
Effects of the Salt-Tolerant Gramineous Forage Echinochloa frumentacea on Biological Improvement and Crop Productivity in Saline–Alkali Land on the Hetao Ningxia Plain in China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Species
2.2. Background of Experimental Sites
2.3. Experimental Design
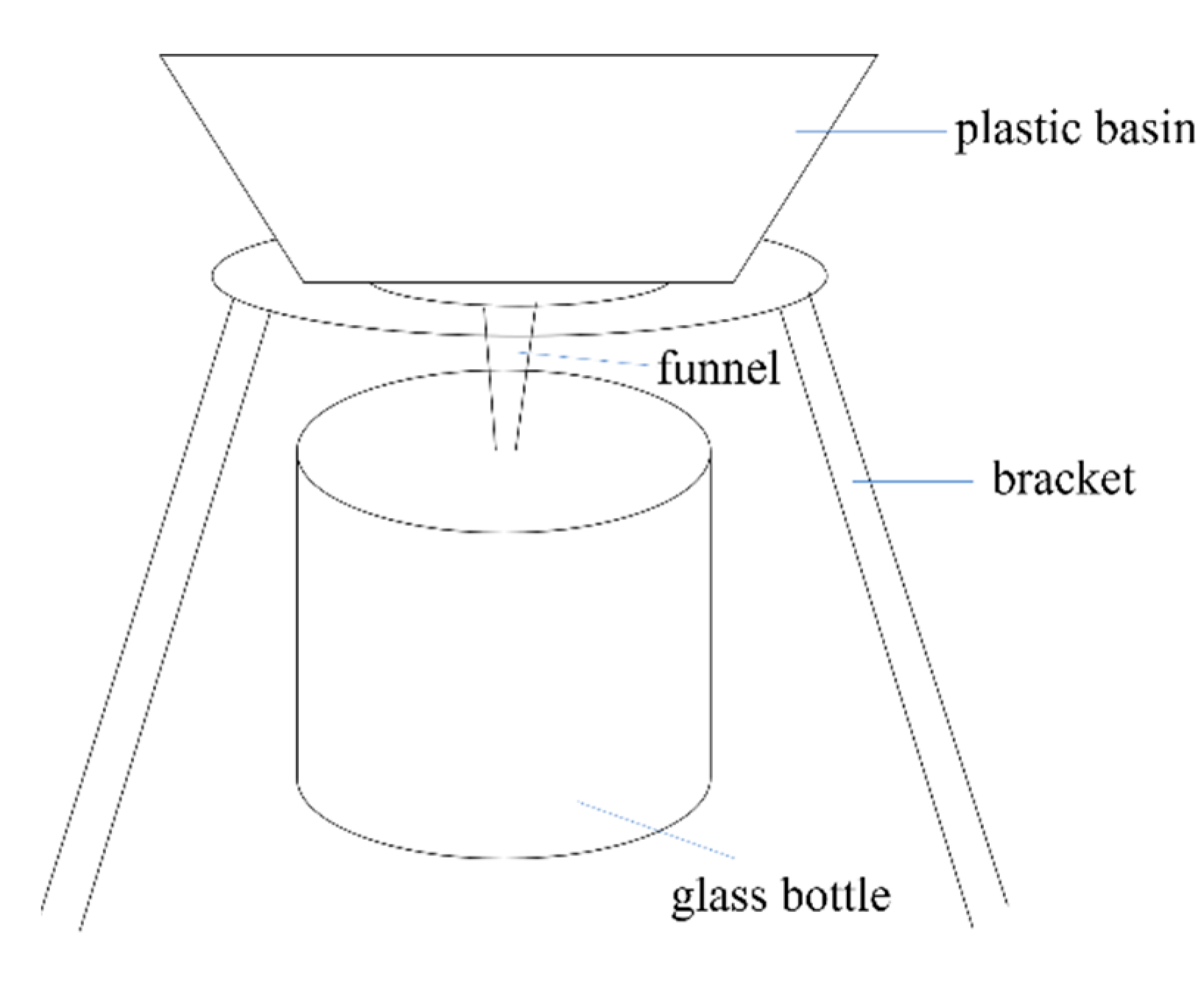
2.3.1. Experiment 1. Leaching and Desalination Experiment of E. frumentacea
2.3.2. Experiment 2. The Experiment for Improving Saline–Alkali Land by Planting E. frumentacea Coupled with Soil Conditioners
- CK: Bare land, control;
- Treatment A: No soil modifier;
- Treatment B: Phosphogypsum (22.5 t·ha−1) + decomposed sheep dung (15 t·ha−1);
- Treatment C: Superphosphate (300 kg·ha−1) + decomposed sheep dung (15 t·ha−1);
- Treatment D: Phosphogypsum (22.5 t·ha−1) + superphosphate (300 kg·ha−1) + decomposed sheep dung (15 t·ha−1);
- Treatment E: Phosphogypsum (30 t·ha−1) + superphosphate (600 kg·ha−1) + decomposed sheep dung (30 t·ha−1);
- Treatment F: Phosphogypsum (30 t·ha−1) + superphosphate (900 kg·ha−1) + decomposed sheep dung (30 t·ha−1).
