
Habitat Modification Alters Food Web Interactions with Focus on Biological Control of Aphids in Apple Orchards

Abstract
:1. Introduction
2. Materials and Methods
2.1. Unconnected Patches
2.1.1. Study Site
2.1.2. Population Dynamic
2.2. Connected Patches: Intercropping Experiment
2.2.1. Experimental Design
2.2.2. Sampling
2.3. Data Analysis
2.3.1. Food Web
2.3.2. Statistical Analyses
3. Results
3.1. Unconnected Patches
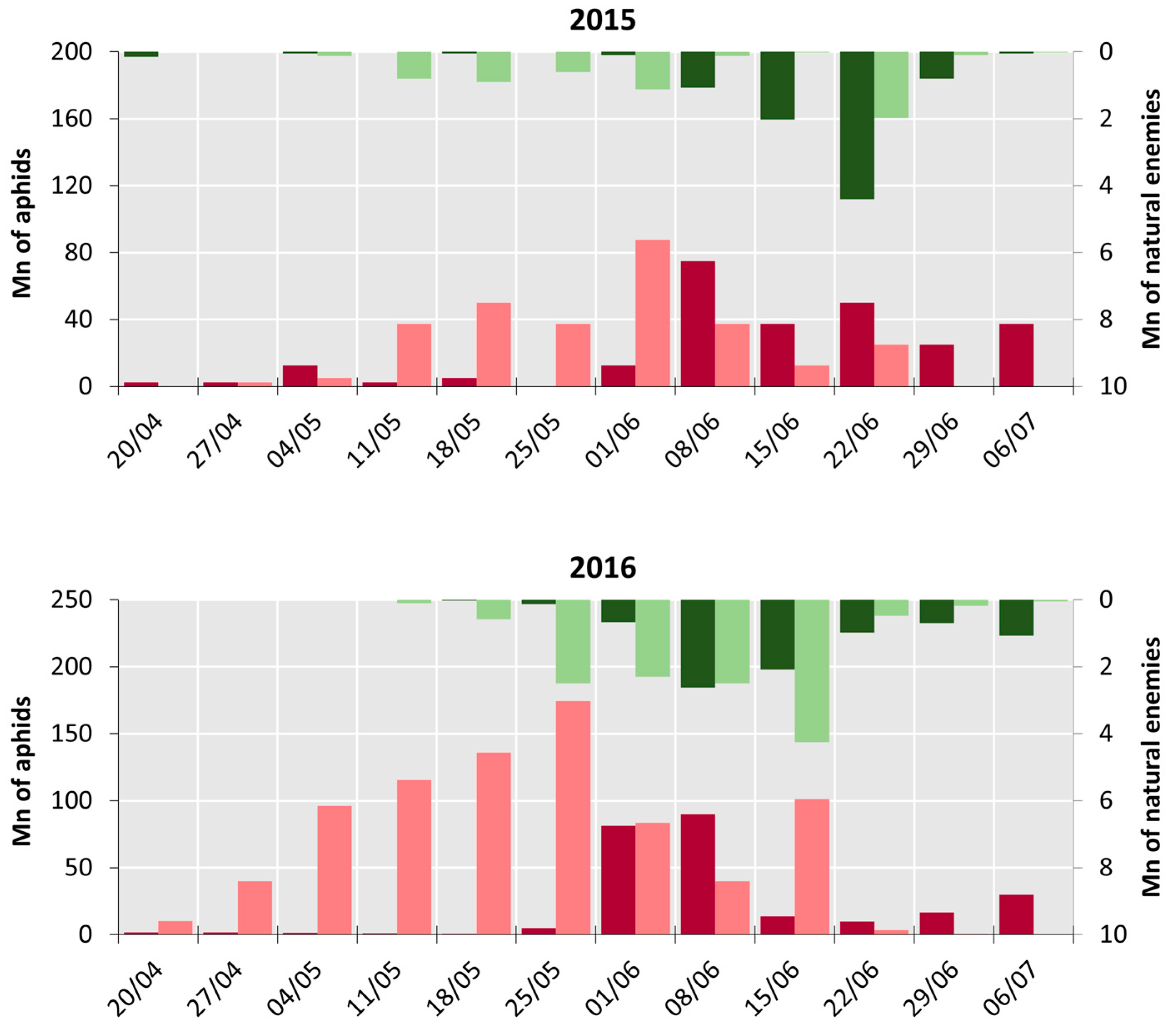
3.1.1. Population Dynamic
3.1.2. Species Interactions
3.2. Connected Patches: Intercropping Experiment
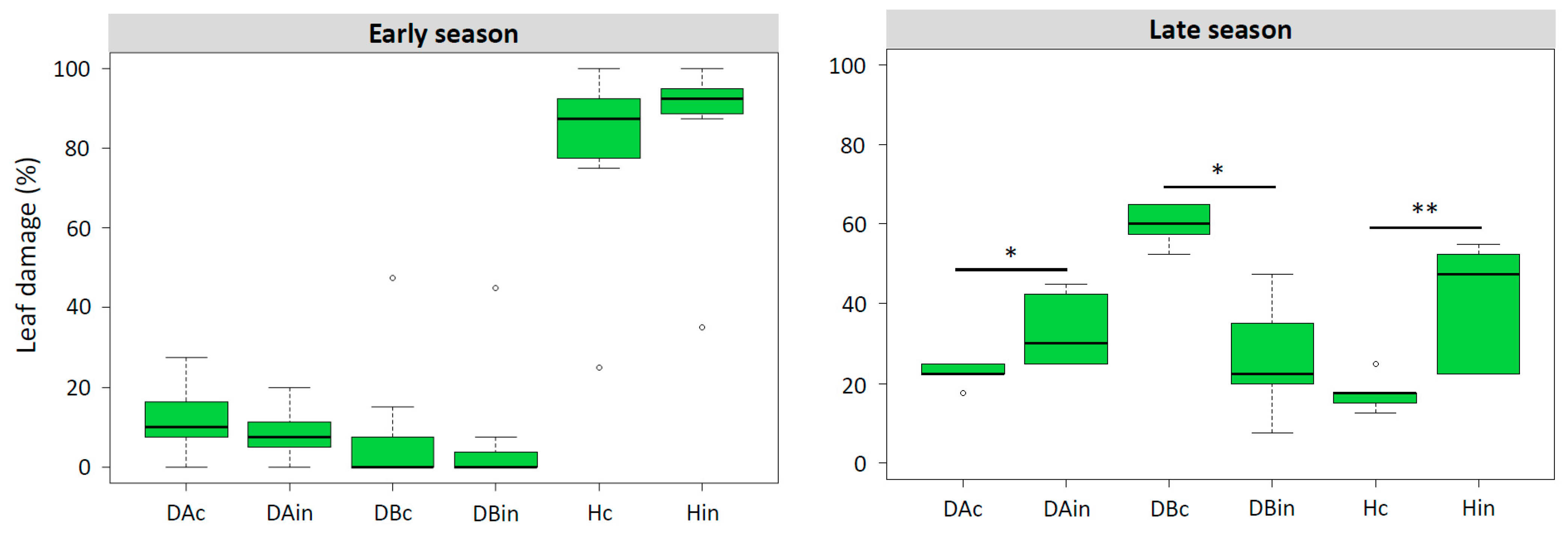
3.2.1. Population Dynamic
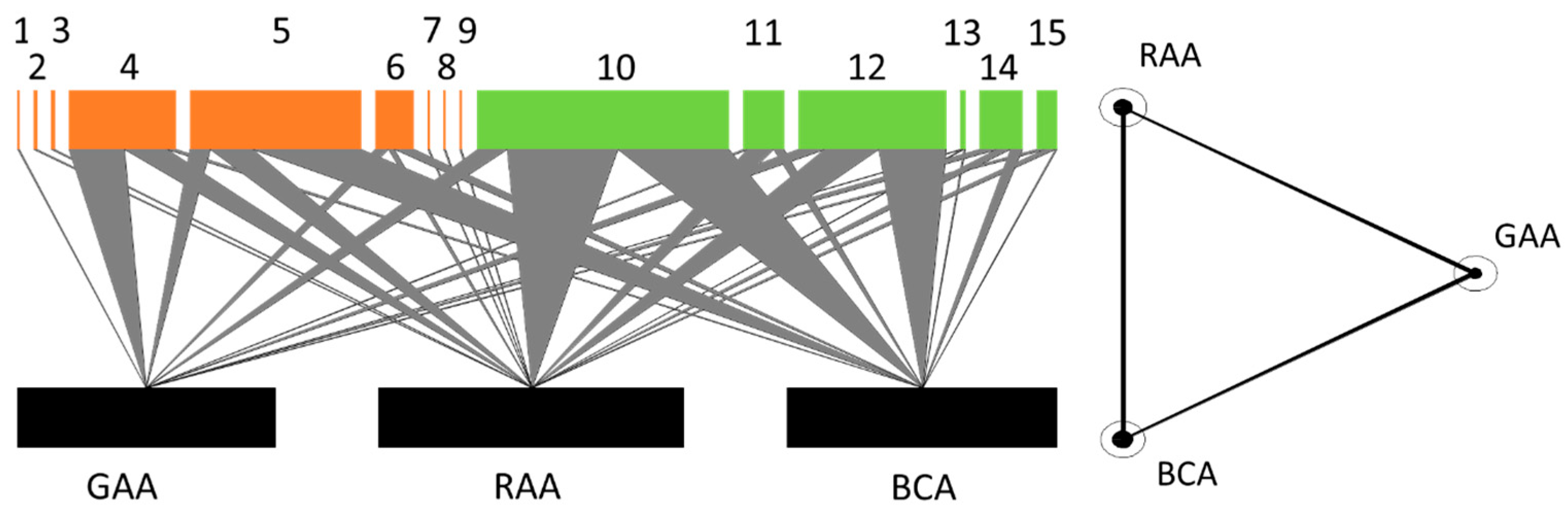
3.2.2. Species Interactions
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muller, C.B.; Godfray, H.C.J. Apparent Competition between Two Aphid Species. J. Anim. Ecol. 1997, 66, 57–64. [Google Scholar] [CrossRef]
- Morris, R.J.; Lewis, O.T.; Godfray, H.C.J. Experimental Evidence for Apparent Competition in a Tropical Forest Food Web. Nature 2004, 428, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Montoya, J.M.; Woodward, G.; Emmerson, M.C.; Solé, R.V. Press Perturbations and Indirect Effects in Real Food Webs. Ecology 2009, 90, 2426–2433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhmedi, A.; Haubruge, E.; D’Hoedt, S.; Francis, F. Quantitative Food Webs of Herbivore and Related Beneficial Community in Non-Crop and Crop Habitats. Biol. Control 2011, 58, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Frost, C.M.; Peralta, G.; Rand, T.A.; Didham, R.K.; Varsani, A.; Tylianakis, J.M. Apparent Competition Drives Community-Wide Parasitism Rates and Changes in Host Abundance across Ecosystem Boundaries. Nat. Commun. 2016, 7, 12644. [Google Scholar] [CrossRef] [Green Version]
- Bastolla, U.; Fortuna, M.A.; Pascual-García, A.; Ferrera, A.; Luque, B.; Bascompte, J. The Architecture of Mutualistic Networks Minimizes Competition and Increases Biodiversity. Nature 2009, 458, 1018–1020. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, Y.; Ma, K. The Ecological Effects of the Ant–Hemipteran Mutualism: A Meta-Analysis. Basic Appl. Ecol. 2012, 13, 116–124. [Google Scholar] [CrossRef]
- Gerardo, N.M.; Parker, B.J. Mechanisms of Symbiont-Conferred Protection against Natural Enemies: An Ecological and Evolutionary Framework. Curr. Opin. Insect Sci. 2014, 4, 8–14. [Google Scholar] [CrossRef]
