
Photosynthesis, Yield and Quality of Soybean (Glycine max (L.) Merr.) under Different Soil-Tillage Systems


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment and Cultivation Management
2.2. Morpho-Physiological Measurements
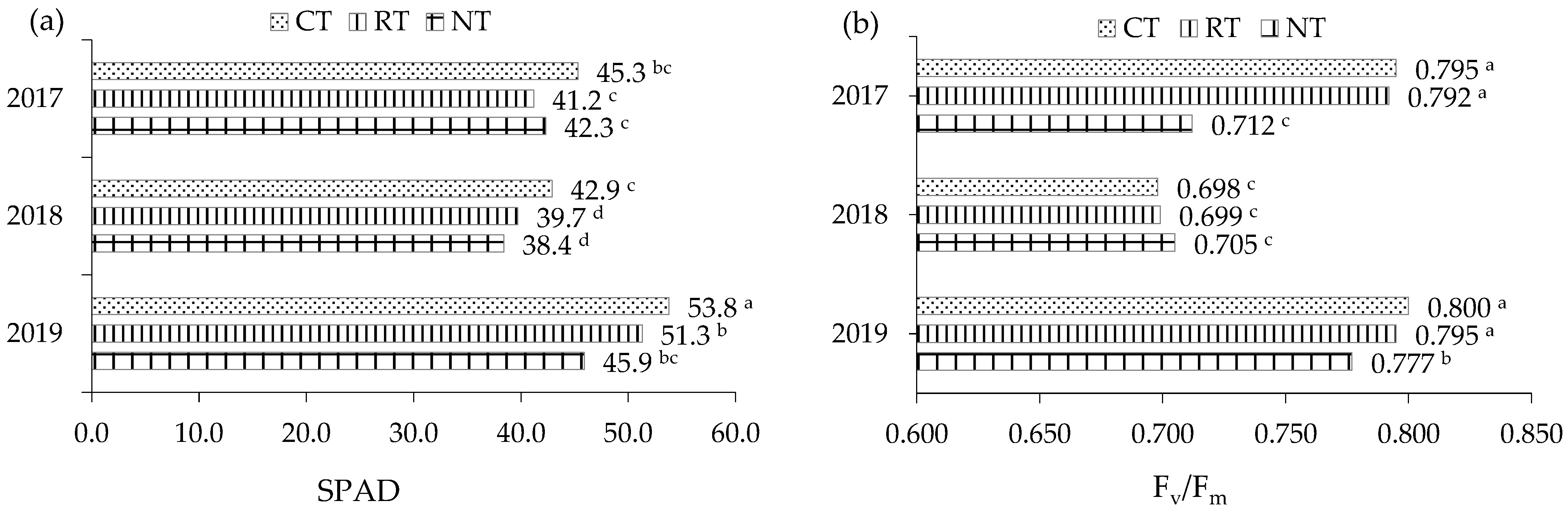
2.2.1. LAI and SPAD
2.2.2. Chlorophyll Fluorescence
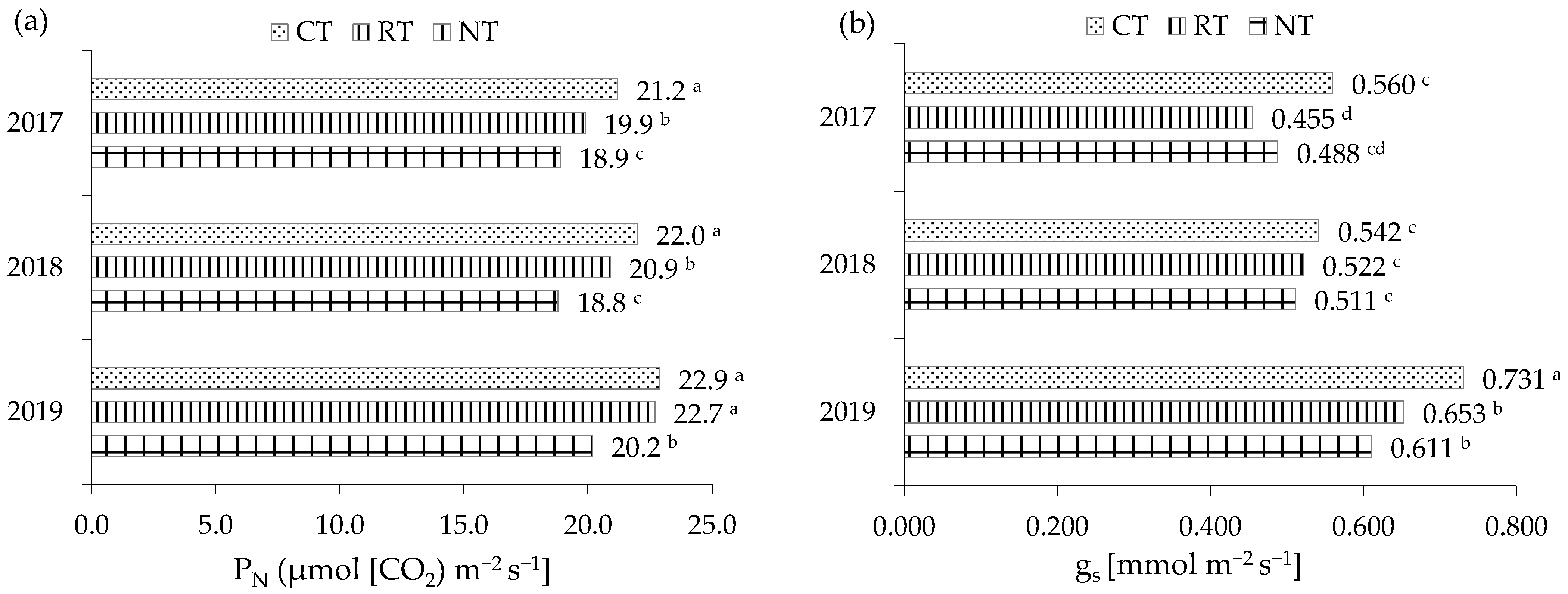
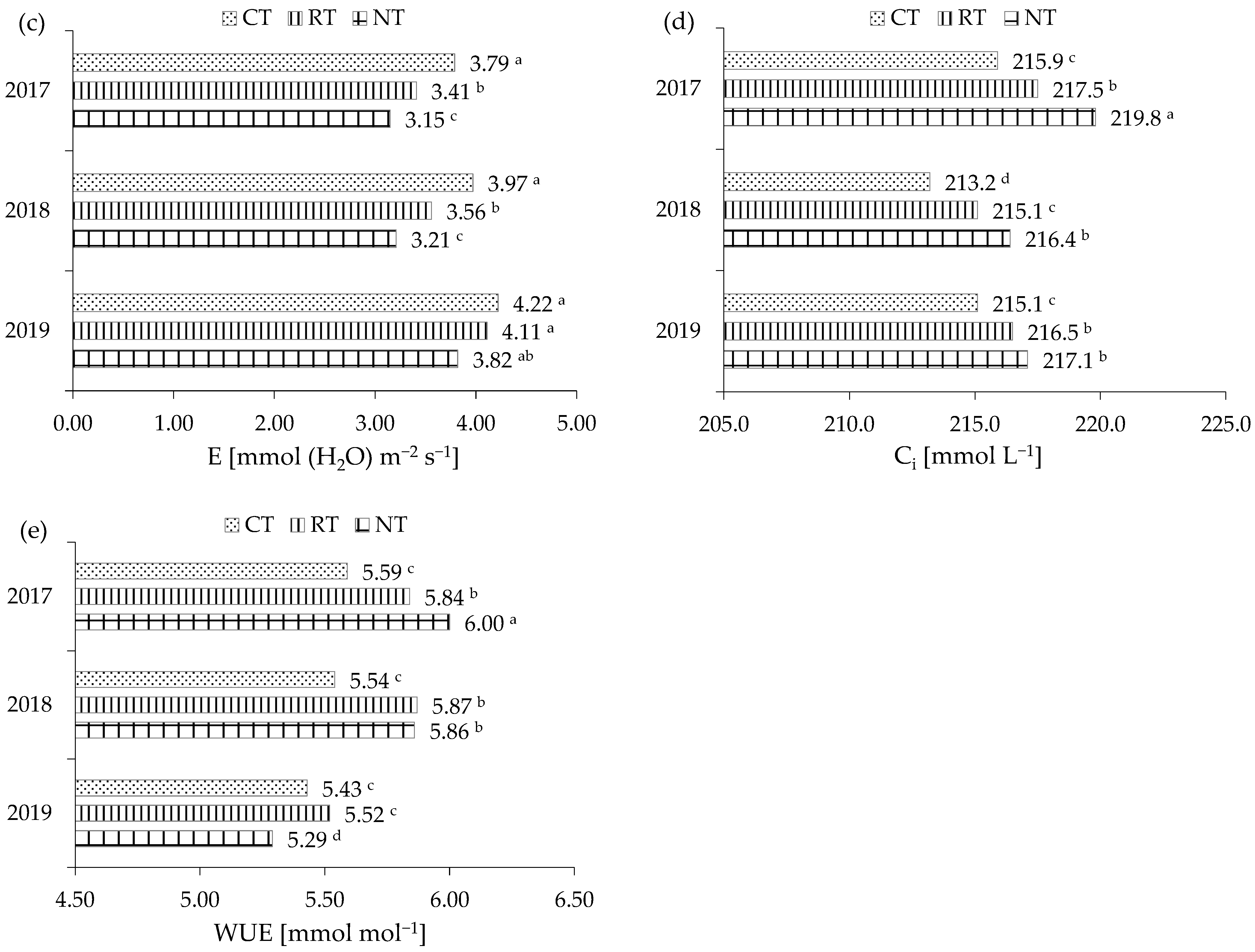
2.2.3. Gas Exchanges
2.3. Laboratory Analysis
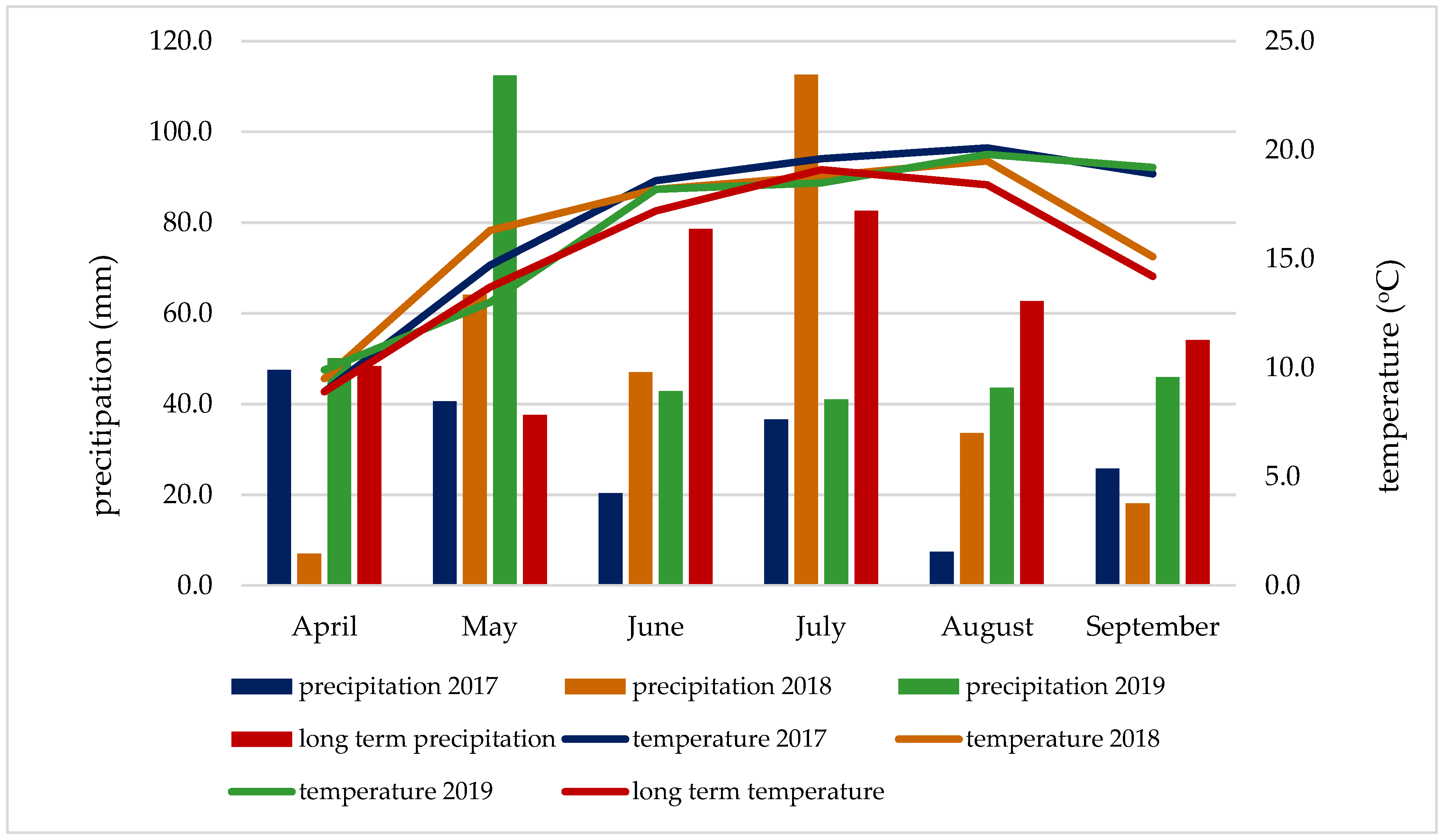
2.4. Soil and Meteorological Conditions
2.5. Statistical Analysis
3. Results and Discussion
3.1. Physiological Parameters
3.2. Protein and Fat Content
3.3. Seed Yield
3.4. Mineral Composition
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Houx, J.H.; Wiebold, W.J.; Fritschi, F.B. Rotation and tillage affect soybean grain composition, yield, and nutrient removal. Field Crop Sci. 2014, 64, 12–21. [Google Scholar] [CrossRef]
- Gawęda, D.; Nowak, A.; Haliniarz, M.; Woźniak, A. Yield and economic effectiveness of soybean grown under different cropping systems. Int. J. Plant Prod. 2020, 14, 475–485. [Google Scholar] [CrossRef] [Green Version]
- Farmaha, S.B.; Fernández, G.F.; Nafziger, D.E. No-till and strip-till soybean Production with surface and subsurface phosphorus and potassium fertilization. Agron. J. 2011, 103, 1862–1869. [Google Scholar] [CrossRef]
- Samarah, N.; Mullen, R.; Cianzio, S. Size distribution and mineral nutrients of soybean seeds in response to drought stress. J. Plant Nutr. 2004, 27, 815–835. [Google Scholar] [CrossRef]