2.3.3. Experiment 3. Effects of Intercropping of E. frumentacea with Leguminous Forages on Productivity and Bacterial Diversity
2.4. Measurements
2.4.1. Soil Sample Collection and Processing
2.4.2. Determination of Soil Chemical Properties
2.4.3. High-Throughput Sequencing of Soil Bacteria
2.4.4. Determination Method of Plant Yields
2.4.5. Evaluation Index of Land Use Efficiency
- LER: This index used to measure yield advantages, which can reflect the land use efficiency of intercropping.
- 2.
- AYL: Compared with monoculture, the relative yield of intercropping under a certain planting ratio is lost or increased. AYL > 0, which means that the processing shows a gain compared with single processing; AYL < 0, indicating that the treatment shows a loss compared with single treatment. The positive and negative of AYLE and AYLL indicate the contribution of E. frumentacea or leguminous forages to gains or losses in the system.
AYLE = (YEL/ZEL)/(YEE/PEE) − 1
AYLL = (YLE/ZLE)/(YLL/PLL) − 1
- 3.
- CR is an index used to evaluate the competition among species. When CR > 1, it indicates that the competitiveness of intercropping crops is greater than that of companion crops.
2.5. Statistical Analysis
3. Results
3.1. Effects of Planting Different Forage Species on Soil Ions under Saline–Alkali Stress
3.2. Effects of Planting E. frumentacea on the Soil pH, Total Water Soluble Salt Content and Soil Nutrients
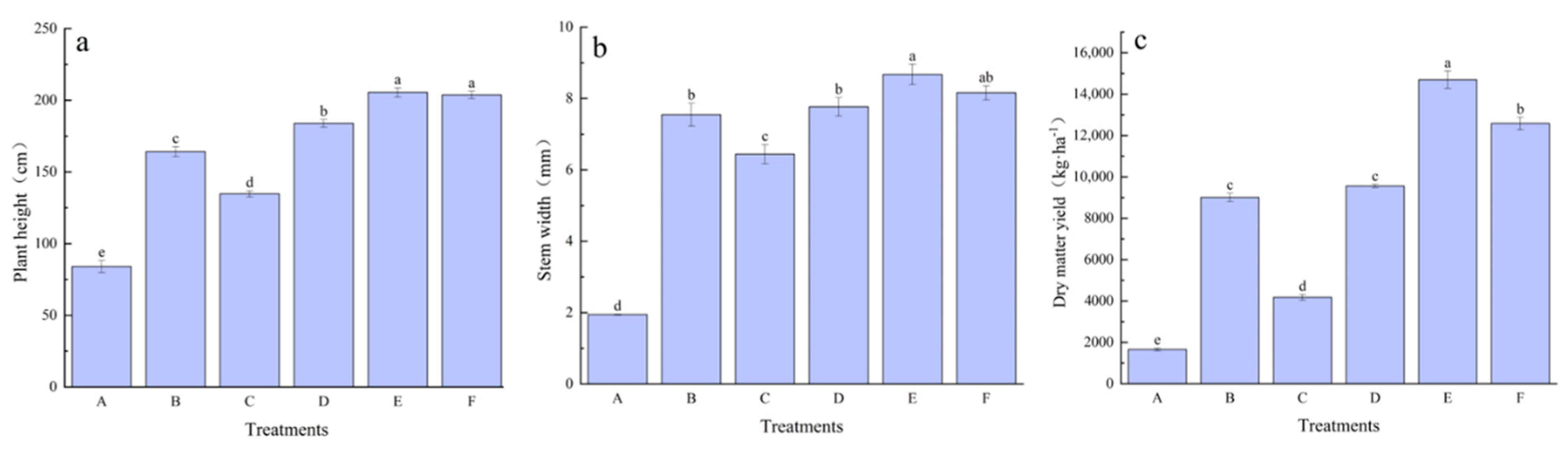
3.3. Effects of Modifiers on Growth and Yields of E. frumentacea
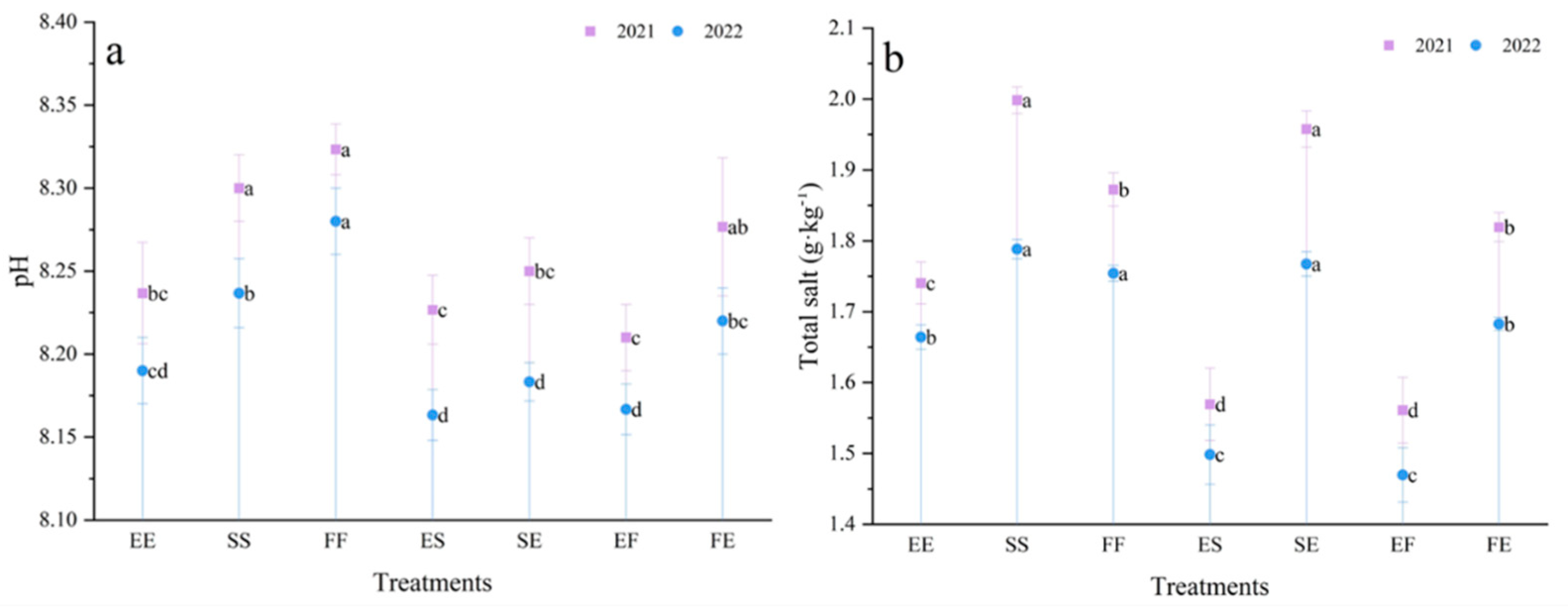
3.4. Effects of Different Planting Patterns on Soil pH and Total Salt
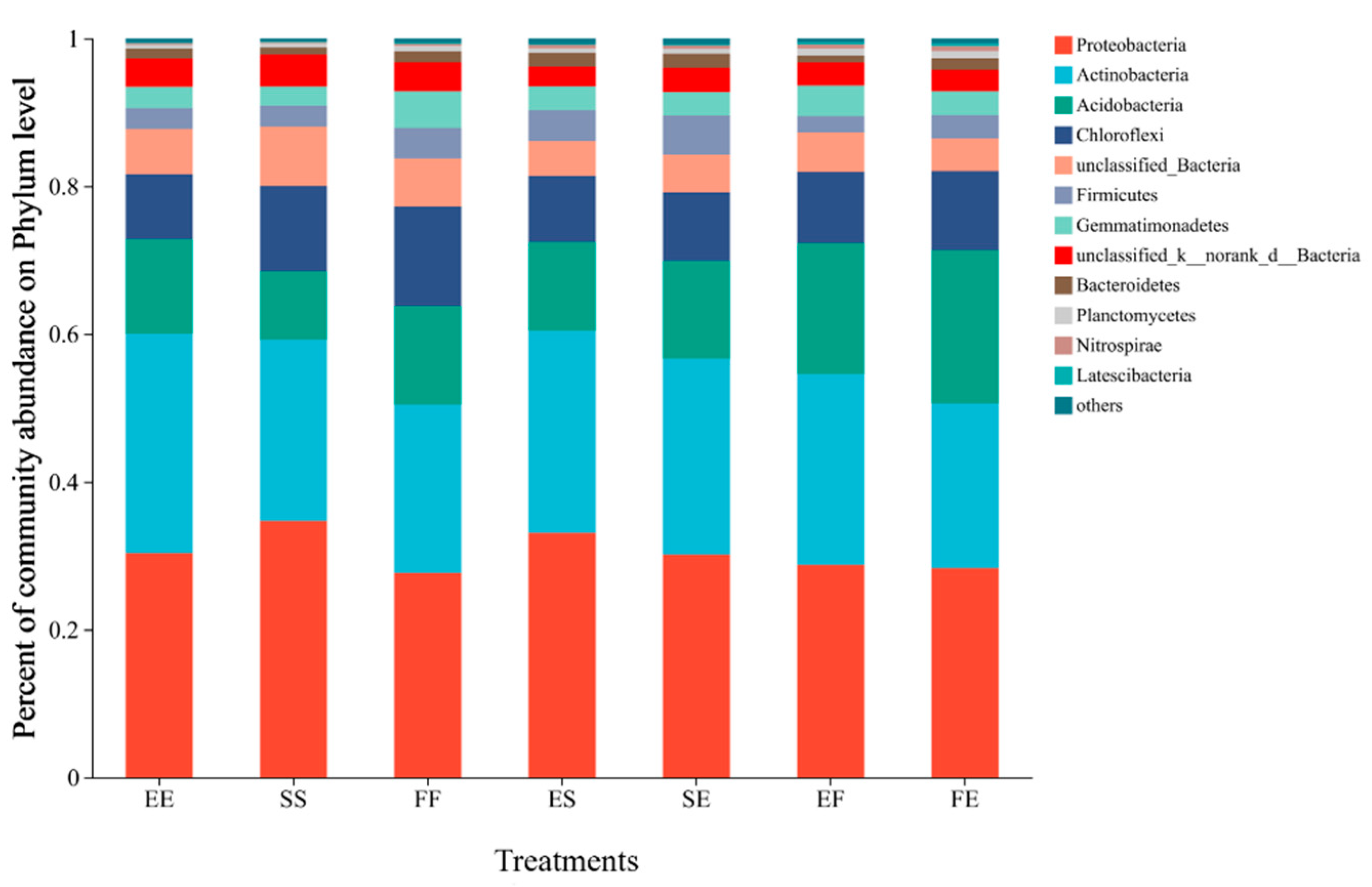
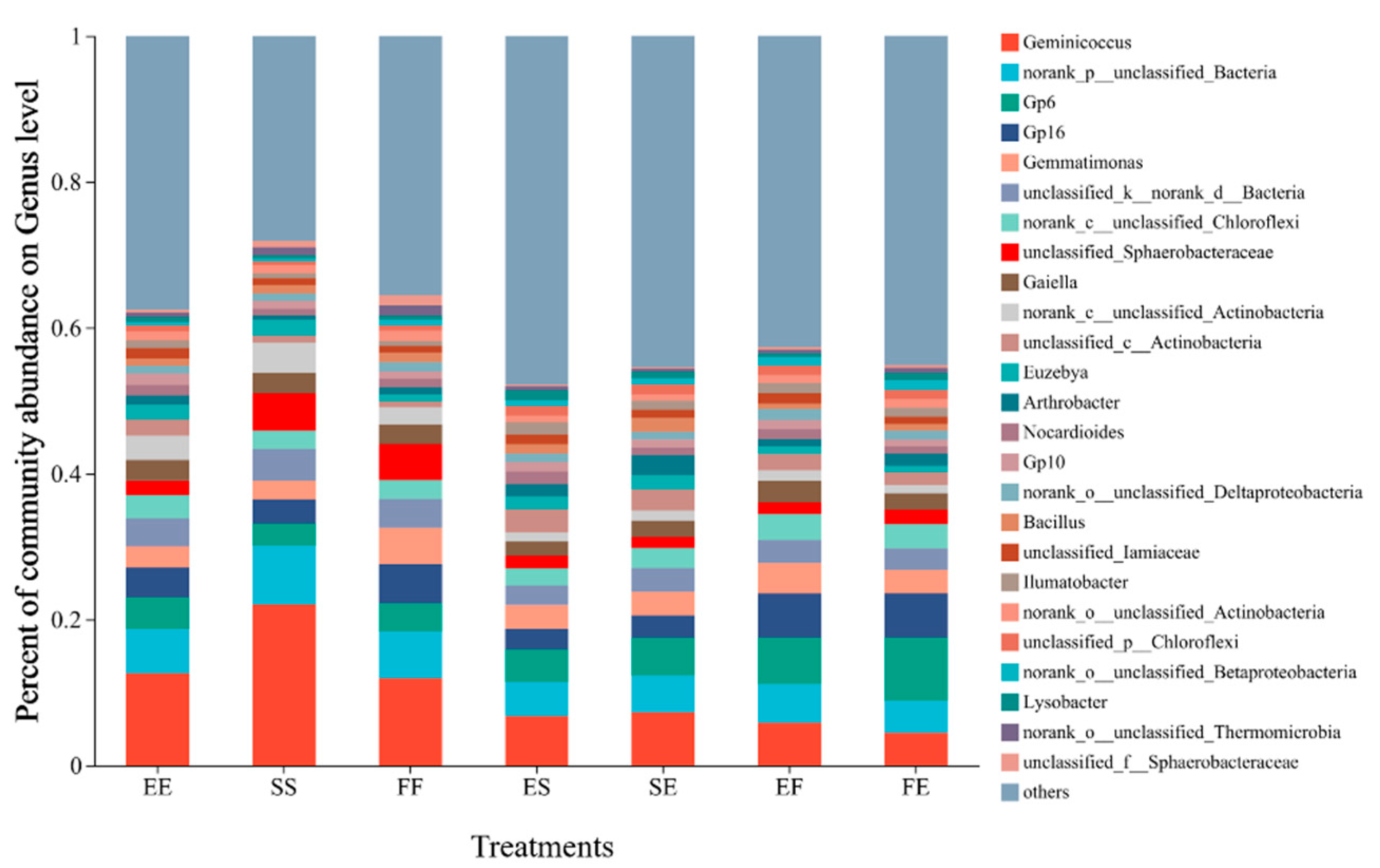
3.5. Effects of Different Planting Patterns on Bacterial Diversity and Community Structure in Rhizosphere Soil
3.6. Effects of Different Planting Patterns on Hay Yields
3.7. Effects of Different Intercropping Modes on Land Use Efficiency
4. Discussion
4.1. Changes in Soil Salt Ions and Chemical Properties after Planting E. frumentacea on Saline–Alkali Land
4.2. Effects of E. frumentacea Intercropping on Soil Bacterial Diversity and Community Structure
4.3. Effects of E. frumentacea Intercropping with Leguminous Forages on Hay Yields and Land Use Efficiency
