
Diet Fermentation Leads to Microbial Adaptation in Black Soldier Fly (Hermetia illucens; Linnaeus, 1758) Larvae Reared on Palm Oil Side Streams

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Screening and Substrate Preparation
2.2. Insect Rearing
2.3. Developmental Parameters
2.4. 16S rRNA Gene and ITS Amplicon Sequencing
2.5. Statistics
3. Results
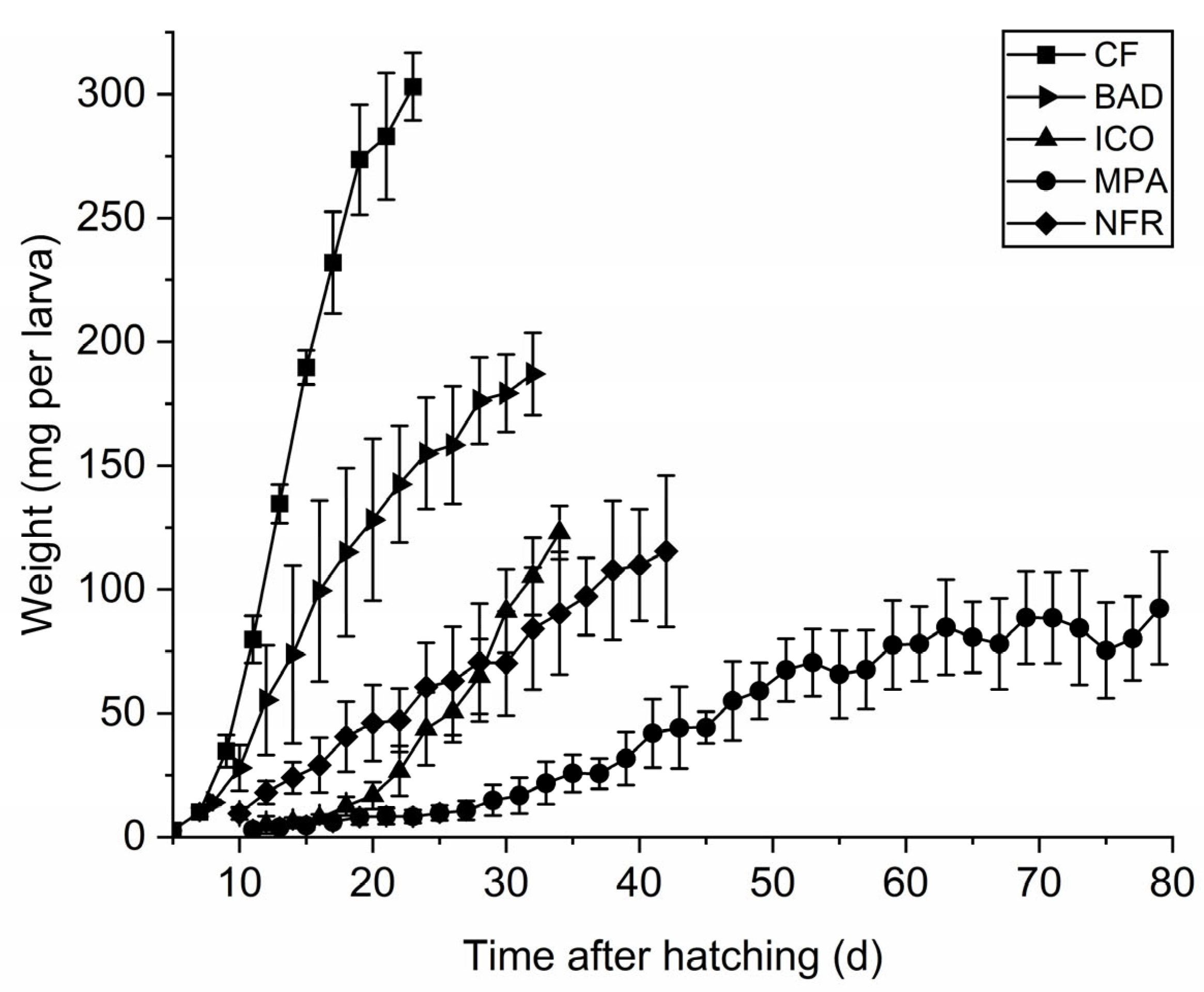
3.1. Fungal Screening and BSF Growth Performance
3.2. Life-History Traits
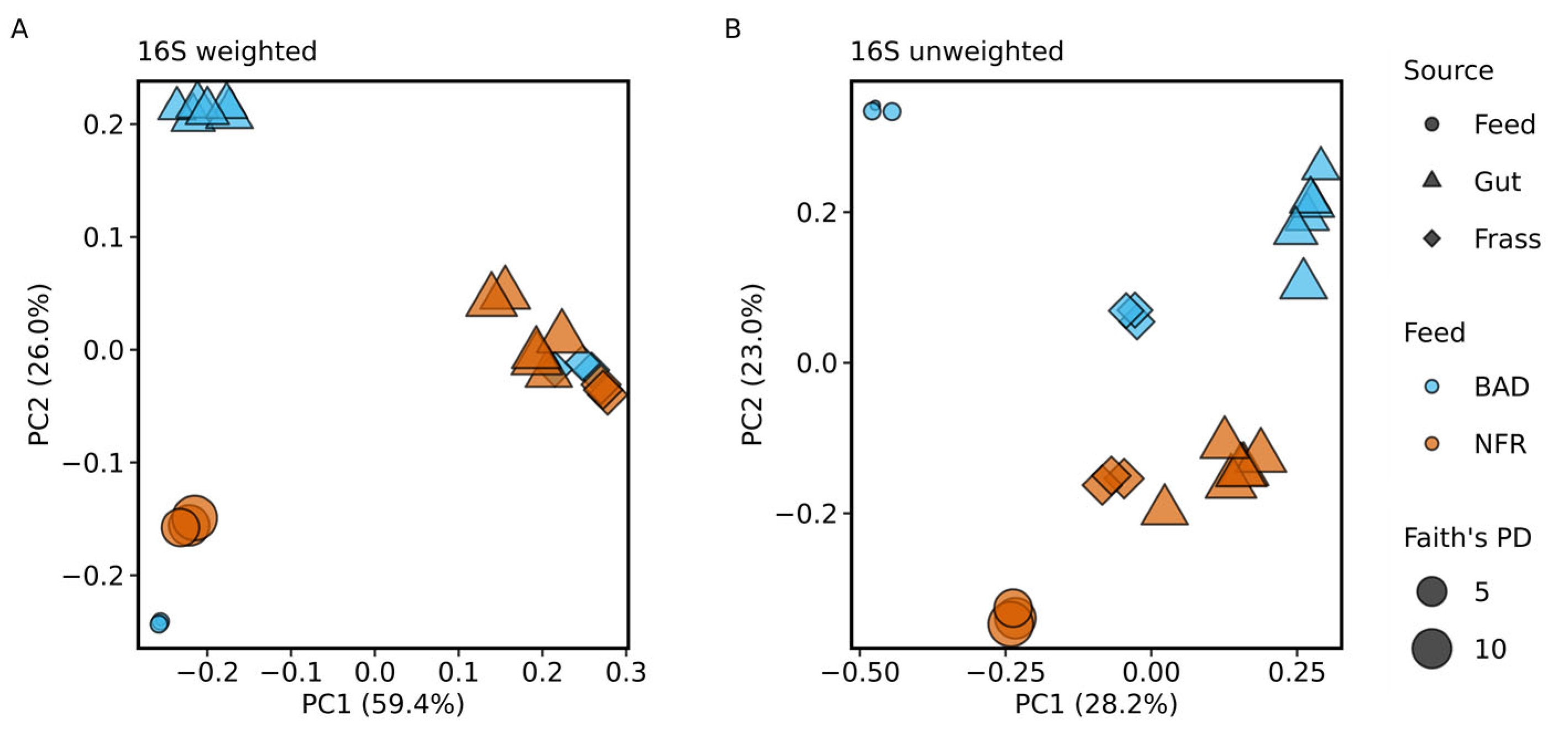
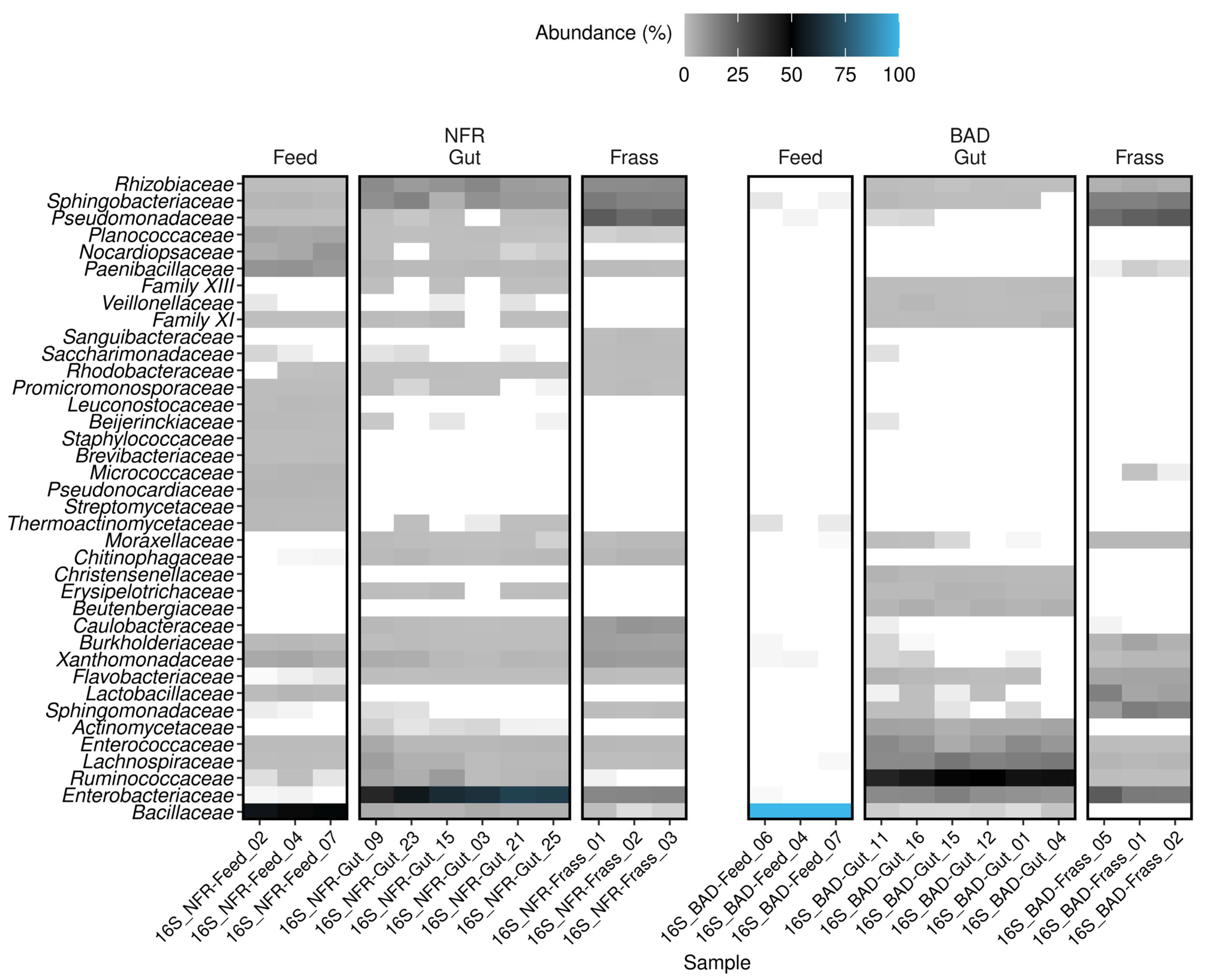
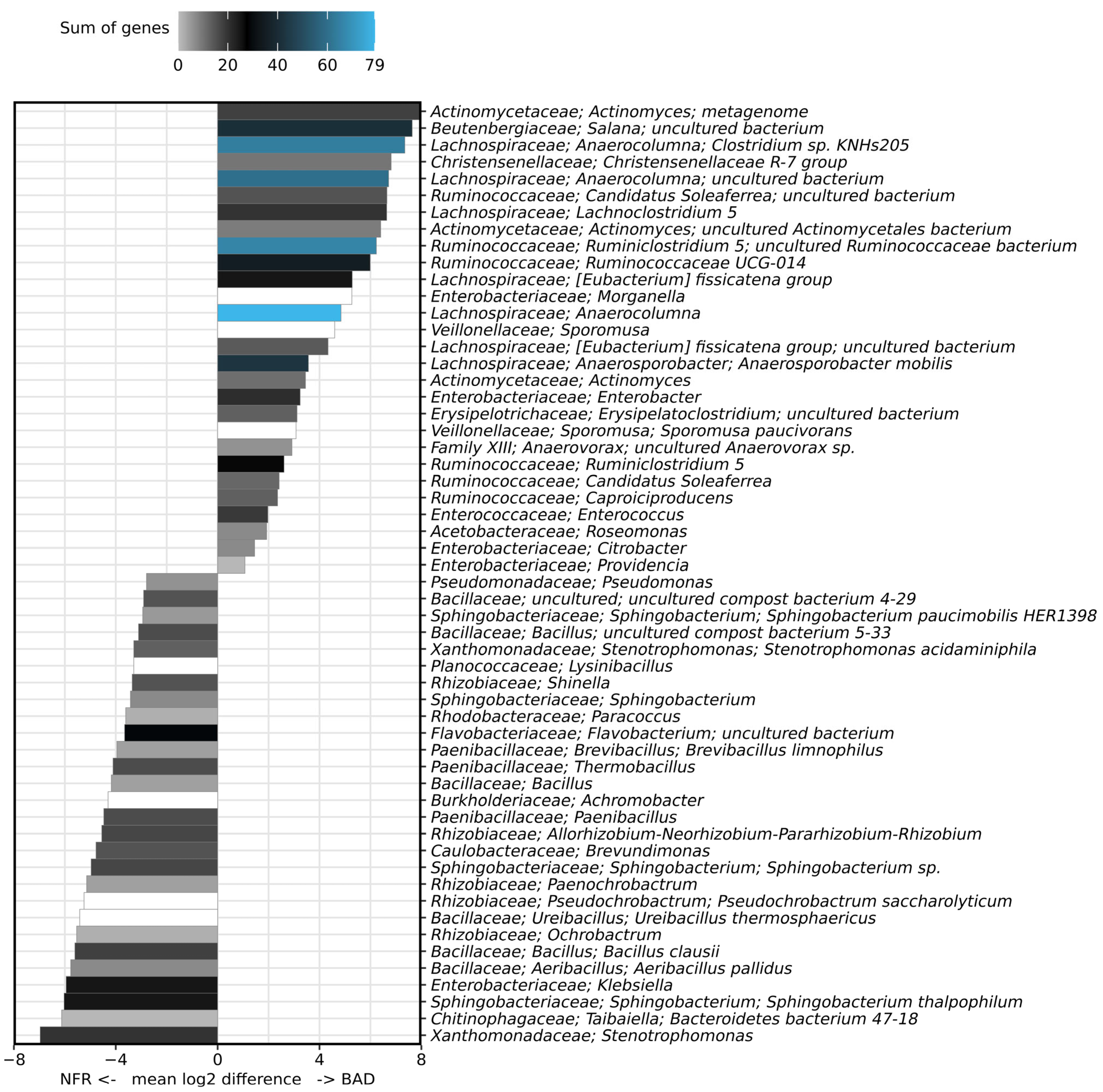
3.3. Taxonomic Composition of the Bacterial Gut Microbiome
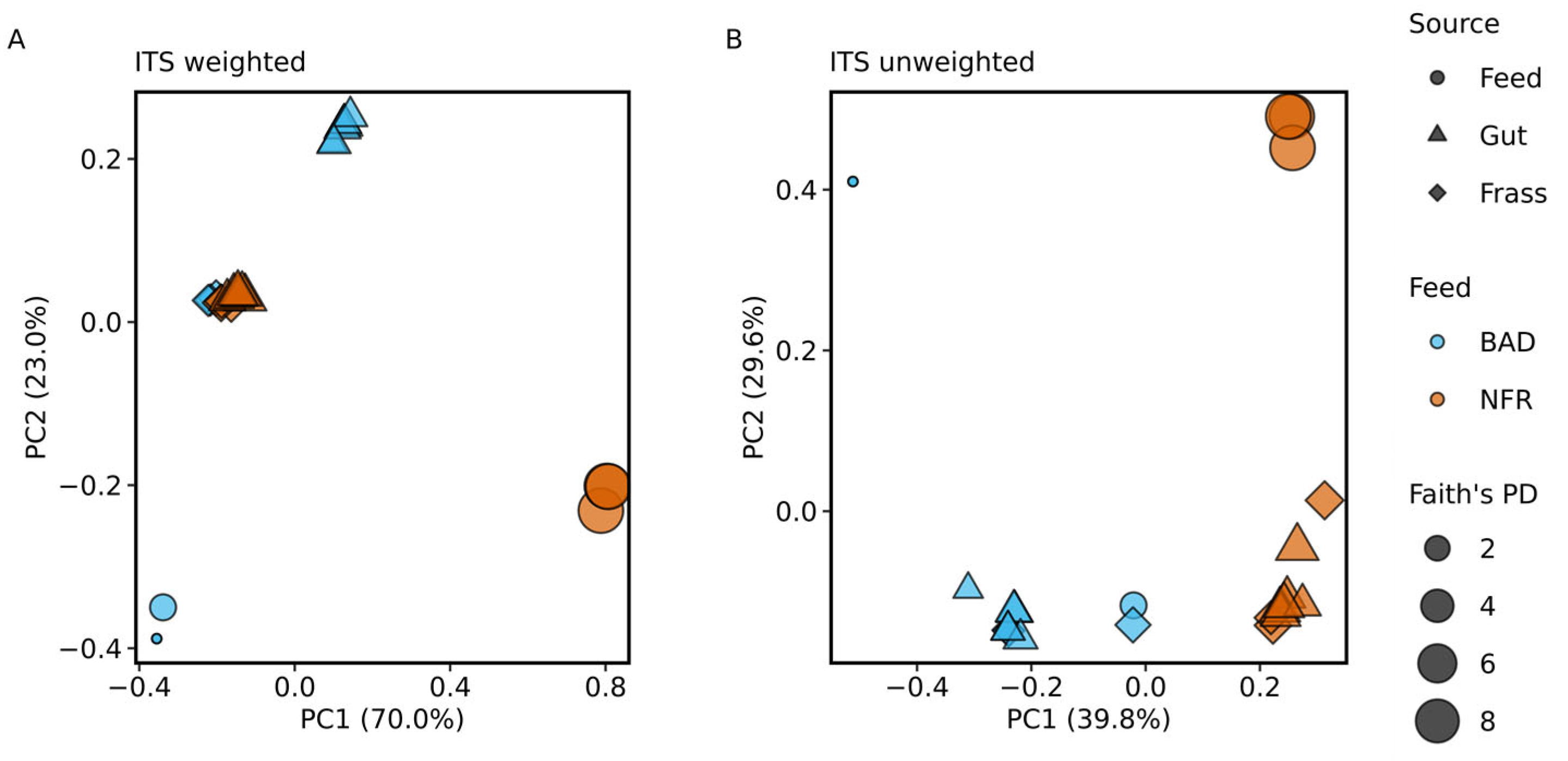
3.4. Taxonomic Composition of the Fungal Gut Microbiome
4. Discussion
4.1. Rationale for the Pretreatment of Palm Oil Side Streams
4.2. Pretreatment by Fermentation Influences BSF Life-History Traits
4.3. Analysis of the Bacterial Communities Associated with BSF
4.4. Analysis of the Fungal Communities Associated with BSF
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lalander, C.; Diener, S.; Zurbrügg, C.; Vinneras, B. Effects of feedstock on larval development and process efficiency in waste treatment with black soldier fly (Hermetia illucens). J. Clean. Prod. 2018, 208, 211–219. [Google Scholar] [CrossRef]
