
Effect of Biochar-Containing Compost on Cucumber Quality and Antibiotic Resistance Genes Abundance in Soil–Cucumber System

Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Materials
2.2. Experimental Setup
2.3. Sampling and Analysis
2.3.1. Soil Sample Collections
2.3.2. Cucumber Sample Collections
2.3.3. Analysis Methods for Each Indicator of Soil and Cucumber
- (1)
- Determination of Soil Physicochemical Properties
- (2)
- Determination of Cucumber Yield and Quality
- (3)
- Determination of Antibiotics in Soil Samples
- (4)
- DNA Extraction from Soil and Cucumber Samples and Q-PCR
- (5)
- Soil Bacterial Community Analysis
2.4. Statistical Analysis
3. Results and Discussion
3.1. Effect of Different Composts on Soil Physicochemical Properties
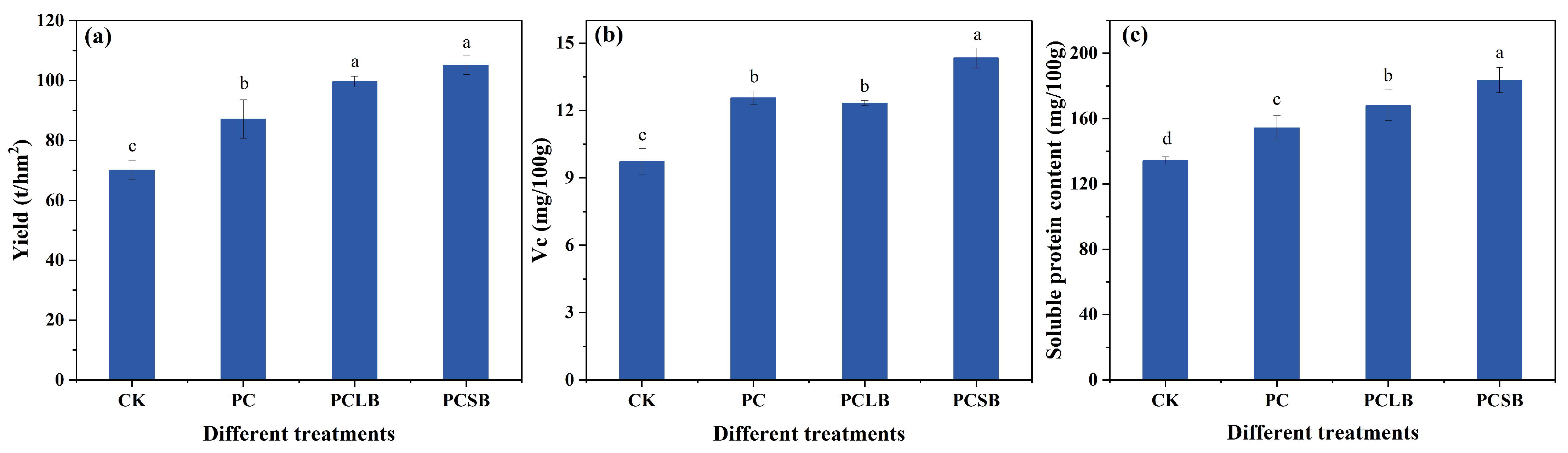
3.2. Effect of Different Composts on Yield and Quality of Cucumber
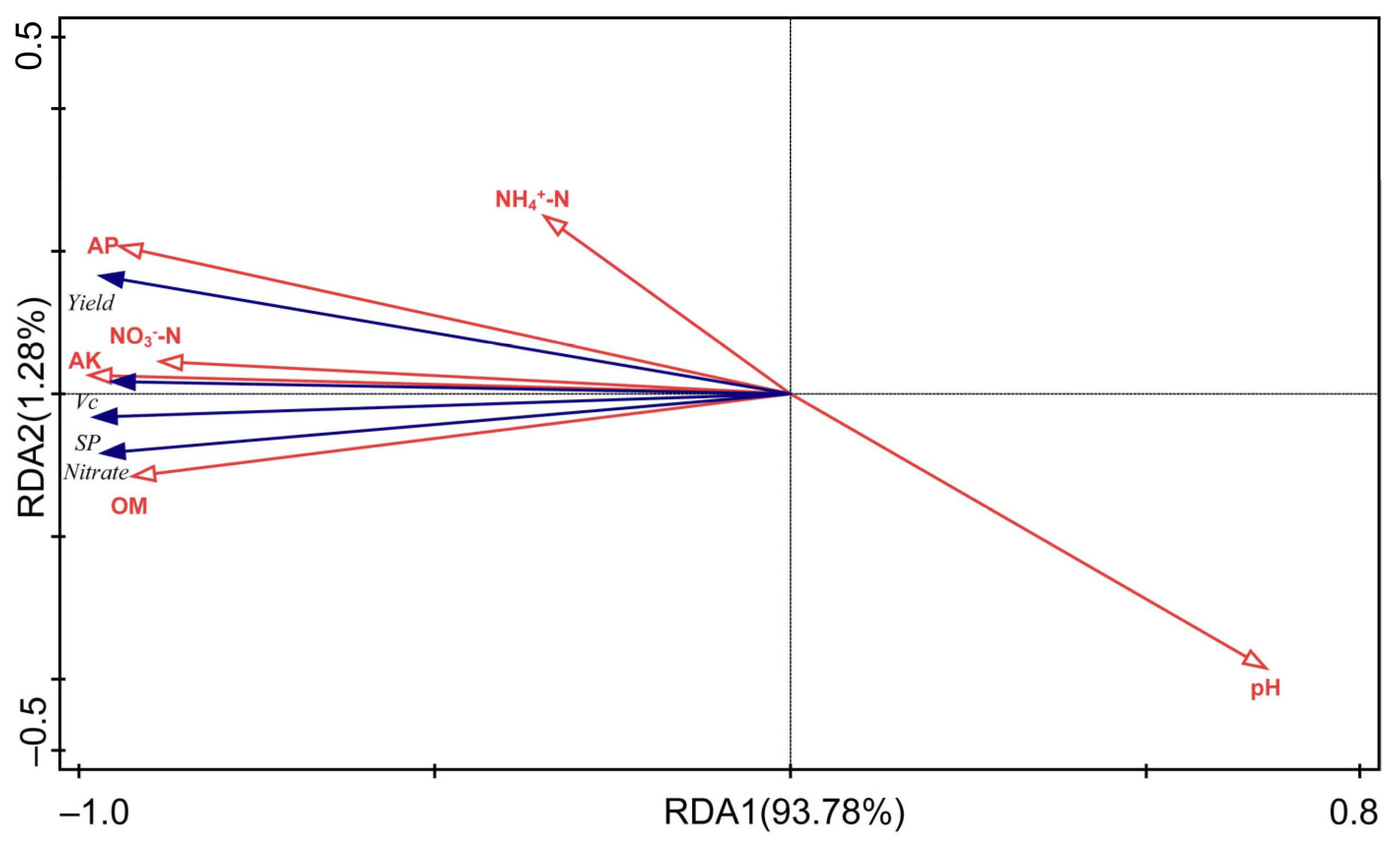
3.3. Relationship between Soil Physicochemical Properties and Cucumber Yield and Quality
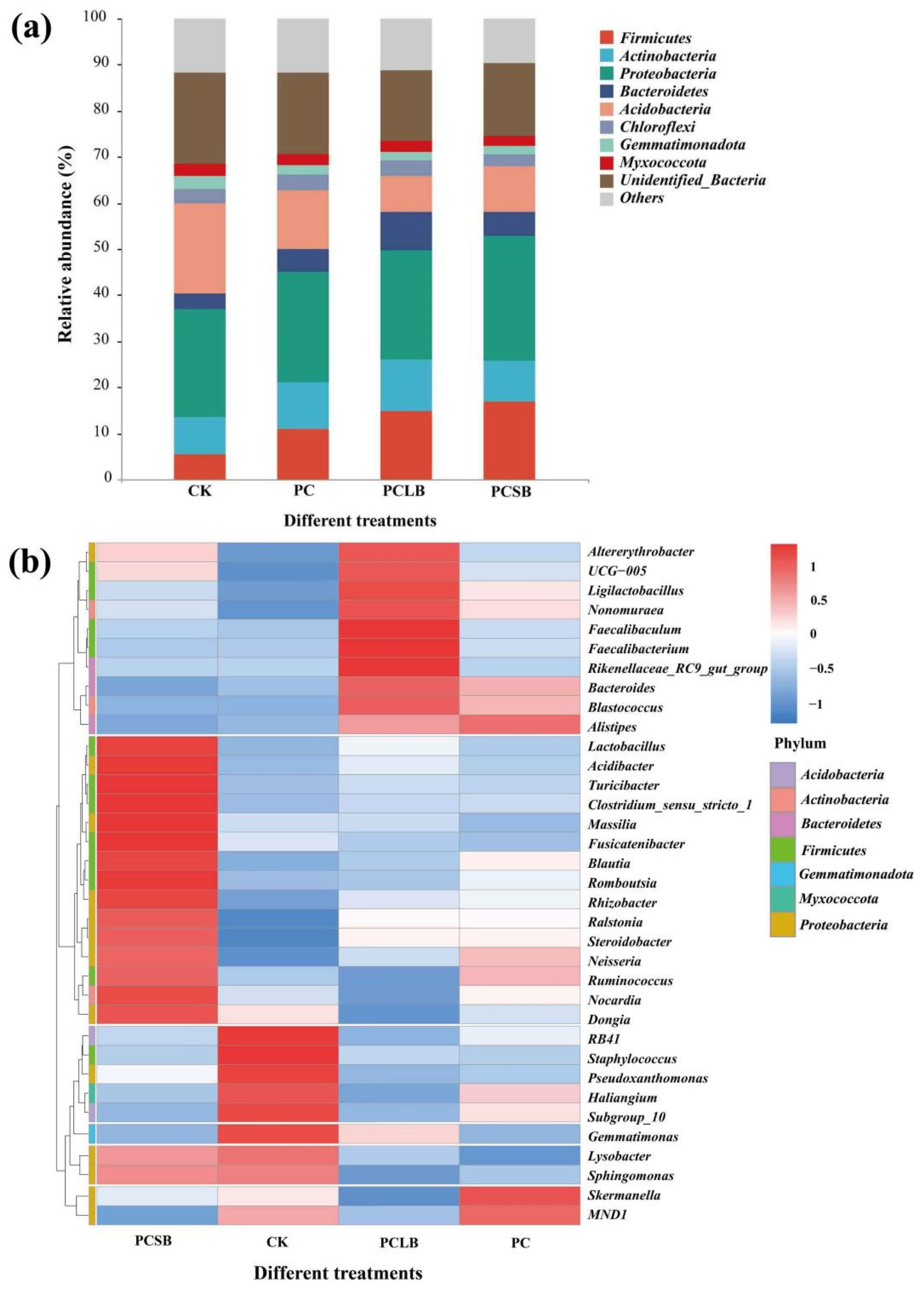
3.4. Effect of Different Composts on Soil Bacterial Community
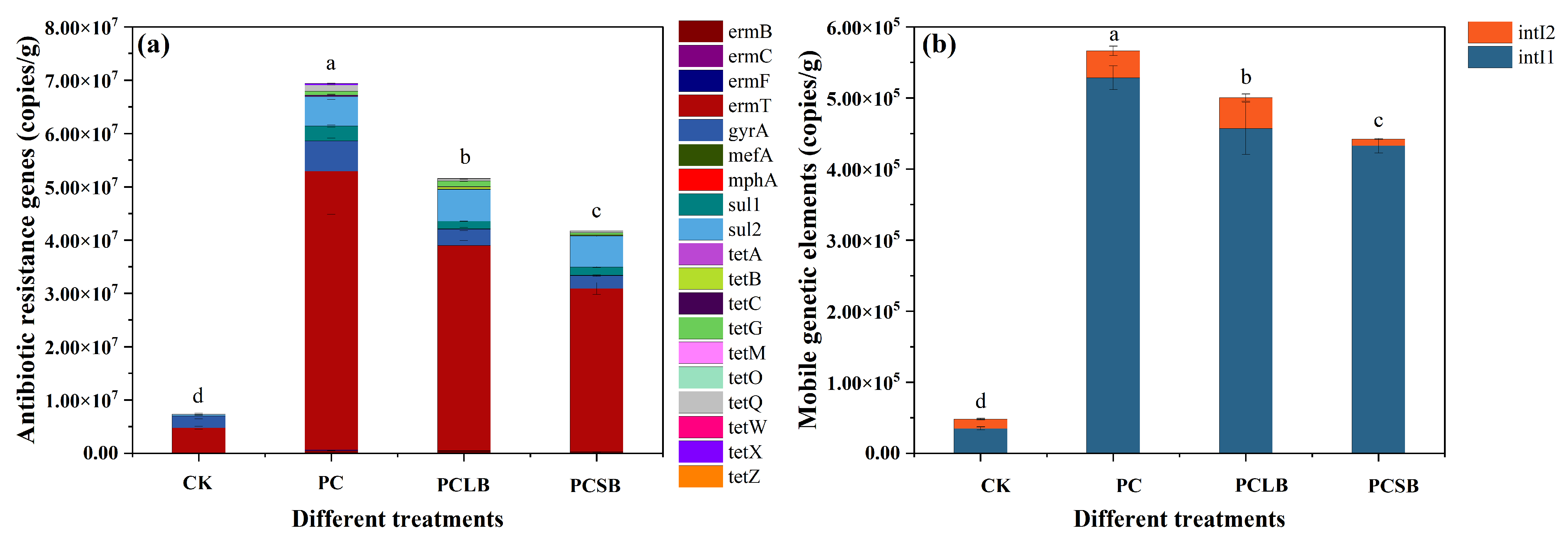
3.5. Effect of Different Composts on Antibiotics and Antibiotic Resistance Genes in Soil
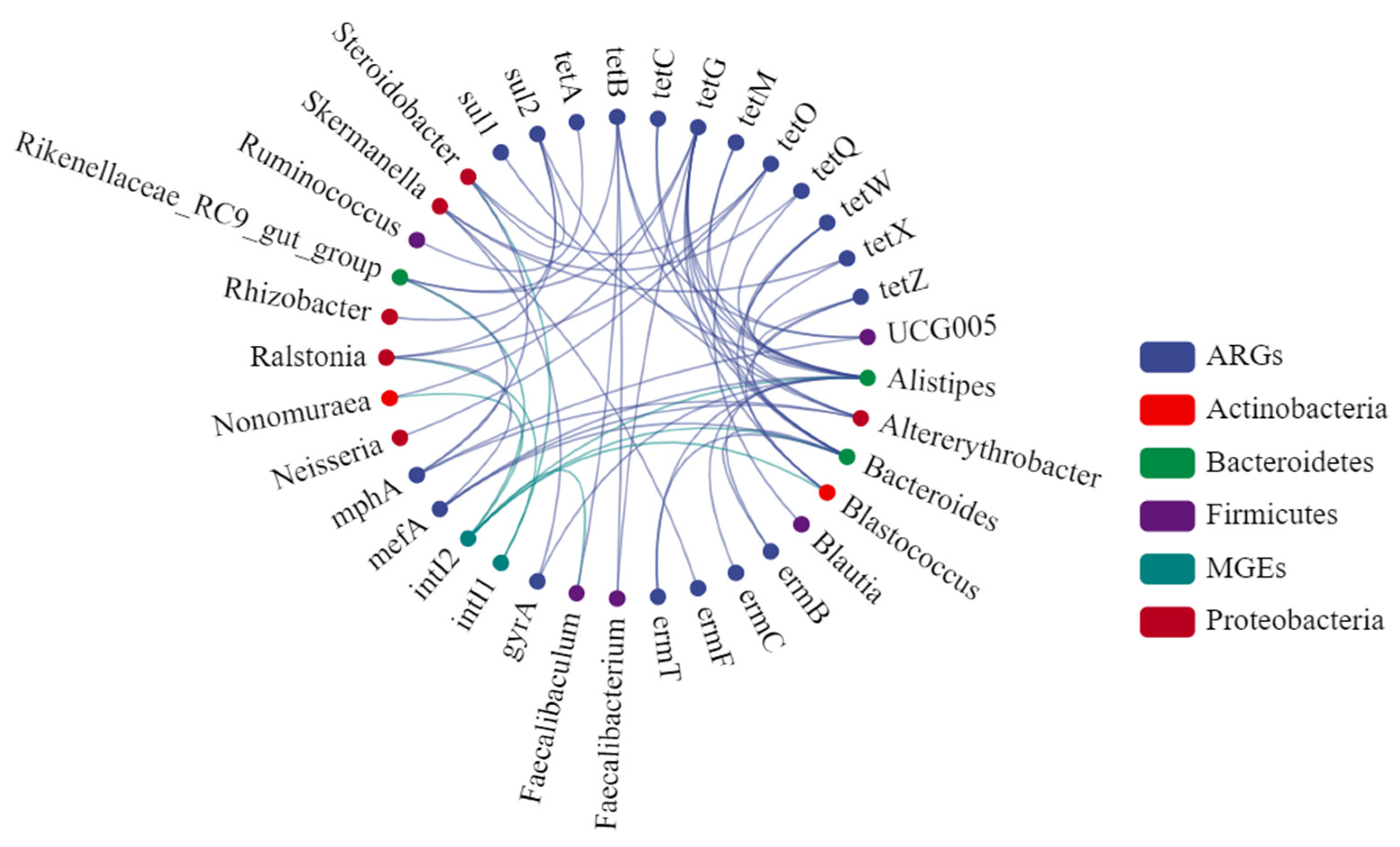
3.6. Relationships between Soil Bacterial Community, Antibiotic Resistance Genes, and Mobile Genetic Elements
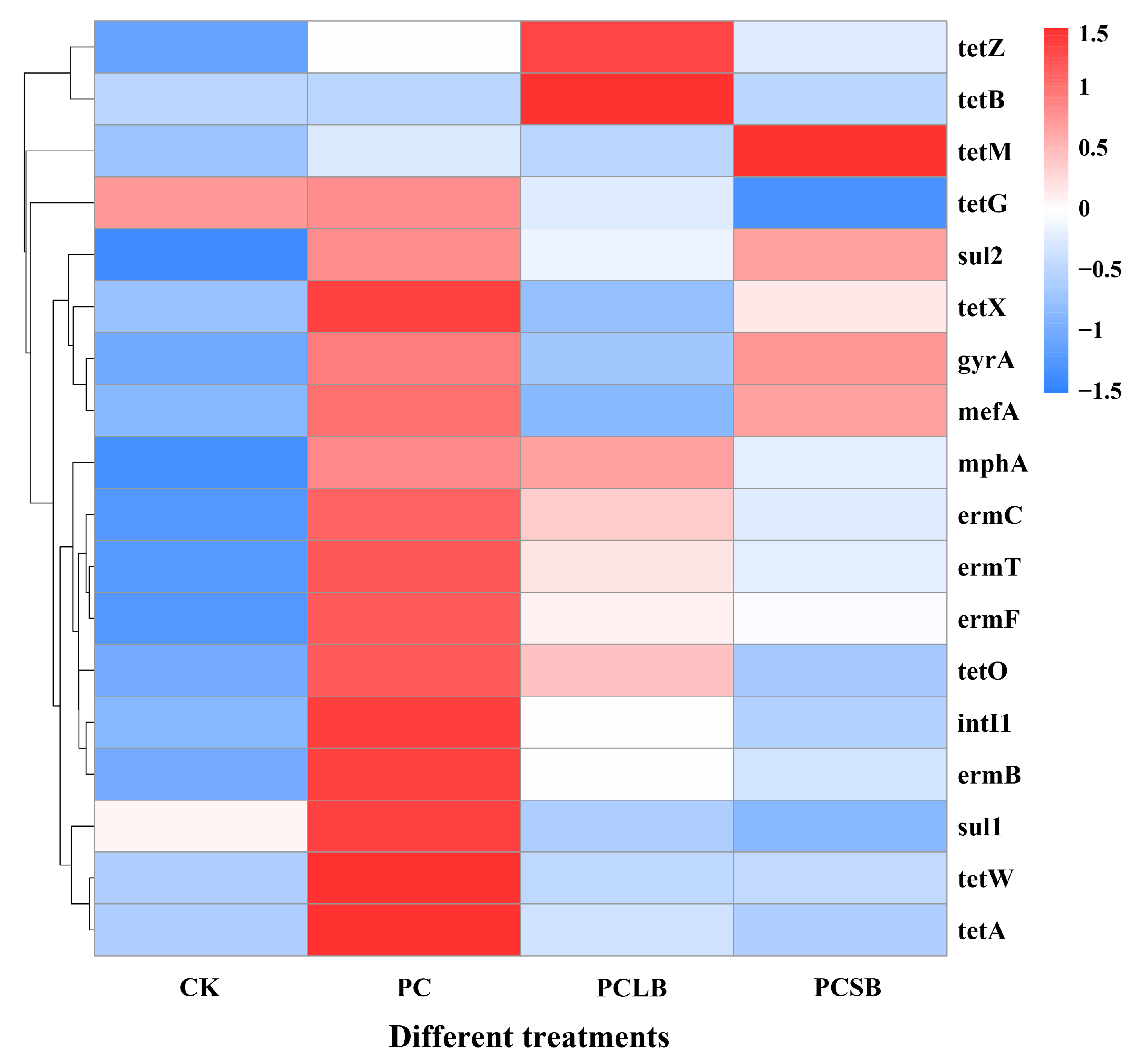
3.7. Effect of Different Composts on Antibiotic Resistance Genes and Mobile Genetic Elements of Cucumber
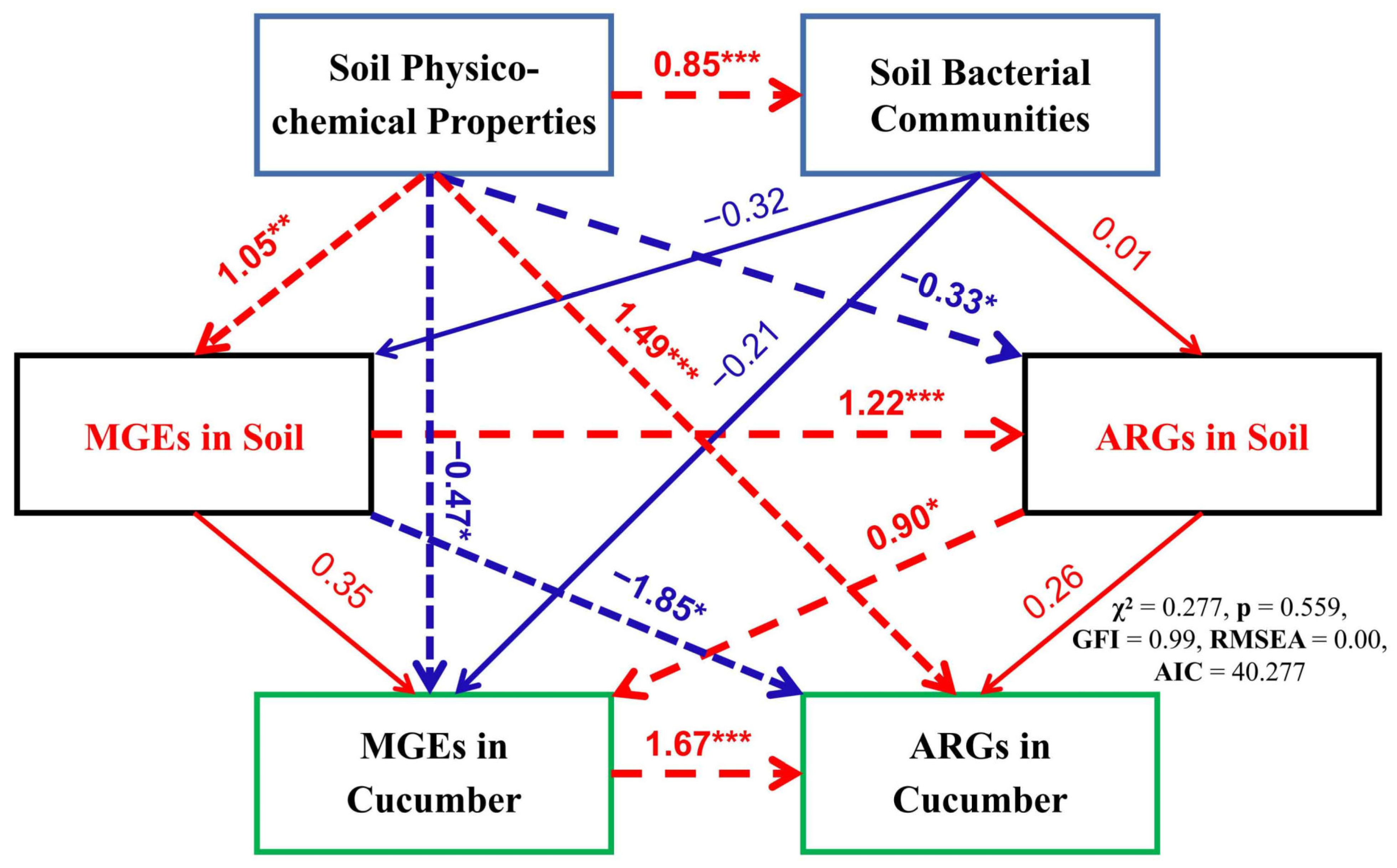
3.8. Mechanisms of Variation in Antibiotic Resistance Genes in Soil–Cucumber Systems
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shao, Y.; Wang, Y.; Yuan, Y.; Xie, Y. A Systematic Review on Antibiotics Misuse in Livestock and Aquaculture and Regulation Implications in China. Sci. Total Environ. 2021, 798, 149205. [Google Scholar] [CrossRef]
- Tian, M.; He, X.; Feng, Y.; Wang, W.; Chen, H.; Gong, M.; Liu, D.; Clarke, J.L.; van Eerde, A. Pollution by Antibiotics and Antimicrobial Resistance in Livestock and Poultry Manure in China, and Countermeasures. Antibiotics 2021, 10, 539. [Google Scholar] [CrossRef]
- Cheng, D.; Feng, Y.; Liu, Y.; Xue, J.; Li, Z. Dynamics of Oxytetracycline, Sulfamerazine, and Ciprofloxacin and Related Antibiotic Resistance Genes During Swine Manure Composting. J. Environ. Manag. 2019, 230, 102–109. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, Y.; Zhou, Y.; Buckley, T.; Wang, H.H. Antibiotic Administration Routes Significantly Influence the Levels of Antibiotic Resistance in Gut Microbiota. Antimicrob. Agents Chemother. 2013, 57, 3659–3666. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Li, H.; Liu, Q.; Ye, C.; Yu, F. Application of Bio-Organic Fertilizer, Not Biochar, in Degraded Red Soil Improves Soil Nutrients and Plant Growth. Rhizosphere 2020, 16, 100264. [Google Scholar] [CrossRef]