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, H.; Lu, H.Y.; Chu, L.; Shao, H.B.; Shi, W.M. Biochar applied with appropriate rates can reduce N leaching, keep N reten-tion and not increase NH3 volatilization in a coastal saline soil. Sci. Total Environ. 2017, 575, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Wang, J.M. Integrated life cycle assessment of improving saline-sodic soil with flue gas desulfurization gypsum. J. Clean. Prod. 2018, 202, 332–341. [Google Scholar] [CrossRef]
- Liu, L.; Long, X.; Shao, H.; Liu, Z.; Tao, Y.; Zhou, Q.; Zong, J. Ameliorants improve saline–alkaline soils on a large scale in northern Jiangsu Province, China. Ecol. Eng. 2015, 81, 328–334. [Google Scholar]
- Zhao, Y.; Wang, S.; Li, Y.; Liu, J.; Zhuo, Y.; Chen, H.; Wang, J.; Xu, L.; Sun, Z. Extensive reclamation of saline-sodic soils with flue gas desulfurization gypsum on the Songnen Plain, Northeast China. Geoderma 2018, 321, 52–60. [Google Scholar] [CrossRef]
- Singh, K. Microbial and Enzyme Activities of Saline and Sodic Soils. Land Degrad. Dev. 2015, 27, 706–718. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, S.; Li, Y.; Bu, K.; Zhang, Y.; Chang, L.; Zhang, Y. Dynamics of saline-alkali land and its ecological regionalization in western Songnen Plain, China. Chin. Geogr. Sci. 2010, 20, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Yan, N.; Marschner, P.; Cao, W.; Zuo, C.; Qin, W. Influence of salinity and water content on soil microorganisms. Int. Soil Water Conserv. Res. 2015, 3, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Kursakova, V.S. The effect of perennial herbs on the physical properties of saline soils. Eurasian Soil Sci. 2006, 39, 748–752. [Google Scholar] [CrossRef]
- Liang, J.; Shi, W. Cotton/halophytes intercropping decreases salt accumulation and improves soil physicochemical properties and crop productivity in saline-alkali soils under mulched drip irrigation: A three-year field experiment. Field Crop. Res. 2020, 262, 108027. [Google Scholar] [CrossRef]
- Li, Y.-H.; Shi, D.-Y.; Li, G.-H.; Zhao, B.; Zhang, J.-W.; Liu, P.; Ren, B.; Dong, S.-T. Maize/peanut intercropping increases photosynthetic characteristics, 13C-photosynthate distribution, and grain yield of summer maize. J. Integr. Agric. 2019, 18, 2219–2229. [Google Scholar] [CrossRef]