- Piper, E.L.; Boote, K.J. Temperature and cultivar effects on soybean seed oil and protein concentrations. J. Am. Oil Chem. Soc. 1999, 76, 1233–1242. [Google Scholar] [CrossRef]
- Wrather, J.A.; Koenning, S.R. Estimates of disease effects on soybean yields in the United States 2003 to 2005. J. Nematol. 2006, 38, 173–180. [Google Scholar] [PubMed]
- Wrather, J.A.; Shannon, G.; Balardin, R.; Carregal, R.; Escobar, R.; Gupta, K.G.; Ma, Z.; Morel, W.; Ploper, D.; Tenuta, A. Effect of diseases on soybean yield in the top eight producing countries in 2006. Plant Health Progress. 2010, 11, 29. [Google Scholar] [CrossRef] [Green Version]
- Thiagalingam, K.; Gould, N.; Watson, P. Effect of tillage on rainfed maize and soybean yield and the nitrogen fertilizer requirements for maize. Soil Tillage Res. 1991, 19, 47–54. [Google Scholar] [CrossRef]
- Adamič, S.; Leskovšek, R. Soybean (Glycine max (L.) Merr.) Growth yield and nodulation in the early transition period from conventional tillage to conservation and no-tillage systems. Agronomy 2021, 11, 2477. [Google Scholar] [CrossRef]
- Sobko, O.; Hartung, J.; Zikeli, S.; Claupein, W.; Gruber, S. Effect of sowing density on grain yield, protein and oil content and plant morphology of soybean (Glycine max L. Merrill). Plant Soil Environ. 2019, 65, 594–601. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.Y.; Shi, S.; Wang, S. A multi-agent model-based evolutionary model of port service value network and decision preferences. Sustainability 2022, 14, 3565. [Google Scholar] [CrossRef]
