
Relationship among Plant Functional Groups, Soil, and Moisture as Basis for Wetland Conservation
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Plant Species Composition
2.3. Soil Sampling
2.4. Water Potential Sampling
2.5. Geospatial Analyses
2.6. Statistical Analysis
3. Results
3.1. Plant Species Composition
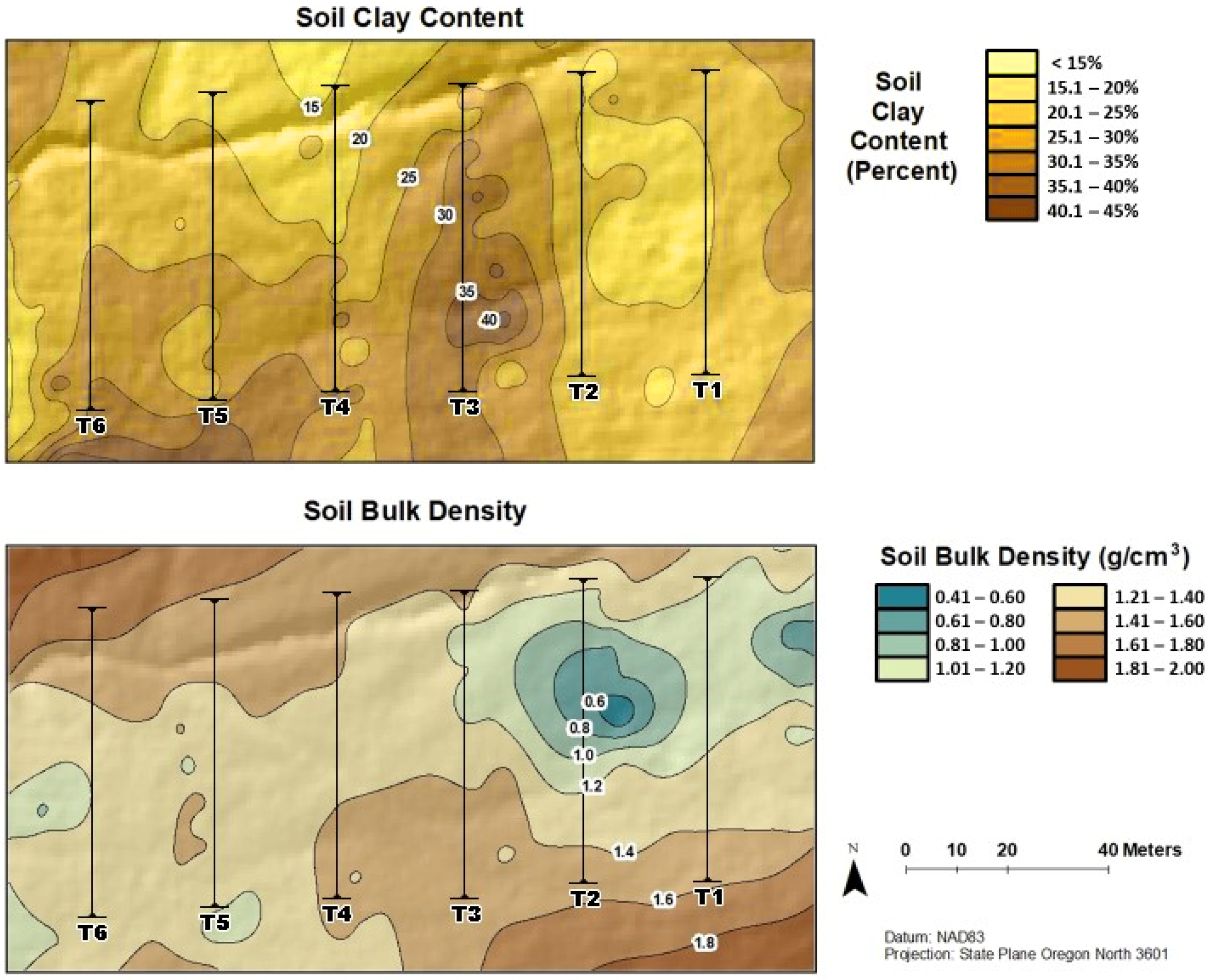
3.2. Soil Characteristics and Moisture Fluctuations
3.3. Plant Water Stress Responses to Drought
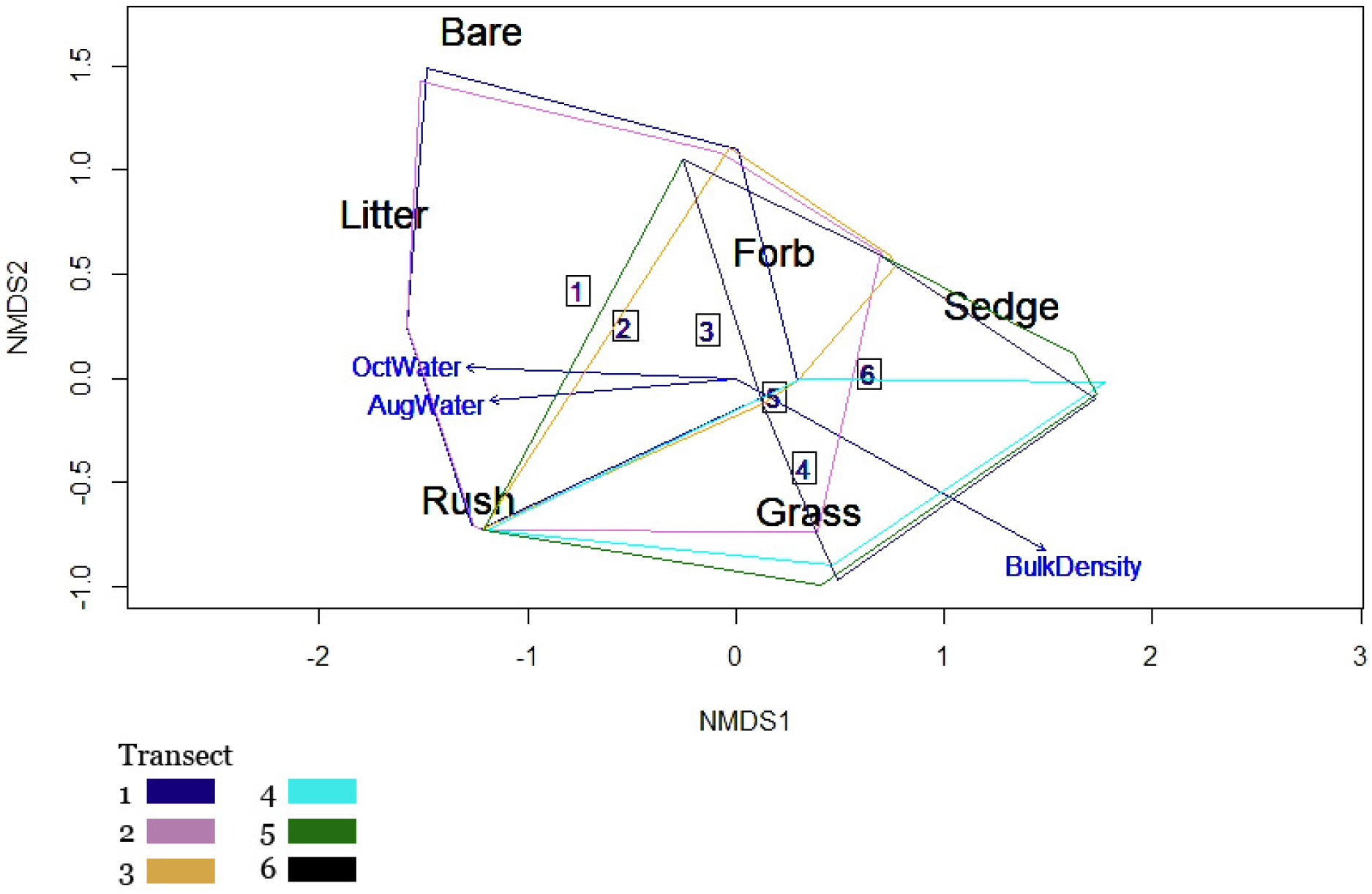
3.4. Functional Group Responses to Soil and Moisture
4. Discussion
4.1. Wetland Plant Species Composition
4.2. Abiotic Factors Affecting Functional Groups
4.3. Wetland Plant Species Responses to Drought
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mitsch, W.; Gosselink, J. Wetlands: Human Use and Science. In Wetlands, 5th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 3–26. [Google Scholar]
- Gilmour, D.M.; Butler, V.L.; O’Connor, J.E.; Davis, E.B.; Culleton, B.J.; Kennett, D.J.; Hodgins, G. Chronology and ecology of late Pleistocene megafauna in the northern Willamette Valley, Oregon. Quat. Res. 2015, 83, 127–136. [Google Scholar] [CrossRef]
- Fickas, K.C.; Cohen, W.B.; Yang, Z. Landsat-based monitoring of annual wetland change in the Willamette Valley of Oregon, USA from 1972 to 2012. Wetl. Ecol. Manag. 2016, 24, 73–92. [Google Scholar] [CrossRef]
- Erwin, K.L. Wetlands and global climate change: The role of wetland restoration in a changing world. Wetl. Ecol. Manag. 2009, 17, 71–84. [Google Scholar] [CrossRef]
- Richardson, C.J. Ecological functions and human values in wetlands: A framework for assessing forestry impacts. Wetlands 1994, 14, 1–9. [Google Scholar] [CrossRef]
- Faulkner, S.; Barrow, W., Jr.; Keeland, B.; Walls, S.; Telesco, D. Effects of conservation practices on wetland ecosystem services in the Mississippi Alluvial Valley. Ecol. Appl. 2011, 21, S31–S48. [Google Scholar] [CrossRef]
- Sueltenfuss, J.P.; Ocheltree, T.W.; Cooper, D.J. Evaluating the realized niche and plant–water relations of wetland species using experimental transplants. Plant Ecol. 2020, 221, 333–345. [Google Scholar] [CrossRef]
- Hao, M.; Zhang, J.; Meng, M.; Chen, H.Y.; Guo, X.; Liu, S.; Ye, L. Impacts of changes in vegetation on saturated hydraulic conductivity of soil in subtropical forests. Sci. Rep. 2019, 9, 8372. [Google Scholar] [CrossRef] [PubMed]
