
Short-Term Effects of Reclamation of Aquaculture Ponds to Paddy Fields on Soil Chemical Properties and Bacterial Communities in Eastern China Coastal Zone

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Information and Sample Collection
2.2. Analysis of Soil Chemical Properties
2.3. DNA Extraction and PCR Amplification
2.4. Bioinformatics Analysis for Raw Sequences
2.5. Prediction of Bacterial Community Function
2.6. Statistical Analysis
3. Results
3.1. Soil Chemical Properties
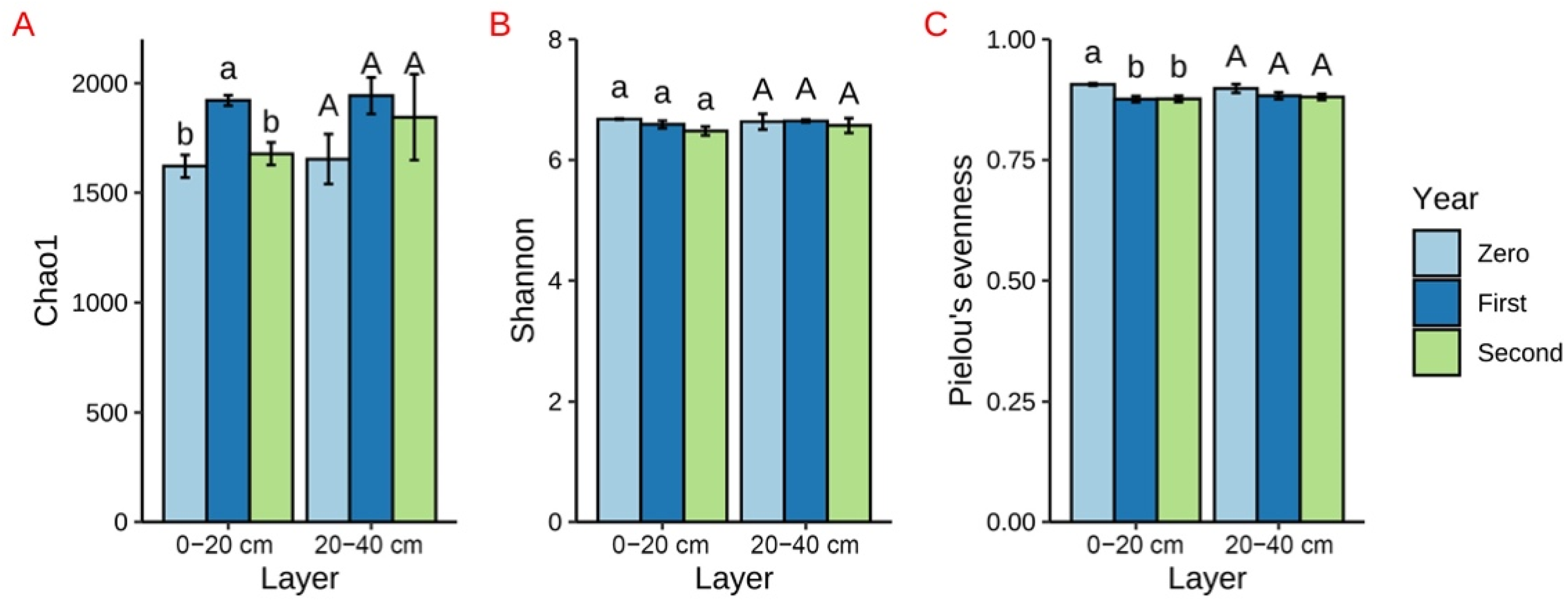
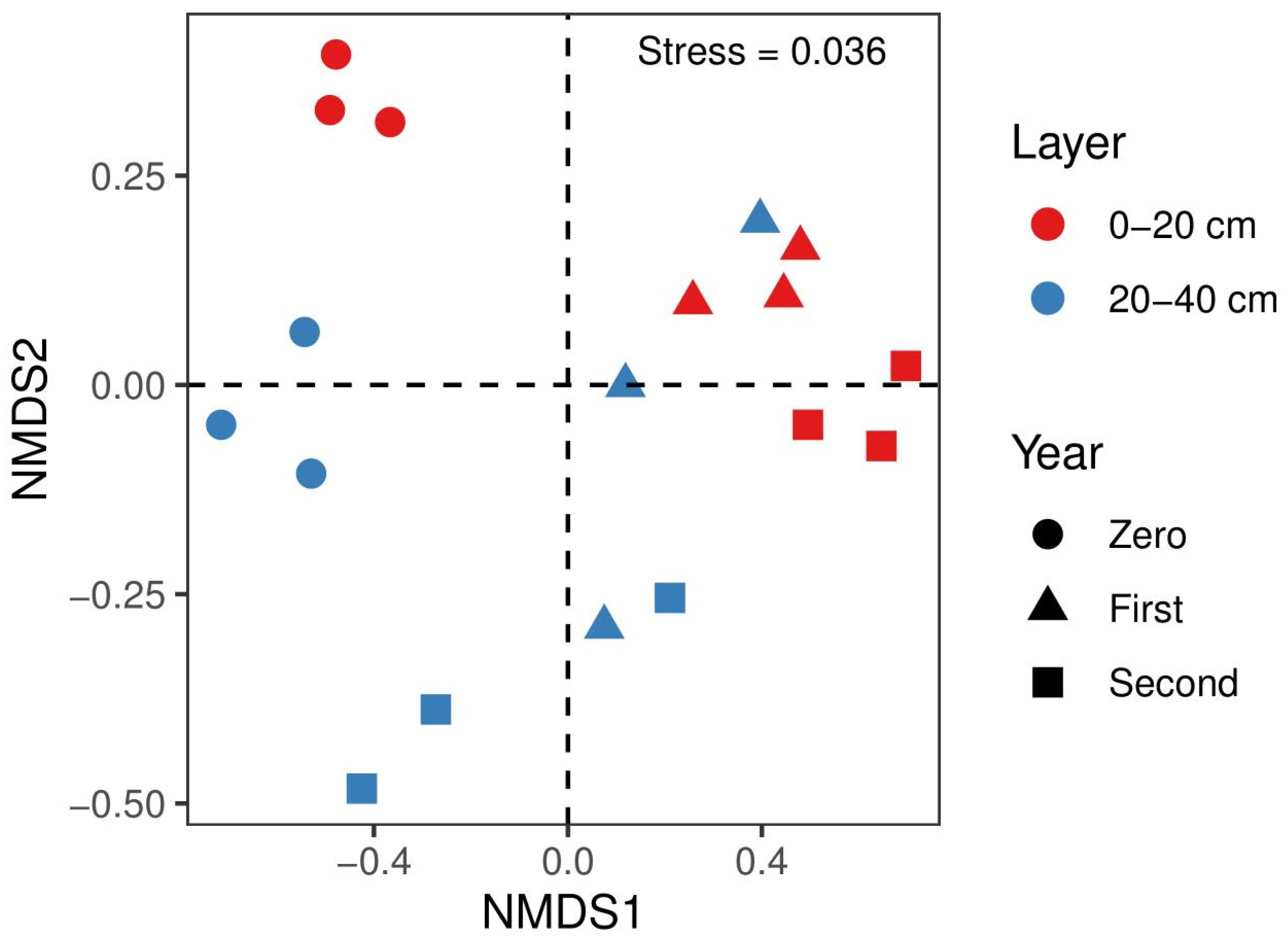
3.2. Diversity and Structure of Bacterial Communities
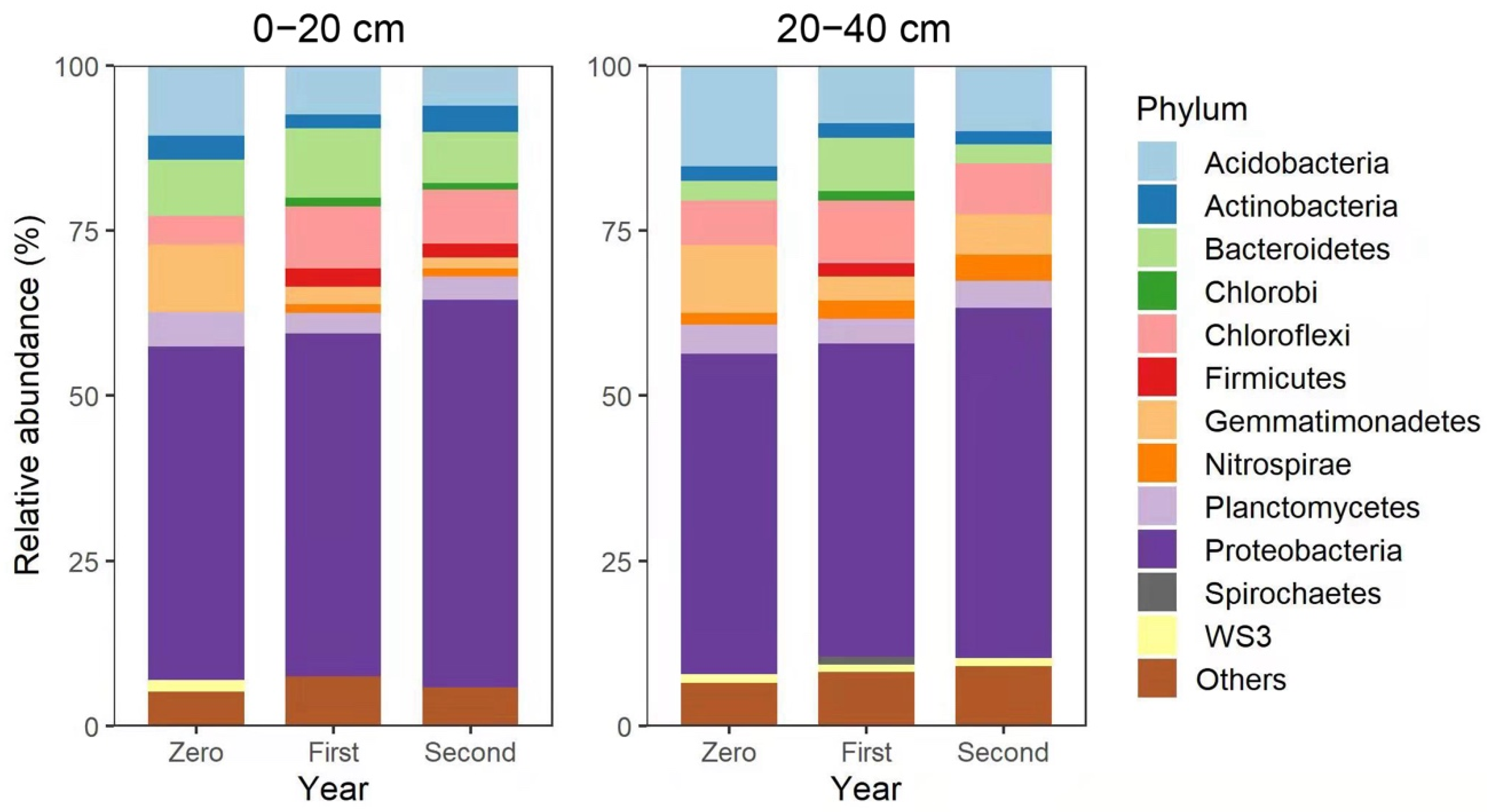
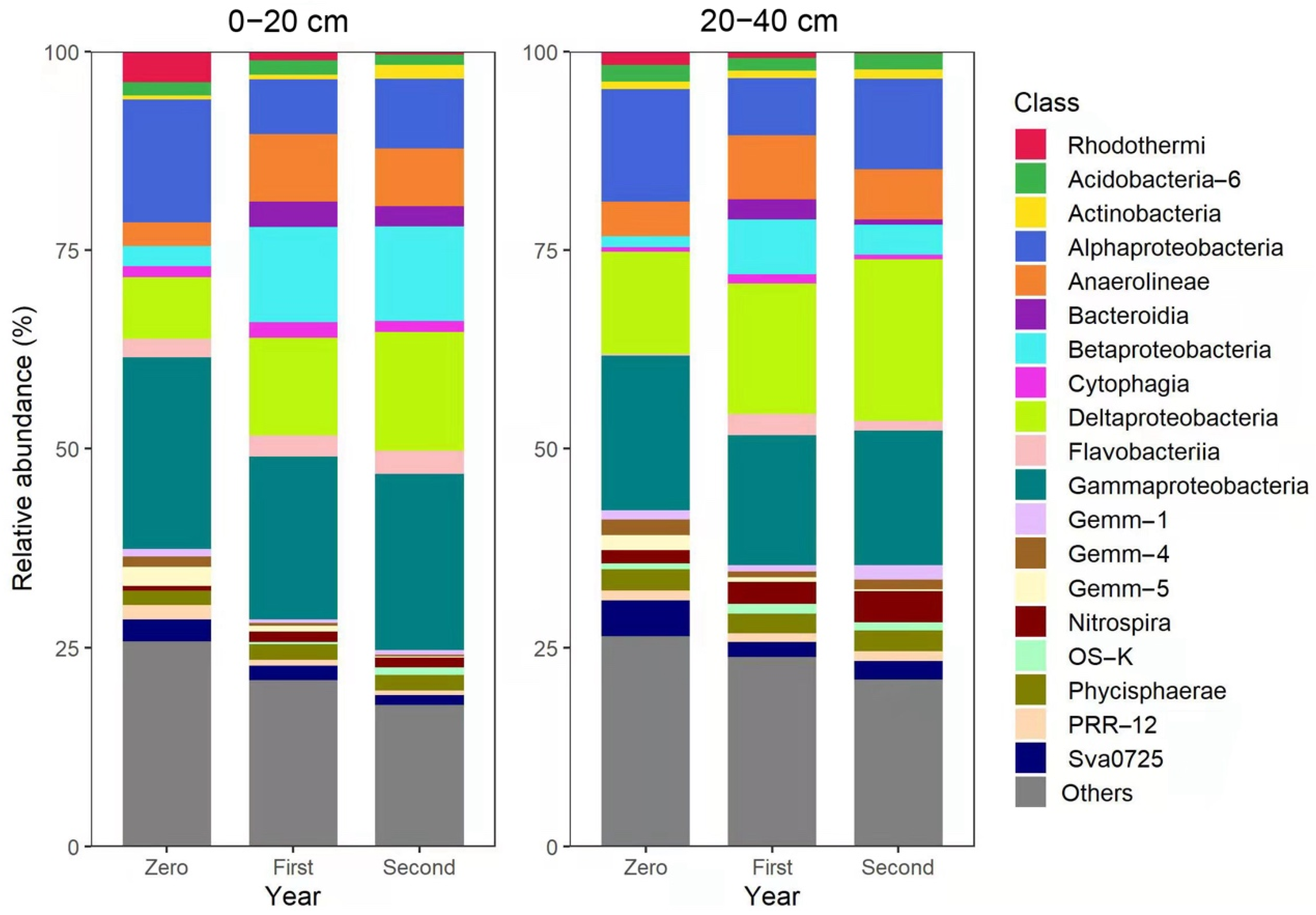
3.3. Composition of Bacterial Communities
3.4. Correlation between Bacterial Communities and Chemical Properties
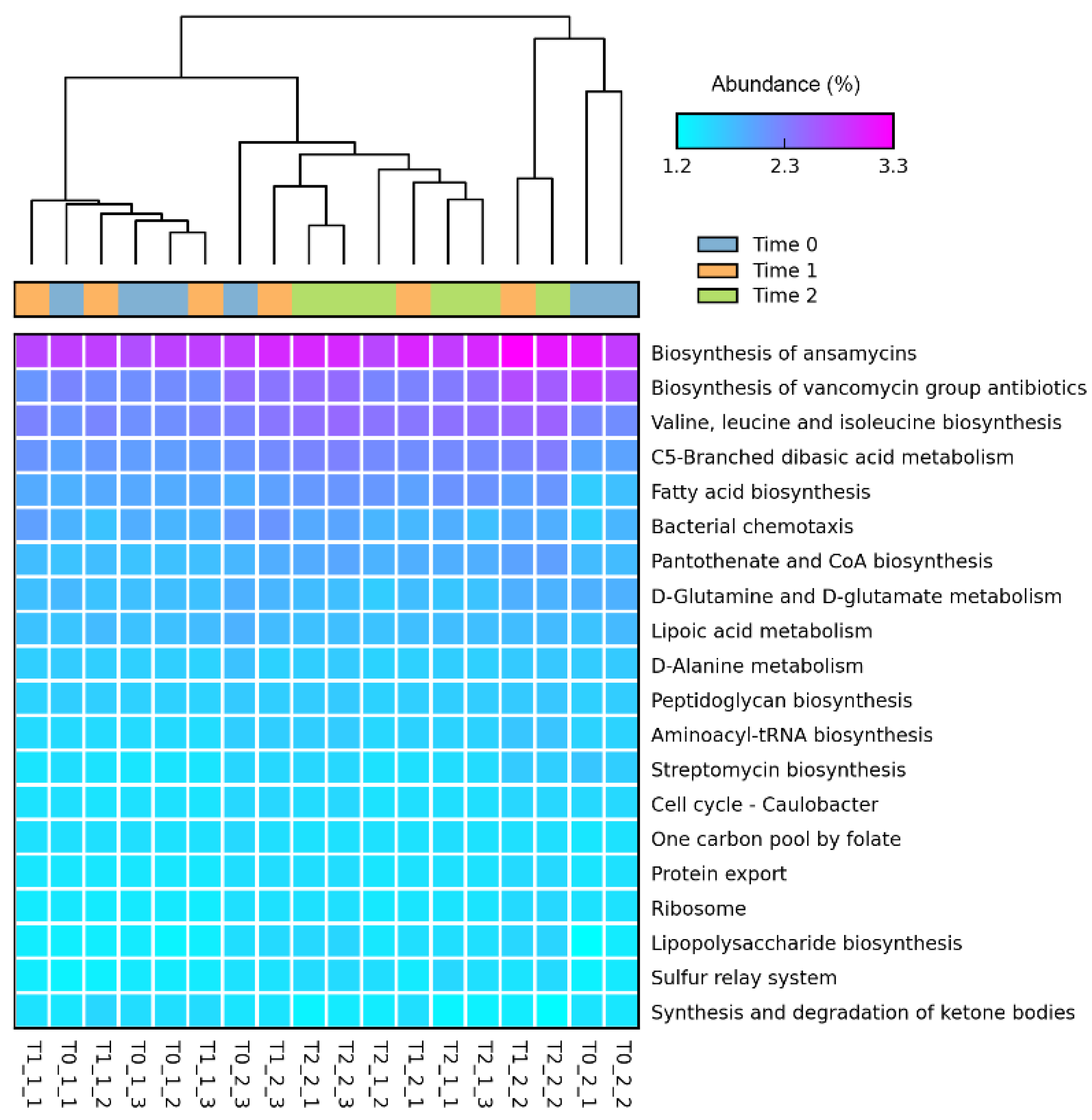
