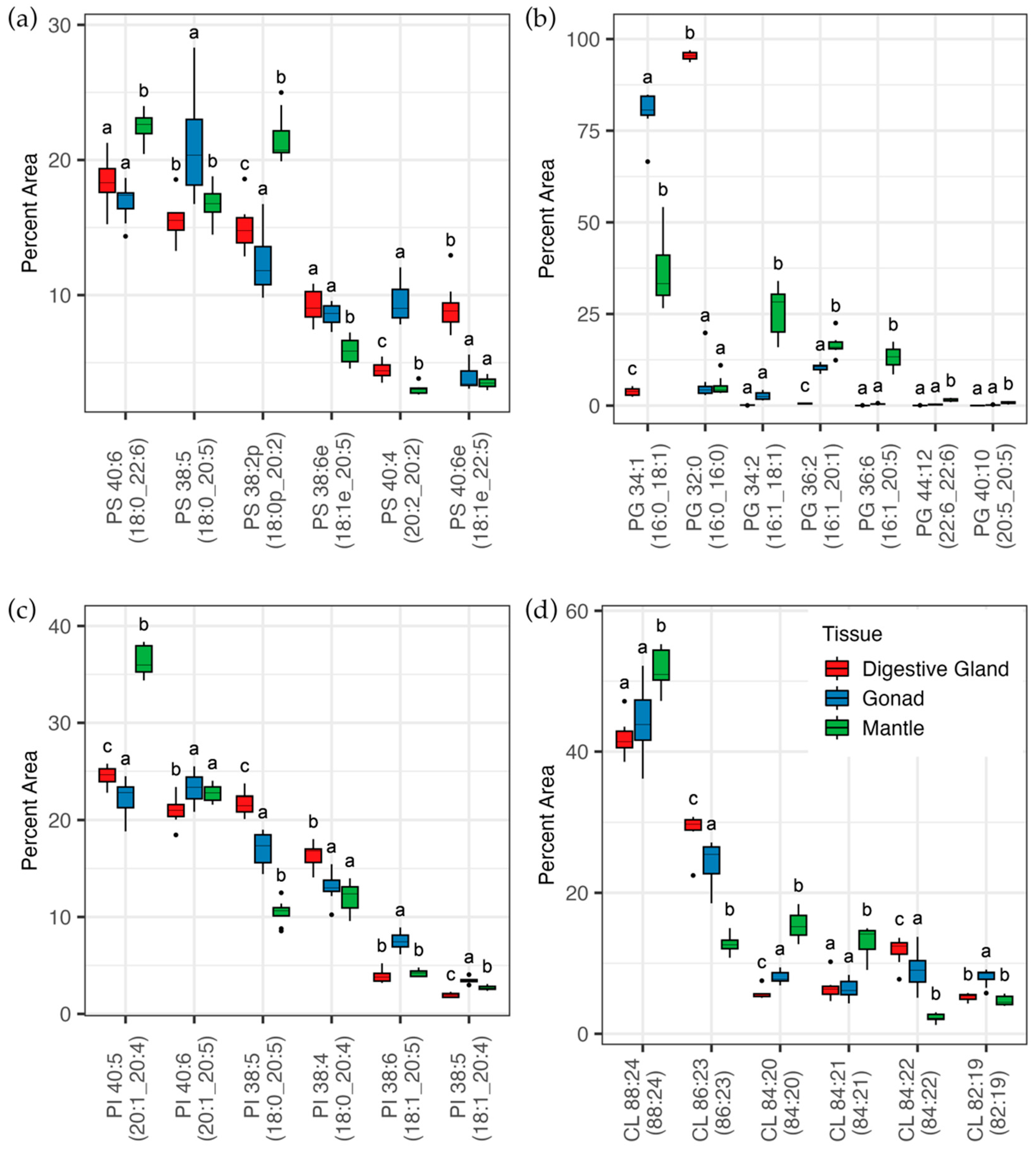
3.2. Lipidomic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aquaculture New Zealand. Mussel Export Statisticss; January 2022 to December 2022. Available online: https://www.aqua.org.nz/exports (accessed on 20 March 2023).
