


Phytoextracts as Crop Biostimulants and Natural Protective Agents—A Critical Review

 , ,
, ,  , ,
, ,  ,
, 
Abstract
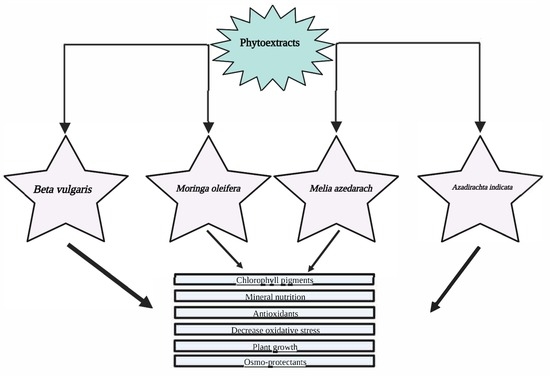
:
1. Introduction
2. Phytoextrants
2.1. Beta vulgaris—Source of Glycinebetaine
2.2. Moringa oleifera—Source of Vitamins and Nutrients
2.3. Citrus sinensis—Source of Ascorbic Acid
2.4. Melia azedarach—Source of Terpenoids
2.5. Azadirachta indica—Source of Secondary Metabolites
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Lemaire, G.; Belanger, G. Allometries in plants as drivers of forage nutritive value: A review. Agriculture 2020, 10, 5. [Google Scholar] [CrossRef] [Green Version]
- Merrington, G. The Good, the Bad and the Ugly: Copper and Arsenic in Soils. In Soil Health: The Foundation of Sustainable Agriculture; Wollongbar Agricultural Institute: Wollongbar, Australia, 2018. [Google Scholar]
- Mahmood, N.; Arshad, M.; Kaechele, H.; Shahzad, M.F.; Ullah, A.; Mueller, K. Fatalism, Climate Resiliency Training and Farmers’ Adaptation Responses: Implications for Sustainable Rainfed-Wheat Production in Pakistan. Sustainability 2020, 12, 1650. [Google Scholar] [CrossRef] [Green Version]
- Sarraf, M.; Vishwakarma, K.; Kumar, V.; Arif, N.; Das, S.; Johnson, R.; Janeeshma, E.; Puthur, J.T.; Aliniaeifard, S.; Chauhan, D.K.; et al. Metal/Metalloid-Based Nanomaterials for Plant Abiotic Stress Tolerance: An Overview of the Mechanisms. Plants 2022, 11, 316. [Google Scholar] [CrossRef] [PubMed]
- Jeyabharathi, S.; Kalishwaralal, K.; Sundar, K.; Muthukumaran, A. Synthesis of zinc oxide nanoparticles (ZnONPs) by aqueous extract of Amaranthus caudatus and evaluation of their toxicity and antimicrobial activity. Mater. Lett. 2017, 209, 295–298. [Google Scholar] [CrossRef]
- Ben Mrid, R.; Benmrid, B.; Hafsa, J.; Boukcim, H.; Sobeh, M.; Yasri, A. Secondary metabolites as biostimulant and bioprotectant agents: A review. Sci. Total Environ. 2021, 777, 146204. [Google Scholar] [CrossRef]
- Iqbal, S.; Khan, H.Z.; Shaheen, H. Growth and yield responses of mungbean (Vigna radiata L.) to different levels of phosphorus application under different tillage systems. Int. J. Agric. Appl. Sci. 2016, 4, 22–27. [Google Scholar]
- Ghani, M.A.; Abbas, M.M.; Ali, B.; Aziz, R.; Qadri, R.W.K.; Noor, A.; Azam, M.; Bahzad, S.; Saleem, M.H.; Abualreesh, M.H.; et al. Alleviating Role of Gibberellic Acid in Enhancing Plant Growth and Stimulating Phenolic Compounds in Carrot (Daucus carota L.) under Lead Stress. Sustainability 2021, 13, 12329. [Google Scholar] [CrossRef]
- Saleem, M.H.; Rizwan, M.; Shah, Z.-U.H.; Depar, N.; Usman, K. Chromium toxicity in plants: Consequences on growth, chromosomal behaviour and mineral nutrient status. Turk. J. Agric. For. 2022, 46, 371–389. [Google Scholar] [CrossRef]
- Mfarrej, M.; Wang, X.; Hamzah Saleem, M.; Hussain, I.; Rasheed, R.; Arslan Ashraf, M.; Iqbal, M.; Sohaib Chattha, M.; Nasser Alyemeni, M. Hydrogen sulphide and nitric oxide mitigate the negative impacts of waterlogging stress on wheat (Triticum aestivum L.). Plant Biol. 2021, 24, 670–683. [Google Scholar] [CrossRef]
- Ma, J.; Saleem, M.H.; Ali, B.; Rasheed, R.; Ashraf, M.A.; Aziz, H.; Ercisli, S.; Riaz, S.; Elsharkawy, M.M.; Hussain, I.; et al. Impact of foliar application of syringic acid on tomato (Solanum lycopersicum L.) under heavy metal stress-insights into nutrient uptake, redox homeostasis, oxidative stress, and antioxidant defense. Front. Plant Sci. 2022, 13, 950120. [Google Scholar] [CrossRef]
- Kour, J.; Kohli, S.K.; Khanna, K.; Bakshi, P.; Sharma, P.; Singh, A.D.; Ibrahim, M.; Devi, K.; Sharma, N.; Ohri, P.; et al. Brassinosteroid Signaling, Crosstalk and, Physiological Functions in Plants Under Heavy Metal Stress. Front. Plant Sci. 2021, 12, 608061. [Google Scholar] [CrossRef] [PubMed]
- Javed, M.T.; Saleem, M.H.; Aslam, S.; Rehman, M.; Iqbal, N.; Begum, R.; Ali, S.; Alsahli, A.A.; Alyemeni, M.N.; Wijaya, L. Elucidating silicon-mediated distinct morpho-physio-biochemical attributes and organic acid exudation patterns of cadmium stressed Ajwain (Trachyspermum ammi L.). Plant Physiol. Biochem. 2020, 157, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Saini, S.; Kaur, N.; Pati, P.K. Phytohormones: Key players in the modulation of heavy metal stress tolerance in plants. Ecotoxicol. Environ. Saf. 2021, 223, 112578. [Google Scholar] [CrossRef] [PubMed]
- Ghafar, M.A.; Akram, N.A.; Saleem, M.H.; Wang, J.; Wijaya, L.; Alyemeni, M.N. Ecotypic Morphological and Physio-Biochemical Responses of Two Differentially Adapted Forage Grasses, Cenchrus ciliaris L. and Cyperus arenarius Retz. to Drought Stress. Sustainability 2021, 13, 8069. [Google Scholar] [CrossRef]
- Kaya, C.; Şenbayram, M.; Akram, N.A.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Sulfur-enriched leonardite and humic acid soil amendments enhance tolerance to drought and phosphorus deficiency stress in maize (Zea mays L.). Sci. Rep. 2020, 10, 6432. [Google Scholar] [CrossRef] [Green Version]
- Sarwar, S.; Akram, N.A.; Saleem, M.H.; Zafar, S.; Alghanem, S.M.; Abualreesh, M.H.; Alatawi, A.; Ali, S. Spatial variations in the biochemical potential of okra [Abelmoschus esculentus L. (Moench)] leaf and fruit under field conditions. PLoS ONE 2022, 17, e0259520. [Google Scholar] [CrossRef]