3.5. Bacterial Functional Genes
4. Discussion
4.1. Effect of Rice Cultivation in Reclaimed Lands on Soil Chemical Properties
4.2. Effect of Rice Cultivation and Its Duration in Reclaimed Lands on Soil Bacterial Community and Bacterial Functional Groups
4.3. Effects of Soil Chemical Properties on Soil Bacterial Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lei, N. Practice of reconstruction soils engineering in Shaanxi province. In IOP Conference Series: Earth and Environmental Science; IOP Publishing Ltd.: Bristol, UK, 2021; Volume 692, p. 042069. [Google Scholar] [CrossRef]
- Long, X.; Liu, L.; Shao, T.; Shao, H.; Liu, Z. Developing and sustainably utilize the coastal mudflat areas in China. Sci. Total Environ. 2016, 569–570, 1077–1086. [Google Scholar] [CrossRef]
- Wang, Z.; Zhuang, J.; Zhao, A.; Li, X. Types, harms and improvement of saline soil in Songnen Plain. In IOP Conference Series: Materials Science and Engineering; IOP Publishing Ltd.: Bristol, UK, 2018; Volume 322, p. 052059. [Google Scholar] [CrossRef]
- Vargas, R.; Pankovoy, E.I.; Balyuk, S.A.; Krasilnikov, P.V.; Hasanhanova, G.M. Handbook for Saline Soil Management: Eurasian Soil Partnership Implementation Plan; FAO: Rome, Italy, 2018. [Google Scholar]
- An, S.; Li, H.; Guan, B.; Zhou, C.; Wang, Z.; Deng, Z.; Zhi, Y.; Liu, Y.; Xu, C.; Fang, S.; et al. China’s natural wetlands: Past problems, current status, and future challenges. AMBIO J. Hum. Environ. 2007, 36, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.; Wang, Z.; Zhang, Y.; Zhang, B.; Chen, L.; Xi, Y.; Xiao, X.; Doughty, R.B.; Liu, M.; Jia, M.; et al. Rapid expansion of coastal aquaculture ponds in China from landsat observations during 1984–2016. Int. J. Appl. Earth Obs. Geoinf. 2019, 82, 101902. [Google Scholar] [CrossRef]
