
Proximate Composition, Predictive Analysis and Allometric Relationships, of the Edible Water Frog (Pelophylax epeiroticus) in Lake Pamvotida (Northwest Greece)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Morphometry
2.3. Proximate Composition
2.4. Statistical Analysis
3. Results
3.1. Sex Ratio
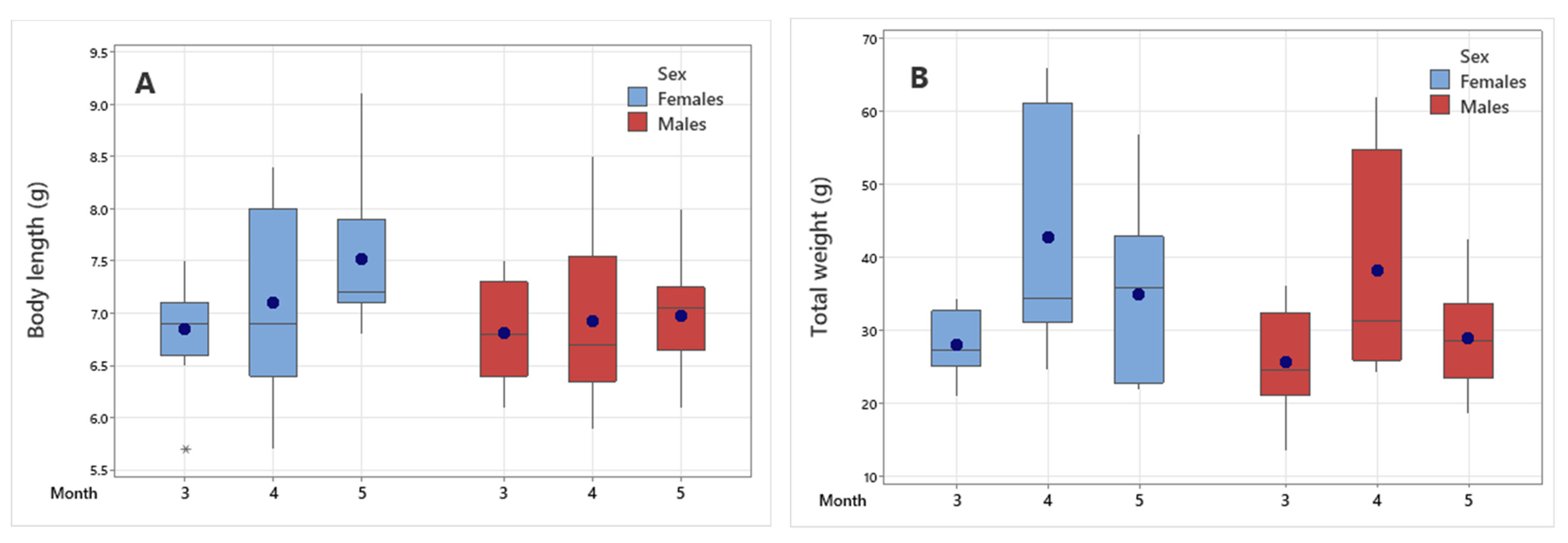
3.2. Population Structure
3.3. Allometry
3.4. Proximate Composition
Predictive Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sofianidou, T.S.; Schneider, H. Distribution range of the Epeirus frog Rana epeirotica (Amphibia: Anura) and the composition of the water frog populations in western Greece. Zool. Anz. 1989, 223, 13–25. [Google Scholar]
- Valakos, E.D. The Amphibians and Reptiles of Greece; Edition Chimaira: Frankfurt, Germany, 2008; ISBN 3899734610. [Google Scholar]
- IUCN SSC Amphibian Specialist Group. Pelophylax Epeiroticus. The IUCN Red List of Threatened Species; IUCN SSC Amphibian Specialist Group: Cambridge, UK, 2020. [Google Scholar]
- Kyriakopoulou-Sklavounou, P.; Sjögren, P.; Jansson, H. Discordant genetic and morphological differentiation between three Rana vidibunda populations from northern Greece. Hereditas 1991, 115, 51–61. [Google Scholar] [CrossRef]
- Herczeg, D.; Vörös, J.; Christiansen, D.G.; Benovics, M.; Mikulíček, P. Taxonomic composition and ploidy level among European water frogs (Anura: Ranidae: Pelophylax) in eastern Hungary. J. Zool. Syst. Evol. Res. 2017, 55, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Pafilis, P.; Kapsalas, G.; Lymberakis, P.; Protopappas, D.; Sotiropoulos, K. Diet composition of the karpathos marsh frog (Pelophylax cerigensis): What does the most endangered frog in europe eat? Anim. Biodivers. Conserv. 2019, 42, 1–8. [Google Scholar] [CrossRef]
- Hermaniuk, A.; Czajkowska, M.; Borkowska, A.; Taylor, J.R.E. Body size variation in hybrids among populations of European water frogs (Pelophylax esculentus complex) with different breeding systems. Amphib. Reptil. 2020, 41, 361–371. [Google Scholar] [CrossRef]
- Sagonas, K.; Karameta, E.; Kotsakiozi, P.; Poulakakis, N. Cross-species testing of nuclear markers in Pelophylax water frogs in Greece and examination of their power to detect genetic admixture. Amphib. Reptil. 2020, 41, 253–259. [Google Scholar] [CrossRef]
- Zhelev, Z.M.; Arnaudova, D.N.; Popgeorgiev, G.S.; Tsonev, S.V. In situ assessment of health status and heavy metal bioaccumulation of adult Pelophylax ridibundus (Anura: Ranidae) individuals inhabiting polluted area in southern Bulgaria. Ecol. Indic. 2020, 115, 106413. [Google Scholar] [CrossRef]
- Katsiyiannis, P.; Tzoras, E. First record of Pelophylax kurtmuelleri preying on Hyla arborea in Greece. Parnass. Arch. 2020, 8, 17–18. [Google Scholar]
- Karatsivou, E. Sustainability Model of Frog Farming in Greece; Unveristy of Thessaly: Volos, Greece, 2014. [Google Scholar]