- Miranda, C.D.; Cammack, J.A.; Tomberlin, J.K. Life-history traits of the black soldier fly, Hermetia illucens (L.) (Diptera: Stratiomyidae), reared on three manure types. Animals 2019, 9, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, M.; Cassar, C.M.; Zurbrügg, C.; Kreuzer, M.; Boulos, S.; Diener, S.; Mathys, A. Biowaste treatment with black soldier fly larvae: Increasing performance through the formulation of biowastes based on protein and carbohydrates. Waste Manag. 2019, 102, 319–329. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; Van Huis, A.; Van Loon, J. Nutrient utilisation by black soldier flies fed with chicken, pig, or cow manure. J. Insects Food Feed 2015, 1, 131–139. [Google Scholar] [CrossRef]
- Müller, A.; Wolf, D.; Gutzeit, H.O. The black soldier fly, Hermetia illucens—A promising source for sustainable production of proteins, lipids and bioactive substances. Z. Naturforsch. 2017, 72, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.; Park, S.; Choi, J.; Jeong, G.; Lee, S.-B.; Choi, Y.; Lee, S.-J. The Intestinal Bacterial Community in the Food Waste-Reducing Larvae of Hermetia illucens. Curr. Microbiol. 2011, 62, 1390–1399. [Google Scholar] [CrossRef]
- Tegtmeier, D.; Hurka, S.; Klüber, P.; Brinkrolf, K.; Heise, P.; Vilcinskas, A. Cottonseed press cake as a potential diet for industrially farmed black soldier fly larvae triggers adaptations of their bacterial and fungal gut microbiota. Front. Microbiol. 2021, 12, 634503. [Google Scholar] [CrossRef] [PubMed]
- Varotto Boccazzi, I.; Ottoboni, M.; Martin, E.; Comandatore, F.; Vallone, L.; Spranghers, T.; Eeckhout, M.; Mereghetti, V.; Pinotti, L.; Epis, S. A survey of the mycobiota associated with larvae of the black soldier fly (Hermetia illucens) reared for feed production. PLoS ONE 2017, 12, e0182533. [Google Scholar] [CrossRef] [Green Version]