- Du, J.B.; Han, T.F.; Gai, J.Y.; Yong, T.W.; Xin, S.; Wang, X.C.; Feng, Y.; Jiang, L.; Kai, S.; Liu, W.; et al. Maize-soybean strip intercropping: Achieved a balance between high productivity and sustainability. J. Integr. Agric. 2018, 17, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Raza, M.A.; Gul, H.; Wang, J.; Yasin, H.S.; Qin, R.; Bin Khalid, M.H.; Naeem, M.; Feng, L.Y.; Iqbal, N.; Gitari, H.; et al. Land productivity and water use efficiency of maize-soybean strip intercropping systems in semi-arid areas: A case study in Punjab Province, Pakistan. J. Clean. Prod. 2021, 308, 127282. [Google Scholar] [CrossRef]
- Stefan, L.; Hartmann, M.; Engbersen, N.; Six, Z.; Schöb, C. Positive Effects of Crop Diversity on Productivity Driven by Changes in Soil Microbial Composition. Front. Microbiol. 2021, 12, 660749. [Google Scholar] [CrossRef] [PubMed]
- Cappelli, S.L.; Domeignoz-Horta, L.A.; Loaiza, V.; Laine, A.-L. Plant biodiversity promotes sustainable agriculture directly and via belowground effects. Trends Plant Sci. 2022, 27, 674–687. [Google Scholar] [CrossRef]
- Upadhyaya, H.D.; Dwivedi, S.L.; Singh, S.K.; Singh, S.; Vetriventhan, M.; Sharma, S. Forming core collections in barnyard, kodo, and little millets using mor-phoagronomic descriptors. Crop Sci. 2014, 54, 2673–2682. [Google Scholar] [CrossRef]
- Muldoon, D. The effect of photoperiod on the growth and development of Echinochloa spp. Millets. Aust. J. Exp. Agric. 1985, 25, 428–433. [Google Scholar] [CrossRef]
- Gandhimeyyan, V.R.; Vanniarajan, C.; Nirmalakumari, A.; Arunachalam, P.; Thiyageshwari, S.; Karthikeyan, A.; Govindaraj, M. Gene effects and heterosis for grain Fe and Zn content in barnyard millet (Echinochloa frumentacea (Roxb.) link). Genetika 2020, 52, 621–639. [Google Scholar] [CrossRef]
- Chandrasekar, B.R.; Ambrose, G.; Jayabalan, N. Influence of biofertilizers and nitrogen source level on the growth and yield of Echinochloa frumentacea (Roxb.) Link. J. Agric. Sci. Technol. 2005, 1, 223–234. [Google Scholar]
- Subramanian, A.; Rajendran, N.R.; Jeyaprakash, P. In vitro and in vivo screening of barnyard millet (Echinochloa frumentacea (roxb.) link) germplasm for salinity tolerance. Plant Arch. 2020, 20, 7389–7397. [Google Scholar]
- Sood, S.; Khulbe, R.K.; Gupta, A.K.; Agrawal, P.K.; Upadhyaya, H.D.; Bhatt, J.C. Barnyard millet—A potential food and feed crop of future. Plant Breed. 2015, 134, 135–147. [Google Scholar] [CrossRef]