- Nagy, C.; Cross, J.V.; Markó, V. Can Artificial Nectaries Outcompete Aphids in Ant-Aphid Mutualism? Applying Artificial Sugar Sources for Ants to Support Better Biological Control of Rosy Apple Aphid, Dysaphis plantaginea Passerini in Apple Orchards. Crop Prot. 2015, 77, 127–138. [Google Scholar] [CrossRef]
- Stadler, B.; Dixon, A.F.G. Ecology and Evolution of Aphid-Ant Interactions. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 345–372. [Google Scholar] [CrossRef]
- Stewart-Jones, A.; Pope, T.W.; Fitzgerald, J.D.; Poppy, G.M. The Effect of Ant Attendance on the Success of Rosy Apple Aphid Populations, Natural Enemy Abundance and Apple Damage in Orchards. Agric. Forest Entomol. 2008, 10, 37–43. [Google Scholar] [CrossRef]
- Muller, C.B.; Adriaanse, I.C.T.; Belshaw, R.; Godfray, H.C.J. The Structure of an Aphid-Parasitoid Community. J. Anim. Ecol. 1999, 68, 346–370. [Google Scholar] [CrossRef]
- Rott, A.S.; Godfray, H.C.J. The Structure of a Leafminer-Parasitoid Community. J. Anim. Ecol. 2000, 69, 274–289. [Google Scholar] [CrossRef]
- Lewis, O.T.; Memmott, J.; Lasalle, J.; Lyal, C.H.C.; Whitefoord, C.; Godfray, H.C.J. Structure of a Diverse Tropical Forest Insect–Parasitoid Community. J. Anim. Ecol. 2002, 71, 855–873. [Google Scholar] [CrossRef]
- Hirao, T.; Murakami, M. Quantitative Food Webs of Lepidopteran Leafminers and Their Parasitoids in a Japanese Deciduous Forest. Ecol. Res. 2008, 23, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Holt, R.D.; Lawton, J.H. The Ecological Consequences of Shared Natural Enemies. Annu. Rev. Ecol. Syst. 1994, 25, 495–520. [Google Scholar] [CrossRef]
- Holt, R.D.; Barfield, M. Impacts of Temporal Variation on Apparent Competition and Coexistence in Open Ecosystems. Oikos 2003, 101, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Chailleux, A.; Mohl, E.K.; Teixeira Alves, M.; Messelink, G.J.; Desneux, N. Natural Enemy-Mediated Indirect Interactions among Prey Species: Potential for Enhancing Biocontrol Services in Agroecosystems. Pest Manag. Sci. 2014, 70, 1769–1779. [Google Scholar] [CrossRef] [Green Version]
- Preisser, E.L.; Bolnick, D.I.; Benard, M.F. Scared to Death? The Effects of Intimidation and Consumption in Predator–Prey Interactions. Ecology 2005, 86, 501–509. [Google Scholar] [CrossRef] [Green Version]