- Kyriakopoulou-Sklavounou, P.; Stylianou, P.; Tsiora, A. A skeletochronological study of age, growth and longevity in a population of the frog Rana ridibunda from southern Europe. Zoology 2008, 111, 30–36. [Google Scholar] [CrossRef]
- Duellman, W.E.; Trueb, L. Biology of Amphibians; JHU Press: Baltimore, MD, USA, 1994; ISBN 080184780X. [Google Scholar]
- Marques, S.M.; Chaves, S.; Gonçalves, F.; Pereira, R. Evaluation of growth, biochemical and bioaccumulation parameters in Pelophylax perezi tadpoles, following an in-situ acute exposure to three different effluent ponds from a uranium mine. Sci. Total Environ. 2013, 445, 321–328. [Google Scholar] [CrossRef]
- Franco-de-Sá, J.F.O.; Val, A.L. Copper toxicity for Scinax ruber and Rhinella granulosa (Amphibia: Anura) of the Amazon: Potential of Biotic Ligand Model to predict toxicity in urban streams. Acta Amaz. 2014, 44, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Taiwo, I.E.; Henry, A.N.; Imbufe, A.P.; Adetoro, O.O. Heavy metal bioaccumulation and biomarkers of oxidative stress in the wild African tiger frog, Hoplobatrachus occipitalis. Afr. J. Environ. Sci. Technol. 2014, 8, 6–15. [Google Scholar]
- Mahmood, I.; Imadi, S.R.; Shazadi, K.; Gul, A.; Hakeem, K.R. Effects of pesticides on environment. In Plant, Soil and Microbes; Springer: Berlin/Heidelberg, Germany, 2016; pp. 253–269. [Google Scholar]
- Jayawardena, U.A.; Angunawela, P.; Wickramasinghe, D.D.; Ratnasooriya, W.D.; Udagama, P.V. Heavy metal–induced toxicity in the Indian green frog: Biochemical and histopathological alterations. Environ. Toxicol. Chem. 2017, 36, 2855–2867. [Google Scholar] [CrossRef] [PubMed]
- Corduk, N.; Hacioglu-Dogru, N.; Gul, C.; Tosunoglu, M. Monitoring of micronuclei and nuclear abnormalities in Pelophylax ridibundus erythrocytes from the Biga Stream (Canakkale, Turkey). Fresenius Environ. Bull. 2018, 27, 147–153. [Google Scholar]
- Pollo, F.; Bionda, C.; Otero, M.; Grenat, P.; Babini, S.; Flores, P.; Grisolia, M.; Salas, N.; Martino, A. Morphological abnormalities in natural populations of the common South American toad Rhinella arenarum inhabiting fluoride-rich environments. Ecotoxicol. Environ. Saf. 2019, 177, 32–38. [Google Scholar] [CrossRef]
- Altherr, S.; Goyenechea, A.; Schubert, D.J. The International Trade in Frogs’legs and Its Ecological Impact; Pro Wildlife Munich (DE): München, Germany, 2011. [Google Scholar]
- Agriculture Organization of the United Nations. The State of World Fisheries and Aquaculture; Food & Agriculture Org.: Rome, Italy, 2000; Volume 3, ISBN 9251044929. [Google Scholar]
- Blé, Y.C.; Yobouet, B.A.; Dadié, A. Consumption, proximate and mineral composition of edible frog Hoplobatrachus occipitalis from midwest areas of Côte d’Ivoire. AJSR 2016, 5, 16–20. [Google Scholar]
- Neveu, A. La raniculture est-elle une alternative à la récolte? Etat actuel en France. Prod. Anim. 2004, 17, 167–175. [Google Scholar] [CrossRef]
- Teixeira, R.D.; Mello, S.C.R.P.; dos Santos, C.A.M.L. The World Market for Frog Legs; Food and Agriculture Organization of the United Nations: Rome, Italy, 2001. [Google Scholar]
