

Assessment of Mound Soils Bacterial Community of the Red Imported Fire Ant, Solenopsis invicta across Guangdong Province of China

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
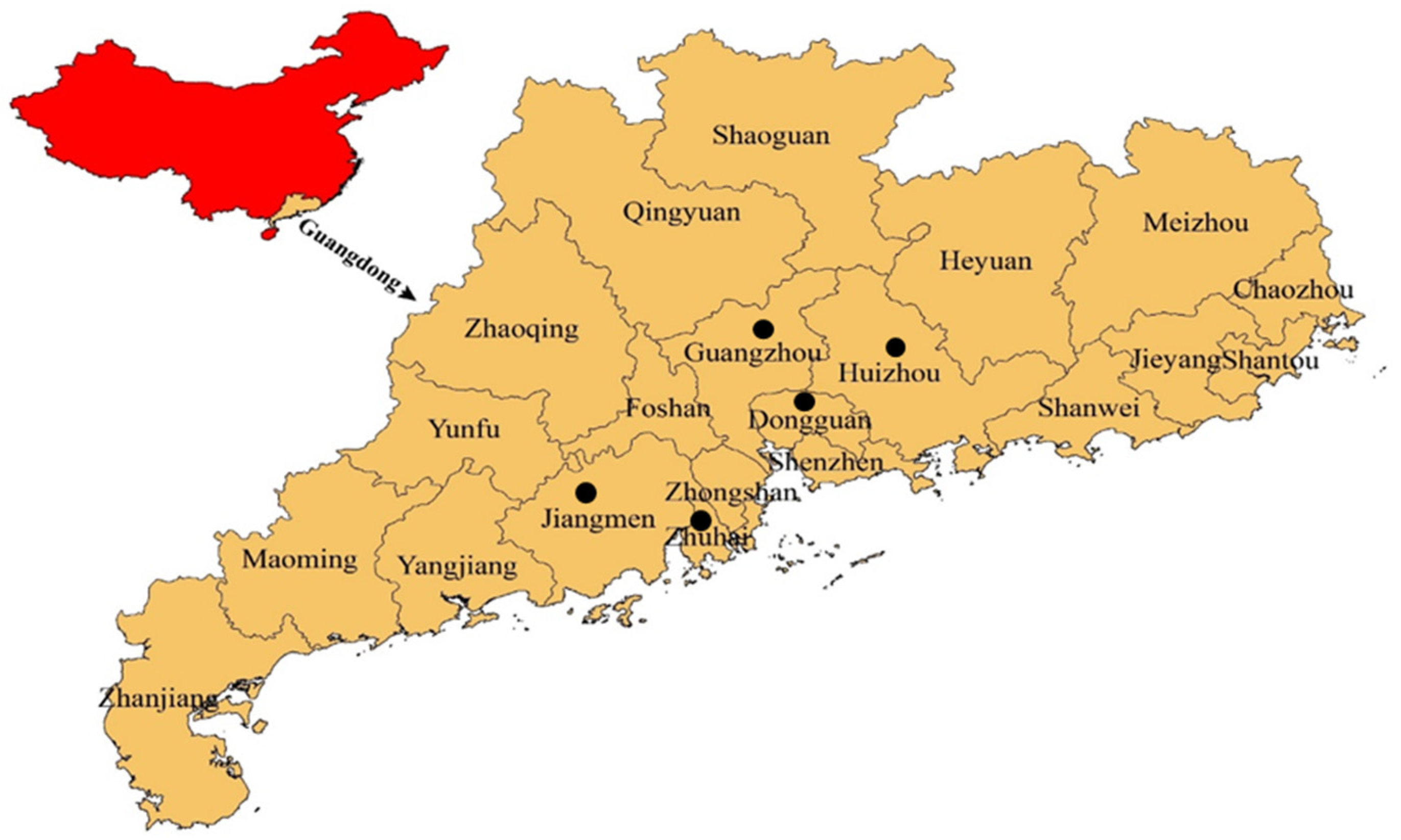
2.1. Sampling Location
2.2. Media Preparation
2.3. Procedure for Bacteria Isolation from Soil Samples
2.4. Bacteria Isolation from Mound Plant Debris
2.5. Bacteria Isolation from Ant Bodies
2.6. Morphological Characterization
2.7. Molecular Identification
2.8. Sequence Alignments and Phylogenetic Analysis
2.9. Statistical Analysis
3. Results
3.1. Morphological Characterization of Isolated Bacterial Strains
3.2. Molecular Identification and Phylogenetic Placement of Isolates
3.3. Analysis of the Diversity, Richness, and Densities of the Culturable Bacterial Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Travanty, N.V.; Vargo, E.L.; Apperson, C.S.; Ponnusamy, L. Colonization by the red imported fire ant, Solenopsis invicta, modifies soil bacterial communities. Microb. Ecol. 2022, 84, 240–256. [Google Scholar] [CrossRef] [PubMed]
- Ishak, H.D.; Plowes, R.; Sen, R.; Kellner, K.; Meyer, E.; Estrada, D.A.; Dowd, S.; Mueller, U.G. Bacterial diversity in Solenopsis invicta and Solenopsis geminata ant colonies characterized by 16S amplicon 454 pyrosequencing. Microb. Ecol. 2011, 61, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Arp, A.P.; Chapman, R.; Crosslin, J.M.; Bextine, B. Low-level detection of Candidatus Liberibacter solanacearum in Bactericera cockerelli (Hemiptera: Triozidae) by 16S rRNA Pyrosequencing. Environ. Entomol. 2013, 42, 868–873. [Google Scholar] [CrossRef] [PubMed]
- Powell, C.M.; Hanson, J.D.; Bextine, B.R. Bacterial community survey of Solenopsis invicta Buren (red imported fire ant) colonies in the presence and absence of Solenopsis invicta virus (SINV). Curr. Microbiol. 2014, 69, 580–585. [Google Scholar] [CrossRef]
- Werren, J.H. Symbionts provide pesticide detoxification. Proc. Natl. Acad. Sci. USA 2012, 109, 8364–8365. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.H.; Husseneder, C.; Hooper-Bùi, L. Culture-independent identification of gut bacteria in fourth-instar red imported fire ant, Solenopsis invicta Buren, larvae. J. Invertebr. Pathol. 2008, 98, 20–33. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Medina, F.; Vinson, S.B.; Coates, C.J. Isolation, characterization, and molecular identification of bacteria from the red imported fire ant (Solenopsis invicta) midgut. J. Invertebr. Pathol. 2005, 89, 203–209. [Google Scholar] [CrossRef]