- Yang, H.Y.; Chen, B.; Barter, M.; Piersma, T.; Zhou, C.F.; Li, F.S.; Zhang, Z.W. Impacts of tidal land reclamation in Bohai Bay, China: Ongoing losses of critical Yellow Sea waterbird staging and wintering sites. Bird Conserv. Int. 2011, 21, 241–259. [Google Scholar] [CrossRef] [Green Version]
- Duan, H.; Zhang, H.; Huang, Q.; Zhang, Y.; Hu, M.; Niu, Y.; Zhu, J. Characterization and environmental impact analysis of sea land reclamation activities in China. Ocean Coast. Manag. 2016, 130, 128–137. [Google Scholar] [CrossRef]
- Zhang, X.; Liao, X.; Huang, L.; Shan, Q.; Hu, A.; Yan, D.; Zhang, J.; Long, X.E. Soil profile rather than reclamation time drives the mudflat soil microbial community in the wheat-maize rotation system of Nantong, China. J. Soils Sediments 2021, 21, 1672–1687. [Google Scholar] [CrossRef]
- Wan, S.; Liao, X.; Zhou, T.T.; Wu, Y.; Hu, A.; Yan, D.; Zhang, J.; Long, X.E. Shift in archaeal community along a soil profile in coastal wheat-maize rotation fields of different reclamation ages. Land Degrad. Dev. 2021, 32, 4162–4173. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, Update 2015: International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; FAO: Rome, Italy, 2015. [Google Scholar]
- Li, Y.; Zhang, J.; Chang, X.S.; Jiang, P.; Zhou, G.; Fu, S.; Yan, E.; Wu, J.; Lin, L. Long-term intensive management effects on soil organic carbon pools and chemical composition in Moso bamboo (Phyllostachys pubescens) forests in subtropical China. Forest Ecol. Mang. 2013, 303, 121–130. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; Circular (No. 939); USDA Press: Washington, DC, USA, 1954.