- Mickleborough, T.D.; Sinex, J.A.; Platt, D.; Chapman, R.F.; Hirt, M. The effects PCSO-524(R), a patented marine oil lipid and omega-3 PUFA blend derived from the New Zealand green lipped mussel (Perna canaliculus), on indirect markers of muscle damage and inflammation after muscle damaging exercise in untrained men: A randomized, placebo controlled trial. J. Int. Soc. Sports Nutr. 2015, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Wood, L.G.; Hazlewood, L.C.; Foster, P.S.; Hansbro, P.M. Lyprinol reduces inflammation and improves lung function in a mouse model of allergic airways disease. Clin. Exp. Allergy 2010, 40, 1785–1793. [Google Scholar] [CrossRef] [PubMed]
- Cobb, C.S.; Ernst, E. Systematic review of a marine nutriceutical supplement in clinical trials for arthritis: The effectiveness of the New Zealand green-lipped mussel Perna canaliculus. Clin. Rheumatol. 2006, 25, 275–284. [Google Scholar] [CrossRef]
- Gibson, S.L.M.; Gibson, R.G. The treatment of arthritis with a lipid extract of Perna canaliculus: A randomized trial. Complement. Ther. Med. 1998, 6, 122–126. [Google Scholar] [CrossRef]
- Cho, S.H.; Jung, Y.B.; Seong, S.C.; Park, H.B.; Byun, K.Y.; Lee, D.C.; Song, E.K.; Son, J.H. Clinical efficacy and safety of Lyprinol, a patented extract from New Zealand green-lipped mussel (Perna Canaliculus) in patients with osteoarthritis of the hip and knee: A multicenter 2-month clinical trial. Eur. Ann. Allergy Clin. Immunol. 2003, 35, 212–216. [Google Scholar]
- Lau, C.S.; Chiu, P.K.Y.; Chu, E.M.Y.; Cheng, I.Y.W.; Tang, W.M.; Man, R.Y.K.; Halpern, G.M. Treatment of knee osteoarthritis with Lyprinol®, lipid extract of the green-lipped mussel—A double-blind placebo-controlled study. Prog. Nutr. 2004, 6, 17–31. [Google Scholar]
- Szechinski, J.; Zawadzki, M. Measurement of pain relief resulting from the administration of Perna canaliculus lipid complex PCSO-524™ as compared to fish oil for treating patients who suffer from osteoarthritis of knee and/or hip joints. Reumatologia 2011, 49, 244–252. [Google Scholar]
- Abshirini, M.; Coad, J.; Wolber, F.M.; von Hurst, P.; Miller, M.R.; Tian, H.S.; Kruger, M.C. Effects of Greenshell™ mussel intervention on biomarkers of cartilage metabolism, inflammatory markers and joint symptoms in overweight/obese postmenopausal women: A randomized, double-blind, and placebo-controlled trial. Front. Med. 2022, 9, 1063336. [Google Scholar] [CrossRef]
- Miller, M.R.; Abshirini, M.; Wolber, F.M.; Tuterangiwhiu, T.R.; Kruger, M.C. Greenshell Mussel Products: A Comprehensive Review of Sustainability, Traditional Use, and Efficacy. Sustainability 2023, 15, 3912. [Google Scholar] [CrossRef]
- Miller, M.R.; Perry, N.; Burgess, E.; Marshall, S. Regiospecific analyses of triacylglycerols of Hoki (Macruronus novaezelandiae) and Greenshell™ mussel (Perna canaliculus). J. Am. Oil Chem. Soc. 2011, 88, 509–515. [Google Scholar] [CrossRef]
- Miller, M.; Pearce, L.; Bettjeman, B. Detailed distribution of lipids in Greenshell™ mussel (Perna canaliculus). Nutrients 2014, 6, 1454–1474. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.R.; Tian, H. Changes in proximate composition, lipid class and fatty acid profile in Greenshell™ mussels (Perna canaliculus) over an annual cycle. Aquac. Res. 2018, 49, 1153–1165. [Google Scholar] [CrossRef]