- Bruno, D.; Bonelli, M.; De Filippis, F.; Di Lelio, I.; Tettamanti, G.; Casartelli, M.; Ercolini, D.; Caccia, S. The Intestinal Microbiota of Hermetia illucens Larvae Is Affected by Diet and Shows a Diverse Composition in the Different Midgut Regions. Appl. Environ. Microbiol. 2019, 85, e01864-18. [Google Scholar] [CrossRef] [Green Version]
- Wynants, E.; Frooninckx, L.; Crauwels, S.; Verreth, C.; De Smet, J.; Sandrock, C.; Wohlfahrt, J.; Van Schelt, J.; Depraetere, S.; Lievens, B.; et al. Assessing the Microbiota of Black Soldier Fly Larvae (Hermetia illucens) Reared on Organic Waste Streams on Four Different Locations at Laboratory and Large Scale. Microb. Ecol. 2018, 77, 913–930. [Google Scholar] [CrossRef]
- Vogel, H.; Müller, A.; Heckel, D.; Gutzeit, H.; Vilcinskas, A. Nutritional immunology: Diversification and diet-dependent expression of antimicrobial peptides in the black soldier fly Hermetia illucens. Dev. Comp. Immunol. 2018, 78, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Klammsteiner, T.; Walter, A.; Bogataj, T.; Heussler, C.D.; Stres, B.; Steiner, F.M.; Schlick-Steiner, B.C.; Arthofer, W.; Insam, H. The Core Gut Microbiome of Black Soldier Fly (Hermetia illucens) Larvae Raised on Low-Bioburden Diets. Front. Microbiol. 2020, 11, 993. [Google Scholar] [CrossRef] [PubMed]
- Klüber, P.; Müller, S.; Schmidt, J.; Zorn, H.; Rühl, M. Isolation of Bacterial and Fungal Microbiota Associated with Hermetia illucens Larvae Reveals Novel Insights into Entomopathogenicity. Microorganisms 2022, 10, 319. [Google Scholar] [CrossRef]