- Urra, J.; Alkorta, I.; Lanzén, A.; Mijangos, I.; Garbisu, C. The Application of Fresh and Composted Horse and Chicken Manure Affects Soil Quality, Microbial Composition and Antibiotic Resistance. Appl. Soil Ecol. 2019, 135, 73–84. [Google Scholar] [CrossRef]
- Ablimit, R.; Li, W.; Zhang, J.; Gao, H.; Zhao, Y.; Cheng, M.; Meng, X.; An, L.; Chen, Y. Altering Microbial Community for Improving Soil Properties and Agricultural Sustainability During a 10-Year Maize-Green Manure Intercropping in Northwest China. J. Environ. Manag. 2022, 321, 115859. [Google Scholar] [CrossRef]
- Liu, P.; Jia, S.; He, X.; Zhang, X.; Ye, L. Different Impacts of Manure and Chemical Fertilizers on Bacterial Community Structure and Antibiotic Resistance Genes in Arable Soils. Chemosphere 2017, 188, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Awad, Y.M.; Kim, S.C.; Abd El-Azeem, S.A.M.; Kim, K.H.; Kim, K.R.; Kim, K.; Jeon, C.; Lee, S.S.; Ok, Y.S. Veterinary Antibiotics Contamination in Water, Sediment, and Soil near a Swine Manure Composting Facility. Environ. Earth Sci. 2014, 71, 1433–1440. [Google Scholar] [CrossRef]
- Chen, C.; Pankow, C.A.; Oh, M.; Heath, L.S.; Zhang, L.; Du, P.; Xia, K.; Pruden, A. Effect of Antibiotic Use and Composting on Antibiotic Resistance Gene Abundance and Resistome Risks of Soils Receiving Manure-Derived Amendments. Environ. Int. 2019, 128, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Tien, Y.C.; Li, B.; Zhang, T.; Scott, A.; Murray, R.; Sabourin, L.; Marti, R.; Topp, E. Impact of Dairy Manure Pre-Application Treatment on Manure Composition, Soil Dynamics of Antibiotic Resistance Genes, and Abundance of Antibiotic-Resistance Genes on Vegetables at Harvest. Sci. Total Environ. 2017, 581–582, 32–39. [Google Scholar] [CrossRef]
- Cheng, J.H.; Tang, X.Y.; Cui, J.F. Distinct Aggregate Stratification of Antibiotic Resistome in Farmland Soil with Long-Term Manure Application. Sci. Total Environ. 2022, 833, 155088. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Chen, Z.; Huang, R.; Cui, Y.; Li, Q.; Zhao, Y.; Wang, X.; Mao, D.; Luo, Y.; Ren, H. Antibiotic Resistance Gene-Carrying Plasmid Spreads into the Plant Endophytic Bacteria Using Soil Bacteria as Carriers. Environ. Sci. Technol. 2021, 55, 10462–10470. [Google Scholar] [CrossRef]
- Zhu, B.; Chen, Q.; Chen, S.; Zhu, Y.G. Does Organically Produced Lettuce Harbor Higher Abundance of Antibiotic Resistance Genes Than Conventionally Produced? Environ. Int. 2017, 98, 152–159. [Google Scholar] [CrossRef]