- Tokur, B.; Gürbüz, R.D.; Özyurt, G. Nutritional composition of frog (Rana esculanta) waste meal. Bioresour. Technol. 2008, 99, 1332–1338. [Google Scholar] [CrossRef]
- Warkentin, I.G.; Bickford, D.; Sodhi, N.S.; Bradshaw, C.J.A. Eating frogs to extinction. Conserv. Biol. 2009, 23, 1056–1059. [Google Scholar] [CrossRef]
- Baygar, T.; Ozgur, N. Sensory and chemical changes in smoked frog (Rana esculanta) leg during cold storage (4 °C ± 1). J. Anim. Vet. Adv. 2010, 9, 588–593. [Google Scholar] [CrossRef]
- Onadeko, A.B.; Egonmwan, R.I.; Saliu, J.K. Edible amphibian species: Local knowledge of their consumption in Southwest Nigeria and their nutritional value. West Afr. J. Appl. Ecol. 2012, 19, 67–76. [Google Scholar]
- Mohneke, M.; Rödel, M.-O. Declining amphibian populations and possible ecological consequences—A review. Salamandra 2009, 45, 203–210. [Google Scholar]
- Omoniyi, L.O.; Ajibola, M.E.; Bifarin, J.O. Demand Analysis for Frog Meat in Ondo State. Glob. J. Sci. Front. Res. Agric. Biol. 2012, 12, 9–12. [Google Scholar]
- Schaefer, G.L.; Cosh, M.H.; Jackson, T.J. The USDA natural resources conservation service soil climate analysis network (SCAN). J. Atmos. Ocean. Technol. 2007, 24, 2073–2077. [Google Scholar] [CrossRef]
- Alkaya, A.; Sereflisan, H.; Dikel, S.; Sereflisan, M. Comparison of Pond-Raised and Wild Female Marsh Frog (Pelophylax ridibundus) with Respect to Proximate Composition and Amino Acids Profiles. Fresenius Environ. Bull. 2018, 27, 6330–6336. [Google Scholar]
- Cagiltay, F.; Erkan, N.; Selcuk, A.; Ozden, O.; Devrim Tosun, D.; Ulusoy, S.; Atanasoff, A. Chemical composition of wild and cultured marsh frog (Rana ridibunda). Bulg. J. Agric. Sci. 2014, 20, 1250–1254. [Google Scholar]
- Afonso, A.M.; Fonseca, A.B.M.; Conte-Junior, C.A.; Mársico, E.T.; de Freitas, M.Q.; Mano, S.B. Cauda de rã: Uma fonte proteica para alimentar o futuro. Bol. Do Inst. Pesca 2017, 43, 112–123. [Google Scholar] [CrossRef]
- Charasrosjanakul, A.; Laohasongkram, K.; Chaiwanichsiri, S. Effects of Garlic and Rosemary Essential Oil and Packaging Conditions on Physical and Chemical Properties of Frog Leg Meat during Refrigerated Storage. In Proceedings of the 5th International Conference on Agriculture, Environment and Biological Sciences (ICAEBS-16), Pattaya, Thailand, 28–29 April 2016. [Google Scholar]
- Özogul, F.; Özogul, Y.; Olgunoglu, A.I.; Boga, E.K. Comparison of fatty acid, mineral and proximate composition of body and legs of edible frog (Rana esculenta). Int. J. Food Sci. Nutr. 2008, 59, 558–565. [Google Scholar] [CrossRef]
- Hatziioannou, M.; Karatsivou, E. Bioeconomic Analysis of a Pilot Commercial Production for Frogs (Lithobates Catesbeianus), In Greece. Oceanogr. Fish. Open Access J. 2020, 12, 137–140. [Google Scholar] [CrossRef]
- Wilson, G.P.; Frogley, M.R.; Roucoux, K.H.; Jones, T.D.; Leng, M.J.; Lawson, I.T.; Hughes, P.D. Limnetic and terrestrial responses to climate change during the onset of the penultimate glacial stage in NW Greece. Glob. Planet. Chang. 2013, 107, 213–225. [Google Scholar] [CrossRef]