- Gilman, S.E.; Urban, M.C.; Tewksbury, J.; Gilchrist, G.W.; Holt, R.D. A Framework for Community Interactions under Climate Change. Trends Ecol. Evol. 2010, 25, 325–331. [Google Scholar] [CrossRef]
- Rico-Gray, V.; Díaz-Castelazo, C.; Ramírez-Hernández, A.; Guimarães, P.R.; Nathaniel Holland, J. Abiotic Factors Shape Temporal Variation in the Structure of an Ant–Plant Network. Arthropod-Plant Interact. 2012, 6, 289–295. [Google Scholar] [CrossRef]
- Walsh, M.R. The Evolutionary Consequences of Indirect Effects. Trends Ecol. Evol. 2013, 28, 23–29. [Google Scholar] [CrossRef]
- Miñarro, M.; Hemptinne, J.-L.; Dapena, E. Colonization of Apple Orchards by Predators of Dysaphis plantaginea: Sequential Arrival, Response to Prey Abundance and Consequences for Biological Control. Biocontrol 2005, 50, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Stutz, S.; Entling, M.H. Effects of the Landscape Context on Aphid-Ant-Predator Interactions on Cherry Trees. Biol. Control 2011, 57, 37–43. [Google Scholar] [CrossRef]
- Dib, H.; Sauphanor, B.; Capowiez, Y. Effect of Management Strategies on Arthropod Communities in the Colonies of Rosy Apple Aphid, Dysaphis Plantaginea Passerini (Hemiptera: Aphididae) in South-Eastern France. Agric. Ecosyst. Environ. 2016, 216, 203–206. [Google Scholar] [CrossRef]
- Alhmedi, A.; Raymaekers, S.; Tomanović, Ž.; Bylemans, D.; Beliën, T. Food Web Structure of Aphids and Their Parasitoids in Belgian Fruit Agroecosystems: Food Webs of Aphids and Parasitoids. Entomol. Sci. 2018, 21, 279–291. [Google Scholar] [CrossRef]
- Retallack, M.; Thomson, L.; Keller, M. Native Insectary Plants Support Populations of Predatory Arthropods for Australian Vineyards. BIO Web. Conf. 2019, 15, 01004. [Google Scholar] [CrossRef]
- Alhmedi, A.; Haubruge, E.; Francis, F. Effect of Stinging Nettle Habitats on Aphidophagous Predators and Parasitoids in Wheat and Green Pea Fields with Special Attention to the Invader Harmonia axyridis Pallas (Coleoptera: Coccinellidae). Entomol. Sci. 2009, 12, 349–358. [Google Scholar] [CrossRef]
- Parolin, P.; Bresch, C.; Poncet, C.; Desneux, N. Functional Characteristics of Secondary Plants for Increased Pest Management. Int. J. Pest Manag. 2012, 58, 369–377. [Google Scholar] [CrossRef]