- Sadiq, M.; Akram, N.A.; Ashraf, M.; Al-Qurainy, F.; Ahmad, P. Alpha-tocopherol-induced regulation of growth and metabolism in plants under non-stress and stress conditions. J. Plant Growth Regul. 2019, 38, 1325–1340. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M.K. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: A review. Ecotoxicol. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef]
- Rehman, M.; Saleem, M.H.; Fahad, S.; Bashir, S.; Peng, D.; Deng, G.; Alamri, S.; Siddiqui, M.H.; Khan, S.M.; Shah, R.A. Effects of rice straw biochar and nitrogen fertilizer on ramie (Boehmeria nivea L.) morpho-physiological traits, copper uptake and post-harvest soil characteristics, grown in an aged-copper contaminated soil. J. Plant Nutr. 2021, 45, 11–24. [Google Scholar] [CrossRef]
- Ahmad, K.; Aslam, M.; Saleem, M.H.; Ijaz, M.; Ul-Allah, S.; El-Sheikh, A.H.M.A.; Adnan, M.; Ali, S. Genetic Diversity and Characterization of Salt Stress Tolerance Traits in Maize (Zea mays L.) Under Normal And Saline Conditions. Pak. J. Bot. 2022, 54, 759–769. [Google Scholar] [CrossRef]
- Zaheer, I.E.; Ali, S.; Saleem, M.H.; Arslan Ashraf, M.; Ali, Q.; Abbas, Z.; Rizwan, M.; El-Sheikh, M.A.; Alyemeni, M.N.; Wijaya, L. Zinc-lysine Supplementation Mitigates Oxidative Stress in Rapeseed (Brassica napus L.) by Preventing Phytotoxicity of Chromium, When Irrigated with Tannery Wastewater. Plants 2020, 9, 1145. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Wang, X.; Haroon, U.; Chaudhary, H.J.; Kamal, A.; Ali, Q.; Saleem, M.H.; Usman, K.; Alatawi, A.; Ali, S.; et al. Antifungal activity of Zinc nitrate derived nano Zno fungicide synthesized from Trachyspermum ammi to control fruit rot disease of grapefruit. Ecotoxicol. Environ. Saf. 2022, 233, 113311. [Google Scholar] [CrossRef] [PubMed]
- Latique, S.; Mrid, R.B.; Kabach, I.; Kchikich, A.; Sammama, H.; Yasri, A.; Nhiri, M.; El Kaoua, M.; Douira, A.; Selmaoui, K. Foliar Application of Ulva rigida Water Extracts Improves Salinity Tolerance in Wheat (Triticum durum L.). Agronomy 2021, 11, 265. [Google Scholar] [CrossRef]
- Ali, Q.; Ahmar, S.; Sohail, M.A.; Kamran, M.; Ali, M.; Saleem, M.H.; Rizwan, M.; Ahmed, A.M.; Mora-Poblete, F.; do Amaral Júnior, A.T. Research advances and applications of biosensing technology for the diagnosis of pathogens in sustainable agriculture. Environ. Sci. Pollut. Res. 2021, 28, 9002–9019. [Google Scholar] [CrossRef] [PubMed]
- Sheoran, V.; Sheoran, A.S.; Poonia, P. Factors affecting phytoextraction: A review. Pedosphere 2016, 26, 148–166. [Google Scholar] [CrossRef]
- Bouchmaa, N.; Mrid, R.B.; Kabach, I.; Zouaoui, Z.; Karrouchi, K.; Chtibi, H.; Zyad, A.; Cacciola, F.; Nhiri, M. Beta vulgaris subsp. maritima: A Valuable Food with High Added Health Benefits. Appl. Sci. 2022, 12, 1866. [Google Scholar] [CrossRef]
- Yolcu, S.; Alavilli, H.; Ganesh, P.; Asif, M.; Kumar, M.; Song, K. An Insight into the Abiotic Stress Responses of Cultivated Beets (Beta vulgaris L.). Plants 2022, 11, 12. [Google Scholar] [CrossRef]
- Nazar, Z.; Akram, N.A.; Saleem, M.H.; Ashraf, M.; Ahmed, S.; Ali, S.; Abdullah Alsahli, A.; Alyemeni, M.N. Glycinebetaine-Induced Alteration in Gaseous Exchange Capacity and Osmoprotective Phenomena in Safflower (Carthamus tinctorius L.) under Water Deficit Conditions. Sustainability 2020, 12, 10649. [Google Scholar] [CrossRef]
- Gou, W.; Tian, L.; Ruan, Z.; Zheng, P.; Chen, F.; Zhang, L.; Cui, Z.; Zheng, P.; Li, Z.; Gao, M. Accumulation of choline and glycinebetaine and drought stress tolerance induced in maize (Zea mays) by three plant growth promoting rhizobacteria (PGPR) strains. Pak. J. Bot. 2015, 47, 581–586. [Google Scholar]
- Yolcu, S.; Alavilli, H.; Ganesh, P.; Panigrahy, M.; Song, K. Salt and drought stress responses in cultivated beets (Beta vulgaris L.) and wild beet (Beta maritima L.). Plants 2021, 10, 1843. [Google Scholar] [CrossRef]
- Romano, A.; Sorgona, A.; Lupini, A.; Araniti, F.; Stevanato, P.; Cacco, G.; Abenavoli, M.R. Morpho-physiological responses of sugar beet (Beta vulgaris L.) genotypes to drought stress. Acta Physiol. Plant. 2013, 35, 853–865. [Google Scholar] [CrossRef]
- Moosavi, S.G.R.; Ramazani, S.H.R.; Hemayati, S.S.; Gholizade, H. Effect of drought stress on root yield and some morpho-physiological traits in different genotypes of sugar beet (Beta vulgaris L.). J. Crop Sci. Biotechnol. 2017, 20, 167–174. [Google Scholar] [CrossRef]
- Mäck, G.; Hoffmann, C.M.; Märländer, B. Nitrogen compounds in organs of two sugar beet genotypes (Beta vulgaris L.) during the season. Field Crops Res. 2007, 102, 210–218. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.; Ashraf, M.; Foolad, M. Improving plant abiotic-stress resistance by exogenous application of osmoprotectants glycine, betaine and proline. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Ali, S.; Chaudhary, A.; Rizwan, M.; Anwar, H.T.; Adrees, M.; Farid, M.; Irshad, M.K.; Hayat, T.; Anjum, S.A. Alleviation of chromium toxicity by glycinebetaine is related to elevated antioxidant enzymes and suppressed chromium uptake and oxidative stress in wheat (Triticum aestivum L.). Environ. Sci. Pollut. Res. 2015, 22, 10669–10678. [Google Scholar] [CrossRef]
- You, L.; Song, Q.; Wu, Y.; Li, S.; Jiang, C.; Chang, L.; Yang, X.; Zhang, J. Accumulation of glycine betaine in transplastomic potato plants expressing choline oxidase confers improved drought tolerance. Planta 2019, 249, 1963–1975. [Google Scholar] [CrossRef]
- Maqsood, A.; Shahbaz, M.; Akram, N.A. Influence of Exogenously Applied Glycinebetaine on Growth and Gas Exchange Characteristics of Maize (Zea mays L.). Pak. J. Agric. Sci. 2006, 43, 36–41. [Google Scholar]