- Romero, J.R.; Kagalou, I.; Imberger, J.; Hela, D.; Kotti, M.; Bartzokas, A.; Albanis, T.; Evmirides, N.; Karkabounas, S.; Papagiannis, J. Seasonal water quality of shallow and eutrophic Lake Pamvotis, Greece: Implications for restoration. Hydrobiologia 2002, 474, 91–105. [Google Scholar] [CrossRef]
- Perdikaris, C.; Georgiadis, C. Co-occurrence of narrow-clawed crayfish (Astacus leptodactylus sensu lato) and noble crayfish (Astacus astacus L.) in the southwestern Balkans: The case of Lake Pamvotida (NW Greece). North-West. J. Zool. 2017, 13, 18–26. [Google Scholar]
- Stefanidis, K.; Kostara, A.; Papastergiadou, E. Implications of human activities, land use changes and climate variability in Mediterranean lakes of Greece. Water 2016, 8, 483. [Google Scholar] [CrossRef]
- Borzée, A.; Park, S.; Kim, A.; Kim, H.-T.; Jang, Y. Morphometrics of two sympatric species of tree frogs in Korea: A morphological key for the critically endangered Hyla suweonensis in relation to H. japonica. Anim. Cells Syst. 2013, 17, 348–356. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis. In Association of Official Analytical Chemistrs, 19th ed.; AOAC: Washington, DC, USA, 2012. [Google Scholar]
- Krishnamoorthy, K.; Lu, F.; Mathew, T. A parametric bootstrap approach for ANOVA with unequal variances: Fixed and random models. Comput. Stat. Data Anal. 2007, 51, 5731–5742. [Google Scholar] [CrossRef]
- Hampton, R.E.; Havel, J.E. Introductory Biological Statistics; Waveland Press: Long Grove, IL, USA, 2006; ISBN 1577663802. [Google Scholar]
- Şahin, M.D.; Aybek, E.C. Jamovi: An easy to use statistical software for the social scientists. Int. J. Assess. Tools Educ. 2019, 6, 670–692. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.K.; Pal, S. Data mining: A prediction for performance improvement of engineering students using classification. arXiv 2012, arXiv:1203.3832. [Google Scholar]
- Demšar, J.; Curk, T.; Erjavec, A.; Gorup, Č.; Hočevar, T.; Milutinovič, M.; Možina, M.; Polajnar, M.; Toplak, M.; Starič, A. Orange: Data mining toolbox in Python. J. Mach. Learn. Res. 2013, 14, 2349–2353. [Google Scholar]
- Cagiltay, F.; Erkan, N.; Tosun, D.; Selcuk, A. Chemical composition of the frog legs (Rana ridibunda). Fleischwirlschaft Int. 2011, 26, 78–81. [Google Scholar]
- Çaklı, Ş.; Kışla, D.; Cadun, A.; Dinçer, T.; Cağlak, E. Determination of Shelf Life in Fried and Boiled Frog Meat Stored in Refrigerator in 3.2 ± 1.08 °C. Ege J. Fish. Aquat. Sci. 2009, 26, 115–119. [Google Scholar]
- Haddad, L.R.; Tejada-Ortigoza, V.; Martín-del-Campo, S.T.; Balderas-León, I.; Morales-de la Peña, M.; Garcia-Amezquita, L.E.; Welti-Chanes, J. Evaluation of nutritional composition and technological functionality of whole American Bullfrog (Lithobates catesbeianus), its skin, and its legs as potential food ingredients. Food Chem. 2022, 372, 131232. [Google Scholar] [CrossRef] [PubMed]