- Dan, B.S. Soil Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Rukun, L. Analysis Methods of Soil Agrochemistry; China Agricultural Science and Technology Press: Beijing, China, 1999; pp. 156–157. (In Chinese) [Google Scholar]
- Litourgidis, A.S.; Vlachostergios, D.N.; Dordas, C.A.; Damalas, C.A. Dry matter yield, nitrogen content, and competition in pea–cereal intercropping systems. Eur. J. Agron. 2011, 34, 287–294. [Google Scholar] [CrossRef]
- MacFall, J.S.; Johnson, G.A.; Kramer, P.J. Comparative water uptake by roots of different ages in seedlings of loblolly pine (Pinus taeda L.). New Phytol. 1991, 119, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Enstone, D.E.; Peterson, C.A.; Ma, F. Root Endodermis and Exodermis: Structure, Function, and Responses to the Environment. J. Plant Growth Regul. 2002, 21, 335–351. [Google Scholar] [CrossRef]
- Xia, J.; Ren, J.; Zhang, S.; Wang, Y.; Fang, Y. Forest and grass composite patterns improve the soil quality in the coastal saline-alkali land of the Yellow River Delta, China. Geoderma 2019, 349, 25–35. [Google Scholar] [CrossRef]
- Peng, B.; Xu, W.; Shao, R.; Feng, G.; Shi, W. Growth of suaeda salsa tin response to salt stress in different habitats. Acta Pratacul. Sin. 2016, 25, 81–90. (In Chinese) [Google Scholar]
- Qadir, M.; Steffens, D.; Yan, F.; Schubert, S. Sodium removal from a calcareous saline-sodic soil through leaching and plant uptake during phytoremediation. Land Degrad. Dev. 2003, 14, 301–307. [Google Scholar] [CrossRef]
- Xie, X.; Pu, L.; Wang, Q.; Zhu, M.; Xu, Y.; Zhang, M. Response of soil physicochemical properties and enzyme activities to long-term reclamation of coastal saline soil, Eastern China. Sci. Total. Environ. 2017, 607–608, 1419–1427. [Google Scholar] [CrossRef]
- Wang, X.; Xu, X.; Lu, A.; Li, X.; Zhang, F. Cultivation of two barnyard varieties improves physicochemical properties of saline-alkali land through mediating rhizospheric microbiome and metabolome. Agronomy 2022, 12, 1322. [Google Scholar] [CrossRef]
- Keshri, J.; Mishra, A.; Jha, B. Microbial population index and community structure in saline–alkaline soil using gene targeted metagenomics. Microbiol. Res. 2013, 168, 165–173. [Google Scholar] [CrossRef]
- Lange, M.; Eisenhauer, N.; Sierra, C.A.; Bessler, H.; Engels, C.; Griffiths, R.I.; Mellado-Vázquez, P.G.; Malik, A.A.; Roy, J.; Scheu, S.; et al. Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 2015, 6, 6707. [Google Scholar] [CrossRef] [Green Version]