- Yang, F.; Tomberlin, J.K.; Jordan, H.R. Starvation Alters Gut Microbiome in Black Soldier Fly (Diptera: Stratiomyidae) Larvae. Front. Microbiol. 2021, 12, 601253. [Google Scholar] [CrossRef]
- Yu, G.; Cheng, P.; Yang, Z.; Chen, Y.; Li, Y.; Tomberlin, J.K. Inoculating Poultry Manure With Companion Bacteria Influences Growth and Development of Black Soldier Fly (Diptera: Stratiomyidae) Larvae. Environ. Entomol. 2011, 40, 30–35. [Google Scholar] [CrossRef]
- Martin, M.M. The Evolution of Insect-Fungus Associations: From Contact to Stable Symbiosis. Am. Zool. 1992, 32, 593–605. [Google Scholar] [CrossRef]
- Correa, Y.; Cabanillas, B.; Jullian, V.; Álvarez, D.; Castillo, D.; Dufloer, C.; Bustamante, B.; Roncal, E.; Neyra, E.; Sheen, P.; et al. Identification and characterization of compounds from Chrysosporium multifidum, a fungus with moderate antimicrobial activity isolated from Hermetia illucens gut microbiota. PLoS ONE 2019, 14, e0218837. [Google Scholar] [CrossRef]
- Schrögel, P.; Wätjen, W. Insects for Food and Feed-Safety Aspects Related to Mycotoxins and Metals. Foods 2019, 8, 288. [Google Scholar] [CrossRef] [Green Version]
- Basiron, Y. Palm oil production through sustainable plantations. Eur. J. Lipid Sci. Technol. 2007, 109, 289–295. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTAT Database. 2018. Available online: http://faostat3.fao.org/home/E (accessed on 15 November 2021).