- De Vita, P.; Di Paolo, E.; Fecondo, G.; Di Fonzo, N.; Pisante, M. No-tillage and conventional tillage effects on durum wheat yield, grain quality and soil moisture content in southern Italy. Soil Tillage Res. 2007, 92, 69–78. [Google Scholar] [CrossRef]
- Hama, J.R.; Kolpin, W.D.; LeFevre, H.G.; Hubbard, E.L.; Powers, M.M.; Strobel, W.B. Exposure and transport of alkaloids and phytoestrogens from soybeans to agricultural soils and streams in the midwestern United States. Environ. Sci. Technol. 2021, 55, 11029–11039. [Google Scholar] [CrossRef] [PubMed]
- Monsefi, A.; Sharma, A.R.; Rang Zan, N.; Behera, U.K.; Das, T.K. Effect of tillage and residue management on productivity of soybean and physic–chemical properties of soil in soybean–wheat cropping system. Int. J. Plant Prod. 2014, 8, 429–440. [Google Scholar] [CrossRef]
- Toliver, D.K.; Toliver, J.A.; Larson, J.A.; Roberts, R.K.; English, B.C.; De La Torre Ugarte, D.G.; West, T.O. Effects of no-till on yields as influenced by crop and environmental factors. Agron. J. 2012, 104, 530–541. [Google Scholar] [CrossRef] [Green Version]
- Biel, W.; Gawęda, D.; Łysoń, E.; Hury, G. Effect of variety and agrotechnical factors on nutritive value of soybean seeds. Acta Agrophys. 2017, 24, 395–404. (In Polish) [Google Scholar]
- Szwejkowska, B. Effect of cultivation intensity on protein content and yields in field pea. Acta Sci. Pol. Agric. 2005, 4, 153–161. (In Polish) [Google Scholar]