- Steinauer, K.; Chatzinotas, A.; Eisenhauer, N. Root exudate cocktails: The link between plant diversity and soil microorgan-isms? Ecol. Evol. 2016, 6, 7387–7396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, J.; Das, A.; Dolan, E.M.; Hu, Z. Ammonia-oxidizing archaea involved in nitrogen removal. Water Res. 2009, 43, 1801–1809. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Zhou, J.; Yang, L.; Gunina, A.; Yang, Y.; Peixoto, L.; Zeng, Z.; Zang, H.; Kuzyakov, Y. Diversified cropping systems benefit soil carbon and nitrogen stocks by increasing aggregate stability: Results of three fractionation methods. Sci. Total. Environ. 2022, 824. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Zhang, Y.; Dong, W.; Zhang, Y.; Zhang, G.; Sun, Z.; Yang, L. Vermicompost can suppress Fusarium oxysporum f. sp. lycopersici via generation of benefi-cial bacteria in a long-term tomato monoculture soil. Plant Soil 2019, 440, 491–505. [Google Scholar] [CrossRef]
- Jo, S.-G.; Kang, Y.-I.; Om, K.-S.; Cha, Y.-H.; Ri, S.-Y. Growth, photosynthesis and yield of soybean in ridge-furrow intercropping system of soybean and flax. Field Crop. Res. 2021, 275, 108329. [Google Scholar] [CrossRef]
- Li, C.; Dong, Y.; Li, H.; Shen, J.; Zhang, F. The Dynamic Process of Interspecific Interactions of Competitive Nitrogen Capture between In-tercropped Wheat (Triticum aestivum L.) and Faba Bean (Vicia faba L.). PLoS ONE 2014, 9, e115804. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Yang, Z.; Dong, S. Interspecific competitiveness affects the total biomass yield in an alfalfa and corn intercropping system. Field Crop. Res. 2011, 124, 66–73. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Saline–Alkali Soil Type | Treatment | Na+ (g·kg−1) | K+ (g·kg−1) | Ca2+ (g·kg−1) | Mg2+ (g·kg−1) | Cl− (g·kg−1) | SO42− (g·kg−1) | CO32− (g·kg−1) | HCO3− (g·kg−1) |
|---|---|---|---|---|---|---|---|---|---|
| Alkalized soil | Bare land (CK) | 0.78 ± 0.03a | 20.06 ± 1.02a | 0.20 ± 0.04a | 0.04 ± 0.01b | 0.47 ± 0.02a | 0.29 ± 0.01a | 0.27 ± 0.03a | 0.24 ± 0.02a |
| E. frumentacea | 0.50 ± 0.01e | 5.00 ± 0.35d | 0.12 ± 0.01e | 0.04 ± 0.00b | 0.40 ± 0.03b | 0.16 ± 0.05e | 0.21 ± 0.06bc | 0.17 ± 0.05b | |
| Barnyard grass | 0.56 ± 0.04d | 5.50 ± 0.29d | 0.15 ± 0.02c | 0.04 ± 0.01b | 0.41 ± 0.01b | 0.17 ± 0.01de | 0.18 ± 0.01c | 0.16 ± 0.01c | |