- Kaya, Y.; Erdem, M.E.; Turan, H. Monthly differentiation in meat yield, Chemical and amino acid composition of wild and cultured brown trout (Salmo trutta forma fario Linneaus, 1758). Turk. J. Fish. Aquat. Sci. 2014, 14, 479–486. [Google Scholar] [CrossRef]
- Şahin, Ş.A.; Başçinar, N.; Kocabaş, M.; Tufan, B.; Köse, S.; Okumuş, I. Evaluation of meat yield, proximate composition and fatty acid profile of cultured brook trout (Salvelinus fontinalis Mitchill, 1814) and black sea trout (Salmo trutta labrax Pallas, 1811) in comparison with their hybrid. Turk. J. Fish. Aquat. Sci. 2011, 11, 261–271. [Google Scholar] [CrossRef]
- Usydus, Z.; Szlinder-Richert, J.; Adamczyk, M.; Szatkowska, U. Marine and farmed fish in the Polish market: Comparison of the nutritional value. Food Chem. 2011, 126, 78–84. [Google Scholar] [CrossRef]
- Ljubojevic, D.; Trbovic, D.; Lujic, J.; Bjelic-Cabrilo, O.; Kostic, D.; Novakov, N.; Cirkovic, M. Fatty acid composition of fishes from Inland Waters. Bulg. J. Agric. Sci. 2013, 19, 62–71. [Google Scholar]
- Al-Anbari, I.H.A.; Abbas, A.A.; Al-Ani, M.T.H. Effect of adding table salt and citric acid on some properties of Iraqi fish (Liza abo mugilidae) after cookingin a microwave oven. Plant Arch. 2019, 19, 3043–3049. [Google Scholar]
- Shirose, L.J.; Brooks, R.J.; Barta, J.R.; Desser, S.S. Intersexual differences in growth, mortality, and size at maturity in bullfrogs in central Ontario. Can. J. Zool. 1993, 71, 2363–2369. [Google Scholar] [CrossRef]
- Tsiora, A.; Kyriakopoulou-Sklavounou, P. A skeletochronological study of age and growth in relation to adult size in the water frog Rana epeirotica. Zoology 2002, 105, 55–60. [Google Scholar] [CrossRef]
- Tsiora, A.; Kyriakopoulou-Sklavounou, P. Female reproductive cycle of the water frog Rana epeirotica in northwestern Greece. Amphib. Reptil. 2002, 23, 269–280. [Google Scholar] [CrossRef]
- Gould, S.J. Allometry and size in ontogeny and phylogeny. Biol. Rev. 1966, 41, 587–638. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Size, shape, and form: Concepts of allometry in geometric morphometrics. Dev. Genes Evol. 2016, 226, 113–137. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P. Heterochrony and allometry: The analysis of evolutionary change in ontogeny. Biol. Rev. 1998, 73, 79–123. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B. Ecological character displacement and the study of adaptation. Proc. Natl. Acad. Sci. USA 2000, 97, 5693–5695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moen, D.S.; Irschick, D.J.; Wiens, J.J. Evolutionary conservatism and convergence both lead to striking similarity in ecology, morphology and performance across continents in frogs. Proc. R. Soc. B Biol. Sci. 2013, 280, 20132156. [Google Scholar] [CrossRef] [Green Version]
- Schmidt-Nielsen, K.; Knut, S.-N. Scaling: Why Is Animal Size So Important? Cambridge University Press: Cambridge, UK, 1984; ISBN 0521319870. [Google Scholar]