- Niemuth, N.D.; Wangler, B.; Reynolds, R.E. Spatial and temporal variation in wet areas of wetlands in the prairie pothole region of North Dakota and South Dakota. Wetlands 2010, 30, 1053–1064. [Google Scholar] [CrossRef]
- Naumburg, E.; Mata-González, R.; Hunter, R.G.; Mclendon, T.; Martin, D.W. Phreatophytic vegetation and groundwater fluctuations: A review of current research and application of ecosystem response modeling with an emphasis on Great Basin vegetation. Environ. Manag. 2005, 35, 726–740. [Google Scholar] [CrossRef]
- Mata-González, R.; Averett, J.P.; Abdallah, M.A.; Martin, D.W. Variations in groundwater level and microtopography influence desert plant communities in shallow aquifer areas. Environ. Manag. 2022, 69, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Vivian, S.G. Microtopographic heterogeneity and floristic diversity in experimental wetland communities. J. Ecol. 1997, 1, 71–82. [Google Scholar] [CrossRef]
- Reis, V.; Hermoso, V.; Hamilton, S.K.; Ward, D.; Fluet-Chouinard, E.; Lehner, B.; Linke, S. A global assessment of inland wetland conservation status. Bioscience 2017, 67, 523–533. [Google Scholar] [CrossRef]
- Bourdeau, P.F. A test of random versus systematic ecological sampling. Ecology 1953, 34, 499–512. [Google Scholar] [CrossRef]
- McGarvey, R.; Burch, P.; Matthews, J.M. Precision of systematic and random sampling in clustered populations: Habitat patches and aggregating organisms. Ecol. Appl. 2016, 26, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Soil Survey Staff. Natural Resources Conservation Service, United States Department of Agriculture Official Soil Series Descriptions. Available online: http://soils.usda.gov (accessed on 5 February 2020).
- Bonham, C.D. Measurements for Terrestrial Vegetation, 2nd ed.; John Wiley & Sons: Fort Collins, CO, USA, 2013. [Google Scholar]
- Gee, G.W.; Bauder, J.W. Particle size analysis by hydrometer: A simplified method for routine textural analysis and a sensitivity test of measurement parameters. Soil Sci. Soc. Am. J. 1979, 43, 1004–1007. [Google Scholar] [CrossRef]
- Lane, P.W. Generalized linear models in soil science. Eur. J. Soil Sci. 2002, 53, 241–251. [Google Scholar] [CrossRef]
- Stroup, W.W. Rethinking the analysis of non-normal data in plant and soil science. Agron. J. 2015, 107, 811–827. [Google Scholar] [CrossRef]
- Guan, Y.; Wei, J.; Zhang, D.; Zu, M.; Zhang, L. To identify the important soil properties affecting dinoseb adsorption with statistical analysis. Sci. World J. 2013, 6, 713–721. [Google Scholar] [CrossRef]