- Kachurina, O.M.; Zhang, H.; Raun, W.R.; Krenzer, E.G. Simultaneous determination of soil aluminum, ammonium- and nitrate-nitrogen using 1 M potassium chloride extraction. Commun. Soil Sci. Plan. 2000, 31, 893–903. [Google Scholar] [CrossRef]
- Ren, G.; Ren, W.; Teng, Y.; Li, Z. Evident bacterial community changes but only slight degradation when polluted with pyrene in a red soil. Front. Microbiol. 2015, 6, 22. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2018. [Google Scholar]
- Oksanen, J. Vegan: Ecological diversity. R Project 2018. Available online: https://cran.r-project.org/web/packages/vegan/vignettes/diversity-vegan.pdf (accessed on 23 January 2022).
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Pu, L.; Zhu, M.; Zhang, J.; Li, P.; Dai, X.; Xu, Y.; Liu, L. Evolution of soil properties following reclamation in coastal areas: A review. Geoderma 2014, 226–227, 130–139. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. Effects of salt stress on growth, mineral nutrition and proline accumulation in relation to osmotic adjustment in rice (Oryza Sativa L.) cultivars differing in salinity resistance. Plant Growth Regul. 1996, 19, 207–218. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.M.; Heuer, S.; Thomson, M.J.; Wissuwa, M. Genetic and genomic approaches to develop rice germplasm for problem soils. Plant Mol. Biol. 2007, 65, 547–570. [Google Scholar] [CrossRef]
- Singh, R.K.; Redoña, E.; Refuerzo, L. Varietal improvement for abiotic stress tolerance in crop plants: Special reference to salinity in rice. In Abiotic Stress Adaptation in Plants: Physiological, Molecular and Genomic Foundation; Pareek, A., Sopory, S.K., Bohnert, H.J., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 387–415. [Google Scholar]
- Xu, Y.; Pu, L.; Zhu, M.; Li, J.; Zhang, M.; Li, P.; Zhang, J. Spatial variation of soil salinity in the coastal reclamation area, Eastern China. J. Coast. Res. 2014, 30, 411–417. [Google Scholar] [CrossRef]
- Xu, Z.; Shao, T.; Lv, Z.; Yue, Y.; Liu, A.; Long, X.; Zhou, Z.; Gao, X.; Rengel, Z. The mechanisms of improving coastal saline soils by planting rice. Sci. Total Environ. 2020, 703, 135529. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Ding, N.; Liu, C.; Lin, Y.; Guo, B. Soil development under different cropping systems in a reclaimed coastal soil chronosequence. Geoderma 2014, 230–231, 50–57. [Google Scholar] [CrossRef]
- Zou, P.; Fu, J.; Cao, Z. Chronosequence of paddy soils and phosphorus sorption–desorption properties. J. Soils Sediments 2011, 11, 249–259. [Google Scholar] [CrossRef]
- Cui, J.; Liu, C.; Li, Z.; Wang, L.; Chen, X.; Ye, Z.; Fang, C. Long-term changes in topsoil chemical properties under centuries of cultivation after reclamation of coastal wetlands in the Yangtze Estuary, China. Soil Tillage Res. 2012, 123, 50–60. [Google Scholar] [CrossRef]
- Kalbitz, K.; Kaiser, K.; Fiedler, S.; Kölbl, A.; Amelung, W.; Bräuer, T.; Cao, Z.; Don, A.; Grootes, P.; Jahn, R.; et al. The carbon count of 2000 years of rice cultivation. Glob. Change Biol. 2013, 19, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Xia, L.; Zhu, Z.; Jiang, L.; Cheng, X.; An, S. Shift in soil organic carbon and nitrogen pools in different reclaimed lands following intensive coastal reclamation on the coasts of Eastern China. Sci. Rep. 2019, 9, 5921. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wu, P.; Yin, A.; Yang, X.; Zhang, X.; Zhang, M.; Gao, C. Organic carbon and total nitrogen dynamics of reclaimed soils following intensive agricultural use in Eastern China. Agric. Ecosyst. Environ. 2016, 235, 193–203. [Google Scholar] [CrossRef]
- Wang, X.; Yan, F.; Su, F. Changes in coastline and coastal reclamation in the three most developed areas of China, 1980–2018. Ocean Coast. Manag. 2021, 204, 105542. [Google Scholar] [CrossRef]