- Bribosia, E.; Bylemans, D.; Migon, M.; Impe, G.V. In-Field Production of Parasitoids of Dysaphis plantaginea by Using the Rowan Aphid Dysaphis sorbi as Substitute Host. Biocontrol 2005, 50, 601–610. [Google Scholar] [CrossRef]
- Cross, J.V.; Cubison, S.; Harris, A.; Harrington, R. Autumn Control of Rosy Apple Aphid, Dysaphis plantaginea (Passerini), with Aphicides. Crop Prot. 2007, 26, 1140–1149. [Google Scholar] [CrossRef]
- Alhmedi, A.; Bylemans, D.; Bangels, E.; Beliën, T. Cultivar-Mediated Effects on Apple—Dysaphis plantaginea Interaction. J. Pest Sci. 2022, 95, 1303–1315. [Google Scholar] [CrossRef]
- Blommers, L.H.M.; Helsen, H.H.M.; Vaal, F.W.N.M. Life History Data of the Rosy Apple Aphid Dysaphis plantaginea (Pass.) (Homopt., Aphididae) on Plantain and as Migrant to Apple. J. Pest Sci. 2004, 77, 155–163. [Google Scholar] [CrossRef]
- Qubbaj, T.; Reineke, A.; Zebitz, C.P.W. Molecular Interactions between Rosy Apple Aphids, Dysaphis plantaginea, and Resistant and Susceptible Cultivars of Its Primary Host Malus domestica. Entomol. Exp. Appl. 2005, 115, 145–152. [Google Scholar] [CrossRef]
- Huang, N.; Enkegaard, A.; Osborne, L.S.; Ramakers, P.M.J.; Messelink, G.J.; Pijnakker, J.; Murphy, G. The Banker Plant Method in Biological Control. Crit. Rev. Plant Sci. 2011, 30, 259–278. [Google Scholar] [CrossRef]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M. Habitat Management to Suppress Pest Populations: Progress and Prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Tomanović, Ž.; Petrović, A.; Janković, M.; Starý, P.; Yovkova, M.; Athanassiou, C.G. Review and Key for the Identification of Parasitoids (Hymenoptera: Braconidae: Aphidiinae) of Aphids Infesting Herbaceous and Shrubby Ornamental Plants in Southeastern Europe. Ann. Entomol. Soc. Am. 2013, 106, 294–309. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Tomanović, Ž.; Starý, P.; Žikić, V.; Petrović-Obradović, O. Parasitoids (Hymenoptera: Braconidae: Aphidiinae) Attacking Aphids Feeding on Solanaceae and Cucurbitaceae Crops in Southeastern Europe: Aphidiine-Aphid-Plant Associations and Key. Ann. Entomol. Soc. Am. 2010, 103, 153–164. [Google Scholar] [CrossRef]
- Rakhshani, E.; Kazemzadeh, S.; Starý, P.; Barahoei, H.; Kavallieratos, N.G.; Ćetković, A.; Popović, A.; Bodlah, l.; Tomanović, Ž. Parasitoids (Hymenoptera: Braconidae: Aphidiinae) of Northeastern Iran: Aphidiine-Aphid-Plant Associations, Key and Description of a New Species. J. Insect Sci. 2012, 12, 1–26. [Google Scholar] [CrossRef]