- Vollmann, J.; Fritz, N.C.; Wagentristl, H.; Ruckenbauer, P. Environmental and genetic variation of soybean seed protein content under Central European growing conditions. J. Sci. Food Agric. 2000, 80, 1300–1306. [Google Scholar] [CrossRef]
- Ohnishi, S.; Miyoshi, T.; Shirai, S. Low temperature stress at different flower developmental stages affects pollen development, pollination, and pod set in soybean. Environ. Exp. Bot. 2010, 69, 56–62. [Google Scholar] [CrossRef]
- Gong, W.Z.; Jiang, C.D.; Wu, Y.S.; Chen, H.H.; Liu, W.Y.; Yang, W.Y. Tolerance vs. avoidance: Two strategies of soybean (Glycine max) seedlings in response to shade in intercropping. Photosynthetica 2015, 53, 259–268. [Google Scholar] [CrossRef]
- Huang, Z.A.; Jiang, D.A.; Yang, Y.; Sun, J.W.; Jin, S.H. Effects of nitrogen deficiency on gas exchange, chlorophyll fluorescence, and antioxidant enzymes in leaves of rice plants. Photosynthetica 2004, 42, 357–364. [Google Scholar] [CrossRef]
- Woo, N.S.; Badger, M.R.; Pogson, B.J.A. Rapid, non-invasive procedure for quantitative assessment of drought survival using chlorophyll fluorescence. Plant Methods 2008, 4, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.; Hyldgaard, B.; Yu, X.Q.; Rosenqvist, E.; Ugarte, R.M.; Yu, S.X.; Wu, Z.; Ottosen, C.O.; Zhao, T. Phenotyping of faba beans (Vicia faba L.) under cold and heat stresses using chlorophyll fluorescence. Euphytica 2018, 214, 68. [Google Scholar] [CrossRef]