| Oat | 0.67 ± 0.04c | 9.90 ± 0.31c | 0.11 ± 0.02d | 0.04 ± 0.01b | 0.41 ± 0.03ab | 0.19 ± 0.08cd | 0.19 ± 0.04c | 0.17 ± 0.07bc | |
| S. europaea | 0.70 ± 0.03b | 17.36 ± 1.73b | 0.18 ± 0.06b | 0.05 ± 0.00a | 0.43 ± 0.01ab | 0.26 ± 0.06b | 0.23 ± 0.02b | 0.15 ± 0.03d | |
| Alfalfa | 0.68 ± 0.07bc | 8.90 ± 1.19c | 0.06 ± 0.03f | 0.05 ± 0.00a | 0.42 ± 0.02ab | 0.21 ± 0.02c | 0.15 ± 0.04d | 0.13 ± 0.03e | |
| Salinized soil | Bare land (CK) | 1.51 ± 0.42a | 54.00 ± 7.23a | 0.26 ± 0.03a | 0.37 ± 0.04a | 2.69 ± 0.23a | 1.20 ± 0.12a | 0.021 ± 0.00a | 0.113 ± 0.02a |
| E. frumentacea | 0.70 ± 0.13e | 38.40 ± 3.81de | 0.16 ± 0.05b | 0.33 ± 0.03b | 2.37 ± 0.16d | 0.94 ± 0.20c | 0.019 ± 0.00bc | 0.083 ± 0.00bc | |
| Barnyard grass | 0.73 ± 0.10d | 39.00 ± 4.97d | 0.19 ± 0.03b | 0.34 ± 0.02ab | 2.40 ± 0.17d | 1.09 ± 0.18b | 0.018 ± 0.00cd | 0.076 ± 0.01bc | |
| Oat | 0.71 ± 0.35e | 43.90 ± 5.54c | 0.20 ± 0.04b | 0.33 ± 0.02b | 2.42 ± 0.10cd | 1.10 ± 0.13ab | 0.017 ± 0.00d | 0.071 ± 0.00bc | |
| S. europaea | 0.87 ± 0.09c | 49.90 ± 3.81b | 0.25 ± 0.02a | 0.36 ± 0.03a | 2.56 ± 0.15b | 1.14 ± 0.19ab | 0.020 ± 0.00ab | 0.091 ± 0.00ab | |
| Alfalfa | 1.05 ± 0.70b | 35.86 ± 5.92e | 0.17 ± 0.06b | 0.32 ± 0.02b | 2.54 ± 0.16bc | 1.05 ± 0.17b | 0.015 ± 0.00e | 0.063 ± 0.00c |
| Treatment | pH | Total Salt | Organic Matter | Available N | Available P | Available K |
|---|---|---|---|---|---|---|
| g·kg−1 | g·kg−1 | mg·kg −1 | mg·kg −1 | mg·kg −1 | ||
| CK | 10.08 ± 0.02a | 2.23 ± 0.03a | 4.93 ± 0.16d | 2.80 ± 0.16ce | 2.23 ± 0.14e | 83.37 ± 2.45f |
| A | 9.92 ± 0.01b | 1.78 ± 0.01b | 5.40 ± 0.32cd | 3.07 ± 0.16bcde | 3.06 ± 0.37e | 90.77 ± 2.64e |
| B | 9.72 ± 0.01c | 1.21 ± 0.02e | 7.20 ± 0.49ab | 4.61 ± 0.27a | 8.22 ± 0.08c | 127.97 ± 3.22c |
| C | 9.22 ± 0.00d | 1.62 ± 0.10c | 6.57 ± 1.57bc | 3.16 ± 0.16bcd | 5.09 ± 0.52d | 104.57 ± 3.55d |
| D | 9.13 ± 0.01e | 1.45 ± 0.03d | 6.78 ± 0.49b | 4.43 ± 0.16a | 15.61 ± 0.08b | 130.13 ± 2.45c |
| E | 8.84 ± 0.02f | 0.92 ± 0.02g | 7.63 ± 0.64ab | 3.26 ± 0.27b | 19.07 ± 1.36a | 152.18 ± 2.04a |
| F | 8.52 ± 0.01g | 1.10 ± 0.03f | 8.26 ± 0.32a | 3.16 ± 0.16bc | 16.90 ± 1.80b | 142.72 ± 1.54b |
| Treatment | Diversity Index | Community Richness Index | ||
|---|---|---|---|---|
| Shannon | Simpson | Chao1 | ACE | |
| EE | 6.221 ± 0.20c | 0.006 ± 0.00b | 2748.178 ± 153.28bc | 2739.791 ± 154.71bc |
