
Stuck in the Caterpillars’ Web: A Half-Century of Biocontrol Research and Application on Gregarious Lepidopteran Pests in Europe

Abstract
:1. Introduction
2. Collective Behavior of Larval Lepidoptera
3. Pestilential or Phytophagous?
4. Criteria for Identifying BC Cases
5. Species-Wise Overview and Current Status
5.1. Hyphantria cunea Drury
5.2. Euproctis chrysorrhoea (L.)
5.3. Malacosoma neustria L.
5.4. Thaumetopoea pityocampa (Denis and Schiffermüller, 1775)
5.5. Thaumetopoea processionea (L.)
5.6. Yponomeuta malinellus (Zeller)
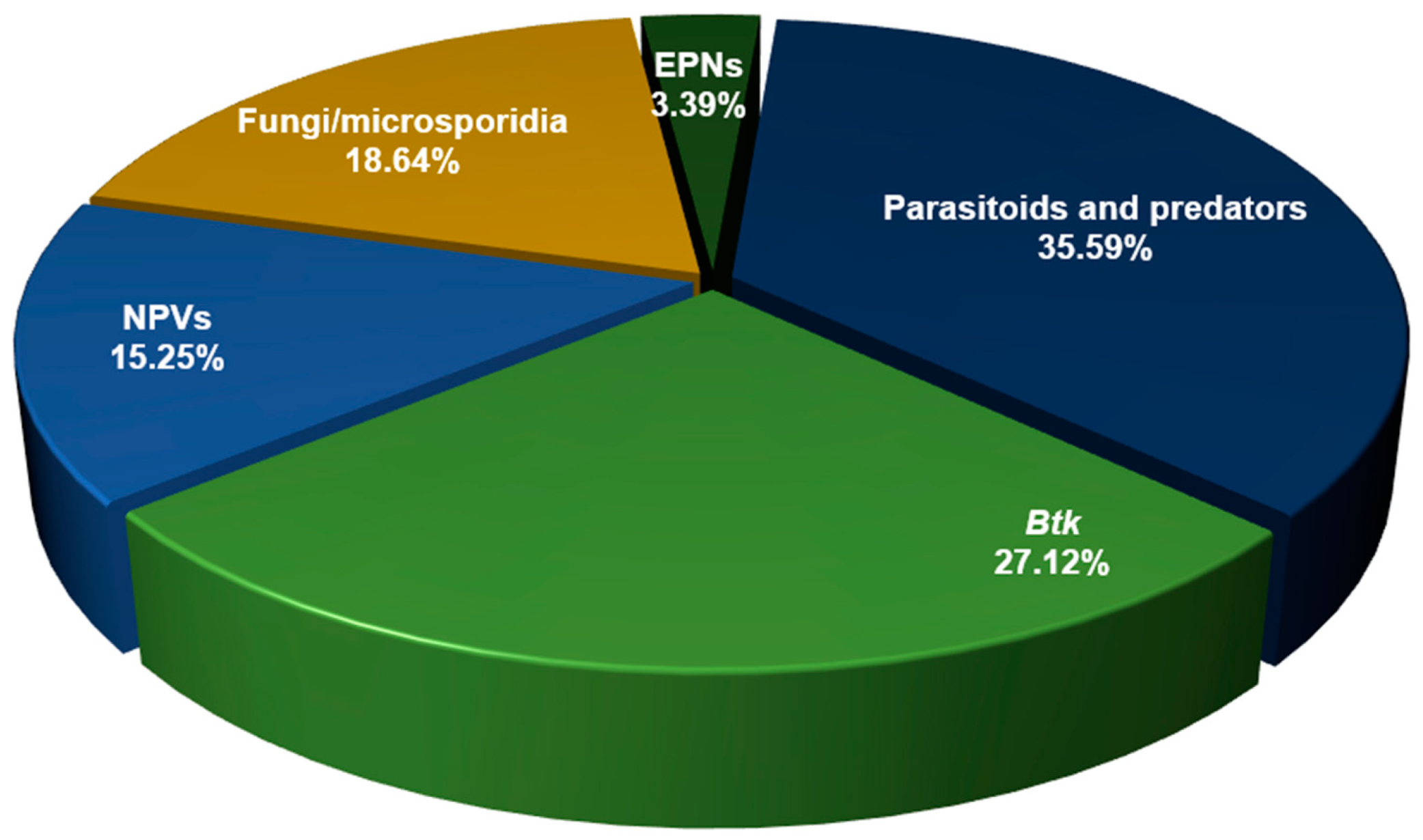
6. Outcomes and BCA Representation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carter, D.J. Pest Lepidoptera of Europe: With Special Reference to the British Isles, 1st ed.; Springer: Dordrecht, The Netherlands, 1984; p. 432. [Google Scholar]
- Oberemok, V.V.; Laikova, K.V.; Gninenko, Y.I.; Zaitsev, A.S.; Nyadar, P.M.; Adeyemi, T.A. A short history of insecticides. J. Plant Prot. Res. 2015, 55, 221–226. [Google Scholar] [CrossRef]
- Casida, J.E.; Quistad, G.B. Golden age of insecticide research: Past, present, or future? Annu. Rev. Entomol. 1998, 43, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.C.; Ida, R.M.M.; Jitendra, K.M.; Robin, B.D.; Wolf-D, D.; Milatovic, D. Insecticides. In Biomarkers in Toxicology, 2nd ed.; Gupta, R.C., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 455–475. [Google Scholar] [CrossRef]
- Taylor, J.M.; Snyder, W.E. Are specialists really safer than generalists for classical biocontrol? BioControl 2021, 66, 9–22. [Google Scholar] [CrossRef]
- DeBach, P. Biological Control of Insect Pests and Weeds, 1st ed.; Cambridge University Press: Cambridge, UK, 1964; p. 844. [Google Scholar]
- Zimmerman, E.C. Insects of Hawaii. Vol. 7. (Macrolepidoptera); University of Hawaii Press: Honolulu, HI, USA, 1958; p. 542. [Google Scholar]
- van Lenteren, J.C.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. BioControl 2018, 63, 39–59. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Tuzun, S.; Kuć, J.A. Proposed definitions related to induced disease resistance. Biocontrol Sci. Technol. 1992, 2, 349–351. [Google Scholar] [CrossRef]
- Nordlund, D.A. Biological control, integrated pest management and conceptual models. Biocontrol News Inf. 1996, 17, 35N–44N. [Google Scholar]
- Eilenberg, J.; Hajek, A.; Lomer, C. Suggestions for unifying the terminology in biological control. BioControl 2001, 46, 387–400. [Google Scholar] [CrossRef]
- Heimpel, G.E.; Mills, N.J. Biological Control: Ecology and Applications, 1st ed.; Cambridge University Press: Cambridge, UK, 2017; p. 386. [Google Scholar]
- Stenberg, J.A.; Sundh, I.; Becher, P.G.; Björkman, C.; Dubey, M.; Egan, P.A.; Friberg, H.; Gil, J.F.; Jensen, D.F.; Jonsson, M.; et al. When is it biological control? A framework of definitions, mechanisms, and classifications. J. Pest Sci. 2021, 94, 665–676. [Google Scholar] [CrossRef]
- Gurr, G.; Wratten, S. Biological Control: Measures of Success, 1st ed.; Springer: Dordrecht, The Netherlands, 2002; p. 429. [Google Scholar] [CrossRef]
- Zeilinger, S.; Omann, M. Trichoderma biocontrol: Signal transduction pathways involved in host sensing and mycoparasitism. Gene Regul. Syst. Biol. 2007, 1, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Nicot, P.C. Classical and Augmentative Biological Control against Diseases and Pests: Critical Status Analysis and Review of Factors Influencing Their Success, 1st ed.; IOBC: St. Paul, MN, USA, 2011; p. 194p. Available online: https://hal.inrae.fr/hal-02809014/document (accessed on 16 December 2022).
- Barratt, B.I.P.; Moran, V.C.; Bigler, F.; van Lenteren, J.C. The status of biological control and recommendations for improving uptake for the future. BioControl 2018, 63, 155–167. [Google Scholar] [CrossRef]
- Lynch, L.; Hokkanen, H.M.T.; Babendreier, D.; Bigler, F.; Burgio, G.; Gao, Z.H.; Kuske, S.; Loomans, A.; Menzler-Hokkanen, I.; Thomas, M.B.; et al. Insect biological control and non-target effects: A European perspective. In Evaluating Indirect Ecological Effects of Biological Control, 1st ed.; Wajnberg, E., Scott, J.K., Quimby, P.C., Eds.; CABI Publishing: Wallingford, UK, 2001; pp. 99–123. Available online: http://sherekashmir.informaticspublishing.com/397/1/9780851994536.pdf (accessed on 16 December 2022).