- Weber, F.J.; Kunz, C.; Peteinatos, G.G.; Santel, H.-J.; Gerhards, R. Utilization of chlorophyll fluorescence imaging technology to detect plant injury by herbicides in sugar beet and soybean. Weed Technol. 2017, 31, 523–535. [Google Scholar] [CrossRef]
- Meier, U. (Ed.) Growth Stages of Mono- and Dicotyledonous Plants. In BBCH Monograph, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Braunschweig, Germany, 2001; pp. 1–204. [Google Scholar]
- Hicks, S.; Lascano, R. Estimation of leaf area index for cotton canopies using the Li-Cor LAI 2000 plant canopy analyser. Agron. J. 1995, 87, 458–464. [Google Scholar] [CrossRef]
- Blackmer, T.M.; Schepers, J.S. Techniques for monitoring crop nitrogen status in corn. Commun. Soil Sci. Plant Anal. 1994, 25, 1791–1800. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Nobel, P.S. Physicochemical and Environmental Plant Physiology; Academic Press: Oxford, UK, 2009. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dąbrowski, P.; et al. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosyn. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Stuart, B.H. Infrared spectroscopy: Fundamentals and applications. In Analytical Techniques in the Sciences; Wiley: New York, NY, USA, 2004. [Google Scholar]
- Reference Base for Soil Resources 2014, Update 2015. In Word Soil Resources Reports No. 106; FAO: Rome, Italy, 2015; pp. 172–173. Available online: https://www.iuss.org/archived-content/archived-int-year-of-soils-2015/archived-working-groups-for-iys/working-group-world-reference-base-for-soil-resources-wrb/ (accessed on 4 February 2022).