- Tufts, D.M.; Bextine, B. Identification of bacterial species in the hemolymph of queen Solenopsis invicta (Hymenoptera: Formicidae). Environ. Entomol. 2009, 38, 1360–1364. [Google Scholar] [CrossRef]
- Peloquin, J.J.; Greenberg, L. Identification of midgut bacteria from fourth instar red imported fire ant larvae, Solenopsis invicta Buren (Hymenoptera: Formicidae). J. Agric. Urban Entomol. 2003, 20, 157–164. [Google Scholar]
- Beard, C.B.; Cordon-Rosales, C.; Durvasula, R.V. Bacterial symbionts of the triatominae and their potential use in control of Chagas disease transmission. Annu. Rev. Entomol. 2002, 47, 123. [Google Scholar] [CrossRef] [Green Version]
- Bextine, B.; Lauzon, C.; Potter, S.; Lampe, D.; Miller, T.A. Delivery of a genetically marked Alcaligenes sp. to the glassy-winged sharpshooter for use in a paratransgenic control strategy. Curr. Microbiol. 2004, 48, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Porter, S.D.; Alonso, L.E. Host specificity of fire ant decapitating flies (Diptera: Phoridae) in laboratory oviposition tests. J. Econ. Entomol. 1999, 92, 110–114. [Google Scholar] [CrossRef]
- Williams, D.F.; Oi, D.H.; Knue, G.J. Infection of red imported fire ant (Hymenoptera: Formicidae) colonies with the entomopathogen Thelohania solenopsae (Microsporidia: Thelohaniidae). J. Econ. Entomol. 1992, 92, 830–836. [Google Scholar] [CrossRef]
- Baird, R.; Woolfolk, S.; Watson, C. Survey of bacterial and fungal associates of black/hybrid imported fire ants from mounds in Mississippi. Southeast Nat. 2007, 6, 615–632. [Google Scholar] [CrossRef]
- Beckham, R.; Bilimoria, S.; Bartell, D. A survey for microorganisms associated with ants in western Texas. Southwest Entomol. 1982, 7, 225–229. [Google Scholar]
- Jouvenaz, D.; Allen, G.; Banks, W.; Wojcik, D.P. A survey for pathogens of fire ants, Solenopsis spp., in the southeastern United States. Fla. Entomol. 1977, 60, 275–279. [Google Scholar] [CrossRef]
- Woolfolk, S.; Stokes, C.E.; Watson, C.; Baker, G.; Brown, R.; Baird, R. Fungi associated with Solenopsis invicta Buren (Red imported fire ant, Hymenoptera: Formicidae) from mounds in Mississippi. Southeast Nat. 2016, 15, 220–234. [Google Scholar] [CrossRef]
- Woolfolk, S.; Stokes, C.E.; Watson, C.; Brown, R.; Baird, R. Bacteria associated with red imported fire ants (Solenopsis invicta) from mounds in Mississippi. Southeast Nat. 2016, 15, 83–101. [Google Scholar] [CrossRef]
- Zettler, J.A.; Mcinnis, T.M., Jr.; Allen, C.R.; Spira, T.P. Biodiversity of fungi in red imported fire ant (Hymenoptera: Formicidae) mounds. Ann. Entomol. Soc. Am. 2002, 95, 487–491. [Google Scholar] [CrossRef] [Green Version]