- Gerber, E.; Schaffner, U. Review of Invertebrate Biological Control Agents Introduced into Europe, 1st ed.; CABI Switzerland: Delémont, Switzerland, 2016; p. 187. [Google Scholar] [CrossRef]
- Kabaluk, J.T.; Svircev, A.M.; Goettel, M.S.; Woo, S.G. The Use and Regulation of Microbial Pesticides in Representative Jurisdictions Worldwide; International Organization for Biological Control of Noxious Animals and Plants (IOBC): St. Paul, MN, USA, 2010; p. 99. Available online: https://www.iobc-global.org/ (accessed on 10 December 2022).
- Fitzgerald, T.D.; Costa, J.T. Collective behavior in social caterpillars. In Information Processing in Social Insects, 1st ed.; Detrain, C., Deneubourg, J.I., Pasteels, J.M., Eds.; Springer Basel AG: Basel, Switzerland, 1999; pp. 379–400. [Google Scholar] [CrossRef]
- Stamp, N.E. Egg deposition patterns in butterflies: Why do some species cluster their eggs rather than deposit them singly? Am. Nat. 1980, 115, 367–380. [Google Scholar] [CrossRef]
- Peterson, S.C. Communication of leaf suitability by gregarious eastern tent caterpillars (Malacosoma americanum). Ecol. Entomol. 1987, 12, 283–289. [Google Scholar] [CrossRef]
- Fitzgerald, T.D.; Webster, F.X. Identification and behavioral assays of the trail pheromone of the forest tent caterpillar, Malacosoma disstria Hübner (Lepidoptera: Lasiocampidae). Can. J. Zool. 1993, 71, 1511–1515. [Google Scholar] [CrossRef]
- Fitzgerald, T.D. The Tent Caterpillars, 1st ed.; Cornell University Press: Ithaca, NY, USA, 1995; p. 317. [Google Scholar] [CrossRef]
- Costa, J.T.; Pierce, N.E. Social evolution in the Lepidoptera: Ecological context and communication in larval societies. In The Evolution of Social Behavior in Insects and Arachnids, 1st ed.; Choe, C.J., Crespi, B.J., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 407–442. [Google Scholar] [CrossRef]
- Ehrlich, P.R. Which Animal Will Invade? In Ecology of Biological Invasions of North America and Hawaii, 1st ed.; Mooney, H.A., Drake, J.A., Eds.; Springer: New York, NY, USA, 1986; pp. 79–95. [Google Scholar] [CrossRef]
- Robinson, G.S.; Ackery, P.R.; Kitching, I.J.; Beccaloni, G.W.; Hernández, L.M. HOSTS–A Database of the World’s Lepidopteran Hostplants; Natural History Museum: London, UK, 2010; Available online: https://www.nhm.ac.uk/our-science/data/hostplants/ (accessed on 9 December 2022).
- Ruf, C. Social Lifestyles in Caterpillars: Behavioral Mechanisms and Ecological Consequences. Doctoral Dissertation, Faculty of Biology, Chemistry & Earth Sciences, University of Bayreuth, Bayreuth, Germany, December 2002. [Google Scholar]
- Bernal, J.S.; Medina, R.F. Agriculture sows pests: How crop domestication, host shifts, and agricultural intensification can create insect pests from herbivores. Curr. Opin. Insect. Sci. 2018, 26, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Thomson, L.J.; Macfadyen, S.; Hoffmann, A.A. Predicting the effects of climate change on natural enemies of agricultural pests. Biol. Control 2010, 52, 296–306. [Google Scholar] [CrossRef]
- Molis, S. The buff-tip moth (Phalera bucephala L.)-a pest of apple trees. Acta Entomol. Litu. 1970, 1, 182–183. [Google Scholar]
- Whitten, M.J. Pest management in 2000: What we might learn from the twentieth century. In Pest Management and the Environment in 2000, 1st ed.; Kadir, A., Barlow, H., Eds.; CAB International and Agricultural Institute of Malaysia: Wallingford, UK, 1992; pp. 9–44. [Google Scholar]
- Kogan, M. Integrated pest management theory and practice. Entomol. Exp. Appl. 1988, 49, 59–70. [Google Scholar] [CrossRef]
- Flint, M.L.; van den Bosch, R. Introduction to Integrated Pest Management, 1st ed.; Springer: New York, NY, USA, 1981; p. 256. [Google Scholar] [CrossRef]
- Deseö, K.V.; Montermini, A.; Cortellini, W. The first outbreak of the fall webworm (Hyphantria cunea Drury; Lepid., Arctiidae) in North Italy: Preliminary observations on the development, diapause and derivation. J. Appl. Entomol. 1986, 101, 201–206. [Google Scholar] [CrossRef]
- Sullivan, G.T.; Karaca, I.; Ozman-Sullivan, S.K.; Kara, K. Tachinid (Diptera: Tachinidae) parasitoids of overwintered Hyphantria cunea (Drury)(Lepidoptera: Arctiidae) pupae in hazelnut plantations in Samsun province, Turkey. J. Entomol. Res. Soc. 2012, 14, 21–30. [Google Scholar]
- Nakonechna, Y.O.; Stankevych, S.V.; Zabrodina, I.V.; Lezhenina, I.P.; Filatov, M.O.; Yushchuk, D.D.; Lutytska, N.V.; Molchanova, O.A.; Melenti, V.O.; Poliakh, V.M.; et al. Distribution area of Hyphantria cunea Drury: The analysis of Ukrainian and world data. Ukr. J. Ecol. 2019, 9, 214–220. [Google Scholar] [CrossRef]
- Gomi, T.; Takeda, M. The transition to a trivoltine life cycle and mechanisms that enforce the voltinism change in Hyphantria cunea Drury (Lepidoptera: Arctiidae) in Kobe. Appl. Entomol. Zool. 1990, 25, 483–489. [Google Scholar] [CrossRef]
- Ge, X.; He, S.; Zhu, C.; Wang, T.; Xu, Z.; Zong, S. Projecting the current and future potential global distribution of Hyphantria cunea (Lepidoptera: Arctiidae) using CLIMEX. Pest Manag. Sci. 2019, 75, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Du, H.; Li, Y.; Li, X.; Sun, L.; Cao, C. Identification and characterization of key genes in insulin signaling pathway as molecular targets for controlling the fall webworm, Hyphantria cunea. Pest Manag. Sci. 2023, 79, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Kean, J.M.; Kumarasinghe, L.B. Predicting the seasonal phenology of fall webworm (Hyphantria cunea) in New Zealand. N. Z. Plant Prot. 2007, 60, 279–285. [Google Scholar] [CrossRef]
- Kelly, M.K.; Entwistle, P.F.; Sterling, P.H.; Speight, M.H.; Laport, R.F. Virus control of the Browntail moth, Euproctis chrysorrhoea. In Proceedings of the Lymantriidae: A Comparison of Features of New and Old World Tussock Moths, New Haven, CT, USA, 26 June–1 July 1988; Wallner, W.E., McManus, K.A., Eds.; Volume 123, p. 427. [Google Scholar]
- Longo, S. Relieves on activity of Telenomus turkarkandas (Szabo’)(Hym.: Scelionidae), an egg parasite of Euproctis chrysorrhoea (L.)(Lep.: Lymantriidae) in Sicily. Phytophaga 1989, 3, 61–72. [Google Scholar]
- Kuznetsov, V.I. Lymantriidae (Liparidae, Orgyidae). In Insects and Mites—Pests of Agricultural Plants, 1st ed.; Kuznetsov, V.I., Ed.; Nauka: St. Petersburg, Russia, 1999; Volume 3, pp. 279–320. [Google Scholar]
- Schaefer, W.P. Attacking Wasps, Polistes and Therion, Penetrate Silk Nests of Fall Webworm. Environ. Entomol. 1977, 6, 591. [Google Scholar] [CrossRef]