- Forsberg, K.J.; Reyes, A.; Wang, B.; Selleck, E.M.; Sommer, M.O.; Dantas, G. The Shared Antibiotic Resistome of Soil Bacteria and Human Pathogens. Science 2012, 337, 1107–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Zhang, Y.; Zhou, J.; Bao, C.; Chen, M.; He, W.; Shi, X. Effects of Composting Pig Manure at Different Mature Stages on Args in Different Types of Soil-Vegetable Systems. J. Environ. Manag. 2022, 321, 116042. [Google Scholar] [CrossRef]
- Xie, W.Y.; Yang, X.P.; Li, Q.; Wu, L.H.; Shen, Q.R.; Zhao, F.J. Changes in Antibiotic Concentrations and Antibiotic Resistome During Commercial Composting of Animal Manures. Environ. Pollut. 2016, 219, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, M.K.; Liu, T.; Chen, H.; Verma, S.; Duan, Y.; Awasthi, S.K.; Wang, Q.; Ren, X.; Zhao, J.; Zhang, Z. The Behavior of Antibiotic Resistance Genes and Their Associations with Bacterial Community During Poultry Manure Composting. Bioresour. Technol. 2019, 280, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Gou, M.; Hu, H.W.; Zhang, Y.J.; Wang, J.T.; Hayden, H.; Tang, Y.Q.; He, J.Z. Aerobic Composting Reduces Antibiotic Resistance Genes in Cattle Manure and the Resistome Dissemination in Agricultural Soils. Sci. Total Environ. 2018, 612, 1300–1310. [Google Scholar] [CrossRef]
- Sharma, P.; Abrol, V.; Sharma, V.; Chaddha, S.; Srinivasa Rao, C.; Ganie, A.Q.; Ingo Hefft, D.; El-Sheikh, M.A.; Mansoor, S. Effectiveness of Biochar and Compost on Improving Soil Hydro-Physical Properties, Crop Yield and Monetary Returns in Inceptisol Subtropics. Saudi J. Biol. Sci. 2021, 28, 7539–7549. [Google Scholar] [CrossRef]
- Liu, L.; Wang, S.; Guo, X.; Wang, H. Comparison of the Effects of Different Maturity Composts on Soil Nutrient, Plant Growth and Heavy Metal Mobility in the Contaminated Soil. J. Environ. Manag. 2019, 250, 109525. [Google Scholar] [CrossRef]
- Selvam, A.; Zhao, Z.; Wong, J.W.C. Composting of Swine Manure Spiked with Sulfadiazine, Chlortetracycline and Ciprofloxacin. Bioresour. Technol. 2012, 126, 412–417. [Google Scholar] [CrossRef]
- Cao, R.; Wang, J.; Ben, W.; Qiang, Z. The Profile of Antibiotic Resistance Genes in Pig Manure Composting Shaped by Composting Stage: Mesophilic-Thermophilic and Cooling-Maturation Stages. Chemosphere 2020, 250, 126181. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cheng, W.; Li, B.; Xu, Y.; Zheng, X. The Fate of Antibiotic Resistance Genes During Co-Composting of Swine Manure with Cauliflower and Corn Straw. Bioresour. Technol. 2020, 300, 122669. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gu, J.; Wang, X.; Song, Z.; Hu, T.; Xie, J.; Guo, H.; Ding, Q.; Xu, L.; Wei, Y.; et al. The Fate of Antibiotic Resistance Genes and Their Influential Factors in Swine Manure Composting with Sepiolite as Additive. Bioresour. Technol. 2022, 347, 126727. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Liu, F.; Tian, Y.; Zhang, J.; Liu, H.; Duan, J.; Bi, W.; Qin, J.; Xu, S. Effect of Biochar on Antibiotics and Antibiotic Resistance Genes Variations During Co-Composting of Pig Manure and Corn Straw. Front. Bioeng. Biotechnol. 