- Koenker, R.; Machado, J. Goodness of fit and related inference processes for quantile regression. J. Am. Stat. Assoc. 1999, 94, 1296–1310. [Google Scholar] [CrossRef]
- Lichvar, R.W.; Banks, D.L.; Kirchner, W.N.; Melvin, N.C. The National Wetland Plant List: 2016 wetland ratings. Phytoneuron 2016, 30, 1–17. [Google Scholar]
- Van Bodegom, P.M.; Douma, J.C.; Witte, J.P.; Ordoñez, J.C.; Bartholomeus, R.P.; Aert, R. Going beyond limitations of plant functional types when predicting global ecosystem–atmosphere fluxes: Exploring the merits of traits-based approaches. Glob. Ecol. Biogeogr. 2012, 21, 625–636. [Google Scholar] [CrossRef]
- Penna, D.; Tromp-van Meerveld, H.J.; Gobbi, A.; Borga, M.; Dalla Fontana, G. The influence of soil moisture on threshold runoff generation processes in an alpine headwater catchment. Hydrol. Earth Syst. Sci. 2011, 15, 689–702. [Google Scholar] [CrossRef]
- Mohanty, B.P.; Skaggs, T.H. Spatio-temporal evolution and time-stable characteristics of soil moisture within remote sensing footprints with varying soil, slope, and vegetation. Adv. Water Resour. 2001, 24, 1051–1067. [Google Scholar] [CrossRef]
- Hlaváčiková, H.; Novák, V.; Holko, L. On the role of rock fragments and initial soil water content in the potential subsurface runoff formation. J. Hydrol. Hydromech. 2015, 63, 71–81. [Google Scholar] [CrossRef]
- Rector, B.G.; Harizanova, V.; Sforza, R.; Widmer, T.; Wiedenmann, R.N. Prospects for biological control of teasels, Dipsacus spp., a new target in the United States. Biol. Control 2006, 36, 1–14. [Google Scholar] [CrossRef]
- Beaton, L.L.; Dudley, S.A. Tolerance of roadside and old field populations of common teasel (Dipsacus fullonum subsp. sylvestris) to salt and low osmotic potentials during germination. AoB Plants 2013, 5, plt001. [Google Scholar] [CrossRef]
- Daddario, J.F.; Bentivegna, D.J.; Tucat, G.; Fernandez, O.A. Environmental factors affecting seed germination of common teasel (Dipsacus fullonum). Planta Daninha 2017, 35, 823–832. [Google Scholar] [CrossRef]
- Mitchell, M.E.; Lishawa, S.C.; Geddes, P.; Larkin, D.J.; Treering, D.; Tuchman, N.C. Time-dependent impacts of cattail invasion in a Great Lakes coastal wetland complex. Wetlands 2011, 31, 1143–1149. [Google Scholar] [CrossRef]
- Wolf, E.; Cooper, D. Restoration of Geomorphic Structure, Hydrologic Regime, and Vegetation in Upper Halstead Meadow; Project Report to Sequoia National Park; Colorado State University: Three Rivers, CA, USA, 2011. [Google Scholar]