- Boyd, K.S.; Drummond, F.; Donahue, C.; Groden, E. Factors influencing the population fluctuations of Euproctis chrysorrhoea (Lepidoptera: Erebidae) in Maine. Environ. Entomol. 2021, 50, 1203–1216. [Google Scholar] [CrossRef]
- Hussain, B.; Sivakumar, G.; Kannan, M.; War, A.R.; Ballal, C.R. First record of a nucleopolyhedrovirus infecting brown-tail moth larvae, Euproctis chrysorrhoea (L.)(Lepidoptera: Lymantriidae) in India. Egypt. J. Biol. Pest Control. 2019, 29, 11. [Google Scholar] [CrossRef]
- Magnoler, A. Bioassay of nucleopolyhedrosis virus against larval instars of Malacosoma neustria. J. Invertebr. Pathol. 1975, 25, 343–348. [Google Scholar] [CrossRef]
- Žikić, V.; Stanković, S.S.; Kavallieratos, N.G.; Athanassiou, C.; Georgiou, P.; Tschorsnig, H.P.; van Achterberg, C. Parasitoids associated with Lymantria dispar (Lepidoptera: Erebidae) and Malacosoma neustria (Lepidoptera: Lasiocampidae) in Greece and comparative analysis of their parasitoid spectrums in Europe. Zool. Anz. 2017, 270, 166–175. [Google Scholar] [CrossRef]
- Chistyakov, Y.A. Family Lasiocampidae. In Butterflies—Pests of the Far Eastern Agriculture. Keys, 1st ed.; Kirpichnikova, V.A., Ler, P.A., Eds.; DVO AN: Vladivostok, Russia, 1988; pp. 153–165. [Google Scholar]
- Xudong, Z.; Lizhi, Z.; Yujiang, Z.; Yan, L.; Wanxing, D. Analysis of the climate factors contributing to the outbreak of Malacosoma neustria testacea Motschulsky in Daxing’an Mountains. For. Pest Dis. 2004, 23, 27–30. [Google Scholar]
- Özbek, H.; Çoruh, S. Larval parasitoids and larval diseases of Malacosoma neustria L. (Lepidoptera: Lasiocampidae) detected in Erzurum Province, Turkey. Turk. J. Zool. 2012, 36, 447–459. [Google Scholar] [CrossRef]
- Jactel, H.; Barbaro, L.; Battisti, A.; Bosc, A.; Branco, M.; Brockerhoff, E.; Castagneyrol, B.; Dulaurent, A.M.; Hódar, J.A.; Jacquet, J.S.; et al. Insect–tree interactions in Thaumetopoea pityocampa. In Processionary Moths and Climate Change: An update, 1st ed.; Roques, A., Ed.; Springer: Dordrecht, The Netherlands, 2015; pp. 265–310. [Google Scholar]
- Campôa, J.; Calvão, T.; Firmino, P.N.; Pimentel, C.S. Disentangling the effects of climate and defoliation on forest growth: The case of an outbreak of a Thaumetopoea pityocampa population with a shifted phenology in a Pinus pinaster monoculture. For. Ecol. Manag. 2021, 498, 119548. [Google Scholar] [CrossRef]
- Groenen, F.; Meurisse, N. Historical distribution of the oak processionary moth Thaumetopoea processionea in Europe suggests recolonization instead of expansion. Agr. For. Entomol. 2012, 14, 147–155. [Google Scholar] [CrossRef]
- Pocock, M.J.; Roy, H.E.; Fox, R.; Ellis, W.N.; Botham, M. Citizen science and invasive alien species: Predicting the detection of the oak processionary moth Thaumetopoea processionea by moth recorders. Biol. Conserv. 2017, 208, 146–154. [Google Scholar] [CrossRef]
- Alonso, C.; Vuorisalo, T.; Wilsey, B.; Honkanen, T. Yponomeuta evonymellus outbreaks in southern Finland: Spatial synchrony but different local magnitudes. Ann. Zool. Fenn. 2000, 37, 179–188. Available online: https://www.jstor.org/stable/23736001.
- Kuhlmann, U.; Carl, K.P.; Mills, N.J. Quantifying the impact of insect predators and parasitoids on populations of the apple ermine moth, Yponomeuta malinellus (Lepidoptera: Yponomeutidae), in Europe. Bull. Entomol. Res. 1998, 88, 165–175. [Google Scholar] [CrossRef]
- Unruh, T.; Short, R.; Herard, F.; Chen, K.; Hopper, K.; Pemberton, R.; Lee, J.H.; Ertle, L.; Swan, K.; Fuester, R.; et al. Introduction and establishment of parasitoids for the biological control of the apple ermine moth, Yponomeuta malinellus (Lepidoptera: Yponomeutidae), in the Pacific Northwest. Biol. Control 2003, 28, 332–345. [Google Scholar] [CrossRef]
- Gencer, L. The parasitoids of Yponomeuta malinellus Zeller (Lepidoptera: Yponomeutidae) in Sivas. Turk. J. Zool. 2003, 27, 43–46. [Google Scholar]
- Tosun, H.; Derya, B.; Erler, F. Potential of entomopathogenic fungi as biological control agents of Yponomeuta malinellus Zeller, 1838 (Lepidoptera: Yponomeutidae). Mediterr. Agric. Sci. 2022, 35, 121–128. [Google Scholar] [CrossRef]
- Maksimov, S.A.; Marushchak, V.N.; Dorofeeva, L.M. On the causes of outbreaks of mass reproduction of the apple ermine moth Yponomeuta malinellus Zell. (Lepidoptera: Yponomeutidae) in the Urals. In Proceedings of the National Academy of Sciences of the Kyrgyz Republic, Bishkek, Kyrgyzstan; 2019; Volume 6, pp. 64–69. Available online: https://www.elibrary.ru/item.asp?id=44653128 (accessed on 30 December 2022).
- Vargas-Osuna, E.; Aldebis, H.K.; Caballero, P.; Lipa, J.J.; Santiago-Alvarez, C. A newly described baculovirus (subgroup B) from Ocnogyna baetica (Rambur) (Lepidoptera: Arctiidae) in Southern Spain. J. Invertebr. Pathol. 1994, 63, 31–36. [Google Scholar] [CrossRef]
- Dijken, M.J.V.; Kole, M.; Lenteren, J.C.V.; Brand, A.M. Host-preference studies with Trichogramma evanescens Westwood (Hym., Trichogrammatidae) for Mamestra brassicae, Pieris brassicae and Pieris rapae. J. Appl. Entomol. 1986, 101, 64–85. [Google Scholar] [CrossRef]
- Costa, J.T.; Fitzgerald, T.D. Social terminology revisited: Where are we ten years later? Ann. Zoo. Fenn. 2005, 42, 559–564. [Google Scholar]
- Ramadan, M.M.; Murai, K.T.; Johnson, T. Host range of Secusio extensa (Lepidoptera: Arctiidae), and potential for biological control of Senecio madagascariensis (Asteraceae). J. Appl. Entomol. 2011, 135, 269–284. [Google Scholar] [CrossRef]
- Chen, Q.; Zhao, H.; Wen, M.; Li, J.; Zhou, H.; Wang, J.; Zhou, Y.; Liu, Y.; Du, L.; Kang, H.; et al. Genome of the webworm Hyphantria cunea unveils genetic adaptations supporting its rapid invasion and spread. BMC Genom. 2020, 21, 242. [Google Scholar] [CrossRef]
- Jaenike, J.; Selander, R.K. On the question of host races in the fall webworm, Hyphantria cunea. Entomol. Exp. Appl. 1980, 27, 31–37. [Google Scholar] [CrossRef]
- Schowalter, T.D.; Ring, D.R. Biology and management of the fall webworm, Hyphantria cunea (Lepidoptera: Erebidae). J. Integr. Pest Manag. 2017, 8, 1–6. [Google Scholar] [CrossRef]
- Ning, J.; Lu, P.; Fan, J.; Ren, L.; Zhao, L. American fall webworm in China: A new case of global biological invasions. Innovation 2021, 3, 100201. [Google Scholar] [CrossRef] [PubMed]
- Department for Environment, Food & Rural Affairs. Rapid Pest Risk Analysis (PRA) for: Hyphantria cunea, November 2015. Available online: https://pra.eppo.int (accessed on 29 November 2022).