2022, 10, 960476. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Chen, J.; Guan, M.; Xia, H.; Lin, L. Effects of Biochars on the Fate of Antibiotics and Their Resistance Genes During Vermicomposting of Dewatered Sludge. J. Hazard. Mater. 2020, 397, 122767. [Google Scholar] [CrossRef]
- Wang, L.; Sun, X.; Chang, Q.; Tao, Y.; Wang, L.; Dong, J.; Lin, Y.; Zhang, Y. Effect of Di-N-Butyl Phthalate (Dbp) on the Fruit Quality of Cucumber and the Health Risk. Environ. Sci. Pollut. Res. Int. 2016, 23, 24298–24304. [Google Scholar] [CrossRef]
- Zhou, S.; Jiang, Z.; Shen, J.; Yao, Q.; Yang, X.; Li, X.; Awasthi, M.K.; Zhang, Z. Biochar-Amended Compost as a Promising Soil Amendment for Enhancing Plant Productivity: A Meta-Analysis Study. Sci. Total Environ. 2023, 879, 163067. [Google Scholar] [CrossRef]
- Tong, Z.; Liu, F.; Sun, B.; Tian, Y.; Zhang, J.; Duan, J.; Bi, W.; Qin, J.; Xu, S. Effect of Biochars with Different Particle Sizes on Fates of Antibiotics and Antibiotic Resistance Genes During Composting of Swine Manure. Bioresour. Technol. 2023, 370, 128542. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Yuan, J.; Chen, H.; Zhao, X.; Wang, S.; Zhu, Y.; Wang, Y. Animal Manures Promoted Soil Phosphorus Transformation Via Affecting Soil Microbial Community in Paddy Soil. Sci. Total Environ. 2022, 831, 154917. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Wang, B.; Feng, Y.; Fu, H.; Feng, Y.; Xie, H.; Xue, L. Livestock Manure-Derived Hydrochar Improved Rice Paddy Soil Nutrients as a Cleaner Soil Conditioner in Contrast to Raw Material. J. Clean. Prod. 2022, 372, 133798. [Google Scholar] [CrossRef]
- Wang, F.; Wang, X.; Song, N. Biochar and Vermicompost Improve the Soil Properties and the Yield and Quality of Cucumber (Cucumis sativus L.) Grown in Plastic Shed Soil Continuously Cropped for Different Years. Agric. Ecosyst. Environ. 2021, 315, 107425. [Google Scholar] [CrossRef]
- Di, J.; Feng, W.; Zhang, W.; Cai, A.; Xu, M. Soil Organic Carbon Saturation Deficit under Primary Agricultural Managements across Major Croplands in China. Ecosyst. Health Sustain. 2017, 3, 1364047. [Google Scholar] [CrossRef] [Green Version]
- Qi, Z. Instruction in Plant Physiological Experiment; China Agricultural Press: Beijing, China, 2003. [Google Scholar]
- Wang, G.; Li, G.; Chang, J.; Kong, Y.; Jiang, T.; Wang, J.; Yuan, J. Enrichment of Antibiotic Resistance Genes after Sheep Manure Aerobic Heap Composting. Bioresour. Technol. 2021, 323, 124620. [Google Scholar] [CrossRef]
- Yin, Y.; Gu, J.; Wang, X.; Song, W.; Zhang, K.; Sun, W.; Zhang, X.; Zhang, Y.; Li, H. Effects of Copper Addition on Copper Resistance, Antibiotic Resistance Genes, and Intl1 During Swine Manure Composting. Front. Microbiol. 2017, 8, 344. [Google Scholar] [CrossRef] [Green Version]