- Wang, G.; Li, F.; Zhang, J.; Zhao, M.; Hui, Z.; Wang, W. Overaccumulation of glycine betaine enhances tolerance of the photosynthetic apparatus to drought and heat stress in wheat. Photosynthetica 2010, 48, 30–41. [Google Scholar] [CrossRef]
- Ma, X.; Wang, Y.; Xie, S.; Wang, C.; Wang, W. Glycinebetaine application ameliorates negative effects of drought stress in tobacco. Russ. J. Plant Physiol. 2007, 54, 472. [Google Scholar] [CrossRef]
- Habib, N.; Ashraf, M.; Ali, Q.; Perveen, R. Response of salt stressed okra (Abelmoschus esculentus Moench) plants to foliar-applied glycine betaine and glycine betaine containing sugarbeet extract. S. Afr. J. Bot. 2012, 83, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Raza, S.H.; Athar, H.R.; Ashraf, M.; Hameed, A. Glycinebetaine-induced modulation of antioxidant enzymes activities and ion accumulation in two wheat cultivars differing in salt tolerance. Environ. Exp. Bot. 2007, 60, 368–376. [Google Scholar] [CrossRef]
- Iqbal, N.; Ashraf, M.; Ashraf, M. Glycinebetaine, an osmolyte of interest to improve water stress tolerance in sunflower (Helianthus annuus L.): Water relations and yield. S. Afr. J. Bot. 2008, 74, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Ali, Q.; Ashraf, M. Induction of drought tolerance in maize (Zea mays L.) due to exogenous application of trehalose: Growth, photosynthesis, water relations and oxidative defence mechanism. J. Agron. Crop Sci. 2011, 197, 258–271. [Google Scholar] [CrossRef]
- Shahbaz, M.; Masood, Y.; Perveen, S.; Ashraf, M. Is foliar-applied glycinebetaine effective in mitigating the adverse effects of drought stress on wheat (Triticum aestivum L.)? J. Appl. Bot. Food Qual. 2012, 84, 192. [Google Scholar]
- Manaf, H.H. Beneficial effects of exogenous selenium, glycine betaine and seaweed extract on salt stressed cowpea plant. Ann. Agric. Sci. 2016, 61, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Tisarum, R.; Theerawitaya, C.; Samphumphung, T.; Takabe, T.; Cha-um, S. Exogenous foliar application of glycine betaine to alleviate water deficit tolerance in two Indica rice genotypes under greenhouse conditions. Agronomy 2019, 9, 138. [Google Scholar] [CrossRef] [Green Version]
- Cha-um, S.; Samphumphuang, T.; Kirdmanee, C. Glycinebetaine alleviates water deficit stress in indica rice using proline accumulation, photosynthetic efficiencies, growth performances and yield attributes. Aust. J. Crop Sci. 2013, 7, 213–218. [Google Scholar]
- Khan, S.U.; Khan, A.; Naveed, S. Effect of exogenously applied kinetin and glycinebetaine on metabolic and yield attributes of rice (Oryza sativa L.) under drought stress. Emir. J. Food Agric. 2015, 27, 75–81. [Google Scholar]
- Cruz, F.; Castro, G.; Silva Júnior, D.; Festucci-Buselli, R.; Pinheiro, H. Exogenous glycine betaine modulates ascorbate peroxidase and catalase activities and prevent lipid peroxidation in mild water-stressed Carapa guianensis plants. Photosynthetica 2013, 51, 102–108. [Google Scholar] [CrossRef]
- Gupta, N.; Thind, S.K. Grain yield response of drought stressed wheat to foliar application of glycine betaine. Indian J. Agric. Res. 2017, 51, 287–291. [Google Scholar]
- Osman, H.S. Enhancing antioxidant–yield relationship of pea plant under drought at different growth stages by exogenously applied glycine betaine and proline. Ann. Agric. Sci. 2015, 60, 389–402. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, A.; Valverde, R.; Chen, T.H.; Murata, N. Transformation of Arabidopsis with the codA gene for choline oxidase enhances freezing tolerance of plants. Plant J. 2000, 22, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Sulpice, R.; Tsukaya, H.; Nonaka, H.; Mustardy, L.; Chen, T.H.; Murata, N. Enhanced formation of flowers in salt-stressed Arabidopsis after genetic engineering of the synthesis of glycine betaine. Plant J. 2003, 36, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.-G.; Du, B.-X.; Zhang, W.-K.; Zhang, J.-S.; Chen, S.-Y. AhCMO, regulated by stresses in Atriplex hortensis, can improve drought tolerance in transgenic tobacco. Theor. Appl. Genet. 2002, 105, 815–821. [Google Scholar] [CrossRef]
- Park, E.J.; Jeknić, Z.; Sakamoto, A.; DeNoma, J.; Yuwansiri, R.; Murata, N.; Chen, T.H. Genetic engineering of glycinebetaine synthesis in tomato protects seeds, plants, and flowers from chilling damage. Plant J. 2004, 40, 474–487. [Google Scholar] [CrossRef]
- Goel, D.; Singh, A.K.; Yadav, V.; Babbar, S.B.; Murata, N.; Bansal, K.C. Transformation of tomato with a bacterial codA gene enhances tolerance to salt and water stresses. J. Plant Physiol. 2011, 168, 1286–1294. [Google Scholar] [CrossRef]
- Quan, R.; Shang, M.; Zhang, H.; Zhao, Y.; Zhang, J. Improved chilling tolerance by transformation with betA gene for the enhancement of glycinebetaine synthesis in maize. Plant Sci. 2004, 166, 141–149. [Google Scholar] [CrossRef]
- Su, J.; Hirji, R.; Zhang, L.; He, C.; Selvaraj, G.; Wu, R. Evaluation of the stress-inducible production of choline oxidase in transgenic rice as a strategy for producing the stress-protectant glycine betaine. J. Exp. Bot. 2006, 57, 1129–1135. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, R.; Kim, M.D.; Back, K.-H.; Kim, H.-S.; Lee, H.-S.; Kwon, S.-Y.; Murata, N.; Chung, W.-I.; Kwak, S.-S. Stress-induced expression of choline oxidase in potato plant chloroplasts confers enhanced tolerance to oxidative, salt, and drought stresses. Plant Cell Rep. 2008, 27, 687–698. [Google Scholar] [CrossRef]