- Santosa, S.J. Palm Oil Boom in Indonesia: From Plantation to Downstream Products and Biodiesel. CLEAN-Soil Air Water 2008, 36, 453–465. [Google Scholar] [CrossRef]
- Rachmawati, R.; Buchori, D.; Hidayat, P.; Hem, S.; Fahmi, M.R. Perkembangan dan Kandungan Nutrisi Larva Hermetia illucens (Linnaeus) (Diptera: Stratiomyidae) pada Bungkil Kelapa Sawit. Jurnal Entomologi Indonesia 2010, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Andlar, M.; Rezić, T.; Marđetko, N.; Kracher, D.; Ludwig, R.; Šantek, B. Lignocellulose degradation: An overview of fungi and fungal enzymes involved in lignocellulose degradation. Eng. Life Sci. 2018, 18, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, P.; Anand, A.; Bajpai, P.K. Bleaching with lignin-oxidizing enzymes. Biotechnol. Annu. Rev. 2006, 12, 349–378. [Google Scholar] [PubMed]
- Diener, S.; Zurbrügg, C.; Tockner, K. Conversion of organic material by black soldier fly larvae: Establishing optimal feeding rates. Waste Manag. Res. J. Sustain. Circ. Econ. 2009, 27, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Lei, Y.; Rehman, K.U.; Yu, Z.; Zhang, J.; Li, W.; Li, Q.; Tomberlin, J.K.; Zheng, L. Dynamic Effects of Initial pH of Substrate on Biological Growth and Metamorphosis of Black Soldier Fly (Diptera: Stratiomyidae). Environ. Entomol. 2018, 47, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Klüber, P.; Bakonyi, D.; Zorn, H.; Rühl, M. Does light color temperature influence aspects of oviposition by the black soldier fly (Diptera: Stratiomyidae)? J. Econ. Entomol. 2020, 113, 2549–2552. [Google Scholar] [CrossRef]
- Barros, L.M.; Gutjahr, A.L.N.; Keppler, R.L.F.; Martins, R.T. Morphological description of the immature stages of Hermetia illucens (Linnaeus, 1758) (Diptera: Stratiomyidae). Microsc. Res. Tech. 2019, 82, 178–189. [Google Scholar] [CrossRef]
- Rozkosný, R. A Biosystematic Study of the European Stratiomyidae (Diptera): Clitellariinae, Hermediinae, Pachygasterinae and Bibliography; Springer: Dordrecht, The Netherlands, 1983. [Google Scholar]
- Sundberg, C.; Al-Soud, W.A.; Larsson, M.; Alm, E.; Yekta, S.S.; Svensson, B.H.; Sørensen, S.J.; Karlsson, A. 454 pyrosequencing analyses of bacterial and archaeal richness in 21 full-scale biogas digesters. FEMS Microbiol. Ecol. 2013, 85, 612–626. [Google Scholar] [CrossRef] [Green Version]
- Ihrmark, K.; Bödeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandström-Durling, M.; Clemmensen, K.E.; et al. New primers to amplify the fungal ITS2 region—Evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Caporaso, J.G. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Abarenkov, K.; Zirk, A.; Piirmann, T.; Pöhönen, R.; Ivanov, F.; Nilsson, R.H.; Kõljalg, U. UNITE QIIME Release for Fungi; UNITE Community: London, UK, 2013. [Google Scholar]
- Werner, J.J.; Koren, O.; Hugenholtz, P.; DeSantis, T.Z.; Walters, W.A.; Caporaso, J.G.; Angenent, L.; Knight, R.; Ley, R. Impact of training sets on classification of high-throughput bacterial 16s rRNA gene surveys. ISME J. 2011, 6, 94–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Amir, A.; Morton, J.T.; Heller, R.; Arias-Castro, E.; Knight, R. Discrete False-Discovery Rate Improves Identification of Differentially Abundant Microbes. mSystems 2017, 2, e00092-17. [Google Scholar] [CrossRef] [Green Version]
- Markowitz, V.M.; Mavrommatis, K.; Ivanova, N.; Chen, I.-M.A.; Chu, K.; Kyrpides, N. IMG ER: A system for microbial genome annotation expert review and curation. Bioinformatics 2009, 25, 2271–2278. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Holmes, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA); Balakrishnan, N., Colton, T., Everitt, B., Piegorsch, W., Ruggeri, F., Teugels, J.L., Eds.; Wiley StatsRef: Statistics Reference Online.
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Lee, S.; Kang, M.; Bae, J.-H.; Sohn, J.-H.; Sung, B.H. Bacterial Valorization of Lignin: Strains, Enzymes, Conversion Pathways, Biosensors, and Perspectives. Front. Bioeng. Biotechnol. 2019, 7, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, Y.; Sudo, K.; Shimizu, K. Enzymatic hydrolysis of woods. II. Effect of grinding and alkali treatment on hydrolysis of woods by Trichoderma viride cellulase. J. Jpn. Wood Res. Soc. 1977, 23, 562–570. [Google Scholar]
- Brune, A. Symbiotic digestion of lignocellulose in termite guts. Nat. Rev. Genet. 2014, 12, 168–180. [Google Scholar] [CrossRef]
- Tartar, A.; Wheeler, M.M.; Zhou, X.; Coy, M.R.; Boucias, D.G.; Scharf, M.E. Parallel metatranscriptome analyses of host and symbiont gene expression in the gut of the termite Reticulitermes flavipes. Biotechnol. Biofuels 2009, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Callegari, M.; Jucker, C.; Fusi, M.; Leonardi, M.G.; Daffonchio, D.; Borin, S.; Savoldelli, S.; Crotti, E. Hydrolytic profile of the culturable gut bacterial community associated with Hermetia illucens. Front. Microbiol. 2020, 11, 1965. [Google Scholar] [CrossRef]
- Martínez, A.T.; Speranza, M.; Ruiz-Dueñas, F.J.; Ferreira, P.; Camarero, S.; Guillén, F.; Martinez, M.J.; Gutiérrez Suárez, A.; del Río Andrade, J.C. Biodegradation of lignocellulosics: Microbial, chemical, and enzymatic aspects of the fungal attack of lignin. Int. Microbiol. 2005, 8, 195–204. [Google Scholar]
- Pérez, J.; Muñoz-Dorado, J.; De La Rubia, T.; Martínez, J. Biodegradation and biological treatments of cellulose, hemicellulose and lignin: An overview. Int. Microbiol. 2002, 5, 53–63. [Google Scholar] [CrossRef]
