
Transporting Tenebrio molitor Eggs: The Effect of Temperature, Humidity and Time on the Hatch Rate

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Colony Information
2.2. Egg Transport Experiments
2.3. Statistical Analysis
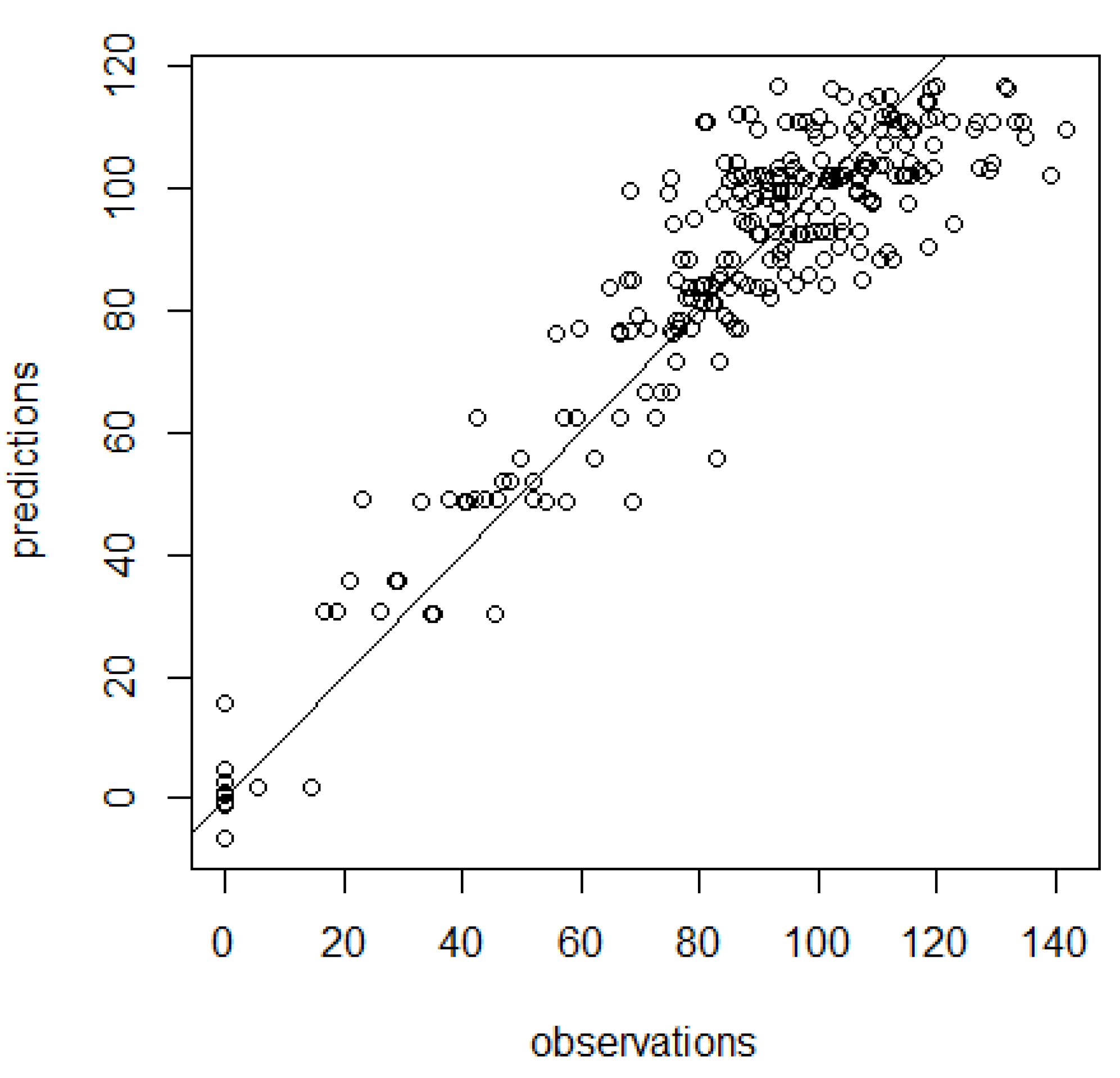
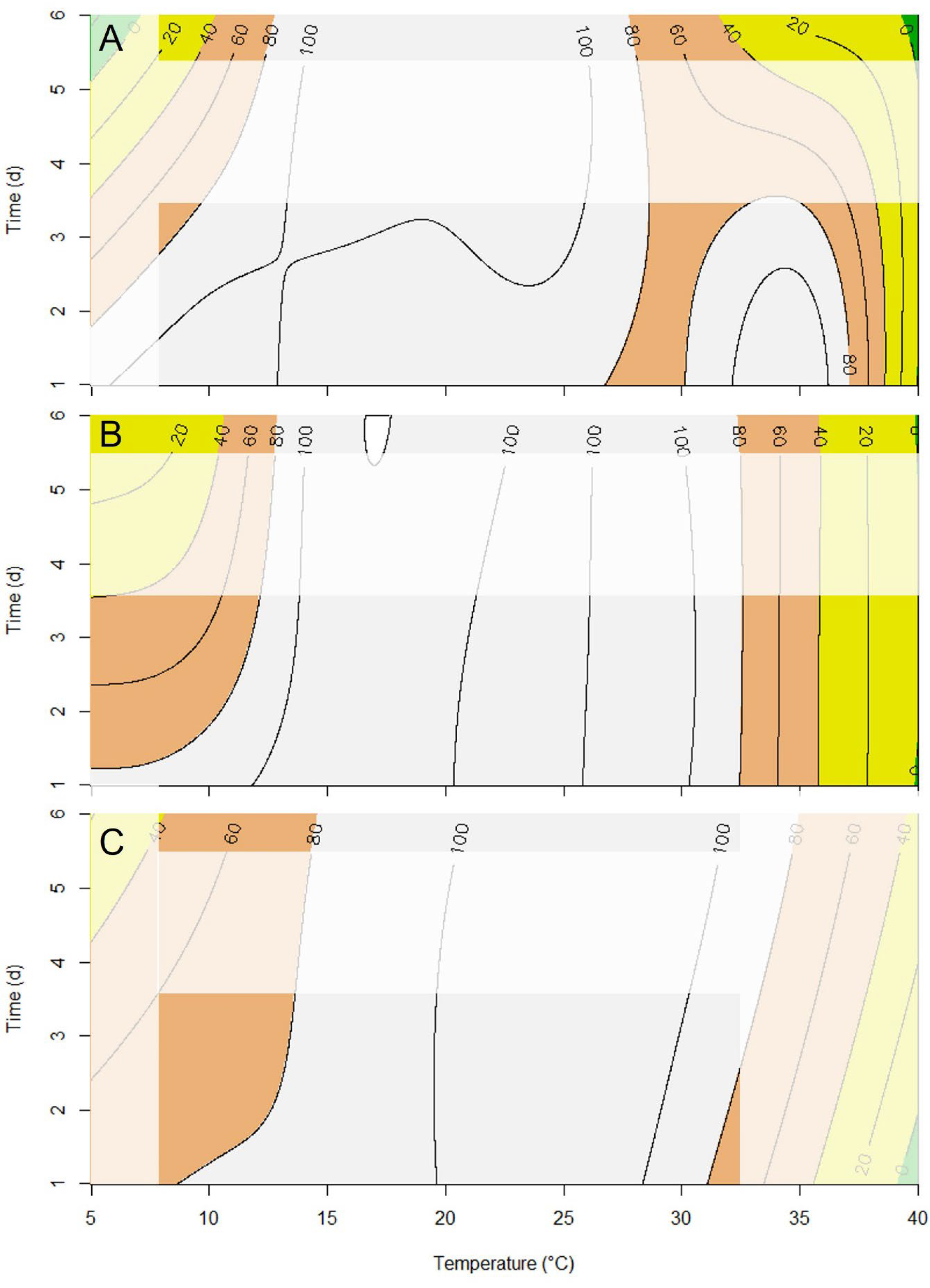
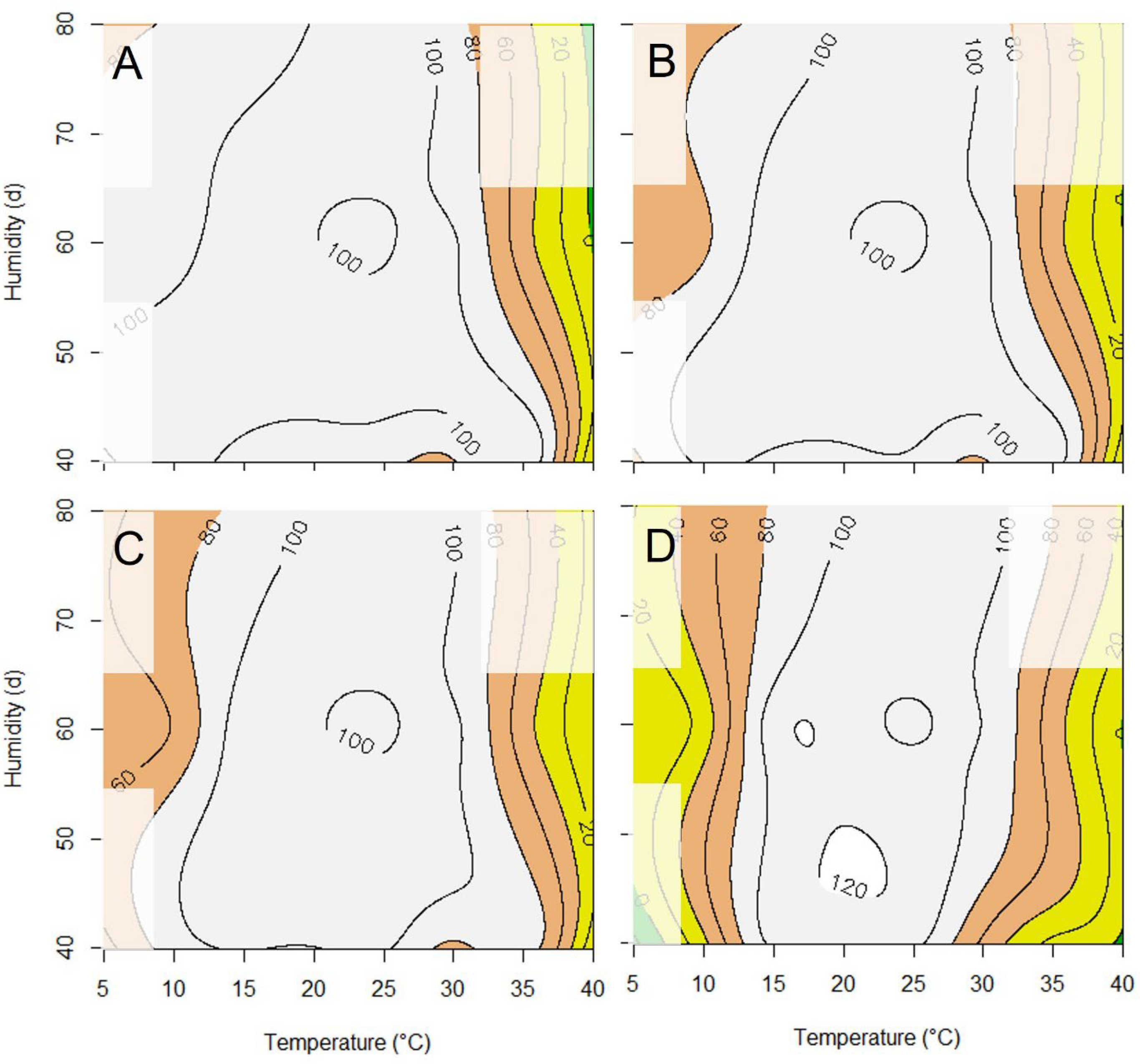
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mansor, M.; Rahman, U.A.; Abidin, M.Z.; Zain, M.; Yusof, M. Variation of Car Cabin Temperature Influenced by Ventilation under Direct Sun Exposure. J. Mech. Eng. Sci. 2014, 6, 1014–1023. [Google Scholar]
- Greenberg, S.; Ar, A. Effects of chronic hypoxia, normoxia and hyperoxia on larval development in the beetle Tenebrio molitor. J. Insect Physiol. 1996, 42, 991–996. [Google Scholar] [CrossRef]
- Punzo, F.; Mutchmor, J.A. Effects of temperature, relative humidity and period of exposure on the survival capacity of Tenebrio molitor (Coleoptera: Tenebrionidae). J. Kans. Entomol. Soc. 1980, 53, 260–270. [Google Scholar]