- Kathuria, H.; Giri, J.; Nataraja, K.N.; Murata, N.; Udayakumar, M.; Tyagi, A.K. Glycinebetaine-induced water-stress tolerance in codA-expressing transgenic indica rice is associated with up-regulation of several stress responsive genes. Plant Biotechnol. J. 2009, 7, 512–526. [Google Scholar] [CrossRef]
- Li, D.; Zhang, T.; Wang, M.; Liu, Y.; Brestic, M.; Chen, T.H.; Yang, X. Genetic engineering of the biosynthesis of glycine betaine modulates phosphate homeostasis by regulating phosphate acquisition in tomato. Front. Plant Sci. 2019, 9, 1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, D.; Zhang, W.; Wang, C.; Meng, Q.; Li, G.; Chen, T.H.; Yang, X. Genetic engineering of the biosynthesis of glycinebetaine leads to alleviate salt-induced potassium efflux and enhances salt tolerance in tomato plants. Plant Sci. 2017, 257, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Tan, W.; Yang, X.-H.; Zhang, H.-X. Plastid-expressed choline monooxygenase gene improves salt and drought tolerance through accumulation of glycine betaine in tobacco. Plant Cell Rep. 2008, 27, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Salaheldeen, M.; Aroua, M.K.; Mariod, A.; Cheng, S.F.; Abdelrahman, M.A.; Atabani, A. Physicochemical characterization and thermal behavior of biodiesel and biodiesel–diesel blends derived from crude Moringa peregrina seed oil. Energy Convers. Manag. 2015, 92, 535–542. [Google Scholar] [CrossRef] [Green Version]
- Hussein, M.; Abou-Baker, N.H. Growth and mineral status of moringa plants as affected by silicate and salicylic acid under salt stress. Int. J. Plant Soil Sci. 2013, 3, 163–177. [Google Scholar] [CrossRef]
- Mahmood, K.T.; Mugal, T.; Haq, I.U. Moringa oleifera: A natural gift-A review. J. Pharm. Sci. Res. 2010, 2, 775. [Google Scholar]
- Kaya, C.; Ashraf, M.; Sonmez, O.; Tuna, A.L.; Polat, T.; Aydemir, S. Exogenous application of thiamin promotes growth and antioxidative defense system at initial phases of development in salt-stressed plants of two maize cultivars differing in salinity tolerance. Acta Physiol. Plant. 2015, 37, 1741. [Google Scholar] [CrossRef]
- Ejaz, B.; Sajid, Z.A.; Aftab, F. Effect of exogenous application of ascorbic acid on antioxidant enzyme activities, proline contents, and growth parameters of Saccharum spp. hybrid cv. HSF-240 under salt stress. Turk. J. Biol. 2012, 36, 630–640. [Google Scholar] [CrossRef]
- Malik, S.; Ashraf, M. Exogenous application of ascorbic acid stimulates growth and photosynthesis of wheat (Triticum aestivum L.) under drought. Soil Environ. 2012, 31, 72–77. [Google Scholar]
- Aziz, A.; Akram, N.A.; Ashraf, M. Influence of natural and synthetic vitamin C (ascorbic acid) on primary and secondary metabolites and associated metabolism in quinoa (Chenopodium quinoa Willd.) plants under water deficit regimes. Plant Physiol. Biochem. 2018, 123, 192–203. [Google Scholar] [CrossRef]
- Sayed, S.; Gadallah, M. Effects of shoot and root application of thiamin on salt-stressed sunflower plants. Plant Growth Regul. 2002, 36, 71–80. [Google Scholar] [CrossRef]
- GHAFFAR, A.; Akram, N.A.; Ashraf, M.; ASHRAF, Y.; Sadiq, M. Thiamin-induced variations in oxidative defense processes in white clover (Trifolium repens L.) under water deficit stress. Turk. J. Bot. 2019, 43, 58–66. [Google Scholar] [CrossRef]
- Semida, W.; Taha, R.; Abdelhamid, M.; Rady, M. Foliar-applied α-tocopherol enhances salt-tolerance in Vicia faba L. plants grown under saline conditions. S. Afr. J. Bot. 2014, 95, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Sadiq, M.; Akram, N.A.; Javed, M.T. Alpha-tocopherol alters endogenous oxidative defense system in mung bean plants under water-deficit conditions. Pak. J. Bot. 2016, 48, 2177–2182. [Google Scholar]
- El-Bassiouny, H.; Sadak, M.S. Impact of foliar application of ascorbic acid and α-tocopherol on antioxidant activity and some biochemical aspects of flax cultivars under salinity stress. Acta Biol. Colomb. 2015, 20, 209–222. [Google Scholar] [CrossRef]
- Gopalakrishnan, L.; Doriya, K.; Kumar, D.S. Moringa oleifera: A review on nutritive importance and its medicinal application. Food Sci. Hum. Wellness 2016, 5, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Stohs, S.J.; Hartman, M.J. Review of the safety and efficacy of Moringa oleifera. Phytother. Res. 2015, 29, 796–804. [Google Scholar] [CrossRef]
- Howladar, S.M. A novel Moringa oleifera leaf extract can mitigate the stress effects of salinity and cadmium in bean (Phaseolus vulgaris L.) plants. Ecotoxicol. Environ. Saf. 2014, 100, 69–75. [Google Scholar] [CrossRef]
- Kerdsomboon, K.; Tatip, S.; Kosasih, S.; Auesukaree, C. Soluble Moringa oleifera leaf extract reduces intracellular cadmium accumulation and oxidative stress in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2016, 121, 543–549. [Google Scholar] [CrossRef]
- Fahey, J.W. Moringa oleifera: A review of the medical evidence for its nutritional, therapeutic, and prophylactic properties. Part 1. Trees Life J. 2005, 1, 1–15. [Google Scholar]
- Khan, T.; Mazid, M.; Mohammad, F. A review of ascorbic acid potentialities against oxidative stress induced in plants. J. Agrobiol. 2011, 28, 97–111. [Google Scholar] [CrossRef]