- Quiroz-Castañeda, R.E.; Balcázar-López, E.; Dantán-González, E.; Martinez, A.; Folch-Mallol, J.; Martínez Anaya, C. Characterization of cellulolytic activities of Bjerkandera adusta and Pycnoporus sanguineus on solid wheat straw medium. Electron. J. Biotechnol. 2009, 12, 5–6. [Google Scholar]
- Quiroz-Castañeda, R.E.; Pérez-Mejía, N.; Martínez-Anaya, C.; Acosta-Urdapilleta, L.; Folch-Mallol, J. Evaluation of different lignocellulosic substrates for the production of cellulases and xylanases by the basidiomycete fungi Bjerkandera adusta and Pycnoporus sanguineus. Biodegradation 2011, 22, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.; Pinto, P.; Bezerra, R.; Dias, A.; Guedes, C.; Cardoso, V.; Cone, J.; Ferreira, L.; Colaço, J.; Sequeira, C. Effect of enzyme extracts isolated from white-rot fungi on chemical composition and in vitro digestibility of wheat straw. Anim. Feed Sci. Technol. 2008, 141, 326–338. [Google Scholar] [CrossRef]
- Klepzig, K.D.; Adams, A.S.; Handelsman, J.; Raffa, K.F. Symbioses: A Key Driver of Insect Physiological Processes, Ecological Interactions, Evolutionary Diversification, and Impacts on Humans. Environ. Entomol. 2009, 38, 67–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shil, R.K.; Mojumder, S.; Sadida, F.F.; Uddin, M.; Sikdar, D. Isolation and Identification of Cellulolytic Bacteria from the Gut of Three Phytophagus Insect Species. Braz. Arch. Biol. Technol. 2014, 57, 927–932. [Google Scholar] [CrossRef]
- Stefanini, I. Yeast-insect associations: It takes guts. Yeast 2018, 35, 315–330. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Li, Y.; Chen, Y.; Fu, C.; Long, W.; Xiao, X.; Liao, H.; Yang, Y. Bamboo lignocellulose degradation by gut symbiotic microbiota of the bamboo snout beetle Cyrtotrachelus buqueti. Biotechnol. Biofuels 2019, 12, 70. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Sheppard, D.C.; Joyce, J.A. Selected Life-History Traits of Black Soldier Flies (Diptera: Stratiomyidae) Reared on Three Artificial Diets. Ann. Entomol. Soc. Am. 2002, 95, 379–386. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Sheppard, D.C. Factors Influencing Mating and Oviposition of Black Soldier Flies (Diptera: Stratiomyidae) in a Colony. J. Entomol. Sci. 2002, 37, 345–352. [Google Scholar] [CrossRef]
- Zheng, L.; Crippen, T.L.; Holmes, L.; Singh, B.; Pimsler, M.; Benbow, M.E.; Tarone, A.; Dowd, S.; Yu, Z.; VanLaerhoven, S.L.; et al. Bacteria Mediate Oviposition by the Black Soldier Fly, Hermetia illucens (L.), (Diptera: Stratiomyidae). Sci. Rep. 2013, 3, 2563. [Google Scholar] [CrossRef] [Green Version]
- Ponnusamy, L.; Wesson, D.M.; Arellano, C.; Schal, C.; Apperson, C.S. Species Composition of Bacterial Communities Influences Attraction of Mosquitoes to Experimental Plant Infusions. Microb. Ecol. 2010, 59, 158–173. [Google Scholar] [CrossRef] [Green Version]
- Sharon, G.; Segal, D.; Zilber-Rosenberg, I.; Rosenberg, E. Symbiotic bacteria are responsible for diet-induced mating preference in Drosophila melanogaster, providing support for the hologenome concept of evolution. Gut Microbes 2011, 2, 190–192. [Google Scholar] [CrossRef] [Green Version]
- Boyce, G.R.; Gluck-Thaler, E.; Slot, J.C.; Stajich, J.E.; Davis, W.J.; James, T.Y.; Cooley, J.R.; Panaccione, D.G.; Eilenberg, J.; De Fine Licht, H.H.; et al. Psychoactive plant- and mushroom-associated alkaloids from two behavior modifying cicada pathogens. Fungal Ecol. 2019, 41, 147–164. [Google Scholar] [CrossRef] [PubMed]
- Cifuentes, Y.; Glaeser, S.P.; Mvie, J.; Bartz, J.-O.; Müller, A.; Gutzeit, H.O.; Vilcinskas, A.; Kampfer, P. The gut and feed residue microbiota changing during the rearing of Hermetia illucens larvae. Antonie van Leeuwenhoek 2020, 113, 1323–1344. [Google Scholar] [CrossRef] [PubMed]
- Holdemann, L.V.; Cato, E.P.; Moore, W.E.C. (Eds.) Anaerobe Laboratory Manual; Anaerobe Laboratory, Virginia Polytechnic Institute and State University: Blacksburg, VA, USA, 1977. [Google Scholar]
- Biddle, A.; Stewart, L.; Blanchard, J.L.; Leschine, S. Untangling the Genetic Basis of Fibrolytic Specialization by Lachnospiraceae and Ruminococcaceae in Diverse Gut Communities. Diversity 2013, 5, 627–640. [Google Scholar] [CrossRef]
- Zheng, L.; Crippen, T.L.; Singh, B.; Tarone, A.M.; Dowd, S.; Yu, Z.; Wood, T.K.; Tomberlin, J.K. A Survey of Bacterial Diversity From Successive Life Stages of Black Soldier Fly (Diptera: Stratiomyidae) by Using 16S rDNA Pyrosequencing. J. Med. Entomol. 2013, 50, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Korniłłowicz-Kowalska, T.; Rybczyńska-Tkaczyk, K. Growth conditions, physiological properties, and selection of optimal parameters of biodegradation of anticancer drug daunomycin in industrial effluents by Bjerkandera adusta CCBAS930. Int. Microbiol. 2019, 23, 287–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynd, L.R.; Weimer, P.J.; van Zyl, W.H.; Pretorius, I.S. Microbial cellulose utilization: Fundamentals and biotechnology. Microbiol. Mol. Biol. Rev. 2002, 66, 506–577. [Google Scholar] [CrossRef] [Green Version]
- Béguin, P. Molecular biology of cellulose degradation. Annu. Rev. Microbiol. 1990, 44, 219–248. [Google Scholar] [CrossRef]
- Million, M.; Raoult, D. Linking gut redox to human microbiome. Hum. Microbiome J. 2018, 10, 27–32. [Google Scholar] [CrossRef]
- Alves, L.M.C.; De Souza, J.A.M.; De Mello Varani, A.; De Macedo Lemos, E.G. The family Rhizobiaceae. In The Prokaryotes—Alphaproteobacteria and Betaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 419–437. [Google Scholar]