- Murphy, K.J.; Mooney, B.D.; Mann, N.J.; Nichols, P.D.; Sinclair, A.J. Lipid, FA, and sterol composition of New Zealand green lipped mussel (Perna canaliculus) and Tasmanian blue mussel (Mytilus edulis). Lipids 2002, 37, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Kabeya, N.; Fonseca, M.M.; Ferrier, D.E.K.; Navarro, J.C.; Bay, L.K.; Francis, D.S.; Tocher, D.R.; Castro, L.F.C.; Monroig, Ó. Genes for de novo biosynthesis of omega-3 polyunsaturated fatty acids are widespread in animals. Sci. Adv. 2018, 4, eaar6849. [Google Scholar] [CrossRef]
- Laudicella, V.A.; Whitfield, P.D.; Carboni, S.; Doherty, M.K.; Hughes, A.D. Application of lipidomics in bivalve aquaculture, a review. Rev. Aquac. 2019, 12, 678–702. [Google Scholar] [CrossRef]
- Siriarchavatana, P.; Kruger, M.C.; Miller, M.R.; Tian, H.S.; Wolber, F.M. The Preventive Effects of Greenshell Mussel (Perna canaliculus) on Early-Stage Metabolic Osteoarthritis in Rats with Diet-Induced Obesity. Nutrients 2019, 11, 1601. [Google Scholar] [CrossRef]
- Eason, C.T.; Adams, S.L.; Puddick, J.; Romanazzi, D.; Miller, M.R.; King, N.; Johns, S.; Forbes-Blom, E.; Hessian, P.A.; Stamp, L.K.; et al. Greenshell Mussels: A Review of Veterinary Trials and Future Research Directions. Vet. Sci. 2018, 5, 36. [Google Scholar] [CrossRef]
- Ulbricht, C.; Chao, W.; Costa, D.; Nguyen, Y.; Seamon, E.; Weissner, W. An Evidence-Based Systematic Review of Green-Lipped Mussel (Perna canaliculus) by the Natural Standard Research Collaboration. J. Diet. Suppl. 2009, 6, 54–90. [Google Scholar] [CrossRef]
- Buchanan, S. Measuring reproductive condition in the Greenshell (TM) mussel Perna canaliculus. N. Z. J. Mar. Freshw. Res. 2001, 35, 859–870. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Rampler, E.; Criscuolo, A.; Zeller, M.; El Abiead, Y.; Schoeny, H.; Hermann, G.; Sokol, E.; Cook, K.; Peake, D.A.; Delanghe, B.; et al. A Novel Lipidomics Workflow for Improved Human Plasma Identification and Quantification Using RPLC-MSn Methods and Isotope Dilution Strategies. Anal. Chem. 2018, 90, 6494–6501. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; Francois, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in H-1 NMR metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef] [PubMed]
- Lee-Chang, K.J.; Taylor, M.C.; Drummond, G.; Mulder, R.J.; Mansour, M.P.; Brock, M.; Nichols, P.D. Docosahexaenoic Acid Is Naturally Concentrated at the sn-2 Position in Triacylglycerols of the Australian Thraustochytrid Aurantiochytrium sp. Strain TC 20. Mar. Drugs 2021, 19, 382. [Google Scholar] [CrossRef]
- Giesbertz, P.; Ecker, J.; Haag, A.; Spanier, B.; Daniel, H. An LC-MS/MS method to quantify acylcarnitine species including isomeric and odd-numbered forms in plasma and tissues. J. Lipid Res. 2015, 56, 2029–2039. [Google Scholar] [CrossRef]
- Facchini, L.; Losito, I.; Cataldi, T.R.I.; Palmisano, F. Ceramide lipids in alive and thermally stressed mussels: An investigation by hydrophilic interaction liquid chromatography-electrospray ionization Fourier transform mass spectrometry. J. Mass Spectrom. 2016, 51, 768–781. [Google Scholar] [CrossRef]