- Sharma, R.; Bhardwaj, R.; Thukral, A.K.; Al-Huqail, A.A.; Siddiqui, M.H.; Ahmad, P. Oxidative stress mitigation and initiation of antioxidant and osmoprotectant responses mediated by ascorbic acid in Brassica juncea L. subjected to copper (II) stress. Ecotoxicol. Environ. Saf. 2019, 182, 109436. [Google Scholar] [CrossRef] [PubMed]
- Fatima, A.; Singh, A.A.; Mukherjee, A.; Agrawal, M.; Agrawal, S.B. Ascorbic acid and thiols as potential biomarkers of ozone tolerance in tropical wheat cultivars. Ecotoxicol. Environ. Saf. 2019, 171, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.A.; Javed, F.; Wahid, A.; Sadia, B. Alleviating effect of exogenous application of ascorbic acid on growth and mineral nutrients in cadmium stressed barley (Hordeum vulgare) seedlings. Int. J. Agric. Biol. 2016, 18, 73–79. [Google Scholar] [CrossRef]
- Ma, J.; Saleem, M.H.; Yasin, G.; Mumtaz, S.; Qureshi, F.F.; Ali, B.; Ercisli, S.; Alhag, S.K.; Ahmed, A.E.; Vodnar, D.C.; et al. Individual and combinatorial effects of SNP and NaHS on morpho-physio-biochemical attributes and phytoextraction of chromium through Cr-stressed spinach (Spinacia oleracea L.). Front. Plant Sci. 2022, 13, 973740. [Google Scholar] [CrossRef]
- Saleem, M.H.; Mfarrej, M.F.B.; Alatawi, A.; Mumtaz, S.; Imran, M.; Ashraf, M.A.; Rizwan, M.; Usman, K.; Ahmad, P.; Ali, S. Silicon Enhances Morpho–Physio–Biochemical Responses in Arsenic Stressed Spinach (Spinacia oleracea L.) by Minimizing Its Uptake. J. Plant Growth Regul. 2022. [Google Scholar] [CrossRef]
- Saleem, M.H.; Wang, X.; Ali, S.; Zafar, S.; Nawaz, M.; Adnan, M.; Fahad, S.; Shah, A.; Alyemeni, M.N.; Hefft, D.I.; et al. Interactive effects of gibberellic acid and NPK on morpho-physio-biochemical traits and organic acid exudation pattern in coriander (Coriandrum sativum L.) grown in soil artificially spiked with boron. Plant Physiol. Biochem. 2021, 167, 884–900. [Google Scholar] [CrossRef]
- Sajid, Z.A.; Aftab, F. Amelioration of salinity tolerance in Solanum tuberosum L. by exogenous application of ascorbic acid. In Vitro Cell. Dev. Biol.-Plant 2009, 45, 540–549. [Google Scholar] [CrossRef]
- Younis, M.E.; Hasaneen, M.N.; Kazamel, A.M. Exogenously applied ascorbic acid ameliorates detrimental effects of NaCl and mannitol stress in Vicia faba seedlings. Protoplasma 2010, 239, 39–48. [Google Scholar] [CrossRef]
- Aly, A.A.; Khafaga, A.F.; Omar, G.N. Improvement the adverse effect of salt stress in Egyptian clover (Trifolium alexandrinum L.) by AsA application through some biochemical and RT-PCR markers. J. Appl. Phytotechnol. Environ. Sanit. 2012, 1, 91–102. [Google Scholar]
- Kamran, M.; Danish, M.; Saleem, M.H.; Malik, Z.; Parveen, A.; Abbasi, G.H.; Jamil, M.; Ali, S.; Afzal, S.; Riaz, M. Application of abscisic acid and 6-benzylaminopurine modulated morpho-physiological and antioxidative defense responses of tomato (Solanum lycopersicum L.) by minimizing cobalt uptake. Chemosphere 2020, 263, 128169. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.; Wang, X.; Saleem, M.H.; Khan, M.H.U.; Afzal, J.; Fiaz, S.; Ali, S.; Ishaq, H.; Khan, A.H.; Rehman, N.; et al. Deciphering Plantago ovata Forsk Leaf Extract Mediated Distinct Germination, Growth and Physio-Biochemical Improvements under Water Stress in Maize (Zea mays L.) at Early Growth Stage. Agronomy 2021, 11, 1404. [Google Scholar] [CrossRef]
- Dolatabadian, A.; Jouneghani, R.S. Impact of exogenous ascorbic acid on antioxidant activity and some physiological traits of common bean subjected to salinity stress. Not. Bot. Horti Agrobot. Cluj-Napoca 2009, 37, 165–172. [Google Scholar]
- Hassan, A.; Amjad, S.F.; Saleem, M.H.; Yasmin, H.; Imran, M.; Riaz, M.; Ali, Q.; Joyia, F.A.; Ahmed, S.; Ali, S. Foliar application of ascorbic acid enhances salinity stress tolerance in barley (Hordeum vulgare L.) through modulation of morpho-physio-biochemical attributes, ions uptake, osmo-protectants and stress response genes expression. Saudi J. Biol. Sci. 2021, 28, 4276–4290. [Google Scholar] [CrossRef] [PubMed]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic acid-a potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front. Plant Sci. 2017, 8, 613. [Google Scholar] [CrossRef] [PubMed]
- Favela-Hernández, J.M.J.; González-Santiago, O.; Ramírez-Cabrera, M.A.; Esquivel-Ferriño, P.C.; Camacho-Corona, M.d.R. Chemistry and Pharmacology of Citrus sinensis. Molecules 2016, 21, 247. [Google Scholar] [CrossRef] [Green Version]
- Salem, M.; Abdel-Ghany, H.M. Effects of dietary orange peel on growth performance of Nile tilapia (Oreochromis niloticus) fingerlings. Aquac. Stud. 2018, 18, 127–134. [Google Scholar] [CrossRef]
- Fukui, K.; Kaneuji, A.; Hirata, H.; Tsujioka, J.-I.; Shioya, A.; Yamada, S.; Kawahara, N. Bilateral spontaneous simultaneous femoral neck occult fracture in a middle-aged man due to osteoporosis and vitamin D deficiency osteomalacia: A case report and literature review. Int. J. Surg. Case Rep. 2019, 60, 358–362. [Google Scholar] [CrossRef]
- Al-Rubae, A.Y. The potential uses of Melia azedarach L. as pesticidal and medicinal plant, review. Am.-Eurasian J. Sustain. Agric. 2009, 3, 185–194. [Google Scholar]
- Han, C.; Chen, J.; Liu, Z.; Chen, H.; Yu, F.; Yu, W. Morphological and Physiological Responses of Melia azedarach Seedlings of Different Provenances to Drought Stress. Agronomy 2022, 12, 1461. [Google Scholar] [CrossRef]
- Rana, A. Melia azedarach: A phytopharmacological review. Pharmacogn. Rev. 2008, 2, 173–179. [Google Scholar]