- Ji, R.; Xie, B.; Li, X.; Gao, Z.; Li, D. Research progress on the invasive species, Hyphantria cunea. Entomol. Knowledg. 2003, 40, 13–18. [Google Scholar]
- Yang, Z.; Zhang, Y. Researches on techniques for biocontrol of the fall webworm, Hyphantria cunea, a severe invasive insect pest to China. Chin. J. Entomol. 2007, 44, 465–471. [Google Scholar]
- Franz, J.M. Biological control of pest insects in Europe. Annu. Rev. Entomol. 1961, 6, 183–200. [Google Scholar] [CrossRef] [PubMed]
- Szirmai, J. Biological defense against Hyphantria cunea Drury by means of viruses. Acta Microbiol. Acad. Sci. Hung. 1957, 4, 31–42. [Google Scholar] [PubMed]
- Warren, L.O.; Tadić, M. The Fall Webworm, Hyphantria cunea, Its Distribution and Natural Enemies: A World List (Lepidoptera: Arctiidae). J. Kans. Entomol. Soc. 1967, 40, 194–202. Available online: http://www.jstor.org/stable/25083620 (accessed on 30 December 2022).
- Szalay-Marzso, L. Biology and control of the fall webworm (Hyphantria cunea Drury) in the middle-and east European countries. EPPO Bull. 1971, 1, 23–31. [Google Scholar] [CrossRef]
- Sullivan, G.; Ozman-Sullivan, S. Tachinid (Diptera) parasitoids of Hyphantria cunea (Lepidoptera: Arctiidae) in its native North America and in Europe and Asia–a literature review. Entomol. Fenn. 2012, 23, 181–192. [Google Scholar] [CrossRef]
- Kuzmanova, I.; Sapundzhieva, K.; Chernev, T. Perspective strains of Bac. thuringiensis for controlling forest pests. Nauchni Tr. 1980, 25, 141–148. [Google Scholar]
- Vuković, S.; Tabaković-Tošić, M.; Nađalin, V.; Đurić, T.; Gološin, B. New approach in the suppression of the weaver tiger moth (Hyphantria cunea Drury) on fruit trees and gypsy moth (Lymantria dispar L.) in forests by bioinsecticide D-Stop based on Bacillus thuringiensis var. kurstaki. In Proceedings of the XII Symposium on plant protection and the application of pesticides, Zlatibor, Serbia, 25–29 November 2002; p. 139. [Google Scholar]
- Tabaković-Tošić, M.; Milanović, S.; Babović, K. Efficiency of the microbiological preparation D-Stop in the control of the fall webworm (Hyphantria cunea Drury). Inst. For. Proc. 2002, 46–47, 101–110. [Google Scholar]
- Tabaković-Tošić, M. Entomopathogenic Bacterium Bacillus thuringiensis ssp. kurstaki, the Important Component of the Integral Protection of Forest Ecosystems; Institute of Forestry: Belgrade, Serbia, 2008; pp. 46–66. [Google Scholar]
- Draganova, S. Virulence of strains of entomopathogenic fungi of the genus Beauveria Vuill. Biotechnol. Biotechnol. Equip. 1990, 4, 22–25. [Google Scholar] [CrossRef]
- Chkhubianishvili, T.; Mikaia, N.; Malania, I.; Kakhadze, M. Susceptibility of entomopathogenic nematodes to the fall webworm Hyphantria cunea Drury (Lepidoptera: Arctiidae). Bull. Georgian Natl. Acad. Sci. 2007, 175, 112–114. [Google Scholar]
- Stîngaci, A. Procedee biotehnologice de producere a preparatelor virale pentru combaterea defoliatorului omida păroasă a dudului (Hyphantria cunea Drury). Stud. Univ. Mold. (Stiinte R. Si Ale Nat.) 2013, 66, 98–104. [Google Scholar]
- Stîngaci, A. Entomopatgenic biopesticide in protecting Lepidoptera diversity in organic farming. In Proceedings of the Microbial Biotechnology, Kishinev, Moldovia, 11–12 October 2018; pp. 103–104. [Google Scholar]
- Stîngaci, A. Evidenţierea legităţilor de declanşare a epizootiilor baculovirale la H. Cunea. In Proceedings of the Protecţia Plantelor-Realizări Şi Perspective, Kishinev, Moldovia, 27–28 October 2020; pp. 197–201. [Google Scholar]
- Stîngaci, A.; Voloşciuc, L. Isolate locale ale baculovirului entomopatogenic ca o tehnologie de formulare inovatoare, care protejează biopesticidul din degradare a radiației ultraviolet. In Proceedings of the Genetica, Fiziologia Şi Ameliorarea Plantelor, Chişinău, Moldova, 4–5 October 2021; pp. 338–341. [Google Scholar]
- Tothill, J.D. Tachinidae and some Canadian hosts. Can. Entomol. 1913, 45, 69–75. [Google Scholar] [CrossRef]
- Burgess, A.F.; Crossman, S.S. Imported Insect Enemies of the Gipsy Moth and the Brown-Tail Moth; USDA: Washington, DC, USA, 1929.
- Burgess, A.F. The Gypsy Moth and the Brown-Tail Moth: A History of the Work for Prevention of Spread and Extermination of These Insects in North America; USDA: Washington, DC, USA, 1944.
- Elkinton, J.S.; Parry, D.; Boettner, G.H. Implicating an introduced generalist parasitoid in the invasive browntail moth’s enigmatic demise. Ecology 2006, 87, 2664–2672. [Google Scholar] [CrossRef]
- Louda, S.M.; Pemberton, R.W.; Johnson, M.T.; Follett, P. Nontarget effects—the Achilles’ heel of biological control? Retrospective analyses to reduce risk associated with biocontrol introductions. Annu. Rev. Entomol. 2003, 48, 365–396. [Google Scholar] [CrossRef] [PubMed]
- Purrini, K.; Weiser, J. Natürliche Feinde des Goldafters, Euproctis chrysorrhoea L., im Gebiet von Kosovo, FSR Jugoslawien. Anz. Für Schädlingskunde Pflanzenschutz Umweltschutz 1975, 48, 11–12. [Google Scholar] [CrossRef]
- Cory, J.S.; Hirst, M.L.; Sterling, P.H.; Speight, M.R. Narrow host range nucleopolyhedrovirus for control of the browntail moth (Lepidoptera: Lymantriidae). Environ. Entomol. 2000, 29, 661–667. [Google Scholar] [CrossRef]
- Delrio, G.; Luciano, P. The parasites and predators of Euproctis chrysorrhoea L. in Sardinia (preliminary note) . In Proceedings of the Atti XIV Congresso Nazionale Italiano di Entomologia, Palermo, Italy, 28 May–1 June 1985; pp. 825–832. [Google Scholar]
- Tabaković-Tošić, M.; Milosavljević, M. Entomopathogenic fungus Entomophaga aulicae as agents in classic biological control of browntail moth in some broadleaf forest in Serbia. IOBC-WPRS Bull. 2017, 129, 88–92. [Google Scholar]
- Tabaković-Tošić, M.; Milosavljević, M.; Jovanović, S.; Lučić, R. First record of entomopathogenic fungus Entomophaga aulicae in the populations of browntail moth in Bosnia and Herzegovina. Sustain. For. Collect. 2019, 79–80, 59–63. [Google Scholar] [CrossRef]
- Frago, E.; Pujade-Villar, J.; Guara, M.; Selfa, J. Hyperparasitism and seasonal patterns of parasitism as potential causes of low top-down control in Euproctis chrysorrhoea L. (Lymantriidae). Biol. Control 2012, 60, 123–131. [Google Scholar] [CrossRef]
- Solter, L.F.; Pilarska, D.K.; Vossbrinck, C.F. Host specificity of microsporidia pathogenic to forest Lepidoptera. Biol. Control 2000, 19, 48–56. [Google Scholar] [CrossRef]
- Pilarska, D.; Linde, A.; Goertz, D.; McManus, M.; Solter, L.; Bochev, N.; Rajkova, M. First report on the distribution of microsporidian infections of browntail moth (Euproctis chrysorrhoea L.) populations in Bulgaria. J. Pest Sci. 2001, 74, 37–39. [Google Scholar] [CrossRef]
- Pilarska, D.; Linde, A.; Solter, L.; Takov, D. New data on the biology of the microsporidium Endoreticulatus schubergi infecting the browntail moth, Euproctis chrysorrhoea (Lepidoptera: Lymantriidae). Acta Zool. Bulg. 2002, 54, 55–62. [Google Scholar]