- Warmerdam, S.; Vickers, J.; Palairet, N. Life Cycle Assessment of New Zealand Mussels and Oysters; Thinkstep Ltd. for Aquaculture New Zealand & Ministry of Primary Industries: Wellington, New Zealand, 2021. [Google Scholar]
- Hernandez, A.B.; Brumbaugh, R.D.; Frederick, P.; Grizzle, R.; Luckenbach, M.W.; Peterson, C.H.; Angelini, C. Restoring the eastern oyster: How much progress has been made in 53 years? Front. Ecol. Environ. 2018, 16, 463–470. [Google Scholar] [CrossRef]
- Bridger, D.; Attrill, M.J.; Davies, B.F.R.; Holmes, L.A.; Cartwright, A.; Rees, S.E.; Cabre, L.M.; Sheehan, E.V. The restoration potential of offshore mussel farming on degraded seabed habitat. Aquac. Fish Fish. 2022, 2, 437–449. [Google Scholar] [CrossRef]
- Willer, D.F.; Aldridge, D.C. Sustainable bivalve farming can deliver food security in the tropics. Nat. Food 2020, 1, 384–388. [Google Scholar] [CrossRef]
- Jónasdóttir, S.H. Fatty acid profiles and production in marine phytoplankton. Mar. Drugs 2019, 17, 151. [Google Scholar] [CrossRef]
- Zhou, C.; Song, H.; Feng, J.; Hu, Z.; Yang, M.J.; Shi, P.; Li, Y.R.; Guo, Y.J.; Li, H.Z.; Zhang, T. Metabolomics and biochemical assays reveal the metabolic responses to hypo-salinity stress and osmoregulatory role of cAMP-PKA pathway in Mercenaria mercenaria. Comput. Struct. Biotechnol. J. 2022, 20, 4110–4121. [Google Scholar] [CrossRef] [PubMed]
- Zhukova, N.V. Fatty Acids of Marine Mollusks: Impact of Diet, Bacterial Symbiosis and Biosynthetic Potential. Biomolecules 2019, 9, 857. [Google Scholar] [CrossRef] [PubMed]
- Laudicella, V.A.; Beveridge, C.; Carboni, S.; Franco, S.C.; Doherty, M.K.; Long, N.; Mitchell, E.; Stanley, M.S.; Whitfield, P.D.; Hughes, A.D. Lipidomics analysis of juveniles’ blue mussels (Mytilus edulis L. 1758), a key economic and ecological species. PLoS ONE 2020, 15, e0223031. [Google Scholar] [CrossRef]
- Yin, F.W.; Zhou, D.Y.; Zhao, Q.; Liu, Z.Y.; Hu, X.P.; Liu, Y.F.; Song, L.; Zhou, X.; Qin, L.; Zhu, B.W.; et al. Identification of glycerophospholipid molecular species of mussel (Mytilus edulis) lipids by high-performance liquid chromatography-electrospray ionization-tandem mass spectrometry. Food Chem. 2016, 213, 344–351. [Google Scholar] [CrossRef]
- Yu, X.M.; Doroghazi, J.R.; Janga, S.C.; Zhang, J.K.; Circello, B.; Griffin, B.M.; Labeda, D.P.; Metcalf, W.W. Diversity and abundance of phosphonate biosynthetic genes in nature. Proc. Natl. Acad. Sci. USA 2013, 110, 20759–20764. [Google Scholar] [CrossRef]
- Metcalf, W.W.; van der Donk, W.A. Biosynthesis of Phosphonic and Phosphinic Acid Natural Products. Annu. Rev. Biochem. 2009, 78, 65–94. [Google Scholar] [CrossRef] [PubMed]
- Kafarski, P. Phosphonates: Their Natural Occurrence and Physiological Role. In Contemporary Topics about Phosphorus in Biology and Materials; David, G.C., Maja Dutour, S., Božana, Č., Helga Füredi, M., Eds.; IntechOpen: Rijeka, Croatia, 2019; pp. 1–19. [Google Scholar]
- Bazan, S.; Mileykovskaya, E.; Mallampalli, V.K.P.S.; Heacock, P.; Sparagna, G.C.; Dowhan, W. Cardiolipin-dependent Reconstitution of Respiratory Supercomplexes from Purified Saccharomyces cerevisiae Complexes III and IV. J. Biol. Chem. 2013, 288, 401–411. [Google Scholar] [CrossRef]