| SS | 5.872 ± 0.11d | 0.010 ± 0.00a | 2509.509 ± 312.92c | 2541.299 ± 352.43c |
| FF | 6.358 ± 0.12bc | 0.005 ± 0.00b | 3013.089 ± 237.52b | 3025.151 ± 247.87b |
| ES | 6.562 ± 0.07ab | 0.004 ± 0.00b | 3791.356 ± 258.01a | 3778.554 ± 182.07a |
| SE | 6.545 ± 0.20ab | 0.005 ± 0.00b | 3877.336 ± 280.03a | 3866.806+260.16a |
| EF | 6.569 ± 0.06ab | 0.004 ± 0.00b | 3647.032 ± 168.69a | 3610.956 ± 187.72a |
| FE | 6.634 ± 0.013a | 0.004 ± 0.00b | 3762.663 ± 316.51a | 3723.070 ± 286.45a |
| Treatment | 2021 | 2022 | F Value | ||
|---|---|---|---|---|---|
| Dry Matter Yield(kg·ha−1) | Dry Matter Yield(kg·ha−1) | Year (df = 1) | Treatment (df = 6) | YT (df = 6) | |
| EE | 16,551.45 ± 276.03aB | 17,014.64 ± 242.21aA | 169.64 *** | 5161.44 *** | 9.57 *** |
| SS | 6421.66 ± 153.67dB | 6871.18 ± 127.13dA | |||
| FF | 9489.78 ± 173.90cB | 10,991.09 ± 161.45cA | |||
| ES | 11,557.69 ± 151.79bB | 12,384.39 ± 182.92bA | |||
| SE | 2502.8 ± 66.53fA | 2681.74 ± 75.19fA | |||
| EF | 11,465.81 ± 183.75bB | 12,293.64 ± 180.00bA | |||
| FE | 4018.90 ± 123.44eB | 4533.81 ± 148.87eA | |||
| Treatment | Year | LER | AYL | CR |
|---|---|---|---|---|
| TES | 2021 | 1.09 ± 0.01a | 0.17 ± 0.02a | 1.80 ± 0.06a |
| 2022 | 1.12 ± 0.02a | 0.23 ± 0.03a | 1.63 ± 0.05b | |
| TEF | 2021 | 1.12 ± 0.02a | 0.24 ± 0.04a | 1.87 ± 0.08a |
| 2022 | 1.14 ± 0.02a | 0.27 ± 0.03a | 1.75 ± 0.07a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Y.; Xie, X.; Wang, X.; Zhu, L.; Qiu, Q.-S.; Xu, X. Effects of the Salt-Tolerant Gramineous Forage Echinochloa frumentacea on Biological Improvement and Crop Productivity in Saline–Alkali Land on the Hetao Ningxia Plain in China. Sustainability 2023, 15, 5319. https://doi.org/10.3390/su15065319
Cheng Y, Xie X, Wang X, Zhu L, Qiu Q-S, Xu X. Effects of the Salt-Tolerant Gramineous Forage Echinochloa frumentacea on Biological Improvement and Crop Productivity in Saline–Alkali Land on the Hetao Ningxia Plain in China. Sustainability. 2023; 15(6):5319. https://doi.org/10.3390/su15065319
Chicago/Turabian StyleCheng, Yunlong, Xiaowei Xie, Xueqin Wang, Lin Zhu, Quan-Sheng Qiu, and Xing Xu. 2023. "Effects of the Salt-Tolerant Gramineous Forage Echinochloa frumentacea on Biological Improvement and Crop Productivity in Saline–Alkali Land on the Hetao Ningxia Plain in China" Sustainability 15, no. 6: 5319. https://doi.org/10.3390/su15065319