- Dhar, S.; Jindal, V.; Jariyal, M.; Gupta, V. Molecular characterization of new isolates of the entomopathogenic fungus Beauveria bassiana and their efficacy against the tobacco caterpillar, Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae). Egypt J. Biol. Pest Control. 2019, 29, 8. [Google Scholar] [CrossRef] [Green Version]
- Parks, O.B.; Kothamasu, K.S.; Ziemba, M.J.; Benner, M.; Cristinziano, M.; Kantz, S.; Leger, D.; Li, J.; Patel, D.; Rabuse, W.; et al. Exposure to cuticular bacteria can alter host behavior in a funnel-weaving spider. Curr. Zool. 2018, 64, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Information Retrieval Ltd.: London, UK, 1999; pp. 95–98. [Google Scholar]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The population biology of invasive species. Annu. Rev. Ecol. Evol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef]
- Taylor, M.; Mediannikov, O.; Raoult, D.; Greub, G. Endosymbiotic bacteria associated with nematodes, ticks and amoebae. FEMS Immunol. Med. Mic. 2012, 64, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Clay, K.; Klyachko, O.; Grindle, N.; Civitello, D.; Oleske, D.; Fuqua, C. Microbial communities and interactions in the lone star tick, Amblyomma americanum. Mol. Ecol. 2008, 17, 4371–4381. [Google Scholar] [CrossRef]
- Firrao, G.; Carraro, L.; Gobbi, E.; Locci, R. Molecular characterization of a phytoplasma causing phyllody in clover and other herbaceous hosts in northern Italy. Eur. J. Plant Pathol. 1996, 102, 817–822. [Google Scholar] [CrossRef]
- Dowd, S.E.; Sun, Y.; Secor, P.R.; Rhoads, D.D.; Wolcott, B.M.; James, G.A.; Wolcott, R.D. Survey of bacterial diversity in chronic wounds using pyrosequencing, DGGE, and full ribosome shotgun sequencing. BMC Microbiol. 2008, 8, 43. [Google Scholar] [CrossRef] [Green Version]
- Andreotti, R.; Pérez de León, A.A.; Dowd, S.E.; Guerrero, F.D.; Bendele, K.G.; Scoles, G.A. Assessment of bacterial diversity in the cattle tick Rhipicephalus (Boophilus) microplus through tag-encoded pyrosequencing. BMC Microbiol. 2011, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Enagbonma, B.J.; Babalola, O.O. Unveiling plant-beneficial function as seen in bacteria genes from termite mound soil. J. Soil Sci. Plant Nutr. 2020, 20, 421–430. [Google Scholar] [CrossRef]
- Enagbonma, B.J.; Ajilogba, C.F.; Babalola, O.O. Metagenomic profiling of bacterial diversity and community structure in termite mounds and surrounding soils. Arch. Microbiol. 2020, 202, 2697–2709. [Google Scholar] [CrossRef]