- Paradies, G.; Paradies, V.; Ruggiero, F.M.; Petrosillo, G. Role of Cardiolipin in Mitochondrial Function and Dynamics in Health and Disease: Molecular and Pharmacological Aspects. Cells 2019, 8, 728. [Google Scholar] [CrossRef]
- Xu, Y.; Malhotra, A.; Ren, M.D.; Schlame, M. The enzymatic function of tafazzin. J. Biol. Chem. 2006, 281, 39217–39224. [Google Scholar] [CrossRef]
- Ahmmed, M.K.; Ahmmed, F.; Tian, H.; Carne, A.; Bekhit, A.E. Marine omega-3 (n-3) phospholipids: A comprehensive review of their properties, sources, bioavailability, and relation to brain health. Compr. Rev. Food Sci. Food Saf. 2020, 19, 64–123. [Google Scholar] [CrossRef]
- Fu, S.S.; Wen, M.; Zhao, Y.C.; Shi, H.H.; Wang, Y.M.; Xue, C.H.; Wei, Z.H.; Zhang, T.T. Short-term supplementation of EPA-enriched ethanolamine plasmalogen increases the level of DHA in the brain and liver of n-3 PUFA deficient mice in early life after weaning. Food Funct. 2022, 13, 1906–1920. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.J.; Wood, T.R. Why Have the Benefits of DHA Not Been Borne Out in the Treatment and Prevention of Alzheimer’s Disease? A Narrative Review Focused on DHA Metabolism and Adipose Tissue. Int. J. Mol. Sci. 2021, 22, 11826. [Google Scholar] [CrossRef] [PubMed]
- AbuMweis, S.; Abu Omran, D.; Al-Shami, I.; Jew, S. The ratio of eicosapentaenoic acid to docosahexaenoic acid as a modulator for the cardio-metabolic effects of omega-3 supplements: A meta-regression of randomized clinical trials. Complement. Ther. Med. 2021, 57, 102662. [Google Scholar] [CrossRef] [PubMed]
- Kariotoglou, D.M.; Mastronicolis, S.K. Phosphonolipids in the mussel Mytilus galloprovincialis. Z. Nat. C 1998, 53, 888–896. [Google Scholar] [CrossRef]
- Tomonaga, N.; Manabe, Y.; Sugawara, T. Digestion of Ceramide 2-Aminoethylphosphonate, a Sphingolipid from the Jumbo Flying Squid Dosidicus gigas, in Mice. Lipids 2017, 52, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Norris, G.H.; Blesso, C.N. Dietary and Endogenous Sphingolipid Metabolism in Chronic Inflammation. Nutrients 2017, 9, 1180. [Google Scholar] [CrossRef]
- Suh, J.H.; Saba, J.D. Sphingosine-1-phosphate in inflammatory bowel disease and colitis-associated colon cancer: The fat’s in the fire. Transl. Cancer Res. 2015, 4, 469–483. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mantle | Digestive Gland | Gonad | Mantle | Digestive Gland | Gonad | |
|---|---|---|---|---|---|---|
| Composition | Whole tissue | Extracted oil | ||||
| Moisture (g/100 g of Organ) | 81.2 | 72.5 | 68.9 | NA | NA | NA |
| Lipid (g/100 g of Organ) | 1.6 | 4.2 | 4.8 | NA | NA | NA |
| Lipid (g/100 g of GSM) | 0.28 | 0.45 | 0.77 | NA | NA | NA |
| Proportion of organ in the Mussel | 16.1 | 17.7 | 10.7 | NA | NA | NA |
| Lipid class content | mg/100 g of GSM | g/100 g of oil | ||||
| Hydrocarbon/nonpolar lipids | 1.9 | 1.9 | 2.1 | 0.7 | 0.4 | 0.3 |
| Free Fatty Acids | 2.7 | 4.0 | 1.3 | 1.0 | 0.9 | 0.2 |
| Triacylglycerols | 27.2 | 138.7 | 219.8 | 9.8 | 31.0 | 28.4 |
| Sterols | 6.7 | 5.4 | 9.2 | 2.4 | 1.2 | 1.2 |
| Polar lipids | 238.2 | 297.3 | 541.9 | 86.2 | 66.4 | 70.1 |