- Lambiase, A. The family Sphingobacteriaceae. In The Prokaryotes—Other Major Lineages of Bacteria and Archaea; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 907–914. [Google Scholar]
- Mayilraj, S.; Stackebrandt, E. The family Paenibacillaceae. In The Prokaryotes—Firmicutes and Tenericutes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 267–280. [Google Scholar]
- Stackebrandt, E. The family Lachnospiraceae. In The Prokaryotes—Firmicutes and Tenericutes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 197–201. [Google Scholar]
- Rainey, F.A. Family V Lachnospiraceae. In Bergey’s Manual of Systematic Bacteriology; De Vos, P., Garrity, G.M., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., Schleifer, K.H., Whitman, W.B., Eds.; Springer: New York, NY, USA, 2009; p. 921. [Google Scholar]
- Greening, C.; Geier, R.; Wang, C.; Woods, L.C.; Morales, S.E.; McDonald, M.J.; Rushton-Green, R.; Morgan, X.C.; Koike, S.; Leahy, S.C.; et al. Diverse hydrogen production and consumption pathways influence methane production in ruminants. ISME J. 2019, 13, 2617–2632. [Google Scholar] [CrossRef]
- Brune, A. Methanogenesis in the digestive tracts of insects and other arthropods. Biogenesis of hydrocarbons. In Handbook of Hydrocarbon and Lipid Microbiology; Stams, A.J.M., Sousa, D.Z., Eds.; Springer: Cham, Switzerland, 2019; pp. 229–260. [Google Scholar]
- Mertenat, A.; Diener, S.; Zurbrügg, C. Black Soldier Fly biowaste treatment—Assessment of global warming potential. Waste Manag. 2019, 84, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Parodi, A.; De Boer, I.J.M.; Gerrits, W.J.J.; Van Loon, J.J.A.; Heetkamp, M.J.W.; Van Schelt, J.; Bolhuis, J.E.; Van Zanten, H.H.E. Bioconversion efficiencies, greenhouse gas and ammonia emissions during black soldier fly rearing—A mass balance approach. J. Clean Prod. 2020, 271, 122488. [Google Scholar] [CrossRef]
- Ermolaev, E.; Lalander, C.; Vinneras, B. Greenhouse gas emissions from small-scale fly larvae composting with Hermetia illucens. Waste Manag. 2019, 96, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.C.d.S.; Diniz, A.G.; Tiago, P.V.; de Oliveira, N.T. Entomopathogenic Fusarium species: A review of their potential for the biological control of insects, implications and prospects. Fungal Biol. Rev. 2020, 34, 41–57. [Google Scholar] [CrossRef]
- Sutherland, J.B.; Pometto, A.L., III; Crawford, D.L. Lignocellulose degradation by Fusarium species. Can. J. Bot. 1983, 61, 1194–1198. [Google Scholar] [CrossRef]
- Huang, Y.; Busk, P.K.; Lange, L. Cellulose and hemicellulose-degrading enzymes in Fusarium commune transcriptome and functional characterization of three identified xylanases. Enzyme Microb. Technol. 2015, 73, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [Green Version]
- Bosch, G.; Van Der Fels-Klerx, H.J.; Rijk, T.C.D.; Oonincx, D.G. Aflatoxin B1 tolerance and accumulation in black soldier fly (Hermetia illucens) and yellow mealworms (Tenebrio molitor). Toxins 2017, 9, 185. [Google Scholar] [CrossRef]
- Langseth, W.; Bernhoft, A.; Rundberget, T.; Kosiak, B.; Gareis, M. Mycotoxin production and cytotoxicity of Fusarium strains isolated from Norwegian cereals. Mycopathologia 1999, 144, 103–113. [Google Scholar] [CrossRef]
- Marasas, W.F.O. Toxigenic fusaria. In Mycotoxins and Animal Foods; Smith, J.E., Henderson, R.S., Eds.; CRC Press: London, UK, 1991. [Google Scholar]
- Dickinson, E.; Harrison, M.; Parker, M.; Dickinson, M.; Donarski, J.; Charlton, A.; Nolan, R.; Rafat, A.; Gschwend, F.; Hallett, J.; et al. From waste to food: Optimizing the breakdown of oil palm waste to provide substrate for insects farmed as animal feed. PLoS ONE 2019, 14, e0224771. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Class | Order | Family | Species | Growth | Optical Lignin Degradation |
|---|---|---|---|---|---|---|
| Ascomycota | Leotiomycetes | Helotiales | Mollisiaceae | Mollisia lividofusca | - | - |
| Mollisia pilosa | - | - | ||||
| Sordariomycetes | Glomerellales | Plectosphaerellaceae | Plectosphaerella sp. | - | - | |
| Hypocreales | Cordycipitaceae | Isaria farinosa | + | - | ||
| Hypocreaceae | Trichoderma longipile | + | - | |||
| Trichoderma minutisporum | + | - | ||||
| Trichoderma polysporum | + | - | ||||
| Xylariales | Xylariaceae | Xylaria longipes | ++ | + | ||
| Basidiomycota | Agaricomycetes | Agaricales | Agaricaceae | Coprinus comatus | + | - |
| Coprinus xanthothrix | ++ | + | ||||
| Crepidotaceae | Crepidotus sp. | - | - | |||
| Marasmiaceae | Marasmius palmivorus | +++ | + | |||
| Mycenaceae | Mycena pseudocorticola | (−) | - | |||
| Omphalotaceae | Omphalotus illudens | (−) | - | |||
| Physalacriaceae | Armillaria bulbosa | + | - | |||
| Armillaria gallica | - | - | ||||
| Armillaria mellea | - | - | ||||
| Armillaria tabescens | - | - | ||||
| Pleurotaceae | Pleurotus calyptratus | + | - | |||
| Pleurotus cornucopiae | - | - | ||||
| Pleurotus eryngii | + | - | ||||
| Pleurotus flabellatus | ++ | - | ||||
| Pleurotus floridanus | ++ | + | ||||
| Pleurotus ostreatus (POS1) | ++ | + | ||||
| Pleurotus ostreatus (POS2) | ++ | + | ||||
| Pleurotus ostreatus (POS3) | + | + | ||||
| Pleurotus ostreatus (POS4) | - | - | ||||
| Pleurotus ostreatus (POS5) | + | - | ||||
| Pleurotus pulmonarius (PPU1) | + | - | ||||
| Pleurotus pulmonarius (PPU2) | + | - | ||||
| Pleurotus salmoneo-stramineus | + | - | ||||
| Pleurotus sapidus | ++ | - | ||||