- Sultana, S.; Asif, H.M.; Akhtar, N.; Waqas, M.; Rehman, S.U. Comprehensive Review on Ethanobotanical Uses, Phytochemistry and Pharmacological Properties of Melia azedarach Linn. Asian J. Pharm. Res. Health Care 2014, 6, 26–32. [Google Scholar]
- Kumar, R.; Singh, R.; Meera, P.S.; Kalidhar, S. Chemical components and insecticidal properties of Bakain (Melia azedarach L.)—A review. Agric. Rev. 2003, 24, 101–115. [Google Scholar]
- Ervina, M. A review: Melia azedarach L. as a potent anticancer drug. Pharmacogn. Rev. 2018, 12, 94–102. [Google Scholar] [CrossRef]
- Singh, B.; Pandya, D.; Mankad, A. A review on different pharmacological & biological activities of Azadirachta indica A. Jusm. and Melia azedarach L. J. Plant Sci. Res. 2020, 36, 53–59. [Google Scholar]
- Akacha, M.; Lahbib, K.; Daami-Remadi, M.; Boughanmi, N.G. Antibacterial, antifungal and anti-inflammatory activities of Melia azedarach ethanolic leaf extract. Bangladesh J. Pharmacol. 2016, 11, 666–674. [Google Scholar] [CrossRef]
- Li, N.; Shao, T.; Zhou, Y.; Cao, Y.; Hu, H.; Sun, Q.; Long, X.; Yue, Y.; Gao, X.; Rengel, Z. Effects of planting Melia azedarach L. on soil properties and microbial community in saline-alkali soil. Land Degrad. Dev. 2021, 32, 2951–2961. [Google Scholar] [CrossRef]
- Klein, A.; Keyster, M.; Ludidi, N. Caffeic acid decreases salinity-induced root nodule superoxide radical accumulation and limits salinity-induced biomass reduction in soybean. Acta Physiol. Plant. 2013, 35, 3059–3066. [Google Scholar] [CrossRef]
- Thind, S.; Barn, G. Caffeic acid and calcium application affects electrolyte leakage, hydrolases and cytosolute contents of heat stressed cotton seedlings. J. Cotton Res. Dev. 2012, 26, 77–80. [Google Scholar]
- El-Soud, W.A.; Hegab, M.M.; AbdElgawad, H.; Zinta, G.; Asard, H. Ability of ellagic acid to alleviate osmotic stress on chickpea seedlings. Plant Physiol. Biochem. 2013, 71, 173–183. [Google Scholar] [CrossRef]
- Saleh, A.M.; Madany, M. Coumarin pretreatment alleviates salinity stress in wheat seedlings. Plant Physiol. Biochem. 2015, 88, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Shakoor, A.; Saleem, M.F.; Anjum, S.A.; Wahid, M.A.; Saeed, M.T. Effect of heat stress and benzoic acid as foliar application on earliness and nutrients uptake in cotton. J. Agric. Res. 2017, 55, 15–28. [Google Scholar]
- Noreen, S.; Ashraf, M.; Hussain, M.; Jamil, A. Exogenous application of salicylic acid enhances antioxidative capacity in salt stressed sunflower (Helianthus annuus L.) plants. Pak. J. Bot. 2009, 41, 473–479. [Google Scholar]
- Gunes, A.; Inal, A.; Alpaslan, M.; Eraslan, F.; Bagci, E.G.; Cicek, N. Salicylic acid induced changes on some physiological parameters symptomatic for oxidative stress and mineral nutrition in maize (Zea mays L.) grown under salinity. J. Plant Physiol. 2007, 164, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Qayyum, A.; Razzaq, A.; Ahmad, M.; Mahmood, I.; Sher, A. Role of foliar application of salicylic acid and l-tryptophan in drought tolerance of maize. J. Anim. Plant Sci. 2012, 22, 768–772. [Google Scholar]
- Li, Q.; Wang, G.; Wang, Y.; Yang, D.; Guan, C.; Ji, J. Foliar application of salicylic acid alleviate the cadmium toxicity by modulation the reactive oxygen species in potato. Ecotoxicol. Environ. Saf. 2019, 172, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Fujita, M. Salicylic acid alleviates copper toxicity in rice (Oryza sativa L.) seedlings by up-regulating antioxidative and glyoxalase systems. Ecotoxicology 2013, 22, 959–973. [Google Scholar] [CrossRef]
- El-Katony, T.M.; El-Bastawisy, Z.M.; El-Ghareeb, S.S. Timing of salicylic acid application affects the response of maize (Zea mays L.) hybrids to salinity stress. Heliyon 2019, 5, e01547. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, I.; Smoleñ, S. Effect of foliar application of salicylic acid on the response of tomato plants to oxidative stress and salinity. J. Elem. 2013, 18, 239–254. [Google Scholar] [CrossRef]
- Husen, A.; Iqbal, M.; Sohrab, S.S.; Ansari, M.K.A. Salicylic acid alleviates salinity-caused damage to foliar functions, plant growth and antioxidant system in Ethiopian mustard (Brassica carinata A. Br.). Agric. Food Secur. 2018, 7, 44. [Google Scholar] [CrossRef] [Green Version]
- Askari, E.; Ehsanzadeh, P. Drought stress mitigation by foliar application of salicylic acid and their interactive effects on physiological characteristics of fennel (Foeniculum vulgare Mill.) genotypes. Acta Physiol. Plant. 2015, 37, 4. [Google Scholar] [CrossRef]
- Cheng, Z.-Y.; Sun, L.; Wang, X.-J.; Sun, R.; An, Y.-Q.; An, B.-L.; Zhu, M.-X.; Zhao, C.-F.; Bai, J.-G. Ferulic acid pretreatment alleviates heat stress in blueberry seedlings by inducing antioxidant enzymes, proline, and soluble sugars. Biol. Plant. 2018, 62, 534–542. [Google Scholar] [CrossRef]
- Amist, N.; Singh, N. Comparative effects of benzoic acid and water stress on wheat seedlings. Russ. J. Plant Physiol. 2018, 65, 709–716. [Google Scholar] [CrossRef]
- Quan, N.T.; Xuan, T.D. Foliar application of vanillic and p-hydroxybenzoic acids enhanced drought tolerance and formation of phytoalexin momilactones in rice. Arch. Agron. Soil Sci. 2018, 64, 1831–1846. [Google Scholar] [CrossRef]
- Alzohairy, M.A. Therapeutics role of Azadirachta indica (Neem) and their active constituents in diseases prevention and treatment. Evid.-Based Complement. Altern. Med. 2016, 2016, 7382506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashmat, I.; Azad, H.; Ahmed, A. Neem (Azadirachta indica A. Juss)—A nature’s drugstore: An overview. Int. Res. J. Biol. Sci. 2012, 1, 76–79. [Google Scholar]