- Tomanović, Ž.; Starý, P.; Kavallieratos, N.G.; Gagić, V.; Plećas, M.; Janković, M.; Rakhshani, E.; Ćetković, A.; Petrović, A. Aphid parasitoids (Hymenoptera: Braconidae: Aphidiinae) in wetland habitats in western Palaearctic: Key and associated aphid parasitoid guilds. Ann. Société Entomol. Fr. 2012, 48, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Wolfram Research, Inc. Mathematica, Version 5.0; Wolfram Research, Inc.: Champaign, IL, USA, 2003. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Development Core Team: Vienna, Austria, 2009. [Google Scholar]
- Zabel, J.; Tscharntke, T. Does Fragmentation of Urtica Habitats Affect Phytophagous and Predatory Insects Differentially? Oecologia 1998, 116, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Branson, D.H. Relationships between Plant Diversity and Grasshopper Diversity and Abundance in the Little Missouri National Grassland. Psyche J. Entomol. 2011, 2011, 748635. [Google Scholar] [CrossRef]
- Altieri, M. The Dynamics of Colonizing Arthropod Communities at the Interface of Abandoned, Organic and Commercial Apple Orchards and Adjacent Woodland Habitats. Agric. Ecosyst. Environ. 1986, 16, 29–43. [Google Scholar] [CrossRef]
- Thomson, L.J.; Hoffmann, A.A. Spatial Scale of Benefits from Adjacent Woody Vegetation on Natural Enemies within Vineyards. Biol. Control 2013, 64, 57–65. [Google Scholar] [CrossRef]
- Corbett, A.; Rosenheim, J.A. Impact of a Natural Enemy Overwintering Refuge and Its Interaction with the Surrounding Landscape. Ecol. Entomol. 1996, 21, 155–164. [Google Scholar] [CrossRef]
- Stefanova, D.; Malchev, S. Preliminary Assessment of Selected Sweet Cherry Hybrids Regarding Their Resistance to Black Cherry Aphid (Myzus cerasi Fabr.) in Bulgaria. Agric. Sci. Technol. 2020, 12, 288–291. [Google Scholar] [CrossRef]
- Mackauer, M.; Völkl, W. Regulation of Aphid Populations by Aphidiid Wasps: Does Parasitoid Foraging Behaviour or Hyperparasitism Limit Impact? Oecologia 1993, 94, 339–350. [Google Scholar] [CrossRef]
- Heimpel, G.E.; Rosenheim, J.A.; Mangel, M. Predation on Adult Aphytis Parasitoids in the Field. Oecologia 1997, 110, 346–352. [Google Scholar] [CrossRef]