- Li, S.X.; Wang, Z.H.; Stewart, B.A. Chapter Five-responses of crop plants to ammonium and nitrate N. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2013; Volume 118, pp. 205–397. [Google Scholar] [CrossRef]
- Ye, L.; Peng, C.; Tang, B.; Wang, S.; Zhao, K.; Peng, Y. Determination effect of influent salinity and inhibition time on partial nitrification in a sequencing batch reactor treating saline sewage. Desalination 2009, 246, 556–566. [Google Scholar] [CrossRef]
- Zhu, H.; Yang, J.; Yao, R.; Wang, X.; Xie, W.; Zhu, W.; Liu, X.; Cao, Y.; Tao, J. Interactive effects of soil amendments (biochar and gypsum) and salinity on ammonia volatilization in coastal saline soil. Catena 2020, 190, 104527. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, R.; Shen, R. Coadaptation of plants to multiple stresses in acidic soils. Soil Sci. 2014, 179, 503–513. [Google Scholar] [CrossRef]
- Hu, A.; Zheng, M.; Sun, L.; Zhao, X.; Shen, R. Ammonium alleviates manganese toxicity and accumulation in rice by down-regulating the transporter gene OsNramp5 through rhizosphere acidification. Front. Plant Sci. 2019, 10, 1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierzynski, G.M. Methods of Phosphorus Analysis for Soils, Sediments, Residuals, and Waters; North Carolina State University: Raleigh, NC, USA, 2000. [Google Scholar]
- Cheng, Z.; Chen, Y.; Zhang, F. Effect of reclamation of abandoned salinized farmland on soil bacterial communities in arid Northwest China. Sci. Total Environ. 2018, 630, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-González, A.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K.; Fierer, N. A global atlas of the dominant bacteria found in soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, Q.; Chen, Y.; Dai, Q.; Hu, J. Dynamic change in enzyme activity and bacterial community with long-term rice cultivation in mudflats. Curr. Microbiol. 2019, 76, 361–369. [Google Scholar] [CrossRef]
- Zheng, W.; Xue, D.; Li, X.; Deng, Y.; Rui, J.; Feng, K.; Wang, Z. The responses and adaptations of microbial communities to salinity in farmland soils: A molecular ecological network analysis. Appl. Soil Ecol. 2017, 120, 239–246. [Google Scholar] [CrossRef]
- Sato, K.; Kato, Y.; Taguchi, G.; Nogawa, M.; Yokota, A.; Shimosaka, M. Chitiniphilus shinanonensis gen. nov., sp. nov., a novel chitin-degrading bacterium belonging to betaproteobacteria. J. Gen. Appl. Microbiol. 2009, 55, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Rocker, D.; Brinkhoff, T.; Grüner, N.; Dogs, M.; Simon, M. Composition of humic acid-degrading estuarine and marine bacterial communities. FEMS Microbiol. Ecol. 2012, 80, 45–63. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Li, J.; Ji, M.; Wu, Q.; Wu, X.; Ma, Y.; Sui, W.; Zhao, L.; Zhang, X. Non-synchronous structural and functional dynamics during the coalescence of two distinct soil bacterial communities. Front. Microbiol. 2019, 10, 1125. [Google Scholar] [CrossRef]
- Rosenfeld, J.S. Functional redundancy in ecology and conservation. Oikos 2002, 98, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Shi, P.; Zhang, Y.; Hu, Z.; Ma, K.; Wang, H.; Chai, T. The response of soil bacterial communities to mining subsidence in the west China aeolian sand area. Appl. Soil Ecol. 2017, 121, 1–10. [Google Scholar] [CrossRef]
