
The Diversity and Composition of Soil Microbial Community Differ in Three Typical Wetland Types of the Sanjiang Plain, Northeastern China


Abstract
:1. Introduction
2. Materials and Methods
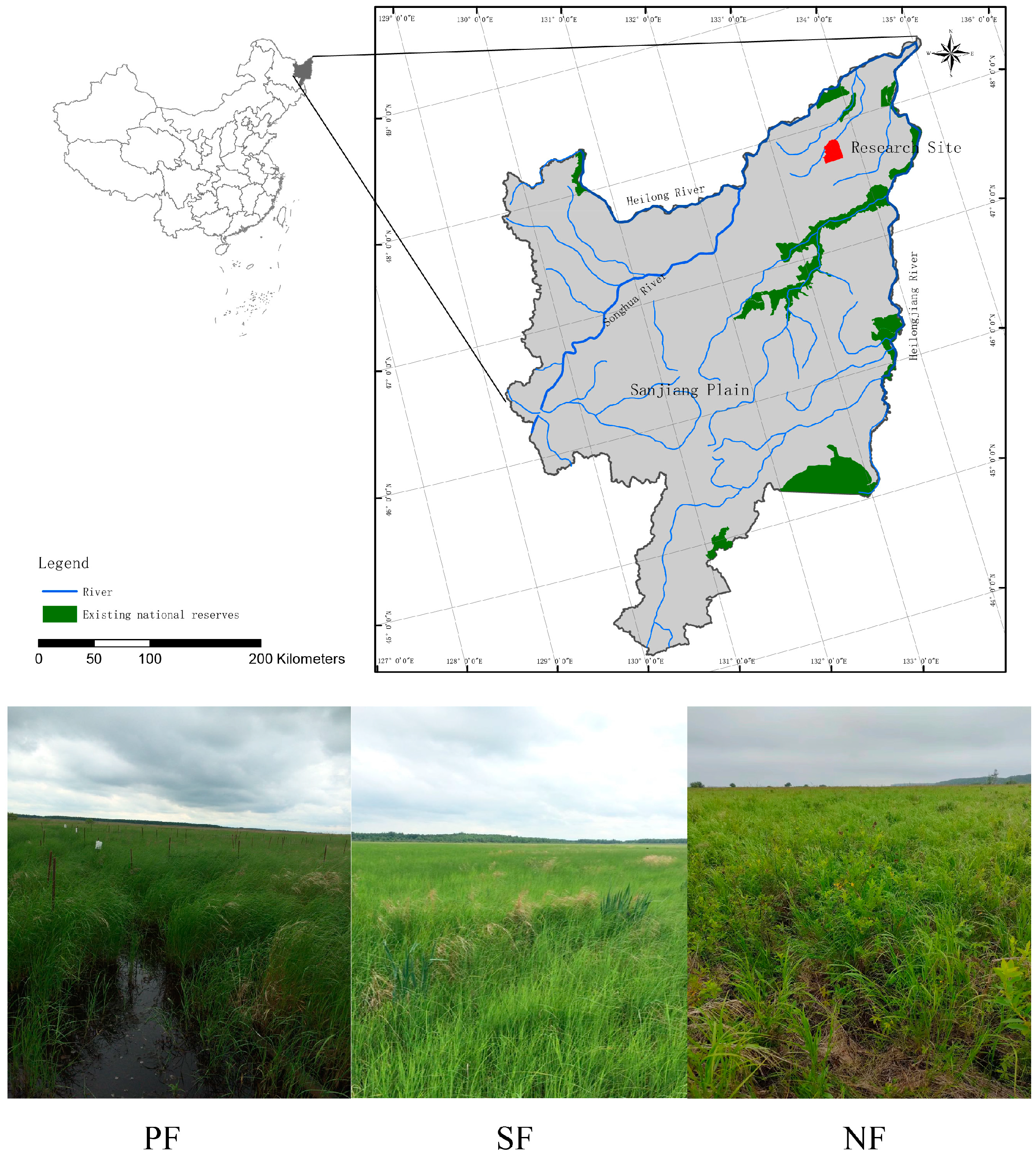
2.1. Study Site
2.2. Sampling Method
2.3. Analysis of Physical and Chemical Properties of Soil
2.4. Phospholipid Fatty Acid Analysis (PLFA)
2.5. Data Analysis
- Shannon-Wiener (H): H = ∑(Pi)(lnPi)
- Simpson (D): D = 1 − ΣPi2
- 3.