- Landis, M.J.; Schraiber, J.G. Pulsed evolution shaped modern vertebrate body sizes. Proc. Natl. Acad. Sci. USA 2017, 114, 13224–13229. [Google Scholar] [CrossRef] [Green Version]
- Levy, D.L.; Heald, R. Biological scaling problems and solutions in amphibians. Cold Spring Harb. Perspect. Biol. 2016, 8, a019166. [Google Scholar] [CrossRef] [Green Version]
- Morton, M.L. Seasonal changes in total body lipid and liver weight in the Yosemite toad. Copeia 1981, 1981, 234–238. [Google Scholar] [CrossRef]
- Schlaghecke, R.; Blüm, V. Seasonal variations in fat body metabolism of the green frog Rana esculenta (L.). Experientia 1978, 34, 1019–1020. [Google Scholar] [CrossRef]
- Tsiora, A.; Kyriakopoulou-Sklavounou, P. Male reproductive cycle of the water frog Rana epeirotica in northwestern Greece. Amphib. Reptil. 2001, 22, 291–302. [Google Scholar]
- Carey, C.; Alexander, M.A. Climate change and amphibian declines: Is there a link? Divers. Distrib. 2003, 9, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Alan Pounds, J.; Bustamante, M.R.; Coloma, L.A.; Consuegra, J.A.; Fogden, M.P.L.; Foster, P.N.; La Marca, E.; Masters, K.L.; Merino-Viteri, A.; Puschendorf, R. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 2006, 439, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Tryjanowski, P.; Sparks, T.; Rybacki, M.; Berger, L. Is body size of the water frog Rana esculenta complex responding to climate change? Naturwissenschaften 2006, 93, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Mosavi, J.; Vaissi, S.; Dastansara, N.; Sharifi, M. Effects of temperature on growth, development and survival in larvae of Pelophylax ridibundus (Pallas, 1771) (Amphibia: Anura): Linking global warming to amphibian development. Acta Zool. Bulg. 2017, 69, 541–546. [Google Scholar]
- Reading, C.J. The effect of winter temperatures on the timing of breeding activity in the common toad Bufo bufo. Oecologia 1998, 117, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Al-Radaideh, Q.A.; Al Nagi, E. Using data mining techniques to build a classification model for predicting employees performance. Int. J. Adv. Comput. Sci. Appl. 2012, 144–151. [Google Scholar]
- Mao, M.; Mi, Z.P.; Yang, Z.S.; Zhou, C.Q. Sexual dimorphism in the limb muscles of the dark-spotted frog, Pelophylax nigromaculata. Herpetol. J. 2014, 24, 147–153. [Google Scholar]
- Andersson, M.B. Sexual Selection; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Liao, W.B. Evolution of sexual size dimorphism in a frog obeys the inverse of Rensch’s rule. Evol. Biol. 2013, 40, 293–299. [Google Scholar] [CrossRef]
- Hedrick, A.V.; Temeles, E.J. The evolution of sexual dimorphism in animals: Hypotheses and tests. Trends Ecol. Evol. 1989, 4, 136–138. [Google Scholar] [CrossRef]
- Shine, R. Sexual selection and sexual dimorphism in the Amphibia. Copeia 1979, 297–306. [Google Scholar] [CrossRef]
- Li, S.T.; Wu, X.; Li, D.Y.; Lou, S.L.; Mi, Z.P.; Liao, W.B. Body size variation of odorous frogs (Odorrana grahami) across altitudinal gradients. Herpetol. J. 2013, 23, 187–192. [Google Scholar]