- Sloey, T.M.; Hester, M.W. Interactions between soil physicochemistry and belowground biomass production in a freshwater tidal marsh. Plant Soil 2016, 401, 397–408. [Google Scholar] [CrossRef]
- Czayka, A. Typha Control and Sedge/Grass Meadow Restoration on a Lake Ontario Wetland. Master’s Thesis, SUNY Brockport, Brockport, NY, USA, 2012. [Google Scholar]
- Mitchell, J.W.; Livingston, G.A. Methods of Studying Plant Hormones and Growth-Regulating Substances; Agriculture Handbook 336; U.S. Government Printing Office: Washington, DC, USA, 1968. [Google Scholar]
- Lillak, R.; Viiralt, R.; Linke, A.; Geherman, V. Integrating efficient grassland farming and biodiversity. In Proceedings of the 13th International Occasional Symposium of the European Grassland Federation, Tartu, Estonia, 29–31 August 2005; Estonian Grassland Society: Tartu, Estonia, 2005. [Google Scholar]
- Perry, L.G.; Galatowitsch, S.M.; Rosen, C.J. Competitive control of invasive vegetation: A native wetland sedge suppresses Phalaris arundinacea in carbon-enriched soil. J. Appl. Ecol. 2004, 41, 151–162. [Google Scholar] [CrossRef]
- Sheley, R. Tolerance of meadow foxtail (Alopecurus pratensis) to two sulfonylurea herbicides. Weed Technol. 2007, 21, 470–472. [Google Scholar] [CrossRef]
- Mata-González, R.; McLendon, T.; Martin, D.W.; Trlica, M.J.; Pearce, R.A. Vegetation as affected by groundwater depth and microtopography in a shallow aquifer area of the Great Basin. Ecohydrology 2012, 5, 54–63. [Google Scholar] [CrossRef]
- Evans, T.L.; Mata-González, R.; Martin, D.W.; McLendon, T.; Noller, J.S. Growth, water productivity, and biomass allocation of Great Basin plants as affected by summer watering. Ecohydrology 2013, 6, 713–721. [Google Scholar] [CrossRef]
- Les, D.H. Aquatic Dicotyledons of North America: Ecology, Life History, and Systematics, 1st ed.; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Burdick, D.M.; Roman, C.T. Salt marsh responses to tidal restriction and restoration. In Tidal Marsh Restoration; Island Press: Washington, DC, USA, 2012; pp. 373–382. [Google Scholar]
- Li, S.; Pezeshki, S.R.; Goodwin, S. Effects of soil moisture regimes on photosynthesis and growth in cattail (Typha latifolia). Acta Oecol. 2004, 25, 17–22. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basal Cover (%) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Forbs | Cycle | Status | T1 | T2 | T3 | T4 | T5 | T6 |