- Margalef (M): M = (S − 1)/lnN
- 4.
- Menhinick (E): E = S/√N
- 5.
- Brillouin (B): B = N − 1lg(N!/n1!n2!…n!)
3. Results
3.1. Physical and Chemical Properties of the Soil in Different Wetland Types
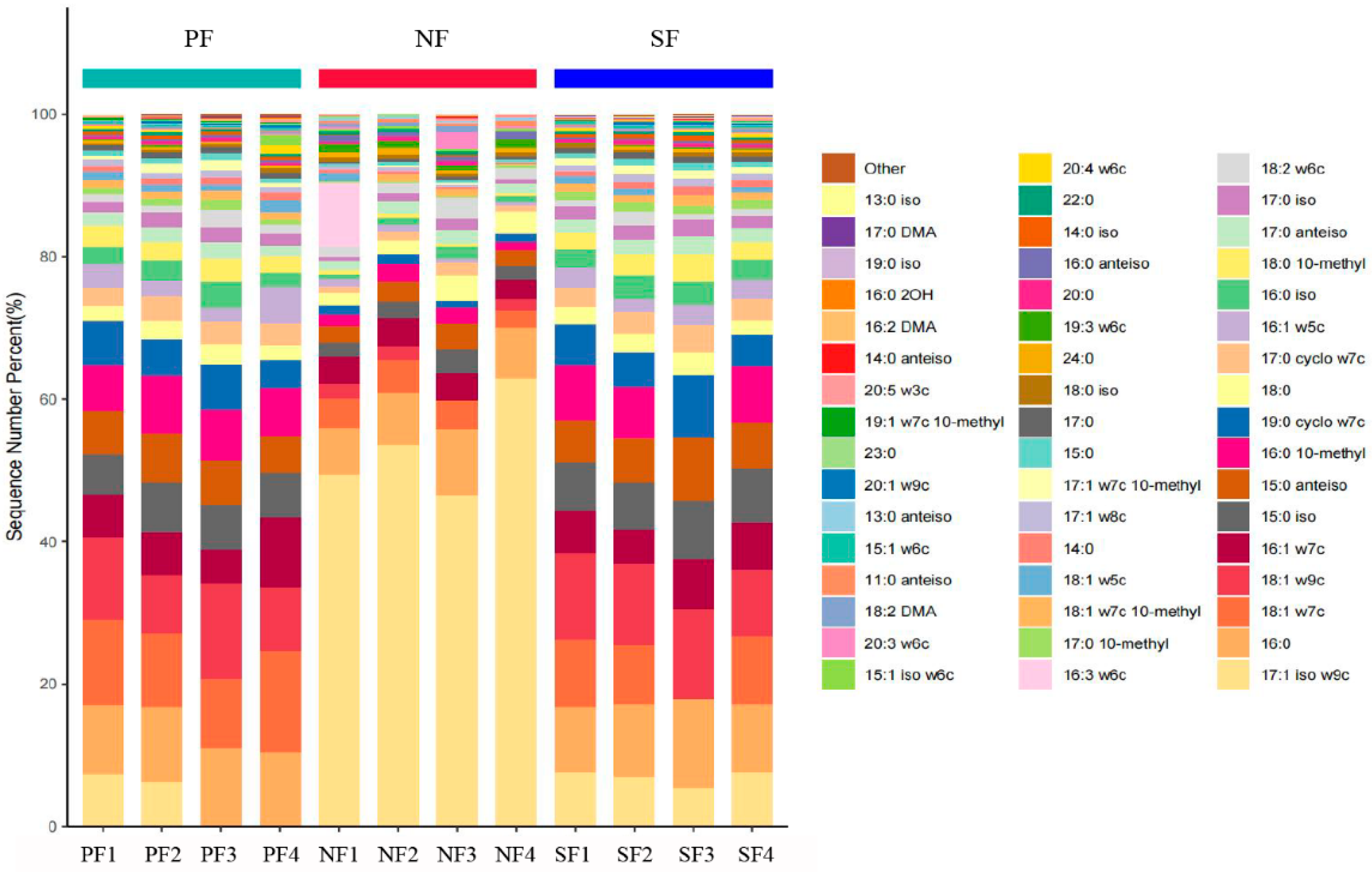
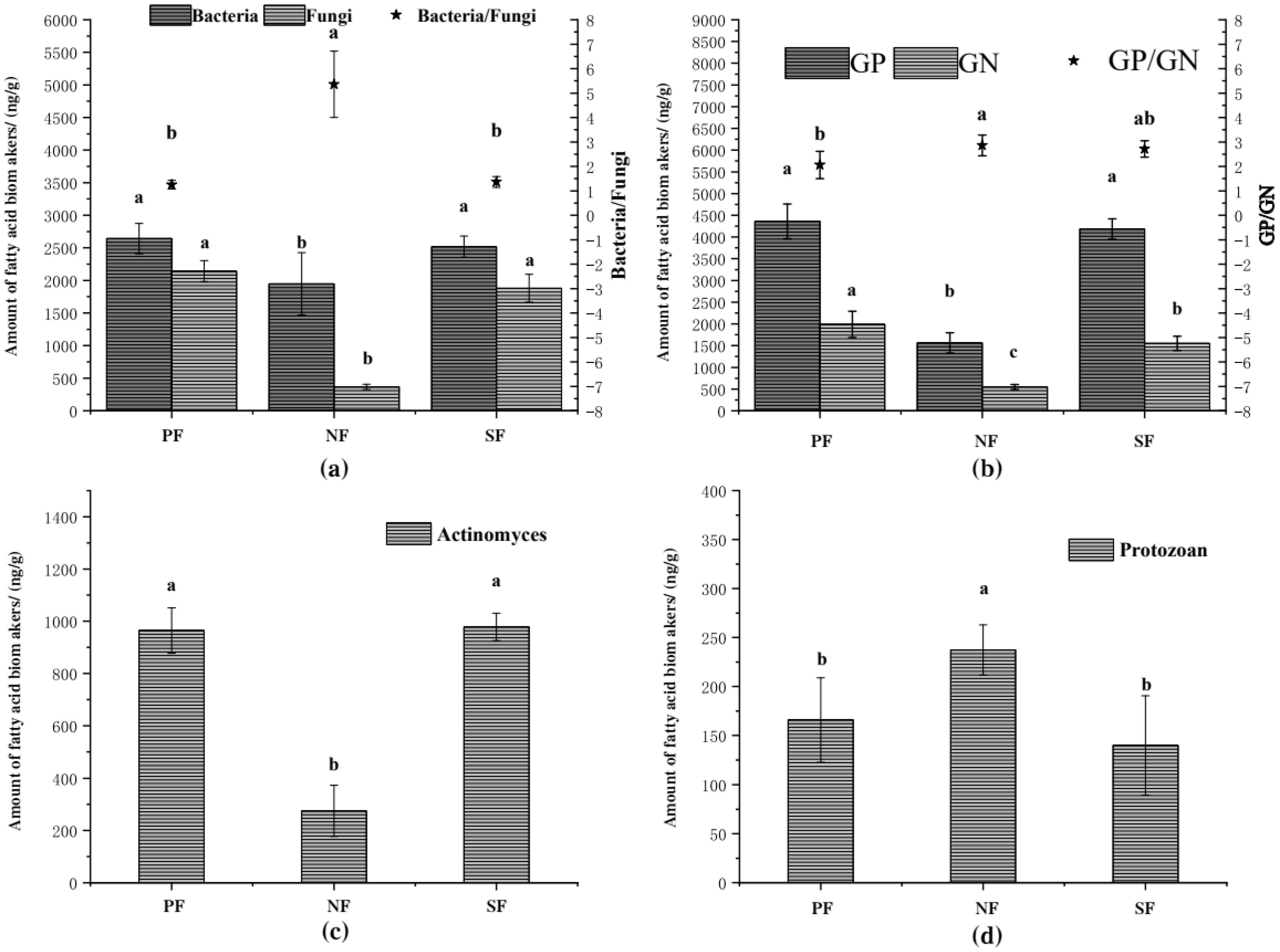
3.2. Relative Abundance of Soil