- Lauber, C.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microb. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; Liu, W.; Long, X.E.; Jia, Y.; Wang, X.; Chen, Y. Different responses of soil bacterial communities to nitrogen addition in moss crust. Front. Microbiol. 2021, 12, 665975. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Ling, N.; Chen, D.; Guo, H.; Wei, J.; Bai, Y.; Shen, Q.; Hu, S. Differential responses of soil bacterial communities to long-term n and p inputs in a semi-arid steppe. Geoderma 2017, 292, 25–33. [Google Scholar] [CrossRef]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dynamics of gemmatimonadetes in soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wang, C.; Yu, W.; Turak, A.; Chen, D.; Huang, Y.; Ao, J.; Jiang, Y.; Huang, Z. Effects of nitrogen and phosphorus inputs on soil bacterial abundance, diversity, and community composition in Chinese fir plantations. Front. Microbiol. 2018, 9, 1543. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Layers/ Reclaimed Times | 0–20 cm | 20–40 cm | ||||
|---|---|---|---|---|---|---|
| Time 0 | Time 1 | Time 2 | Time 0 | Time 1 | Time 2 | |
| pH | 8.54 ± 0.01 a | 8.56 ± 0.04 a | 8.39 ± 0.02 b | 8.68 ± 0.02 B | 8.96 ± 0.04 A | 8.91 ± 0.01 A |
| EC (ds m−1) | 6.42 ± 0.27 a | 1.30 ± 0.03 b | 1.13 ± 0.08 b | 3.57 ± 0.03 A | 1.57 ± 0.04 B | 1.47 ± 0.02 B |
| TC (g kg−1) | 2.87 ± 0.15 b | 3.62 ± 0.05 a | 3.56 ± 0.19 a | 2.48 ± 0.08 C | 2.79 ± 0.03 B | 3.15 ± 0.02 A |
| TN (g kg−1) | 0.60 ± 0.02 a | 0.63 ± 0.01 a | 0.57 ± 0.02 a | 0.60 ± 0.00 A | 0.60 ± 0.03 A | 0.59 ± 0.01 A |
| WSOC (mg kg−1) | 46.74 ± 1.23 b | 66.65 ± 0.51 a | 71.12 ± 4.19 a | 24.97 ± 0.07 B | 29.87 ± 1.88 A | 32.87 ± 0.46 A |
| NH4+ (mg kg−1) | 22.52 ± 1.39 a | 15.28 ± 0.81 b | 10.27 ± 0.30 c | 29.82 ± 0.89 A | 17.31 ± 2.02 B | 12.44 ± 0.86 C |
| NO3– (mg kg−1) | 3.06 ± 0.07 a | 2.58 ± 0.02 b | 2.15 ± 0.02 c | 2.83 ± 0.03 A | 2.58 ± 0.05 B | 2.38 ± 0.02 C |
| AP (mg kg−1) | 10.34 ± 0.76 a | 9.17 ± 0.16 a | 7.18 ± 0.33 b | 6.52 ± 0.23 B | 8.41 ± 0.28 A | 8.67 ± 0.26 A |
| pH | EC | WSOC | AP | TC | TN | NH4+ | NO3− | ||
|---|---|---|---|---|---|---|---|---|---|
| Diversity indexes | Chao1 | 0.39 ** | −0.45 ** | −0.03 | 0.11 | 0.18 | 0.15 | −0.29 * | −0.22 |
| Shannon | 0.19 | 0.33 * | −0.29 * | 0.19 | −0.33 * | 0.13 | 0.34 * | 0.43 ** | |
| Pielou’s evenness | −0.05 | 0.75 ** | −0.35 ** | 0.16 | −0.54 ** | 0.02 | 0.62 ** | 0.68 ** | |
| Major phyla | Proteobacteria | −0.48 ** | −0.29 * | 0.60 ** | −0.04 | 0.58 ** | −0.16 | −0.55 ** | −0.57 ** |
| Acidobacteria | 0.24 | 0.49 ** | −0.72 ** | −0.28 * | −0.79 ** | 0.12 | 0.78 ** | 0.60 ** | |
| Gemmatimonadetes | 0.11 | 0.78 ** | −0.56 ** | 0.01 | −0.70 ** | 0.14 | 0.73 ** | 0.75 ** | |
| Bacteroidetes | −0.40 ** | 0.00 | 0.65 ** | 0.55 ** | 0.52 ** | 0.14 | −0.23 | 0.04 | |
| Chloroflexi | 0.23 | −0.80 ** | 0.16 | −0.24 | 0.37 ** | −0.05 | −0.44 ** | −0.56 ** | |
| Planctomycetes | 0.02 | 0.77 ** | −0.40 ** | 0.20 | −0.52 ** | −0.13 | 0.51 ** | 0.62 ** | |
| Nitrospirae | 0.66 ** | −0.42 ** | −0.46 ** | −0.22 | −0.11 | 0.03 | −0.32 * | −0.27 * | |
| Actinobacteria | −0.66 ** | 0.31 * | 0.42 ** | 0.06 | 0.26 | −0.29 * | −0.12 | 0.02 | |
| Firmicutes | −0.14 | −0.67 ** | 0.58 ** | 0.02 | 0.61 ** | −0.12 | −0.51 ** | −0.57 ** | |
| WS3 | 0.20 | 0.71 ** | −0.50 ** | 0.23 | −0.55 ** | 0.10 | 0.51 ** | 0.66 ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, A.; Lv, M.; Jiang, S.; Xu, L.; Guo, S.; Zhao, N.; Zheng, M.; Liu, B.; Li, J. Short-Term Effects of Reclamation of Aquaculture Ponds to Paddy Fields on Soil Chemical Properties and Bacterial Communities in Eastern China Coastal Zone. Sustainability 2022, 14, 1613. https://doi.org/10.3390/su14031613
Hu A, Lv M, Jiang S, Xu L, Guo S, Zhao N, Zheng M, Liu B, Li J. Short-Term Effects of Reclamation of Aquaculture Ponds to Paddy Fields on Soil Chemical Properties and Bacterial Communities in Eastern China Coastal Zone. Sustainability. 2022; 14(3):1613. https://doi.org/10.3390/su14031613
Chicago/Turabian StyleHu, Anyong, Min Lv, Shumiao Jiang, Lingyi Xu, Shiyang Guo, Na Zhao, Manman Zheng, Bo Liu, and Jinbiao Li. 2022. "Short-Term Effects of Reclamation of Aquaculture Ponds to Paddy Fields on Soil Chemical Properties and Bacterial Communities in Eastern China Coastal Zone" Sustainability 14, no. 3: 1613. https://doi.org/10.3390/su14031613