- Sidor, C.; Dusanic, L.; Vujin, M. Most important diseases of European gold tail Euproctis chrysorrhoea L., provoked by microorganisms in a period 1971–1974 in SR Macedonia. Acta Entom. Jugosl. 1975, 11, 125–134. [Google Scholar]
- Sterling, P.H.; Speight, M.R. Comparative mortalities of the brown-tail moth, Euproctis chrysorrhoea (L.) (Lepidoptera: Lymantriidae), in south-east England. Bot. J. Linn. 1989, 101, 69–78. [Google Scholar] [CrossRef]
- Sterling, P.H.; Kelly, P.M.; Speight, M.R.; Entwistle, P.F. The generation of secondary infection cycles following the introduction of nuclear polyhedrosis virus to a population of the Brown-tail moth, Euproctis chrysorrhoea L. (Lep., Lymantriidae). J. Appl. Entomol. 1988, 106, 302–311. [Google Scholar] [CrossRef]
- Kelly, P.M.; Speight, M.R.; Entwistle, P.F. Mass production and purification of Euproctis chrysorrhoea (L.) nuclear polyhedrosis virus. J. Virol. Methods 1989, 25, 93–99. [Google Scholar] [CrossRef]
- Speight, M.R.; Kelly, P.M.; Sterling, P.H.; Entwistle, P.F. Field application of a nuclear polyhedrosis virus against the Brown-tail moth, Euproctis chrysorrhoea (L.) (Lep., Lymantriidae). J. Appl. Entomol. 1992, 113, 295–306. [Google Scholar] [CrossRef]
- Ciornei, C.; Netoiu, C.; Taur, I.; Vals, S.; Barca, M.; Vladescu, D. The use of viral preparations in biological control of defoliators Lymantria dispar (L.) and Euproctis chrysorrhoea (L.) in Romanian deciduous forests. In Proceedings of the IUFRO Working Party 7.03.10 Workshop on “Methodology of Forest Insect and Disease Survey in Central Europe”, Gmunden, Austria, 11–14 September 2006; Volume 14, pp. 284–291. [Google Scholar]
- Kniest, F.M.; Hoffman, J.R. Brown-tail moth, Euproctis chrysorrhoea, an indigenous pest of parks and public in the Benelux countries (Lepidoptera: Lymantriidae). Gt. Lakes Entomol. 2017, 17, 111–112. [Google Scholar]
- Lesko, K. Environment-friendly control measures against Euproctis chrysorrhoea. Erdészeti Kut. 1984, 76–77, 315–320. (In Hungarian) [Google Scholar]
- Currado, I.; Brussino, G. Control experiments with Bacillus thuringiensis Berl. in the forests of Piedmont. Dif. Delle Piante 1985, 8, 339–343. [Google Scholar]
- Canivet, J.-P.; Nef, L.; Lebrun, P. Utilisation combinée de Bacillus thuringiensis et d’insecticides chimiques à doses réduites contre Euproctis chrysorrhoea. Z. Für Angew. Entomol. 2009, 86, 85–97. [Google Scholar] [CrossRef]
- Miętkiewski, R. The possibility of using selected species of fungi for the control of larvae of the brown-tail (Euproctis chrysorrhoea L.). Rocz. Nauk. Rol. E (Ochr. Roślin) 1984, 14, 102–111. [Google Scholar] [CrossRef]
- Petrov, S. Checklist of Entomophaguses of the Scelionidae Family (Hymenoptera, Platygastroidea) Parasites on Eggs of Polyphagous Butterflies. Bulg. J. Agric. Sci. 2012, 11, 30–35. [Google Scholar]
- Magnoler, A. Field evaluation of a baculovirus against the larvae of Malacosoma neustria L. in Sardinia. Dif. Delle Piante 1985, 8, 329–337. [Google Scholar]
- Zanati, E.M. Promising egg parasites of Malacosoma neustria. Gorsko Stop. 1978, 34, 39–43. [Google Scholar]
- Jankevica, L.; Zarins, I. Biological control of Malacosoma neustria L. population with Latvian isolate of nuclear polyhedrosis virus. In Microbial insecticides: Novelty or necessity? In Proceedings of the Symposium Held at the University of Warwick, Coventry, UK, 17 July 1997; pp. 285–288. [CrossRef]
- Jankevica, L.; Tenbergs, G.; Zarinš, I. Biological control of the European tent caterpillar Malacosoma neustria L. (Lepidoptera, Lasiocampidae) population with different virus formulations. Latv. Entomol. 1998, 36, 11–16. [Google Scholar]
- Jankevica, L.; Zarinš, I. Virulence of Malacosoma neustria nucleopolyhedrovirus Latvian isolates. Latv. Entomol. 1999, 37, 40–45. [Google Scholar]
- Jankevica, L. Persistence and accumulation of Malacosoma neustria nuclear polyhedrosis virus in the ecosystem after virus application. IOBC/wprs Bull. 2000, 23, 249–252. [Google Scholar]
- Jankevica, L. Increasing the susceptibility of the Malacosoma neustria larvae to nucleopolyhedrovirus by a synergistic additive. Latv. Entomol. 2003, 40, 64–66. [Google Scholar]
- Draganova, S.; Takov, D.; Pilarska, D.; Doychev, D.; Mirchev, P.; Georgiev, G. Fungal pathogens on some lepidopteran forest pests in Bulgaria. Acta Zool. Bulg. 2013, 65, 179–186. [Google Scholar]
- Ruiu, L.; Mannu, R.; Falchi, G.; Braggio, A.; Luciano, P. Evaluation of different Bacillus thuringiensis sv kurstaki formulations against Lymantria dispar and Malacosoma neustria larvae infesting Quercus suber trees. Redia 2013, 96, 27–31. [Google Scholar]
- Ruiu, L.; Mannu, R.; Falchi, G.; Braggio, A.; Luciano, P. Two years of efficacy trials with different Bacillus thuringiensis sv kurstaki formulations against Lymantria dispar and Malacosoma neustria. IOBC/wprs Bull. 2014, 101, 205–211. [Google Scholar]
- Tabakovic-Tosic, M.; Milosavljevic, M. Studies on Non-target Phyllophagous Lepidoptera in Some Oak Forests in Đerdap National Park as Potential host of Entomophaga maimaiga. In Proceedings of the International Conference on Agriculture, Forests, Food Sciences and Technologies, Izmir, Turkey, 2–5 April 2018. [Google Scholar]
- Georgieva, M.; Takov, D.; Georgiev, G.; Pilarska, D.; Pilarski, P.; Mirchev, P.; Humber, R. Studies on non-target phyllophagous insects in oak forests as potential hosts of Entomophaga maimaiga (Entomophthorales: Entomophthoraceae) in Bulgaria. Acta Zool. Bulg. 2014, 66, 115–120. [Google Scholar]
- Werno, J.; Lamy, M.; Vincendeau, P. Caterpillar hairs as allergens. Lancet 1993, 342, 936–937. [Google Scholar] [CrossRef]
- Rossi, J.P.; Imbault, V.; Lamant, T.; Rousselet, J. A citywide survey of the pine processionary moth Thaumetopoea pityocampa spatial distribution in Orléans (France). Urban For. Urban Green. 2016, 20, 71–80. [Google Scholar] [CrossRef]
- Roques, A.; Rousselet, J.; Avcı, M.; Avtzis, D.N.; Basso, A.; Battisti, A.; Ben Jamaa, M.L.; Bensidi, A.; Berardi, L.; Berretima, W.; et al. Climate warming and past and present distribution of the processionary moths (Thaumetopoea spp.) in Europe, Asia Minor and North Africa. In Processionary Moths and Climate Change: An Update; Springer: Dordrecht, The Netherlands, 2015; pp. 81–161. [Google Scholar] [CrossRef]
- De Boer, J.G.; Harvey, J.A. Range-Expansion in Processionary Moths and Biological Control. Insects 2020, 11, 267. [Google Scholar] [CrossRef]
- Tsankov, G.; Schmidt, G.H.; Mirchev, P. Impact of parasitoids in egg-batches of Thaumetopoea pityocampa (Den. & Schiff.) in Algeria. Boll. Di Zool. Agrar. E Di Bachic. 1995, 27, 53–60. [Google Scholar]
- Tsankov, G.; Schmidt, G.H.; Mirchev, P. Parasitism of egg-batches of the pine processionary moth Thaumetopoea pityocampa (Den. & Schiff.) (Lep., Thaumetopoeidae) in various regions of Bulgaria. J. Appl. Entomol. 1996, 120, 93–105. [Google Scholar] [CrossRef]