- Halliday, T. Intersexual selection and alternative mating behaviour. Amphib. Biol. 1995, 2, 419–468. [Google Scholar]
- Liao, W.B.; Zeng, Y.; Yang, J.D. Sexual size dimorphism in anurans: Roles of mating system and habitat types. Front. Zool. 2013, 10, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Number of Individuals | Proportion | χ² Goodness of Fit | |||
|---|---|---|---|---|---|---|
| Female | Observed | 31.0 | 0.36 | χ² | df | p |
| Expected | 42.5 | 0.50 | 6.22 | 1 | <0.05 | |
| Male | Observed | 54.0 | 0.64 | |||
| Expected | 42.5 | 0.50 | ||||
| Morphometric Relationships | Equation Comparison | ||||||
|---|---|---|---|---|---|---|---|
| Sampling Site | Equation | N | R2 | t-Test | Allometry | Slopes | Intercepts |
| Total population | TW = 0.0793345 × BL 3.08249 | 85 | 84.4% | ns | Positive | ||
| Females | TW = 0.147744 × BL 2.78335 | 31 | 85.7% | ns | Negative | ns | ns |
| Males | TW = 0.0446303 × BL 3.36766 | 54 | 85.6% | ns | Positive | ||
| Total population | BL = 2.62897 × FL 0.723272 | 85 | 93.3% | *** | Negative | ||
| Females | BL = 2.2427 × FL 0.839318 | 31 | 96.3% | ns | Negative | *** | *** |
| Males | BL = 2.9084 × FL 0.647612 | 54 | 95.6% | *** | Negative | ||
| Total population | BL = 2.90683 × TL 0.696432 | 85 | 92.8% | *** | Negative | ||
| Females | BL = 3.13275 × TL 0.632722 | 31 | 94.7% | *** | Negative | ns | ns |
| Males | BL = 2.70909 × TL 0.758638 | 54 | 96.5% | *** | Negative | ||
| Total population | BL = 6.14082 × SED 0.70285 | 85 | 87.8% | ** | Negative | ||
| Females | BL = 5.81649 × SED 0.95948 | 31 | 92.9% | ns | Negative | *** | ns |
| Males | BL = 6.25962 × SED 0.592945 | 54 | 92.5% | *** | Negative | ||
| Total population | MHL = 1.24333 × MHW 0.958627 | 85 | 94.8% | ns | Negative | ||
| Females | MHL = 1.41502 × MHW 0.829075 | 31 | 96.8% | ns | Negative | ns | ns |
| Total population | MHL = 1.24333 × MHW 0.958627 | 85 | 94.8% | ns | Negative | ||
| Origin | Species | Moisture | Ash | Protein | Fat | Origin | Reference |
|---|---|---|---|---|---|---|---|
| Cultured | Pelophylax ridibundus | 74.79 ± 0.25 *** | 1.00 ± 0.15 ns | 22.95 ± 0.16 *** | 0.93 ± 0.20 *** | Turkey | [34] |
| Pelophylax ridibundus | 79.15 ± 0.77 *** | 1.03 ± 0.01 ns | 19.69 ± 0.17 *** | 0.71 ± 0.03 ** | Turkey | [33] | |
| Lithobates catesbeianus | 84.81 ± 0.12 *** | 0.94 ± 0.27 ns | 15.24 ± 0.67 ns | 0.06 ± 0.0 * | Brazil | [35] | |
| Holobatrachus rugulosus | 79.47 ± 0.12 ** | 0.77 ± 0.05 ** | 21.89 ± 0.16 *** | 0.8 ± 0.07 *** | Thailand | [36] | |
| Pelophylax epeiroticus | 81.5 ± 3.63 | 1.10 ± 0.3 | 15.93 ± 3.32 | 0.25 ± 0.13 | Greece | Present study | |
| Wild | Pelophylax ridibundus | 79.37 ± 0.50 ** | 1.37 ± 0.23 * | 18.52 ± 0.70 *** | 0.74 ± 0.15 ** | Turkey | [34] |
| Pelophylax ridibundus | 81.67 ± 0.11 ns | 0.82 ± 0.05 ** | 16.58 ± 0.14 ns | 0.89 ± 0.07 *** | Turkey | [50] | |
| Pelophylax ridibundus | 81.2 ± 2.12 ns | 1.26 ± 0.06 ns | 18.88 ± 0.48 *** | 0.82 ± 0.03 *** | Turkey | [33] | |