| Psathyrellaceae | Coprinellus flocculosus | - | - | |||
| Strophariaceae | Agrocybe aegerita | + | - | |||
| Agrocybe perfecta | - | - | ||||
| Hypholoma fasciculare | - | - | ||||
| Pholiota lignicola | - | - | ||||
| Auriculariales | Auriculariaceae | Auricularia mesenterica | - | - | ||
| Gloeophyllales | Gloeophyllaceae | Gloeophyllum trabeum | + | - | ||
| Hymenochaetales | Hymenochaetaceae | Inonotus dryadeus | + | - | ||
| Polyporales | Fomitopsidaceae | Fomitopsis pinicola | - | - | ||
| Laetiporus sulphureus | - | - | ||||
| Piptoporus betulinus | - | - | ||||
| Irpicaceae | Irpex consors | +++ | + | |||
| Irpex vellereus | ++ | - | ||||
| Meruliaceae | Bjerkandera adusta | +++ | + | |||
| Ceriporiopsis rivulosa | - | - | ||||
| Emmia lacerata | + | - | ||||
| Phlebia radiata | + | - | ||||
| Phanerochaetaceae | Byssomerulius corium | + | - | |||
| Phanerochaete chrysosporium | ++ | + | ||||
| Polyporaceae | Dichomitus albidofuscus | + | - | |||
| Dichomitus campestris | ++ | - | ||||
| Dichomitus squalens | + | - | ||||
| Fomes fomentarius | - | - | ||||
| Ganoderma lucidum | - | - | ||||
| Ganoderma sp. | - | - | ||||
| Lentinus crinitus | - | - | ||||
| Lenzites betulinus | + | - | ||||
| Microporus affinis | + | - | ||||
| Pycnoporus cinnabarinus | - | - | ||||
| Pycnoporus coccineus | + | - | ||||
| Pycnoporus sanguineus | - | - | ||||
| Trametes versicolor (TVE1) | ++ | + | ||||
| Trametes versicolor (TVE2) | - | - | ||||
| Sparassidaceae | Sparassis crispa | - | - | |||
| Russulales | Bondarzewiaceae | Heterobasidion annosum | - | - | ||
| Peniophoraceae | Peniophora lycii | + | - | |||
| Stereaceae | Stereum sp. | ++ | - |
| Parameters | Sampling Size | CF 8 | NFR | BAD | |
|---|---|---|---|---|---|
| Hatching time (d) | ≥50% | 3.0 ± 0.8 a | 3.3 ± 0.5 a | 3.0 ± 0.0 a | |
| Larval development (d) 1 | ≥50% | 22.3 ± 0.5 a | 41.3 ± 0.9 b | 30.3 ± 1.7 c | |
| Prepupa-pupa (d) | n = 300 | 11.3 ± 1.7 a | 8.0 ± 0.0 a | 7.7 ± 1.3 a | |
| Intrapuparial metamorphosis (d) 2 | n = 300 | 10.7 ± 0.5 a | 10.0 ± 0.8 a | 9.7 ± 0.9 a | |
| Adult preoviposition period (d) 3 | n = 60 | 7.7 ± 0.8 a | 9.6 ± 1.0 a | 6.1 ± 1.0 a | |
| Total preoviposition period (d) 4 | n = 60 | 52.0 ± 2.0 a | 69.0 ± 2.6 b | 53.7 ± 1.6 a | |
| Total development (d) 5 | n = 300 | 47.3 ± 1.7 a | 62.7 ± 1.9 b | 50.7 ± 2.1 a | |
| Oviposition period (d) | n = 60 | 7.3 ± 2.1 a | 4.0 ± 2.2 a | 4.00 ± 0.8 a | |
| Oviposition span (min-max d) | n = 60 | 5–10 | 2–7 | 3–9 | |
| Successful development larva-prepupa (%) | n = 300 | 100.0 ± 0.0 a | 61.0 ± 16.8 ab | 81.3 ± 1.0 b | |
| Successful development prepupa-pupa (%) | n = 300 | 90.7 ± 0.5 a | 98.2 ± 0.5 b | 99.5 ± 0.7 b | |
| Successful development pupa-adult (%) | n = 300 | 97.0 ± 2.4 a | 98.7 ± 1.0 a | 100.0 ± 0.0 a | |
| Adult longevity (d) | ♂ | n = 60 | 17.7 ± 0.5 a | 13.3 ± 0.5 b | 13.0 ± 0.8 b |
| ♀ | n = 60 | 14.7 ± 0.2 a | 11.7 ± 0.5 b | 11.3 ± 0.5 b | |
| total | n = 120 | 16.2 ± 0.2 a | 12.7 ± 0.5 b | 12.0 ± 0.00 b | |
| Adult longevity (min-max d) | n = 120 | 4–25 | 7–24 | 5–19 | |
| Final larval weight (mg) | n = 150 | 303.0 ± 13.6 a | 149.3 ± 7.2 b | 187.0 ± 16.7 c | |
| Final larval length (mm) | n = 150 | 26.0 ± 0.1 a | 20.4 ± 0.1 b | 22.6 ± 0.1 c | |
| Weight prepupa (mg) | n = 150 | 219.6 ± 18.7 a | 130.3 ± 2.3 b | 179.7 ± 2.7 c | |
| Length prepupa (mm) | n = 150 | 23.4 ± 0.1 a | 19.0 ± 0.7 b | 20.8 ± 0.1 c | |
| Weight pupa (mg) | n = 150 | 169.0 ± 10.7 a | 121.0 ± 7.3 b | 151.6 ± 4.9 ab | |
| Length pupa (mm) | n = 150 | 22.7 ± 0.1 a | 20.1 ± 0.5 b | 21.1 ± 0.3 ab | |
| Weight adult (mg) | ♂ | n = 90 | 89.4 ± 8.5 a | 67.6 ± 6.6 a | 77.7 ± 2.9 a |
| ♀ | n = 90 | 103.6 ± 8.3 a | 83.5 ± 10.4 a | 94.9 ± 4.7 a | |
| total | n = 180 | 98.1 ± 8.0 a | 75.5 ± 8.0 a | 87.2 ± 2.3 a | |
| Length adult (mm) 6 | ♂ | n = 90 | 16.8 ± 0.3 a | 15.2 ± 0.3 b | 16.1 ± 0.1 a |
| ♀ | n = 90 | 17.5 ± 0.3 a | 16.4 ± 0.7 b | 17.3 ± 0.2 a | |
| total | n = 180 | 17.2 ± 0.2 a | 15.8 ± 0.5 b | 16.8 ± 0.1 a | |
| Sex ratio (♀/♂) | n = 180 | 1.5 ± 0.2 a | 1.0 ± 0.2 a | 1.3 ± 0.4 a | |
| Fertility (egg clutches/10 females) | n = 60 | 7.8 ± 0.9 a | 1.4 ± 0.4 b | 8.2 ± 0.9 a | |
| Egg clutch size (eggs/clutch) | n = 10 | 676.0 ± 59.6 a | 259.9 ± 59.7 b | 541.3 ± 59.1 c | |
| Span of egg clutch size (min-max eggs) | n = 10 | 548–763 | 172–334 | 375–589 | |
| Egg clutch weight (mg) | n = 10 | 16.5 ± 3.8 a | 4.6 ± 1.3 b | 11.1 ± 1.3 c | |
| Egg weight (mg/egg) 7 | n = 10 | 0.024 ± 0.005 a | 0.018 ± 0.002 a | 0.021 ± 0.001 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klüber, P.; Tegtmeier, D.; Hurka, S.; Pfeiffer, J.; Vilcinskas, A.; Rühl, M.; Zorn, H. Diet Fermentation Leads to Microbial Adaptation in Black Soldier Fly (Hermetia illucens; Linnaeus, 1758) Larvae Reared on Palm Oil Side Streams. Sustainability 2022, 14, 5626. https://doi.org/10.3390/su14095626
Klüber P, Tegtmeier D, Hurka S, Pfeiffer J, Vilcinskas A, Rühl M, Zorn H. Diet Fermentation Leads to Microbial Adaptation in Black Soldier Fly (Hermetia illucens; Linnaeus, 1758) Larvae Reared on Palm Oil Side Streams. Sustainability. 2022; 14(9):5626. https://doi.org/10.3390/su14095626
Chicago/Turabian StyleKlüber, Patrick, Dorothee Tegtmeier, Sabine Hurka, Janin Pfeiffer, Andreas Vilcinskas, Martin Rühl, and Holger Zorn. 2022. "Diet Fermentation Leads to Microbial Adaptation in Black Soldier Fly (Hermetia illucens; Linnaeus, 1758) Larvae Reared on Palm Oil Side Streams" Sustainability 14, no. 9: 5626. https://doi.org/10.3390/su14095626