- Gryta, A.; Frąc, M.; Oszust, K. Genetic and Metabolic Diversity of Soil Microbiome in Response to Exogenous Organic Matter Amendments. Agronomy 2020, 10, 546. [Google Scholar] [CrossRef] [Green Version]
- Oldfield, T.L.; Sikirica, N.; Mondini, C.; López, G.; Kuikman, P.J.; Holden, N.M. Biochar, Compost and Biochar-Compost Blend as Options to Recover Nutrients and Sequester Carbon. J. Environ. Manag. 2018, 218, 465–476. [Google Scholar] [CrossRef]
- Agegnehu, G.; Bass, A.M.; Nelson, P.N.; Muirhead, B.; Wright, G.; Bird, M.I. Biochar and Biochar-Compost as Soil Amendments: Effects on Peanut Yield, Soil Properties and Greenhouse Gas Emissions in Tropical North Queensland, Australia. Agric. Ecosyst. Environ. 2015, 213, 72–85. [Google Scholar] [CrossRef]
- Siedt, M.; Schäffer, A.; Smith, K.E.C.; Nabel, M.; Roß-Nickoll, M.; van Dongen, J.T. Comparing Straw, Compost, and Biochar Regarding Their Suitability as Agricultural Soil Amendments to Affect Soil Structure, Nutrient Leaching, Microbial Communities, and the Fate of Pesticides. Sci. Total Environ. 2021, 751, 141607. [Google Scholar] [CrossRef]
- Guo, Z.; Wan, S.; Hua, K.; Yin, Y.; Chu, H.; Wang, D.; Guo, X. Fertilization Regime Has a Greater Effect on Soil Microbial Community Structure Than Crop Rotation and Growth Stage in an Agroecosystem. Appl. Soil Ecol. 2020, 149, 103510. [Google Scholar] [CrossRef]
- Ho, T.T.K.; Tra, V.T.; Le, T.H.; Nguyen, N.K.Q.; Tran, C.S.; Nguyen, P.T.; Vo, T.D.H.; Thai, V.N.; Bui, X.T. Compost to Improve Sustainable Soil Cultivation and Crop Productivity. Case Stud. Chem. Environ. Eng. 2022, 6, 100211. [Google Scholar] [CrossRef]
- Wang, X.; Cammeraat, E.L.H.; Cerli, C.; Kalbitz, K. Soil Aggregation and the Stabilization of Organic Carbon as Affected by Erosion and Deposition. Soil Biol. Biochem. 2014, 72, 55–65. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and Fulvic Acids as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Agegnehu, G.; Bass, A.M.; Nelson, P.N.; Bird, M.I. Benefits of Biochar, Compost and Biochar–Compost for Soil Quality, Maize Yield and Greenhouse Gas Emissions in a Tropical Agricultural Soil. Sci. Total Environ. 2016, 543, 295–306. [Google Scholar] [CrossRef]
- Guo, X.; Liu, H.; Zhang, J. The Role of Biochar in Organic Waste Composting and Soil Improvement: A Review. Waste Manag. 2020, 102, 884–899. [Google Scholar] [CrossRef]
- Hua, K.; Wang, D.; Guo, Z. Soil Organic Carbon Contents as a Result of Various Organic Amendments to a Vertisol. Nutr. Cycl. Agroecosyst. 2017, 108, 135–148. [Google Scholar] [CrossRef]
- Zhang, Y.; Cong, J.; Lu, H.; Li, G.; Qu, Y.; Su, X.; Zhou, J.; Li, D. Community Structure and Elevational Diversity Patterns of Soil Acidobacteria. J. Environ. Sci. 2014, 26, 1717–1724. [Google Scholar] [CrossRef]