- Skowera, B. Changes of hydrothermal conditions in the Polish area (1971–2010). Fragm. Agron. 2014, 31, 74–87. (In Polish) [Google Scholar]
- Peng, Y.; Gitelson, A.A. Remote estimation of gross primary productivity in soybean and maize based on total crop chlorophyll content. Remote Sens. Environ. 2012, 117, 440–448. [Google Scholar] [CrossRef]
- Xiong, D.; Chen, J.; Yu, T.; Gao, W.; Ling, X.; Li, Y.; Peng, S.; Huang, J. SPAD-based leaf nitrogen estimation is impacted by environmental factors and crop leaf characteristics. Sci. Rep. 2015, 5, 13389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiozza, M.V.; Parmley, K.A.; Higgins, R.H.; Singh, A.K.; Miguez, F.E. Comparative prediction accuracy of hyperspectral bands for different soybean crop variables: From leaf area to seed composition. Field Crops Res. 2021, 271, 108260. [Google Scholar] [CrossRef]
- Tang, J.H.; Su, L.L.; Li, Y.J.; Xu, W.X.; Peng, J.L. Effects of different tillage methods on photosynthetic characteristics, dry matter production and economic benefit of double cropping soybean. Chin. J. Appl. Ecol. 2016, 27, 182–190. [Google Scholar]
- Sabo, M.; Jug, D.; Jug, I. Dynamic chloroplast pigments concentration in leaves of soybean (Glycine max [L.] Merr.) under reduced tillage. Arch. Agron. Soil Sci. 2005, 51, 291–297. [Google Scholar] [CrossRef]
- Murchie, H.E.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badr, A.; Brüggemann, W. Special issue in honour of Prof. Reto, J. Strasser–Comparative analysis of drought stress response of maize genotypes using chlorophyll fluorescence measurements and leaf relative water content. Photosynthetica 2020, 58, 638–645. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Iqbal, N.; Brestic, M.; Raza, M.A.; Pang, T.; Langham, D.R.; Safdar, M.E.; Ahmeda, S.; Wena, B.; Gao, Y.; et al. Changes in morphology, chlorophyll fluorescence performance and Rubisco activity of soybean in response to foliar application of ionic titanium under normal light and shade environment. Sci. Total Environ. 2019, 658, 626–637. [Google Scholar] [CrossRef]
- Khalid, M.H.B.; Raza, M.A.; Yu, H.Q.; Sun, F.A.; Zhang, Y.Y.; Lu, F.Z.; Si, L.; Iqbal, N.; Khan, I.; Fu, F.L.; et al. Effect of shade treatments on morphology, photosynthetic and chlorophyll fluorescence characteristics of soybeans (Glycine max L. Merr.). Appl. Ecol. Environ. Res. 2019, 17, 2551–2569. [Google Scholar] [CrossRef]
- Gratani, L.; Catoni, R.; Pirone, G.; Frattaroli, R.A. Physiological and morphological leaf trait variations in two Apennine plant species in response to different altitudes. Photosynthetica 2012, 50, 15–23. [Google Scholar] [CrossRef]
- Gavili, E.; Moosavi, A.A.; Kamgar Haghighi, A.A. Does biochar mitigate the adverse effects of drought on the agronomic traits and yield components of soybean? Ind. Crop. Prod. 2019, 128, 445–454. [Google Scholar] [CrossRef]
- Savvides, A.M.; Fotopoulos, V. Two inexpensive and non-destructive techniques to correct for smaller-than-gasket leaf area in gas exchange measurements. Front. Plant Sci. 2018, 9, 548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buczek, J.; Migut, D.; Jańczak-Pieniązek, M. Effect of Soil Tillage Practice on Photosynthesis, Grain Yield and Quality of Hybrid Winter Wheat. Agriculture 2021, 11, 479. [Google Scholar] [CrossRef]
- Harrison, E.L.; Cubas, L.A.; Hepworth, J.E.G.C. The influence of stomatal morphology and distribution on photosynthetic gas exchange. Plant J. 2020, 101, 768–779. [Google Scholar] [CrossRef] [Green Version]
- Bojarszczuk, J. The influence of soil tillage system on changes in gas exchange parameters of Pisum sativum L. Agronomy 2021, 11, 1000. [Google Scholar] [CrossRef]