- Bouwma, A.; Ahrens, M.; DeHeer, C.; DeWayne Shoemaker, D. Distribution and prevalence of Wolbachia in introduced populations of the fire ant Solenopsis invicta. Insect Mol. Biol. 2006, 15, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Van Borm, S.; Wenseleers, T.; Billen, J.; Boomsma, J.J. Wolbachia in leafcutter ants: A widespread symbiont that may induce male killing or incompatible matings. J. Evol. Biol. 2001, 14, 805–814. [Google Scholar] [CrossRef]
- Medina, F.; Li, H.; Vinson, S.B.; Coates, C.J. Genetic transformation of midgut bacteria from the red imported fire ant (Solenopsis invicta). Curr. Microbiol. 2009, 58, 478–482. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Isolation | Taxa (Isolate Number) | Location of Isolation | GenBank Accession No. |
|---|---|---|---|
| Ant body | Vagococcus fluvialis D121 | Dongguan | OP811464 |
| Ant body | Vagococcus fluvialis D141 | Dongguan | - |
| Ant body | D142-Unidentified bacterium sp. | Dongguan | - |
| Ant body | D181-Unidentified bacterium sp. | Dongguan | - |
| Ant body | D232-Unidentified bacterium sp. | Dongguan | - |
| Ant body | Myroides odoratimimus H231 | Huizhou | OP811465 |
| Ant body | Lactococcus taiwanensis H182 | Huizhou | - |
| Ant body | Myroides odoratus H191 | Huizhou | - |
| Ant body | H242-Unidentified bacterium sp. | Huizhou | - |
| Ant body | Pseudomonas monteilii G911 | Guangzhou | OP811474 |
| Ant body | Lactococcus garvieae G1012 | Guangzhou | OP811476 |
| Ant body | Myroides odoratimimus G2112 | Guangzhou | OP811479 |
| Ant body | Pseudomonas mosselii G2212 | Guangzhou | OP811480 |
| Ant body | Acinetobacter calcoaceticus G812 | Guangzhou | - |
| Ant body | Myroides marinus G1113 | Guangzhou | - |
| Ant body | G441-Unidentified bacterium sp. | Guangzhou | - |
| Ant body | G512-Unidentified bacterium sp. | Guangzhou | - |
| Ant body | G632-Unidentified bacterium sp. | Guangzhou | - |
| Ant body | G811-Unidentified bacterium sp. | Guangzhou | - |
| Ant body | G912-Unidentified bacterium sp. | Guangzhou | - |
| Ant body | G921-Unidentified bacterium sp. | Guangzhou | - |
| Ant body | G2111-Unidentified bacterium sp. | Guangzhou | - |
| Ant body | Myroides odoratus J4312 | Jiangmen | OP811481 |
| Ant body | Enterobacter asburiae J7312 | Jiangmen | OP811482 |
| Ant body | Enterobacter cloacae J7322 | Jiangmen | OP811483 |
| Ant body | Pantoea dispersa Z241 | Zhuhai | OP811466 |
| Ant body | Acidovorax radicis Z331 | Zhuhai | OP811468 |
| Ant body | Enterococcus saccharolyticus Z412 | Zhuhai | OP811469 |
| Ant body | Myroides odoratus Z421 | Zhuhai | OP811470 |
| Ant body | Myroides odoratus Z442 | Zhuhai | OP811471 |
| Ant body | Providencia rettgeri Z511 | Zhuhai | OP811472 |
| Ant body | Enterobacter cloacae Z721 | Zhuhai | OP811473 |
| Ant body | Enterobacter cloacae Z631 | Zhuhai | - |