- Shen, Y.; Stedtfeld, R.D.; Guo, X.; Bhalsod, G.D.; Jeon, S.; Tiedje, J.M.; Li, H.; Zhang, W. Pharmaceutical Exposure Changed Antibiotic Resistance Genes and Bacterial Communities in Soil-Surface- and Overhead-Irrigated Greenhouse Lettuce. Environ. Int. 2019, 131, 105031. [Google Scholar] [CrossRef]
- Heuer, H.; Schmitt, H.; Smalla, K. Antibiotic Resistance Gene Spread Due to Manure Application on Agricultural Fields. Curr. Opin. Microbiol. 2011, 14, 236–243. [Google Scholar] [CrossRef]
- Tashiro, Y.; Kanda, K.; Asakura, Y.; Kii, T.; Cheng, H.; Poudel, P.; Okugawa, Y.; Tashiro, K.; Sakai, K. A Unique Autothermal Thermophilic Aerobic Digestion Process Showing a Dynamic Transition of Physicochemical and Bacterial Characteristics from the Mesophilic to the Thermophilic Phase. Appl. Environ. Microbiol. 2018, 84, e02537-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Wang, Z.; Xu, T.; Feng, Z.; Liu, J.; Luo, L.; He, Y.; Xiao, Y.; Peng, H.; Zhang, Y.; et al. The Fate of Antibiotic Resistance Genes and Their Influential Factors During Excess Sludge Composting in a Full-Scale Plant. Bioresour. Technol. 2021, 342, 126049. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Niu, X.; Zhang, N.; Li, T. Effect of Biochar-Immobilized Sphingomonas Sp. Pj2 on Bioremediation of Pahs and Bacterial Community Composition in Saline Soil. Chemosphere 2021, 279, 130427. [Google Scholar] [CrossRef] [PubMed]
- Barrios, R.E.; Khuntia, H.K.; Bartelt-Hunt, S.L.; Gilley, J.E.; Schmidt, A.M.; Snow, D.D.; Li, X. Fate and Transport of Antibiotics and Antibiotic Resistance Genes in Runoff and Soil as Affected by the Timing of Swine Manure Slurry Application. Sci. Total Environ. 2020, 712, 136505. [Google Scholar] [CrossRef]
- D’Alessio, M.; Durso, L.M.; Miller, D.N.; Woodbury, B.; Ray, C.; Snow, D.D. Environmental Fate and Microbial Effects of Monensin, Lincomycin, and Sulfamethazine Residues in Soil. Environ. Pollut. 2019, 246, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, A.; Saez, J.M.; Davila Costa, J.S.; Colin, V.L.; Fuentes, M.S.; Cuozzo, S.A.; Benimeli, C.S.; Polti, M.A.; Amoroso, M.J. Actinobacteria: Current Research and Perspectives for Bioremediation of Pesticides and Heavy Metals. Chemosphere 2017, 166, 41–62. [Google Scholar] [CrossRef]
- Liu, W.; Ling, N.; Guo, J.; Ruan, Y.; Wang, M.; Shen, Q.; Guo, S. Dynamics of the Antibiotic Resistome in Agricultural Soils Amended with Different Sources of Animal Manures over Three Consecutive Years. J. Hazard. Mater. 2021, 401, 123399. [Google Scholar] [CrossRef]
- Wang, F.; Han, W.; Chen, S.; Dong, W.; Qiao, M.; Hu, C.; Liu, B. Fifteen-Year Application of Manure and Chemical Fertilizers Differently Impacts Soil Args and Microbial Community Structure. Front. Microbiol. 2020, 11, 62. [Google Scholar] [CrossRef]