- Errico, S.; Dimatteo, S.; Moliterni, S.; Baldacchino, F. Effects of long-lasting cold storage on Tenebrio molitor larvae (Coleoptera: Tenebrionidae). J. Insects Food Feed. 2021, 7, 1111–1116. [Google Scholar] [CrossRef]
- Dreyer, M.; Hörtenhuber, S.; Zollitsch, W.; Jäger, H.; Schaden, L.-M.; Gronauer, A.; Kral, I. Environmental life cycle assessment of yellow mealworm (Tenebrio molitor) production for human consumption in Austria—A comparison of mealworm and broiler as protein source. Int. J. Life Cycle Assess. 2021, 26, 2232–2247. [Google Scholar] [CrossRef]
- Punzo, F.; Mutchmor, J.A. Changes in temperature tolerance as a function of relative humidity during the larval stage of Tenebrio molitor (Coleoptera: Tenebrionidae). J. Kans. Entomol. Soc. 1978, 51, 207–212. [Google Scholar]
- Allen, J.L.; Clusella-Trullas, S.; Chown, S.L. The effects of acclimation and rates of temperature change on critical thermal limits in Tenebrio molitor (Tenebrionidae) and Cyrtobagous salviniae (Curculionidae). J. Insect Physiol. 2012, 58, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Rebaudo, F.; Rabhi, V.B. Modeling temperature-dependent development rate and phenology in insects: Review of major developments, challenges, and future directions. Entomol. Exp. Appl. 2018, 166, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Coudron, C.; Deruytter, D.; Craeye, S.; Bleyaert, P. Entomoponics: Combining insect rearing and greenhouse vegetable production—A case study with Tenebrio molitor and high-wire cucumber cultivation. J. Insects Food Feed. 2022, 8, 427–437. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deruytter, D.; Coudron, C.L.; Claeys, J. Transporting Tenebrio molitor Eggs: The Effect of Temperature, Humidity and Time on the Hatch Rate. Sustainability 2023, 15, 6231. https://doi.org/10.3390/su15076231
Deruytter D, Coudron CL, Claeys J. Transporting Tenebrio molitor Eggs: The Effect of Temperature, Humidity and Time on the Hatch Rate. Sustainability. 2023; 15(7):6231. https://doi.org/10.3390/su15076231
Chicago/Turabian StyleDeruytter, David, Carl L. Coudron, and Jonas Claeys. 2023. "Transporting Tenebrio molitor Eggs: The Effect of Temperature, Humidity and Time on the Hatch Rate" Sustainability 15, no. 7: 6231. https://doi.org/10.3390/su15076231