- Yao, X.; Li, C.; Li, S.; Zhu, Q.; Zhang, H.; Wang, H.; Yu, C.; St. Martin, K.S.; Xie, F. Effect of shade on leaf photosynthetic capacity, light-intercepting, electron transfer and energy distribution of soybeans. Plant Growth Regul. 2017, 83, 409–416. [Google Scholar] [CrossRef]
- Lawlor, D. Photosynthesis, productivity and environment. J. Exp. Bot. 1995, 46, 1449–1461. Available online: www.jstor.org/stable/23694991 (accessed on 1 February 2022). [CrossRef]
- Liu, F.L.; Christian, R.J.; Mathias, N.A. Drought stress effect on carbohydrate concentration in soybean leaves and pods during early reproductive development: Its implication in altering pod set. Field Crop. Res. 2004, 86, 1–13. [Google Scholar] [CrossRef]
- Bărdaş, M.; Chețan, F.; Şimon, A.; Deac, V.; Popa, A.; Oltean, V. Influence of classical and conservative tillage systems on physiological parameters and yield on soybean culture in the Transylvanian Plain. ProEnvironment 2019, 12, 208–215. [Google Scholar]
- Lamptey, S.; Li, L.; Xie, J.; Coulter, J.A. Tillage system affects soil water and photosynthesis of plastic-mulched maize on the semiarid Loess Plateau of China. Soil Tillage Res. 2020, 196, 104479. [Google Scholar] [CrossRef]
- Prior, S.S.; Runion, G.B.; Runion, B.G.; Rogers, H.H. Elevated atmospheric carbon dioxide effects on soybean and sorghum gas exchange in conventional and no-tillage systems. J. Environ. Qual. 2010, 39, 596–608. [Google Scholar] [CrossRef] [Green Version]
- Popović, V.; Vidić, M.; Jocković, D.; Ikanović, J.; Jaksic, S.; Cvijanoviv, G. Variability and correlations between yield components of soybean (Glycine max (L.) Merr.). Genetics 2012, 4, 33–45. [Google Scholar] [CrossRef]
- Fecák, P.; Šariková, D.; Cerný, I. Influence of tillage system and starting N fertilization on seed yield and quality of soybean Glycine max (L.) Merrill. Plant Soil Environ. 2010, 56, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Rodrígues, J.L.; Gamero, C.; Fernándes, J.C.; Mirás-Avalos, J. Effects of different soil tillage systems and coverages on soybean crop in the Botucatu Region in Brazil. Span. J. Agric. Res. 2009, 7, 173. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, F.G.; Brouder, M.S.; Volenec, J.J.; Beyrouty, A.C.; Hoyum, R. Root and shoot growth, seed composition, and yield components of no-till rainfed soybean under variable potassium. Plant Soil. 2009, 322, 125–138. [Google Scholar] [CrossRef]
- Szostak, B.; Głowacka, A.; Klebaniuk, R.; Kiełtyka-Dadasiewicz, A. Mineral composition of traditional non-GMO soybean cultivars in relation to nitrogen fertilization. Sci. World J. 2020, 2020, 9374564. [Google Scholar] [CrossRef]
- Jarecki, W.; Bobrecka-Jamro, D. Effect of fertilization with nitrogen and seed inoculation with nitragina on seed quality of soya bean (Glycine max (L) Merrill). Acta Sci. Pol. Agric. 2015, 14, 51–59. Available online: www.agricultura.acta.utp.edu.pl (accessed on 27 January 2022).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tillage System | Soil Treatment | ||
|---|---|---|---|
| After-Harvest | Autumn | Spring | |
| CT | shallow plowing (10–12 cm deep), harrowing | plowing (25–30 cm deep) | combined tillage unit (cultivator and string roller) |
| RT | disking (13–15 cm deep) | combined tillage unit (cultivator and string roller) | |
| NT | glyphosate at dose of 4.0 dm3 ha−1 | no plowing | sown directly into the stubble with a seeder with double disc coulters |
| Years | P | K | Mg | Fe | Zn | Mn | Cu |
|---|---|---|---|---|---|---|---|
| [mg kg−1] | |||||||
| 2017 | 203.0 | 274.1 | 26.2 | 2277.0 | 13.8 | 398.0 | 6.1 |
| 2018 | 130.2 | 181.0 | 51.2 | 2514.0 | 13.9 | 252.1 | 6.3 |
| 2019 | 74.0 | 251.2 | 55.7 | 2219.0 | 12.7 | 262.8 | 6.8 |
| Year | Month | Mean | |||||
|---|---|---|---|---|---|---|---|
| April | May | June | July | August | September | ||
| 2017 | 1.25 rd | 1.00 d | 0.38 ed | 0.61 vd | 0.23 ed | 0.37 ed | 0.65 vd |