- Cerqueira, F.; Christou, A.; Fatta-Kassinos, D.; Vila-Costa, M.; Bayona, J.M.; Piña, B. Effects of Prescription Antibiotics on Soil- and Root-Associated Microbiomes and Resistomes in an Agricultural Context. J. Hazard. Mater. 2020, 400, 123208. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Wang, Y.; Wang, F.; Xia, S.; Zhao, J. Struvite-Supported Biochar Composite Effectively Lowers Cu Bio-Availability and the Abundance of Antibiotic-Resistance Genes in Soil. Sci. Total Environ. 2020, 724, 138294. [Google Scholar] [CrossRef]
- Wang, F.H.; Qiao, M.; Chen, Z.; Su, J.Q.; Zhu, Y.G. Antibiotic Resistance Genes in Manure-Amended Soil and Vegetables at Harvest. J. Hazard. Mater. 2015, 299, 215–221. [Google Scholar] [CrossRef]
- Martínez, J.L.; Coque, T.M.; Baquero, F. What Is a Resistance Gene? Ranking Risk in Resistomes. Nat. Rev. Microbiol. 2015, 13, 116–123. [Google Scholar] [CrossRef]
- Lian, F.; Yu, W.; Zhou, Q.; Gu, S.; Wang, Z.; Xing, B. Size Matters: Nano-Biochar Triggers Decomposition and Transformation Inhibition of Antibiotic Resistance Genes in Aqueous Environments. Environ. Sci. Technol. 2020, 54, 8821–8829. [Google Scholar] [CrossRef]
- Ye, M.; Sun, M.; Feng, Y.; Wan, J.; Xie, S.; Tian, D.; Zhao, Y.; Wu, J.; Hu, F.; Li, H.; et al. Effect of Biochar Amendment on the Control of Soil Sulfonamides, Antibiotic-Resistant Bacteria, and Gene Enrichment in Lettuce Tissues. J. Hazard. Mater. 2016, 309, 219–227. [Google Scholar] [CrossRef]
- Xu, S.; Duan, Y.; Zou, S.; Liu, H.; Luo, L.; Wong, J.W.C. Evaluations of Biochar Amendment on Anaerobic Co-Digestion of Pig Manure and Sewage Sludge: Waste-to-Methane Conversion, Microbial Community, and Antibiotic Resistance Genes. Bioresour. Technol. 2022, 346, 126400. [Google Scholar] [CrossRef] [PubMed]
- Phares, C.A.; Akaba, S. Co-Application of Compost or Inorganic Npk Fertilizer with Biochar Influences Soil Quality, Grain Yield and Net Income of Rice. J. Integr. Agric. 2022, 21, 3600–3610. [Google Scholar] [CrossRef]
- Teodoro, M.; Trakal, L.; Gallagher, B.N.; Šimek, P.; Soudek, P.; Pohořelý, M.; Beesley, L.; Jačka, L.; Kovář, M.; Seyedsadr, S.; et al. Application of Co-Composted Biochar Significantly Improved Plant-Growth Relevant Physical/Chemical Properties of a Metal Contaminated Soil. Chemosphere 2020, 242, 125255. [Google Scholar] [CrossRef]
- Han, X.M.; Hu, H.W.; Chen, Q.L.; Yang, L.Y.; Li, H.L.; Zhu, Y.G.; Li, X.Z.; Ma, Y.B. Antibiotic Resistance Genes and Associated Bacterial Communities in Agricultural Soils Amended with Different Sources of Animal Manures. Soil Biol. Biochem. 2018, 126, 91–102. [Google Scholar] [CrossRef]