- Neycee, M.; Nematzadeh, G.; Dehestani, A.; Alavi, M. Assessment of antifungal effects of shoot extracts in chinaberry (Melia azedarach) against 5 phytopathogenic fungi. Int. J. Agric. Crop Sci. 2012, 4, 474–477. [Google Scholar]
- Faccin-Galhardi, L.C.; Yamamoto, K.A.; Ray, S.; Ray, B.; Linhares, R.E.C.; Nozawa, C. The in vitro antiviral property of Azadirachta indica polysaccharides for poliovirus. J. Ethnopharmacol. 2012, 142, 86–90. [Google Scholar] [CrossRef]
- Paul, R.; Prasad, M.; Sah, N.K. Anticancer biology of Azadirachta indica L. (neem): A mini review. Cancer Biol. Ther. 2011, 12, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Koona, S.; Budida, S. Antibacterial Potential of the Extracts of the Leaves of Azadirachta indica Linn. Not. Sci. Biol. 2011, 3, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.D.; Sarwar, M.S.; Dewan, S.M.R.; Hossain, M.S.; Shahid-Ud-Daula, A.; Islam, M.S. Investigation of total phenolic content and antioxidant activities of Azadirachta indica roots. Avicenna J. Phytomed. 2014, 4, 97. [Google Scholar] [PubMed]
- Khan, Z.I.; Mansha, A.; Saleem, M.H.; Tariq, F.; Ahmad, K.; Ahmad, T.; Farooq Awan, M.U.; Abualreesh, M.H.; Alatawi, A.; Ali, S. Trace Metal Accumulation in Rice Variety Kainat Irrigated with Canal Water. Sustainability 2021, 13, 13739. [Google Scholar] [CrossRef]
- Goel, N.; Paul, P.K. Polyphenol oxidase and lysozyme mediate induction of systemic resistance in tomato, when a bioelicitor is used. J. Plant Prot. Res. 2015, 55, 343–350. [Google Scholar] [CrossRef]
- Dheeba, B.; Niranjana, R.; Sampathkumar, P.; Kannan, K.; Kannan, M. Efficacy of neem (Azadirachta indica) and tulsi (Ocimum sanctum) leaf extracts against early blight of tomato. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2015, 85, 327–336. [Google Scholar] [CrossRef]
- Shah, F.M.; Razaq, M.; Ali, A.; Han, P.; Chen, J. Comparative role of neem seed extract, moringa leaf extract and imidacloprid in the management of wheat aphids in relation to yield losses in Pakistan. PLoS ONE 2017, 12, e0184639. [Google Scholar] [CrossRef] [Green Version]
- Riaz, T.; Nawaz Khan, S.; Javaid, A. Management of corm-rot disease of gladiolus by plant extracts. Nat. Prod. Res. 2010, 24, 1131–1138. [Google Scholar] [CrossRef]
- Prasad, S.M.; Dwivedi, R.; Singh, R.; Singh, M.; Singh, D. Neem Leaf Aqueous Extract Induced Growth, Pigments, and Photosynthesis Responses of Cyanobacterium Nostoc muscorum. Philipp. J. Sci. 2007, 136, 75. [Google Scholar]
- Tariq, F.; Wang, X.; Saleem, M.H.; Khan, Z.I.; Ahmad, K.; Saleem Malik, I.; Munir, M.; Mahpara, S.; Mehmood, N.; Ahmad, T.; et al. Risk Assessment of Heavy Metals in Basmati Rice: Implications for Public Health. Sustainability 2021, 13, 8513. [Google Scholar] [CrossRef]

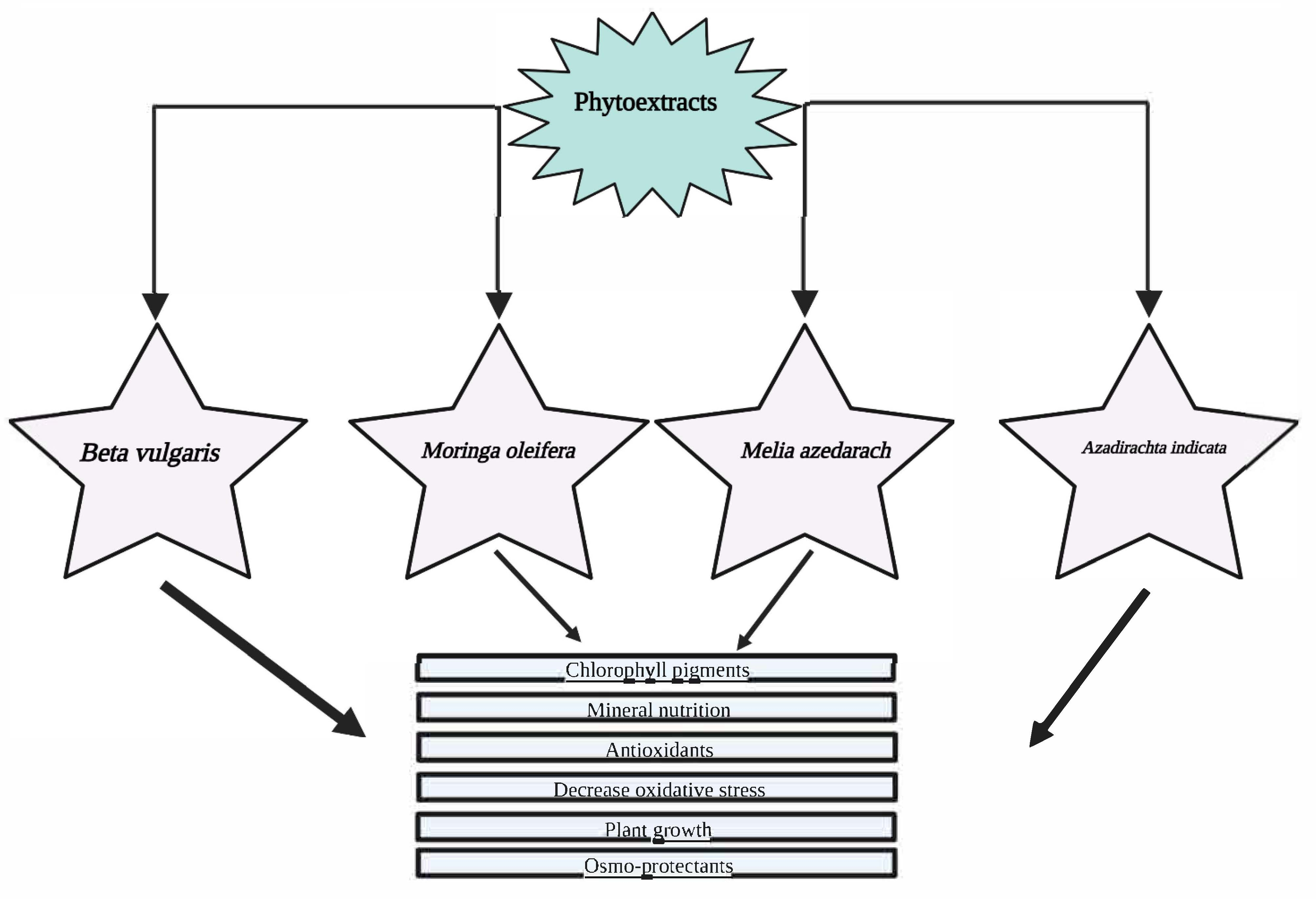{kind=link}
| Type of Stress | Mode of Application | Concentration | Plant/Species | Effects | Reference |
|---|---|---|---|---|---|
| Salt | Foliar spray | 50 mM | Okra | Enhanced growth, gaseous exchange and mineral nutrients uptake under saline stress conditions | [41] |
| Salt | Foliar spray | 50 and 100 mM | Wheat | Accumulation of GB, in or outflux of nutrients, activities of SOD, POD and CAT enzymes | [42] |
| Drought | Foliar and pre-sowing | 50 and 100 mM | Sunflower | Improvement in water status and turgor potential of cells/tissues under water stress conditions | [43] |
| Drought | Foliar spray | 30 mM | Maize | Enhanced sugars, oil proteins, fiber, Ash, moisture, GB, micro and macro nutrients in seeds of maize | [44] |
| Drought | Foliar spray | 50 and 100 mM | Wheat | High biomass production, shoot length, transpiration rate, root P, N and shoot K+ under varying water regimes | [45] |
| Salt | Foliar spray | 5 and 10 mM | Cowpea | Improved plant growth, yield production and biochemical constituents under saline conditions | [46] |
| Drought | Foliar spray | 100 mM | Rice | Improved growth, yield, chlorophyll pigments and leaf fluorescence | [47] |
| Drought | Foliar spray | 100 mM | Rice | Improved chlorophyll, carotenoids, leaf fluorescence and yield attributes | [48] |
| Drought | Foliar spray | 100 mM | Rice | Increased proline, soluble sugar, starch, paddy yield and yield/plant under water stress conditions | [49] |
| Drought | Foliar spray | 25 and 50 mM | Carapa guianensis | Improved GB accumulation, and activities of CAT and APX enzymes | [50] |
| Drought | Foliar spray | 100 mM | Wheat | Improvement in proline and GB accumulaton | [51] |
| Drought | Foliar spray | 4 mM | Pea | Increased soluble proteins, yield as well as activities of SOD, APX and CAT enzymes | [52] |
| Stress | Plant in Which Transferred | Gene | Effects | Reference |
|---|---|---|---|---|
| Freezing stress | Arabidopsis thaliana | CodA | Enhanced tolerance against stress | [53] |
| Salt stress | Arabidopsis thaliana | CodA | Increase in accumulation of GB under stress condition | [54] |
| Drought and salt stress | Tobacco | AhCMO | Tolerance against stress | [55] |
| Chilling stress | Tomato | Cod A | Increase in accumulation of GB | [56] |
| Salt and water stress | Tomato | CodA | Improved RWC, chlorophyll, proline and GB | [57] |
| Chilling stress | Maize | CodA | Increased germination, GB, photosynthesis, soluble sugars and aminoacids | [58] |