| Cirsium arvense | P | FAC | 0 | 0 | 0 | 0 | 0.8 | 0 |
| Daucus carota | B | FACU | 0 | 0 | 0 | 4.9 | 0 | 0 |
| Dipsacus fullonum | B | FAC | 20.6 | 49.6 | 52 | 27.2 | 21.5 | 41.3 |
| Galium aparine | A | FACU | 0.8 | 0 | 0 | 0 | 1.6 | 0.8 |
| Leucanthemum vulgare | P | FACU | 0 | 0 | 0 | 0.8 | 0 | 0 |
| Malva neglecta | A | FACU | 0 | 0 | 0 | 0 | 1.6 | 0 |
| Myosotis laxa | A | OBL | 1.6 | 0 | 0.8 | 1.6 | 0.8 | 0.82 |
| Typha latifolia | P | OBL | 2.5 | 4.9 | 0 | 0 | 0 | 0 |
| Veronica americana | P | OBL | 0 | 0.8 | 0 | 0.8 | 0 | 0 |
| Vicia sativa | A | FACU | 0 | 0 | 0 | 0 | 0 | 0.8 |
| Vicia tetrasperma | A | FACU | 0 | 4.9 | 7.4 | 11.5 | 2.4 | 7.4 |
| Grasses | ||||||||
| Alopecurus pratensis | P | FAC | 1.6 | 1.6 | 7.4 | 30.5 | 34.7 | 14 |
| Holcus lanatus | P | FAC | 1.6 | 0 | 1.6 | 0.8 | 2.4 | 0.8 |
| Phalaris arundinacea | P | FACW | 0 | 0 | 0 | 0 | 0 | 11.5 |
| Poa pratensis | P | FAC | 1.6 | 2.4 | 0.8 | 2.4 | 0 | 1.6 |
| Rushes | ||||||||
| Juncus effusus | P | FACW | 17.3 | 12.4 | 6.6 | 7.4 | 8.2 | 4.9 |
| Juncus patens | P | FACW | 4.9 | 5.8 | 2.4 | 4.9 | 1.6 | 0.8 |
| Sedges | ||||||||
| Carex amplifolia | P | OBL | 0 | 2.5 | 0 | 0 | 0 | 0 |
| Carex densa | P | OBL | 0 | 0 | 0 | 0 | 4.1 | 0.8 |
| Carex feta | P | FACW | 0 | 0 | 0.8 | 0 | 0 | 0 |
| Carex pellita | P | OBL | 0 | 0.8 | 0 | 0 | 0 | 9.1 |
| Carex stipata | P | OBL | 1.6 | 4.1 | 13.2 | 5.8 | 13.2 | 3.3 |
| Schoenoplectus acutus | P | OBL | 0 | 0 | 0 | 0 | 6.6 | 0 |
| Scirpus microcarpus | P | OBL | 27.2 | 4.9 | 0 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aslan, F.; Mata-González, R.; Prado-Tarango, D.E.; Hovland, M.; Stemke, J.; Ochoa, C.G. Relationship among Plant Functional Groups, Soil, and Moisture as Basis for Wetland Conservation. Sustainability 2023, 15, 14377. https://doi.org/10.3390/su151914377
Aslan F, Mata-González R, Prado-Tarango DE, Hovland M, Stemke J, Ochoa CG. Relationship among Plant Functional Groups, Soil, and Moisture as Basis for Wetland Conservation. Sustainability. 2023; 15(19):14377. https://doi.org/10.3390/su151914377
Chicago/Turabian StyleAslan, Fevziye, Ricardo Mata-González, David Eduardo Prado-Tarango, Matthew Hovland, Jenessa Stemke, and Carlos G. Ochoa. 2023. "Relationship among Plant Functional Groups, Soil, and Moisture as Basis for Wetland Conservation" Sustainability 15, no. 19: 14377. https://doi.org/10.3390/su151914377