- Jechalke, S.; Kopmann, C.; Rosendahl, I.; Groeneweg, J.; Weichelt, V.; Krögerrecklenfort, E.; Brandes, N.; Nordwig, M.; Ding, G.C.; Siemens, J.; et al. Increased Abundance and Transferability of Resistance Genes after Field Application of Manure from Sulfadiazine-Treated Pigs. Appl. Environ. Microbiol. 2013, 79, 1704–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physicochemical Properties | Compost Samples | ||
|---|---|---|---|
| PC | PCLB | PCSB | |
| pH | 7.51 ± 0.04 | 7.63 ± 0.06 | 7.68 ± 0.04 |
| EC mS/cm | 4.19 ± 0.05 | 3.43 ± 0.13 | 3.88 ± 0.02 |
| TN g/kg | 22.14 ± 0.85 | 20.30 ± 0.84 | 21.24 ± 0.53 |
| TP g/kg | 21.35 ± 0.82 | 18.82 ± 0.21 | 19.46 ± 0.40 |
| TK g/kg | 7.15 ± 0.13 | 7.92 ± 0.30 | 10.19 ± 0.22 |
| AP mg/kg | 159.46 ± 0.83 | 165.06 ± 2.76 | 174.11 ± 0.93 |
| AK mg/kg | 13,192.45 ± 124.70 | 13,903.77 ± 64.46 | 14,654.30 ± 59.41 |
| TOC g/kg | 288.41 ± 0.91 | 296.47 ± 4.12 | 318.58 ± 2.16 |
| Physicochemical Properties | Treatment | |||
|---|---|---|---|---|
| CK | PC | PCLB | PCSB | |
| Organic matter (g/kg) | 15.34 ± 0.15 c | 16.32 ± 0.11 b | 17.13 ± 0.41 b | 18.71 ± 0.92 a |
| AP (mg/kg) | 61.34 ± 1.96 c | 96.71 ± 3.76 b | 102.58 ± 1.24 b | 113.86 ± 7.34 a |
| AK (mg/kg) | 544.86 ± 7.71 d | 609.26 ± 6.16 c | 638.52 ± 2.96 b | 671.99 ± 25.95 a |
| NH4+-N (mg/kg) | 1.43 ± 0.10 d | 6.15 ± 1.04 a | 4.56 ± 0.97 b | 2.96 ± 0.08 c |
| NO3−-N (mg/kg) | 35.90 ± 0.60 c | 42.97 ± 0.74 b | 47.08 ± 0.89 b | 56.15 ± 5.47 a |
| pH | 8.72 ± 0.02 a | 8.70 ± 0.05 a | 8.69 ± 0.03 a | 8.67 ± 0.01 a |
| Large aggregates (%) | 57.24 ± 0.87 c | 61.34 ± 1.45 b | 62.53 ± 0.52 b | 67.24 ± 0.90 a |
| Small aggregates (%) | 13.48 ± 0.25 ab | 14.07 ± 1.51 a | 11.97 ± 0.61 b | 12.73 ± 0.57 ab |
| Micro-aggregates (%) | 29.28 ± 1.00 a | 24.59 ± 2.96 b | 25.51 ± 1.00 b | 20.03 ± 1.47 c |
| Bulk density (g/cm3) | 1.03 ± 0.01 a | 1.01 ± 0.02 a | 0.98 ± 0.01 b | 0.97 ± 0.01 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, Z.; Liu, F.; Rajagopalan, U.M.; Sun, B.; Tian, Y.; Zuo, Q.; Zhang, J.; Duan, J.; Bi, W.; Qin, J.; et al. Effect of Biochar-Containing Compost on Cucumber Quality and Antibiotic Resistance Genes Abundance in Soil–Cucumber System. Sustainability 2023, 15, 9563. https://doi.org/10.3390/su15129563
Tong Z, Liu F, Rajagopalan UM, Sun B, Tian Y, Zuo Q, Zhang J, Duan J, Bi W, Qin J, et al. Effect of Biochar-Containing Compost on Cucumber Quality and Antibiotic Resistance Genes Abundance in Soil–Cucumber System. Sustainability. 2023; 15(12):9563. https://doi.org/10.3390/su15129563
Chicago/Turabian StyleTong, Zhenye, Fenwu Liu, Uma Maheswari Rajagopalan, Bo Sun, Yu Tian, Qihui Zuo, Jingzhi Zhang, Jiaze Duan, Wenlong Bi, Junmei Qin, and et al. 2023. "Effect of Biochar-Containing Compost on Cucumber Quality and Antibiotic Resistance Genes Abundance in Soil–Cucumber System" Sustainability 15, no. 12: 9563. https://doi.org/10.3390/su15129563