| 2018 | 0.21 ed | 1.30 rd | 0.95 d | 1.87 rh | 0.65 vd | 0.36 ed | 0.89 d |
| 2019 | 1.74 rh | 2.60 vh | 0.98 d | 0.68 vd | 0.85 d | 0.92 d | 1.30 rd |
| long term | 1.76 rh | 1.85 rh | 1.60 o | 1.58 o | 1.25 rd | 1.00 d | 1.51 o |
| Specification | SPAD | LAI | Fv/Fm | Fv/F0 | PI |
|---|---|---|---|---|---|
| Tillage (T) | |||||
| CT | 47.3 a | 5.62 a | 0.764 a | 3.44 a | 5.54 a |
| RT | 44.1 b | 4.89 b | 0.762 a | 3.37 a | 5.28 a |
| NT | 42.2 b | 5.08 b | 0.731a | 3.03 b | 4.22 b |
| Year (Y) | |||||
| 2017 | 40.3 b | 4.99 b | 0.766 a | 2.42 b | 4.49 b |
| 2018 | 42.9 b | 5.19 ab | 0.701 a | 3.63 a | 5.11 b |
| 2019 | 50.3 a | 5.41 a | 0.791 a | 3.79 a | 5.44 a |
| Mean | 44.5 | 5.20 | 0.753 | 3.28 | 5.01 |
| ANOVA | |||||
| T | *** | ** | ns | * | * |
| Y | * | * | ns | * | * |
| T × Y | *** | ns | ns | ** | ** |
| Specification | PN (μmol (CO2) m−2 s−1) | gs (mol (H2O) m−2 s−1) | E (mmol (H2O) m−2 s−1) | Ci (mmol L−1) | WUE (mmol mol−1) |
|---|---|---|---|---|---|
| Tillage (T) | |||||
| CT | 22.0 a | 0.611 a | 3.99 a | 214.7 a | 5.52 b |
| RT | 21.2 a | 0.543 b | 3.69 ab | 216.4 a | 5.74 a |
| NT | 19.3 b | 0.537 b | 3.39 b | 217.8 a | 5.71 a |
| Year (Y) | |||||
| 2017 | 20.0 b | 0.501 b | 3.45 b | 217.7 a | 5.81 a |
| 2018 | 20.6 b | 0.525 b | 3.58 b | 216.2 a | 5.76 a |
| 2019 | 21.9 a | 0.665 a | 4.05 a | 214.9 a | 5.41 b |
| Mean | 20.8 | 0.564 | 3.69 | 216.3 | 5.66 |
| ANOVA | |||||
| T | *** | ** | ** | ** | ** |
| Y | * | * | *** | ns | ** |
| T × Y | * | * | *** | *** | *** |
| Specification | Protein Content (% DM) | Protein Yield (kg ha−1) | Fat Content (% DM) | Fat Yield (kg ha−1) | Seed Yield (t ha−1) |
|---|---|---|---|---|---|
| Tillage (T) | |||||
| CT | 34.1 a | 1179.0 a | 22.8 b | 794.0 a | 3.47 a |
| RT | 34.5 a | 1080.9 ab | 22.4 b | 705.6 a | 3.14 b |
| NT | 32.6 b | 998.8 b | 24.4 a | 755.3 a | 3.08 b |
| Year (Y) | |||||
| 2017 | 35.7 a | 1036.8 a | 22.1 c | 640.5 b | 2.90 c |
| 2018 | 33.9 b | 1075.0 a | 23.4 b | 739.3 ab | 3.17 b |
| 2019 | 31.6 c | 1146.8 a | 24.2 a | 875.1 a | 3.62 a |
| Mean | 33.7 | 1082.6 | 23.2 | 751.6 | 3.23 |
| ANOVA | |||||
| T | ** | ** | ** | ns | *** |
| Y | *** | ns | * | ** | *** |
| T × Y | ns | ns | ** | ns | ** |
| Specification | P | K | Ca | Mg | Fe | Cu | Mn | Zn |
|---|---|---|---|---|---|---|---|---|
| [g kg−1 DM] | [mg kg−1 DM] | |||||||
| Tillage (T) | ||||||||
| CT | 6.3 a | 15.0 b | 0.8 a | 2.0 a | 115.4 a | 20.9 a | 21.5 a | 52.1 a |
| RT | 6.5 b | 18.2 a | 1.0 a | 2.4 a | 114.9 a | 20.1 a | 20.4 a | 50.3 a |
| NT | 8.1 b | 14.8 b | 0.7 a | 2.1 a | 117.7 a | 19.8 a | 19.6 a | 49.9 a |
| Year (Y) | ||||||||
| 2017 | 6.4 b | 14.6 b | 0.6 a | 1.5 a | 114.3 a | 26.9 a | 24.3 a | 57.9 a |
| 2018 | 7.0 b | 16.0 b | 0.8 a | 2.3 a | 117.4 a | 16.4 b | 20.4 b | 52.1 b |
| 2019 | 7.5 a | 17.5 a | 1.0 a | 2.6 a | 116.2 a | 17.4 b | 16.8 c | 42.3 c |
| Mean | 7.0 | 16.0 | 0.8 | 2.1 | 116.0 | 20.2 | 20.5 | 50.8 |
| ANOVA | ||||||||
| T | ** | ** | ns | ns | ns | ns | ns | ns |
| Y | ** | ** | ns | ns | ns | ** | *** | *** |
| T × Y | ns | ns | ns | ns | ns | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buczek, J.; Bobrecka-Jamro, D.; Jańczak-Pieniążek, M. Photosynthesis, Yield and Quality of Soybean (Glycine max (L.) Merr.) under Different Soil-Tillage Systems. Sustainability 2022, 14, 4903. https://doi.org/10.3390/su14094903
Buczek J, Bobrecka-Jamro D, Jańczak-Pieniążek M. Photosynthesis, Yield and Quality of Soybean (Glycine max (L.) Merr.) under Different Soil-Tillage Systems. Sustainability. 2022; 14(9):4903. https://doi.org/10.3390/su14094903
Chicago/Turabian StyleBuczek, Jan, Dorota Bobrecka-Jamro, and Marta Jańczak-Pieniążek. 2022. "Photosynthesis, Yield and Quality of Soybean (Glycine max (L.) Merr.) under Different Soil-Tillage Systems" Sustainability 14, no. 9: 4903. https://doi.org/10.3390/su14094903