- Tsankov, G.; Douma-Petridou, E.; Mirchev, P.; Georgiev, G.; Koutsaftikis, A. Spectrum of egg parasitoids and rate of parasitism of egg batches of the pine processionary moth Thaumetopoea pityocampa (Den. & Schiff.) in the Northern Peloponnes/Greece. J. Entomol. Res. Soc. 1999, 1, 1–8. [Google Scholar]
- Schmidt, G.H.; Tsankov, G.; Mirchev, P. Notes on the egg parasitoids of Thaumetopoea pityocampa (Den. & Schiff.) (Insecta Lepidoptera Thaumetopoeidae) collected on the Greek island Hydra. Bolletiona Zool. Agrar. Di Bachic. Ser II 1997, 29, 91–99. [Google Scholar]
- Mirchev, P.; Tsankov, G. Checklist of the parasitoids and predators on Thaumetopoea spp. (Lepidoptera: Notodontidae). Acta Entomol. Bulg. 2005, 11, 82–96. [Google Scholar]
- Mirchev, P.; Georgiev, G.; Boyadzhiev, P.; Matova, M. Impact of entomophages on density of Thaumetopoea pityocampa in egg stage near Ivaylovgrad, Bulgaria. Acta Zool. Bulg. 2012, 4, 103–110. [Google Scholar]
- Mirchev, P.; Georgiev, G.; Boyadzhiev, P. Comparative studies on egg parasitoids of pine processionary moth (Thaumetopoea pityocampa) in a 20-year period. For. Sci. 2015, 2, 105–114. [Google Scholar]
- Battisti, A.; Bernardi, M.; Ghiraldo, C. Predation by the hoopoe (Upupa epops) on pupae of Thaumetopoea pityocampa and the likely influence on other natural enemies. BioControl 2000, 45, 311–323. [Google Scholar] [CrossRef]
- Ince, I.A.; Demir, I.; Demirbag, Z.; Nalcacioglu, R. A cytoplasmic polyhedrosis virus isolated from the pine processionary caterpillar, Thaumetopoea pityocampa. J. Microbiol. Biotechnol. 2007, 17, 632–637. [Google Scholar] [PubMed]
- Sevim, A.; Demir, I.; Demirbağ, Z. Molecular characterization and virulence of Beauveria spp. from the pine processionary moth, Thaumetopoea pityocampa (Lepidoptera: Thaumetopoeidae). Mycopathologia 2010, 170, 269–277. [Google Scholar] [CrossRef]
- Masutti, L.; Battisti, A. Thaumetopoea pityocampa (Den. & Schiff.) in Italy: Bionomics and perspectives of integrated control. J. Appl. Entomol. 1990, 110, 229–234. [Google Scholar] [CrossRef]
- Battisti, A.; Longo, S.; Tiberi, R.; Triggiani, O. Results and perspectives in the use of Bacillus thuringiensis Berl. var. kurstaki and other pathogens against Thaumetopoea pityocampa (Den. et Schiff.) in Italy (Lep., Thaumetopoeidae). Anz. Schädlingskd. Pfl. Umwelt 1998, 71, 72–76. [Google Scholar] [CrossRef]
- Triggiani, O.; Tarasco, E. Preliminary attempts to control overwintering populations of Thaumetopoea pityocampa (Den. et Schiff.) (Lepidoptera: Thaumetopoeidae) with Steinernema feltiae (Filipjev, 1934) (Nematoda: Steinernematidae). Entomologica 2001, 35, 7–15. [Google Scholar] [CrossRef]
- Colacci, M.; Kavallieratos, N.G.; Athanassiou, C.G.; Boukouvala, M.C.; Rumbos, C.I.; Kontodimas, D.C.; Pardo, D.; Sancho, J.; Benavent-Fernández, E.; Gálvez-Settier, S.; et al. Management of the pine processionary moth, Thaumetopoea pityocampa (Lepidoptera: Thaumetopoeidae), in urban and suburban areas: Trials with trunk barrier and adhesive barrier trap devices. J. Econ. Entomol. 2018, 111, 227–238. [Google Scholar] [CrossRef]
- Trematerra, P.; Colacci, M.; Athanassiou, C.G.; Kavallieratos, N.G.; Rumbos, C.I.; Boukouvala, M.C.; Nikolaidou, A.J.; Kontodimas, D.C.; Benavent-Fernández, E.; Gálvez-Settier, S. Evaluation of mating disruption for the control of Thaumetopoea pityocampa (Lepidoptera: Thaumetopoeidae) in suburban recreational areas in Italy and Greece. J. Econ. Entomol. 2019, 112, 2229–2235. [Google Scholar] [CrossRef]
- Barta, M.; Horáková, M.K.M.; Georgieva, M.; Mirchev, P.; Zaemdzhikova, G.; Pilarska, D.; Takov, D.; Todorov, M.; Hubenov, Z.; Pilarski, P.; et al. Entomopathogenic fungi (Ascomycota: Hypocreales) as natural antagonists of the pine processionary moth Thaumetopoea pityocampa (Denis & Schiffermüller, 1775) (Lepidoptera: Notodontidae) in Bulgaria. Acta Zool. Bulg. 2020, 15, 89–96. [Google Scholar]
- Csóka, G.; Hirka, A.; Szőcs, L.; Móricz, N.; Rasztovits, E.; Pödör, Z. Weather-dependent fluctuations in the abundance of the oak processionary moth, Thaumetopoea processionea (Lepidoptera: Notodontidae). Eur. J. Entomol. 2018, 115, 249–255. [Google Scholar] [CrossRef]
- Ceianu, I.; Dissescu, G. The role of entomophagous insects in an outbreak area of Thaumetopoea processionea treated with bacterial preparations. Cerc. Ecol. Anim. Lucrari Prim. Ses. Stiint. Tara Ecol. Anim. 1969, 1966, 183–190. [Google Scholar]
- Sands, R.J.; Kitson, J.J.N.; Raper, C.M.; Jonusas, G.; Straw, N. Carcelia iliaca (Diptera: Tachinidae), a specific parasitoid of the oak processionary moth (Lepidoptera: Thaumetopoeidae), new to Great Britain. Br. J. Entomol. Nat. Hist. 2015, 28, 225–228. [Google Scholar]
- Zeegers, T. Tachinid flies (Diptera: Tachinidae) from Dutch oak processionary. Entomolog. Ber. 1997, 57, 73–78. [Google Scholar]
- Bin, F.; Tiberi, R. Preliminary notes on the egg parasites of Thaumetopoea processionea (L.) in central Italy (Hym., Chalcidoidea; Lep., Thaumetopoeidae). Redia 1983, 66, 449–459. [Google Scholar]
- Mirchev, P.; Georgiev, G.; Balov, S.; Kirilova, M.; Georgieva, A. Distribution of Thaumetopoea processionea (L.) in Bulgaria. Silva Balc. 2011, 12, 71–80. [Google Scholar]
- Wagenhoff, E.; Veit, H. Five years of continuous Thaumetopoea processionea monitoring: Tracing population dynamics in an arable landscape of South-Western Germany. Gesunde Pflanz. 2011, 63, 51–61. [Google Scholar] [CrossRef]
- Hoch, G.; Verucchi, S.; Schopf, A. Microsporidian pathogens of the oak processionary moth, Thaumetopoea processionea (L.) (Lep., Thaumetopoeidae), in eastern Austria’s oak forests. Mitt. Dtsch. Ges. Allg. Angew. Ent. 2008, 16, 225–228. [Google Scholar]
- Pascual, J.A.; Robredo, F.; Galante, E. Aerial ULV applications of alpha-cypermethrin, diflubenzuron and Bacillus thuringiensis against the oak processionary moth (Thaumetopoea processionea) (Lep., Thaumetopoeidae). Bol. San. Veg. Plagas 1990, 16, 585–591. [Google Scholar]
- Flemming, G. First experiences at control of Thaumetopoea processionea in the north-western region of the Land Brandenburg in the year 1996. Gesunde Pflanz. 1997, 49, 54–57. [Google Scholar]
- Roversi, P.F.; Faggi, M.; Rumine, P. Bacillus thuringiensis var. kurstaki against Thaumetopoea processionea L.: Field trials in Quercus cerris L. woods. IOBC/wprs Bull. 2005, 28, 203–209. [Google Scholar]
- Roversi, P.F. Aerial spraying of Bacillus thuringiensis var. kurstaki for the control of Thaumetopoea processionea in Turkey oak woods. Phytoparasitica 2008, 36, 175–186. [Google Scholar] [CrossRef]
- Straw, N.A.; Forster, J. The effectiveness of ground-based applications of Bacillus thuringiensis var. kurstaki for controlling oak processionary moth Thaumetopoea processionea (Lepidoptera: Thaumetopoeidae). Ann. Appl. Biol. 2021, 181, 48–57. [Google Scholar] [CrossRef]
- Groenen, F. Variation of Thaumetopoea processionea (Notodontidae: Thaumetopoeinae) in Europe and the Middle East. Entomolog. Ber. 2010, 70, 77–82. [Google Scholar]