| Ant body | Z311-Unidentified bacterium sp. | Zhuhai | - |
| Ant body | Z332-Unidentified bacterium sp. | Zhuhai | - |
| Mound soil | Enterobacter cloacae G922 | Guangzhou | OP811475 |
| Mound soil | Myroides odoratimimus G1612 | Guangzhou | OP811478 |
| Mound soil | Z411-Unidentified bacterium sp. | Zhuhai | - |
| Plant debris | Vagococcus fluvialis G1322 | Guangzhou | OP811477 |
| Plant debris | G722-Unidentified bacterium sp. | Guangzhou | - |
| Plant debris | Lysinibacillus fusiformis J8321 | Jiangmen | OP811484 |
| Plant debris | J1011-Unidentified bacterium sp. | Jiangmen | - |
| Plant debris | Myroides odoratimimus Z312 | Zhuhai | OP811467 |
| Plant debris | Z422-Unidentified bacterium sp. | Zhuhai | - |
| Location | Dongguan | Huizhou | Guangzhou | Zhuhai | Jiangmen | H’ | J’ |
| 11.4 (1) | 9.1 (3) | 38.6 (8) | 29.5 (7) | 11.4 (4) | 1.44 | 0.89 | |
| Substrates | Soil | Plant debris | Ant body | H’ | J’ | ||
| 6.8 (2) | 13.6 (3) | 79.5 (15) | 0.64 | 0.58 |
| Substrates | CC |
|---|---|
| Ant body–Mound soil | 0.13 |
| Ant body–Plant debris | 0.25 |
| Plant debris–Mound soil | 0.12 |
| Location | Substrates | CC |
|---|---|---|
| Dongguan | Ant body–Mound soil | - |
| Ant body–Plant debris | - | |
| Plant debris–Mound soil | - | |
| Guangzhou | Ant body–Mound soil | 0.17 |
| Ant body–Plant debris | 0.33 | |
| Plant debris–Mound soil | 0.5 | |
| Huizhou | Ant body–Mound soil | - |
| Ant body–Plant debris | - | |
| Plant debris–Mound soil | - | |
| Zhuhai | Ant body–Mound soil | 0.14 |
| Ant body–Plant debris | 0.14 | |
| Plant debris–Mound soil | - | |
| Jiangmen | Ant body–Mound soil | - |
| Ant body–Plant debris | - | |
| Plant debris–Mound soil | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bamisile, B.S.; Nie, L.; Siddiqui, J.A.; Ramos Aguila, L.C.; Akutse, K.S.; Jia, C.; Xu, Y. Assessment of Mound Soils Bacterial Community of the Red Imported Fire Ant, Solenopsis invicta across Guangdong Province of China. Sustainability 2023, 15, 1350. https://doi.org/10.3390/su15021350
Bamisile BS, Nie L, Siddiqui JA, Ramos Aguila LC, Akutse KS, Jia C, Xu Y. Assessment of Mound Soils Bacterial Community of the Red Imported Fire Ant, Solenopsis invicta across Guangdong Province of China. Sustainability. 2023; 15(2):1350. https://doi.org/10.3390/su15021350
Chicago/Turabian StyleBamisile, Bamisope Steve, Lei Nie, Junaid Ali Siddiqui, Luis Carlos Ramos Aguila, Komivi Senyo Akutse, Chunsheng Jia, and Yijuan Xu. 2023. "Assessment of Mound Soils Bacterial Community of the Red Imported Fire Ant, Solenopsis invicta across Guangdong Province of China" Sustainability 15, no. 2: 1350. https://doi.org/10.3390/su15021350