- Colfer, R.G.; Rosenheim, J.A. Predation on Immature Parasitoids and Its Impact on Aphid Suppression. Oecologia 2001, 126, 292–304. [Google Scholar] [CrossRef]
- Snyder, W.E.; Ives, A.R. Generalist Predators Disrupt Biological Control by a Specialist Parasitoid. Ecology 2001, 82, 705–716. [Google Scholar] [CrossRef]
- Mailleux, A.-C.; Deneubourg, J.-L.; Detrain, C. Regulation of Ants’ Foraging to Resource Productivity. Proc. Biol. Sci. 2003, 270, 1609–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devigne, C.; Detrain, C. Foraging Responses of the Aphid Tending Ant Lasius niger to Spatio-Temporal Changes in Aphid Colonies Cinara Cedri. Acta Zool. Sin. 2005, 51, 161–166. [Google Scholar]
- Harmon, J.P.; Andow, D.A. Behavioral Mechanisms Underlying Ants’ Density-Dependent Deterrence of Aphid-Eating Predators. Oikos 2007, 116, 1030–1036. [Google Scholar] [CrossRef]
- Miller, K.E.; Polaszek, A.; Evans, D.M. A Dearth of Data: Fitting Parasitoids into Ecological Networks. Trends Parasitol. 2021, 37, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.W.; England, S. Indirect Interactions in Biological Control of Insects: Pests and Natural Enemies in Alfalfa. Ecol. Appl. 1996, 6, 920–930. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Natural Enemies (Code) | |
|---|---|
| Parasitoids Aphidius ervi (1), A. matricariae (2), A. urticae (3), Binodoxys angelicae (4), Ephedrus persicae (5), E. plagiator (6), Lipolexis gracilis (7), Praon abjectum (8), Toxares deltiger (9). | Predators Cecidomyiidae (10), Forficulidae (11), Syrphidae (12), Chrysopidae (13), Coccinellidae (14), Anthocoridae (15). |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhmedi, A.; Belien, T.; Bylemans, D. Habitat Modification Alters Food Web Interactions with Focus on Biological Control of Aphids in Apple Orchards. Sustainability 2023, 15, 5978. https://doi.org/10.3390/su15075978
Alhmedi A, Belien T, Bylemans D. Habitat Modification Alters Food Web Interactions with Focus on Biological Control of Aphids in Apple Orchards. Sustainability. 2023; 15(7):5978. https://doi.org/10.3390/su15075978
Chicago/Turabian StyleAlhmedi, Ammar, Tim Belien, and Dany Bylemans. 2023. "Habitat Modification Alters Food Web Interactions with Focus on Biological Control of Aphids in Apple Orchards" Sustainability 15, no. 7: 5978. https://doi.org/10.3390/su15075978