- Cosic, H.T.; Colombel, E.; Tabone, E.; Cosic, B.; Venard, M. An Egg Parasitoid in the Oak Processionary Moth Management; IOBC-OILB: Hammamet, Tunisia, 2022. [Google Scholar]
- Parker, D.J.; Schmidt, A.C. Apple ermine moth, Hyponomeuta malinellus. Report for Agriculture Agri-Food Canada, Plant Health Division. 159. E. Junnikkala (1960) Life history and insect enemies of Hyponomeuta malinellus Zell (Lep., Hyponomeutidae) in Finland. Ann. Zool. Soc. 1998, 21, 1–44. [Google Scholar]
- Junnikkala, E. Life History and Insect Enemies of Hyponomeuta malinellus Zell. (Lep., Hyponomeutidae) in Finland. Anz. Schädlingskd. 1961, 34, 176. [Google Scholar] [CrossRef]
- Mason, P.G.; Huber, J.T. Biological Control Programmes in Canada, 1981–2000; CABI Publishing: New York, NY, USA, 2002. [Google Scholar]
- Dobesberger, E.; Garland, J.A. Plant Health Risk Assessment Yponomeuta malinellus (Apple Ermine Moth); PRA request No. 98-48, 2009 (Update No. 2); Canadian Food Inspection Agency: Ottawa, ON, Canada, 2009. [Google Scholar]
- Cossentine, J.E.; Kuhlmann, U. Status of Ageniaspis fuscicollis (Hymenoptera: Encyrtidae), an introduced parasitoid of the apple ermine moth (Lepidoptera: Yponomeutidae). Can. Entomol. 2000, 132, 685–689. [Google Scholar] [CrossRef]
- Cossentine, J.; Kuhlmann, U. Yponomeuta malinellus Zeller, Apple Ermine Moth (Lepidoptera: Yponomeutidae). In Biological Control Programmes in Canada 1981–2000; Mason, P.G., Huber, J.T., Eds.; CABI Publishing: New York, NY, USA, 2002; pp. 275–278. [Google Scholar]
- Smith, R. Biological Control of the Apple Ermine Moth in Southwestern British Columbia; Report prepared for the British Columbia Ministry of Agriculture; Fisheries and Food and Agriculture and Agri-food Canada: Ottawa, ON, Canada, 1990. [Google Scholar]
- Hérard, F.; Prévost, G. Suitability of Yponomeuta malinellus and Y. cagnagellus (Lepidoptera: Yponomeutidae) as hosts of Diadegma armillata (Hymenoptera: Ichneumonidae). Environ. Entomol. 1997, 26, 933–938. [Google Scholar] [CrossRef]
- Affolter, F.; Carl, K.P. The Natural Enemies of the Apple Ermine Moth Yponomeuta malinellus in Europe; Report of the CAB International; Institute of Biological Control: Delémont, Switzerland, 1986. [Google Scholar]
- Hoch, G.; Pilarska, D.K.; Dobart, N. Effect of midgut infection with the microsporidium Endoreticulatus schubergi on carbohydrate and lipid levels in Lymantria dispar larvae. J. Pest Sci. 2009, 82, 351–356. [Google Scholar] [CrossRef]
- Žikić, V.; Lotfalizadeh, H.; Schwarz, M.; Stanković, S.S.; Lazarević, M.; Kos, K.; Rakhshani, E.; Tschorsnig, H.P. Parasitoids of European species of the genus Yponomeuta Latreille 1796 (Lepidoptera: Yponomeutidae): New findings with an updated checklist. Phytoparasitica 2018, 46, 617–626. [Google Scholar] [CrossRef]
- Piekarska-Boniecka, H.; Rzańska-Wieczorek, M.; Siatkowski, I.; Barczak, T. Parasitisation of Yponomeuta malinellus feeding on Crataegus monogyna in the allotment gardens in the city of Poznań, Poland. Plant Prot. Sci. 2022, 58, 150–157. [Google Scholar] [CrossRef]
- Cory, J.S.; Myers, H.J. The ecology and evolution of insect baculoviruses. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 239–272. [Google Scholar] [CrossRef]
- Tabakovic-Tosic, M. Chapter 32: Biological control of harmful forest insects: The biological efficiency of microbial insecticides and possibility of their application in forest ecosystems. In Biopesticides in Organic Farming—Recent Advances, Section 15: Management of the Diseases of Crops through Biotic Biopesticides (Natural enemies) Biological Control; Awasthi, L.P., Ed.; Taylor & Francis Group: Abingdon, UK; CRC Press: Boca Raton, FL, USA, 2021; pp. 203–206, 404. [Google Scholar]
- Waage, J.K.; Greathead, D.J. Biological control: Challenges and opportunities. Philos. Trans. R. Soc. 1988, 318, 111–128. [Google Scholar] [CrossRef]


{kind=link}
| Taxon 1 | Host Plant 2 |
|---|---|
| Yponomeutidae | |
| Yponomeuta cagnagella (Hübner) * | Euonymus sp. |
| Y. evonymella (L.) * | Prunus padus, P. serotina |
| Y. gigas Rebel * | Salix sp., Populus sp. |
| Y. irrorella (Hübner) | Euonymus sp. |
| Y. mahalebella Guenée * | Prunus mahaleb |
| Y. malinellus Zeller ** | Malus sp., Pyrus sp. |
| Y. padella (L.) * | Rosaceae |
| Y. plumbella (Denis and Schiffermüller) * | Euonymus sp. |
| Y. rorrella (Hübner) * | Salix sp., Populus sp. |
| Y. sedella Treitschke | Crassulaceae |
| Scythropia crataegella L. * | Rosaceae |
| Lasiocampidae | |
| Malacosoma alpicola Staudinger | polyphagous on herbs |
| M. castrensis (L.) | polyphagous on herbs |
| M. franconica (Denis and Schiffermüller) * | polyphagous on herbs |
| M. laurae Lajonquière | polyphagous on herbs |
| M. neustria (L.) ** | polyphagous on woody Aceraceae, Rosaceae, Salicaceae, Fagaceae, Ulmaceae and Oleaceae |
| Eriogaster arbusculae Freyer | alpine Salicaceae, Betulacae, Rosacae and Ericaceae |
| E. catax (L.) | Prunus spinosa, Crataegus sp., Quercus sp., Salix sp., Ulmus sp. |
| E. lanestris (L.) * | woody Rosaceae and Betulaceae, Corylaceae, Salicaceae and Tiliaceae |
| E. rimicola (Denis and Schiffermüller) Chondrostega vandalicia (Millière) C. pastrana Lederer | Quercus sp. polyphagous on herbs polyphagous on herbs |
| Notodontidae | |
| Phalera bucephala (L.) * | Fagaceae, Ulmaceae, Aceraceae, Betulaceae, Salicaceae, Tiliaceae, Caprifoliaceae |
| P. bucephaloides (Ochsenheimer) * | Quercus sp. |
| T. pinivora (Treitschke) * | Pinaceae, Cupressaceae |
| T. pityocampa (Denis and Schiffermüller) ** | Pinaceae |
| T. processionea (L.) ** | Fagaceae, Juglandaceae |
| T. solitaria (Freyer) ** | Pistacia sp. |
| T. wilkinsoni Tams ** | Pinaceae, Cupressaceae |
| Erebidae | |
| Ocnogyna corsica (Rambur) | polyphagous on herbs and grasses |
| O. baetica **(Rambur) | Fabaceae, Apiaceae, Poaceae |
| Euproctis chrysorrhoea (L.) ** | polyphagous on woody Rosaceae, also on Aceraceae, Betulaceae, Fagaceae, Juglandaceae, Oleaceae, Eleagnaceae, Vitaceae, Grossulariaceae and others |
| E. similis Fuessly * | polyphagous on woody Rosaceae, also on Betulaceae and Fagaceae |
| Noctuidae | |
| Oxicesta chamoenices (Herrich-Schäffer) | Euphorbia sp. |
| O. geographica (Fabricius) | Euphorbia sp. |
| O. serratae (Zerny) | Euphorbia sp. |
| Endromidae | |
| Endromis versicolora (L.) | Betulaceae, Rosaceae, Tiliaceae |
| Saturniidae | |
| Saturnia pavonia (L.) | polyphagous in herb and shrub layer |
| S. pavoniella (Scopoli) | polyphagous in herb and shrub layer |
| S. spini (Denis and Schiffermüller) | Prunus spinosa and other Rosaceae |
| Nymphalidae | |
| Aglais io L. | Urtica sp. |
| A. urticae L. | Urtica sp. |
| A. ichnusa (Bonelli) | Urtica sp. |
| M. arduinna (Esper) | Centaurea sp. |
| M. athalia (Rottemburg) | Plantaginaceae, Scrophulariaceae |
| M. aurelia Nickerl | Plantaginaceae |
| M. britomartis Assmann | Plantaginaceae, Scrophulariaceae |