| Salt stress | Rice | COX | Increased endogenous GB accumulation | [59] |
| Oxidative, salt and drought stress | Potato | CodA | Reduced membrane damage, high biomass production and RWC | [60] |
| Water stress | Rice | CodA | Protected photosynthetic machinery | [61] |
| Low phosphate | Tomato | CodA | Enhanced enzymes activity and phosphate uptake | [62] |
| Salt stress | Tomato | CodA | Regulation of transporters and ions channels | [63] |
| Salt stress | Tobacco | BADH | Protected enzymes and improved photosynthesis | [64] |
| Vitamins | Levels | Stress | Crops/Species | Effects | References |
|---|---|---|---|---|---|
| Thiamine | 25, 50, 75, 100, 125 and 150 mg/L | Salt | Maize | Reduced Na+ concentration, MDA, H2O2, RMP while improving N, P, Ca2+, and K+, growth, chlorophyll and the activities of CAT, SOD and POD | [68] |
| Ascorbic acid | 0.1 and 0.5 1 mM | Salt | Saccharum spp. | Improved growth, activity of POD and SOD as well as proline contents | [69] |
| AsA | 0.5 and 1 mM | Drought | Wheat | Enhanced net photosynthesis rate, chlorophyll and growth | [70] |
| AsA | 150 mg/L | Drought | Quinoa | Improved growth, RMP, Proline, GB, AsA, TSP, amino acids, total soluble sugars, reducing and non-reducing sugars activities of SOD and POD enzymes | [71] |
| Thiamine | 5 and 10 mg/L | Salt | Sunflower | Reduced leaf water potential, improved RWC, chlorophyll, total amino acids, dry mass and concentration of K+ | [72] |
| Thiamine | 50 and 100 mM | Drought | White clover | Improved biomass, shoot root length and chlorophyll pigments | [73] |
| Tocopherol | 0.25, 0.5 and 1 mM | Salt | Vicia faba | Increased growth, leaf area, yield, RWC and nutrients uptake | [74] |
| Tocopherol | 100, 200 and 300 mg/L | Drought | Mung bean | Improved plant height, total soluble proteins, ascorbic acid, amino acids, activities of POD and CAT enzymes while reducing MDA contents | [75] |
| AsA and Tocopherol | 400 mg/L | Salt | Flax | Reduced peroxidation and polyphenol oxidase while accumulating proline, antioxidants and carbohydrates | [76] |
| Phenolic Compounds | Levels | Stress | Crops/Species | Effects | Reference |
|---|---|---|---|---|---|
| Caffeic acid | 100 µM | Salt | Soybean | Decreased superoxide radical, improved cell viability, SOD, growth, manganese SOD isoforms and Cu/Zn SOD isoforms | [109] |
| Caffeic acid | 10 and 20 mg/L | Heat | Cotton | Decreased electrolyte leakage and amino acids, increased alpha and beta amylase activity | [110] |
| Ellagic acid | 50 ppm | Osmotic | Chickpea | Enhanced germination growth, Proline, GB, flavonoids, GSH, CAT, POX, SOD and GR while lowering MDA, H2O2, and electrolyte leakage | [111] |
| Coumarin | 50 ppm | Salt | Wheat | Improved osmolytes, soluble sugars, K+/Na+ and antioxidants | [112] |
| Benzoic acid | 0.25, 0.50, 0.75 and 1 mM | Heat | Cotton | Improved N, P, K and Z uptake | [113] |
| Salicylic acid | 100, 200 and 300 mg/L | Salt | Sunflower | Improved biomass, growth and photosynthetic rate | [114] |
| Salicylic acid | 0.1, 0.5 and 1 mM | Salt | Maize | Increased growth and uptake of N, mg, Fe, Mn and Cu while inhibiting Na+ and Cl- | [115] |
| Salicylic acid | 100, 150 and 200 ppm | Drought | Maize | Improved chlorophyll, RWC, K content and leaf membrane stability | [116] |
| Salicylic acid | 600 µMS | Cd toxicity | Potato | Increased RWC, chlorophyll, proline, CAT, SOD, APX, GR decreased MDA, H2O2, O2- | [117] |
| Salicylic acid | 100 µM | Cu toxicity | Rice | Improved RWC, chlorophyll, AsA and redox ratio | [118] |
| Salicylic acid | 1 mM | Salt | Maize | Increased sugar, proline, while decreasing K+ and phenolic contents | [119] |
| Salicylic acid | 0.01% | Salt | Tomato | Increased AsA while decreasing phenolic compounds and amino acids | [120] |
| Salicylic acid | 0.5 mM | Salt | Mustard | Modulated cell redox balance and increased the activities of enzymes | [121] |
| Salicylic acid | 0.5 and 1 mM | Drought | Fennel | Increased water potential, RWC, osmolytes, chlorophyll, carotenoids and seed essential contents | [122] |
| Ferulic acid | 0.6 mM | Heat | Blueberry | Increased proline, soluble sugars, RWC, transcription of genes encoding cu/zn SOD, CAT, GR, while decreasing H2O2, MDA, SO2− | [123] |
| Cinnamic acid | 0.5, 1 and 1.5 mM | Drought | Wheat | Improved proline, SOD, APX, guiacol peroxidase | [124] |
| Vannilic acid and p-hydroxybenzoic acids | 25 and 50 µM | Drought | Rice | Improved flavonoids, phenolics, activities of antioxidants | [125] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akram, N.A.; Saleem, M.H.; Shafiq, S.; Naz, H.; Farid-ul-Haq, M.; Ali, B.; Shafiq, F.; Iqbal, M.; Jaremko, M.; Qureshi, K.A. Phytoextracts as Crop Biostimulants and Natural Protective Agents—A Critical Review. Sustainability 2022, 14, 14498. https://doi.org/10.3390/su142114498
Akram NA, Saleem MH, Shafiq S, Naz H, Farid-ul-Haq M, Ali B, Shafiq F, Iqbal M, Jaremko M, Qureshi KA. Phytoextracts as Crop Biostimulants and Natural Protective Agents—A Critical Review. Sustainability. 2022; 14(21):14498. https://doi.org/10.3390/su142114498
Chicago/Turabian StyleAkram, Nudrat Aisha, Muhammad Hamzah Saleem, Sidra Shafiq, Hira Naz, Muhammad Farid-ul-Haq, Baber Ali, Fahad Shafiq, Muhammad Iqbal, Mariusz Jaremko, and Kamal Ahmad Qureshi. 2022. "Phytoextracts as Crop Biostimulants and Natural Protective Agents—A Critical Review" Sustainability 14, no. 21: 14498. https://doi.org/10.3390/su142114498