| Fatty acid content (mg/100 g of GSM) | mg/100g of GSM | % Fatty acids | ||||
| C14:0 myristic acid | 9.3 | 16.9 | 30.9 | 4.2 | 4.7 | 5.0 |
| C15:0 pentadecanoic acid | 1.2 | 1.4 | 3.2 | 0.6 | 0.4 | 0.5 |
| C16:0 palmitic acid | 32.6 | 49.4 | 98.7 | 14.7 | 13.8 | 16.0 |
| C16:1 palmitoleic acid | 16.2 | 36.7 | 55.8 | 7.3 | 10.3 | 9.0 |
| C16:2n4 hexadecadienoic acid | 1.3 | 4.2 | 4.9 | 0.6 | 1.2 | 0.8 |
| C17:0 heptadecanoic acid | 1.5 | 1.8 | 4.0 | 0.7 | 0.5 | 0.6 |
| C18:0 stearic acid | 8.7 | 13.6 | 22.7 | 3.9 | 3.8 | 3.7 |
| C18:1n7 vaccenic acid | 6.0 | 9.5 | 19.5 | 2.7 | 2.7 | 3.2 |
| C18:1n9c oleic acid | 2.6 | 4.9 | 6.8 | 1.2 | 1.4 | 1.1 |
| C18:1t elaidic acid | 0.2 | 0.2 | 0.9 | 0.1 | 0.1 | 0.1 |
| C18:2n6c linoleic acid | 3.8 | 4.9 | 9.3 | 1.7 | 1.4 | 1.5 |
| C18:3n3 alpha linolenic acid (ALA) | 1.4 | 2.4 | 4.9 | 0.7 | 0.7 | 0.8 |
| C18:3n6 gamma linolenic (GLA) | 0.3 | 0.6 | 1.3 | 0.1 | 0.2 | 0.2 |
| C18:3n4 octadecatrienoic acid | 4.1 | 5.0 | 10.4 | 1.9 | 1.4 | 1.7 |
| C18:4n3 stearidonic acid (SDA) | 3.9 | 9.1 | 16.3 | 1.8 | 2.5 | 2.6 |
| C20:0 arachidic acid | 0.1 | 0.3 | 0.3 | 0.0 | 0.1 | 0.0 |
| C20:1 gadoleic acid | 0.9 | 0.9 | 1.4 | 0.4 | 0.3 | 0.2 |
| C20:3n3 cis-11, 14, 17-eicosatrienoic acid | 0.1 | 0.5 | 0.8 | 0.0 | 0.1 | 0.1 |
| C20:3n6 cis-8, 11, 14-eicosatrienoic acid | 0.6 | 1.2 | 2.5 | 0.3 | 0.3 | 0.4 |
| C20:4n3 eicosatetraenoic acid | 0.6 | 1.9 | 2.3 | 0.3 | 0.5 | 0.4 |
| C20:4n6 arachidonic acid (AA) | 4.5 | 5.3 | 8.2 | 2.1 | 1.5 | 1.3 |
| C20:5n3 eicosapentaenoic acid (EPA) | 45.5 | 94.9 | 157.9 | 20.6 | 26.5 | 25.5 |
| C22:5n3 docosapentaenoic acid (DPA) | 3.7 | 6.7 | 9.1 | 1.7 | 1.9 | 1.5 |
| C22:6n3 docosahexaenoic acid (DHA) | 28.2 | 30.7 | 67.0 | 12.8 | 8.6 | 10.8 |
| Fatty acid classes | mg/100 g of GSM | % Fatty acids | ||||
| ∑SFA | 53.6 | 83.7 | 160.1 | 24.2 | 23.4 | 25.9 |
| ∑MUFA | 26.0 | 52.2 | 84.4 | 11.8 | 14.6 | 13.6 |
| ∑PUFA | 98.2 | 167.3 | 294.6 | 44.4 | 46.7 | 47.6 |
| ∑n-3 PUFA | 83.5 | 146.1 | 258.1 | 37.8 | 40.8 | 41.7 |
| ∑n-6 PUFA | 9.3 | 12.0 | 21.2 | 4.2 | 3.4 | 3.4 |
| Other Fatty acids | mg/100 g of GSM | mg/100 g of GSM | ||||
| 16:0 FALD | 0.6 | 0.6 | 0.9 | 0.3 | 0.2 | 0.2 |
| 18:0 FALD | 13.5 | 13.6 | 15.6 | 6.1 | 3.8 | 2.5 |
| 16:0 OH | 1.8 | 0.6 | 8.1 | 0.8 | 0.2 | 1.3 |
| 20:2 NMI | 3.5 | 5.7 | 10.8 | 1.6 | 1.6 | 1.7 |
| 22:2 NMI | 4.2 | 4.8 | 7.4 | 1.9 | 1.3 | 1.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taylor, M.C.; Roberts, R.D.; Miller, M.R. A Lipidomic Profile of a Sustainable Source of Omega-3 Long-Chain Polyunsaturated Fatty Acids, Greenshell Mussels™, Perna canaliculus. Sustainability 2023, 15, 7586. https://doi.org/10.3390/su15097586
Taylor MC, Roberts RD, Miller MR. A Lipidomic Profile of a Sustainable Source of Omega-3 Long-Chain Polyunsaturated Fatty Acids, Greenshell Mussels™, Perna canaliculus. Sustainability. 2023; 15(9):7586. https://doi.org/10.3390/su15097586
Chicago/Turabian StyleTaylor, Matthew C., Rodney D. Roberts, and Matthew R. Miller. 2023. "A Lipidomic Profile of a Sustainable Source of Omega-3 Long-Chain Polyunsaturated Fatty Acids, Greenshell Mussels™, Perna canaliculus" Sustainability 15, no. 9: 7586. https://doi.org/10.3390/su15097586