| M. cinxia L. | Plantaginaceae, Asteraceae, Scrophulariaceae |
| M. deione Geyer | Plantaginaceae |
| M. diamina (Lang) | Scrophulariaceae, Valerianaceae, Plantaginaceae |
| M. didyma (Esper) | Scrophulariaceae, Valerianaceae, Plantaginaceae |
| M. parthenoides Keferstein | Plantaginaceae, Scrophulariaceae |
| M. phoebe (Denis and Schiffermüller) | Asteraceae |
| M. telona Frühstorfer | Cirsium sp., Centaurea sp. |
| M. trivia (Denis and Schiffermüller) | Verbascum sp. |
| M. varia Meyer-Dür | Plantaginaceae, Scrophulariaceae |
| Euphydryas aurinia (Rottemburg) | Caprifoliaceae, Gentianaceae, Dipsacaceae |
| E. cynthia (Denis and Schiffermüller) | Plantago alpina, Viola calcarata and others |
| E. desfontainii (Godart) | Dipsacaceae |
| E. iduna (Dalman) | Scrophulariaceae, Plantaginaceae |
| E. intermedia (Ménétriés) | Caprifoliaceae, Scrophulariaceae |
| E. maturna (L.) | Fraxinus excelsior, Scrophulariaceae, Plantaginaceae, Valerianaceae, Oleaceae, Adoxaceae |
| Araschnia levana (L.) | Urticaceae |
| Nymphalis antiopa (L.) | Salicaceae, Betulaceae, Ulmaceae, Aceraceae, Oleaceae, Cannabaceae, Moraceae, Corylaceae, Rosaceae, Tiliaceae, Urticacae |
| N. polychloros (L.) | Salicaceae, Ulmaceae, Rosaceae, |
| N. vaualbum (Denis and Schiffermüller) | Betulacae, Salicacae, Ulmaceae |
| N. xanthomelas (Esper) | Salicaceae, Ulmaceae, Anacardiaceae |
| Pieridae | |
| Pieris brassicae (L.) ** | Brassicaceae, Resedaceae, Solanaceae, Tropaeolaceae |
| Aporia crataegi (L.) ** | woody Rosaceae and Betulaceae, Juglandaceae, Fagaceae, Salicaceae, Ericaceae |
| Taxon | Geographical Origin | Market Sector * | Outbreak Region(s) ** References |
|---|---|---|---|
| Hyphantria cunea (Erebidae) | Nearctic | pome fruits, stone fruits, grapes, vegetables, ornamental trees, forestry | Europe [36,37] |
| Asia [38,39,40,41] | |||
| New Zealand [42] | |||
| Euproctis chrysorrhoea (Erebidae) | Europe | pome fruits, stone fruits, forestry | Europe [43,44,45] |
| North America [46,47] Asia [48] | |||
| Malacosoma neustria (Lasiocampidae) | Paleartic | pome fruits, ornamental shrubs, stone fruits, ornamental trees | Europe [49,50] |
| Asia [51,52,53] | |||
| Thaumetopoea pityocampa (Notodontidae) | Paleartic | forestry, livestock farming | Europe [54,55] |
| Thaumetopoea processionea (Notodontidae) | Europe | forestry, livestock farming | Europe [56,57] |
| Yponomeuta malinellus (Yponomeutidae) | Paleartic, Indomalayan | pome fruit, orchards, ornamental trees | Europe [58] |
| North America [59,60] | |||
| Asia [61,62,63] |
| Taxon | Research | Application | |||
|---|---|---|---|---|---|
| RNR | BCAS | OPT | FE | OFI | |
| H. cunea | / | 1 [83] | 4 [80,84,85,86,87,88,89] | 1 [81,82,83] | / |
| E. chrysorrhoea | 7 [93,94,95,96,97,98,99,100,104] | / | 3 [106,107,127] | 4 [108,111,112,114] | 2 [109,113] |
| M. neustria | 3 [50,117,123] | / | 1 [49] | 3 [116,118,119,120,121,122,124] | 1 [125] |
| T. pityocampa | 3 [132,133,134,135,136,137,138,144,147] | / | / | 2 [83,144] | 2 [142,143] |
| T. processionea | 7 [150,151,152,153,154,155,163] | / | 1 [162] | 2 [156,158] | 2 [157,160] |
| Y. malinellus | 4 [163,171,172,173,174] | 2 [59,60] | 2 [169,170] | / | 2 [165,166] |
| Subcategory | Reference(s) * | Objective | Outcome ** |
|---|---|---|---|
| Biological control agent selection (BCAS) | |||
| 1 | [59] | selecting agents for intracontinental introduction, export | successful |
| 2 | [60] | selecting agents for intracontinental introduction, export | successful |
| 3 | [81] | comparing novel bioinsecticide efficiency to the standard | successful, commercially applied |
| Optimization and susceptibility (OPT) | |||
| 1 | [80] | selecting perspective strains | discontinued |
| 2 | [84] | pathogenicity testing | successful, not implemented * |
| 3 | [85] | pathogenicity testing | successful, not implemented * |
| 4 | [86,87,88,89] | larvicidal potential and formulation testing | successful, commercially applied |
| 5 | [106] | transmission and persistence testing | successful, continued |
| 6 | [107] | mass production | successful, cost-ineffective |
| 7 | [127] | confirming host specificity | successful, implemented |
| 8 | [49] | investigating relative susceptibility | successful, implemented |
| 9 | [162] | selecting agents and mass production | unknown |
| 10 | [169] | susceptibility in different phases of development | successful, implementation unknown |
| 11 | [170] | collecting BCAs from alternate host | successful, implementation unknown |
| Field experiments (FE) | |||
| 1 | [81,82,83] | bioinsecticide efficiency testing | successful, commercially applied |
| 2 | [108] | formulation efficiency testing | successful, application unknown |
| 3 | [114] | strain efficiency testing | unknown |
| 4 | [111] | synergistic effects investigation | recommendations for further research |
| 5 | [112] | efficiency testing | successful, implemented |
| 6 | [116] | laboratory trial confirmation | successful |
| 7 | [118,119,120,121,122] | pathogenicity, susceptibility, synergistic effects | successful, continued |
| 8 | [124] | formulation testing, comparison to standard | successful, commercially applied |
| 9 | [83] | bioinsecticide efficiency testing | successful, commercially applied |
| 10 | [144] | pathogenicity testing | recommendations for further research |
| 11 | [156] | susceptibility, comparison with standards | further adjustments recommended |
| 12 | [158] | commercial formulation testing | successful, recommendations for further research |
| Open field implementation (OFI) | |||
| 1 | [109] | formulation efficiency testing | successful, implementation unknown |
| 2 | [113] | dosage variation testing | unknown |
| 3 | [125] | impact evaluation | successful, continued |
| 4 | [142] | report | successful, continued |
| 5 | [143] | methodology variation testing | recommendations for further research |
| 6 | [157] | routine efficiency estimation, commercial | successful, continued |
| 7 | [160] | commercial formulation testing | successful, recommendations for further research |
| 8 | [165] | agent introduction | successful, continued |
| 9 | [166] | agent introduction | successful, continued |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trajković, A.; Žikić, V. Stuck in the Caterpillars’ Web: A Half-Century of Biocontrol Research and Application on Gregarious Lepidopteran Pests in Europe. Sustainability 2023, 15, 2881. https://doi.org/10.3390/su15042881
Trajković A, Žikić V. Stuck in the Caterpillars’ Web: A Half-Century of Biocontrol Research and Application on Gregarious Lepidopteran Pests in Europe. Sustainability. 2023; 15(4):2881. https://doi.org/10.3390/su15042881
Chicago/Turabian StyleTrajković, Aleksandra, and Vladimir Žikić. 2023. "Stuck in the Caterpillars’ Web: A Half-Century of Biocontrol Research and Application on Gregarious Lepidopteran Pests in Europe" Sustainability 15, no. 4: 2881. https://doi.org/10.3390/su15042881