| Pelophylax esculentus | 79.72 ± 0.94 * | 0.56 ± 0.02 *** | 19.23 ± 0.15 *** | 0.68 ± 0.03 ** | Turkey | [37] | |
| Pelophylax esculentus | 79.47 ± 0.59 ** | 1.83 ± 0.16 *** | 22.21 ± 1.19 *** | 1.05 ± 0.34 *** | Turkey | [28] | |
| Pelophylax esculentus | 78.83 ± 0.48 *** | 0.85 ± 0.02 ** | 18.83 ± 0.48 *** | 1.04 ± 0.00 *** | Turkey | [51] | |
| Lithobates catesbeianus | 80.77 ± 0.74 ns | 0.63 ± 0.04 *** | 17.01 ± 0.12 ns | 0.61 ± 0.07 ** | Mexico | [52] | |
| Lithobates catesbeianus | 80.02 ± 0.13 * | 0.58 ± 0.06 *** | 19.31 ± 0.06 *** | 0.71 ± 0.05 ** | Thailand | [36] |
| Species | Moisture | Ash | Protein | Fat | Origin | Reference |
|---|---|---|---|---|---|---|
| Pelophylax epeiroticus | 81.5 ± 3.63 | 1.10 ± 0.3 | 15.93 ± 3.32 | 0.25 ± 0.13 | Greece | Present study |
| Salmo trutta fario | 78.3 *** | 1.16 ± 0.03 ns | 17.36 ± 0.37 * | 2.71 ± 0.21 *** | Turkey | [53] |
| Salvelinus fontinalis | 76.54 *** | 1.89 ± 0.04 *** | 16.31 ± 0.09 ns | 2.33 ± 0.02 *** | Turkey | [54] |
| Oreochromis niloticus | 81.2 ± 1.0 ns | 0.5 ± 0.2 *** | 16.4 ± 0.60 ns | 2.00 ± 0.60 *** | Poland | [55] |
| Cyprinus carpio | 77.7 ± 2.6 *** | 0.6 ± 0.3 *** | 16.7 ± 0.80 ns | 5.1 ± 3.00 *** | Poland | [55] |
| Cyprinus carpio | 73.73 ± 0.24 *** | 0.88 ± 0.05 * | 16.69 ± 0.40 ns | 7.13 ± 0.10 *** | Serbia | [56] |
| Oncorhynchus mykiss | 73.00 ± 1.5 *** | 0.80 ± 0.2 ** | 18.90 ± 0.80 *** | 7.4 ± 1.60 *** | Poland | [55] |
| Liza abu | 80.2 ns | 1.10 ns | 19.60 *** | 1.40 *** | Iraq | [57] |
| Aspius aspius | 78.51 ± 0.2 *** | 1.16 ± 0.05 ns | 18.07 ± 0.09 *** | 2.78 ± 0.11 *** | Serbia | [56] |
| Abramis brama | 78.66 ± 0.19 *** | 0.80 ± 0.04 ** | 17.59 ± 0.22 * | 3.24 ± 0.15 *** | Serbia | [56] |
| Barbus barbus | 72.39 ± 0.29 *** | 1.33 ± 0.03 * | 18.61 ± 0.37 *** | 7.78 ± 0.15 *** | Serbia | [56] |
| Acipenser ruthenus | 75.38 ± 0.35 *** | 0.93 ± 0.09 ns | 17.54 ± 0.23 * | 5.39 ± 0.14 *** | Serbia | [56] |
| Esox lucius | 79.00 ± 0.14 ** | 0.64 ± 0.03 *** | 18.43 ± 0.21 *** | 1.61 ± 0.01 *** | Serbia | [56] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatziioannou, M.; Kougiagka, E.; Karapanagiotidis, I.; Klaoudatos, D. Proximate Composition, Predictive Analysis and Allometric Relationships, of the Edible Water Frog (Pelophylax epeiroticus) in Lake Pamvotida (Northwest Greece). Sustainability 2022, 14, 3150. https://doi.org/10.3390/su14063150
Hatziioannou M, Kougiagka E, Karapanagiotidis I, Klaoudatos D. Proximate Composition, Predictive Analysis and Allometric Relationships, of the Edible Water Frog (Pelophylax epeiroticus) in Lake Pamvotida (Northwest Greece). Sustainability. 2022; 14(6):3150. https://doi.org/10.3390/su14063150
Chicago/Turabian StyleHatziioannou, Marianthi, Efkarpia Kougiagka, Ioannis Karapanagiotidis, and Dimitris Klaoudatos. 2022. "Proximate Composition, Predictive Analysis and Allometric Relationships, of the Edible Water Frog (Pelophylax epeiroticus) in Lake Pamvotida (Northwest Greece)" Sustainability 14, no. 6: 3150. https://doi.org/10.3390/su14063150