Microbial PLFA in the Wetlands of the Sanjiang Plain
3.3. Soil Diversity Index Analysis of the Different Wetland Types
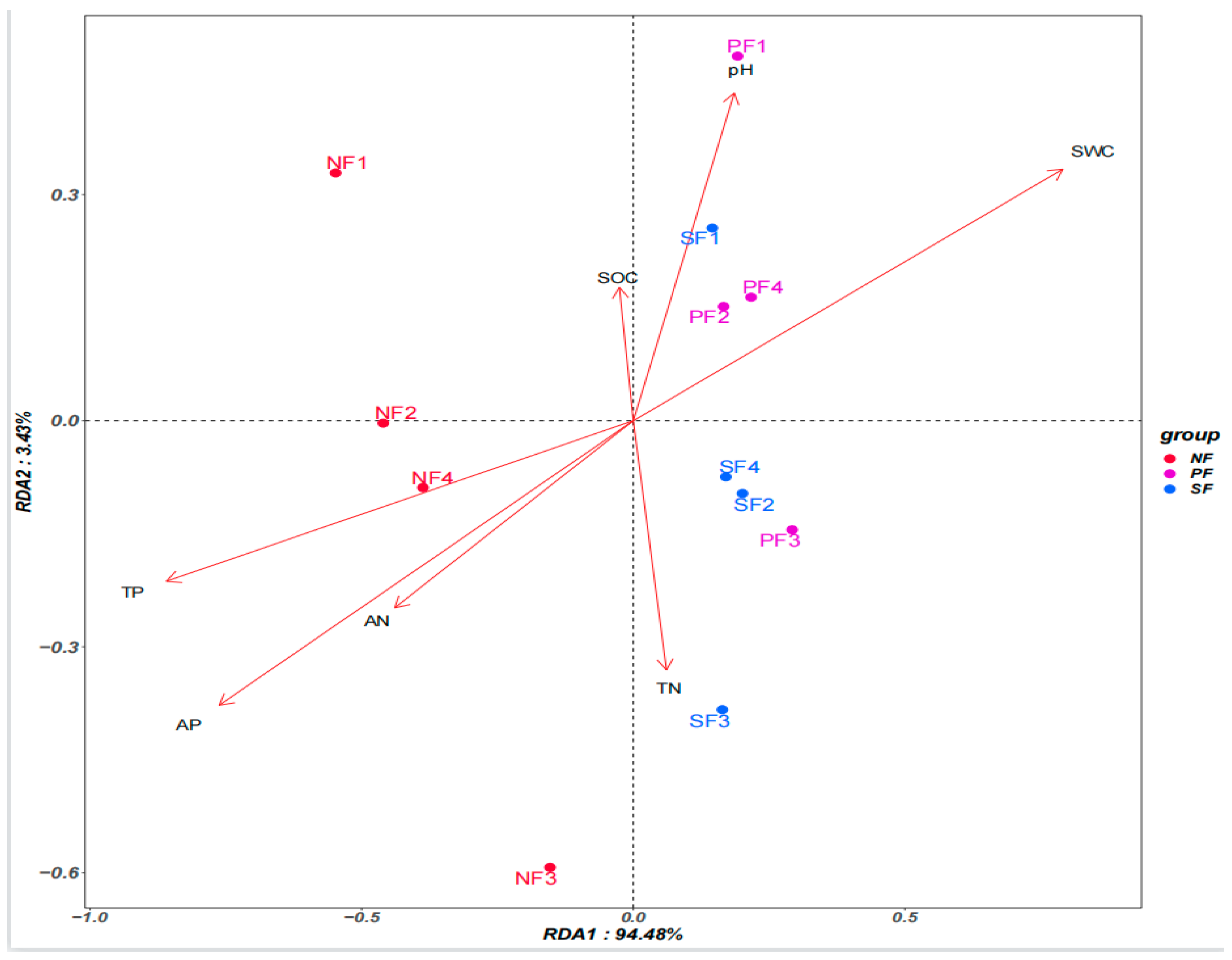
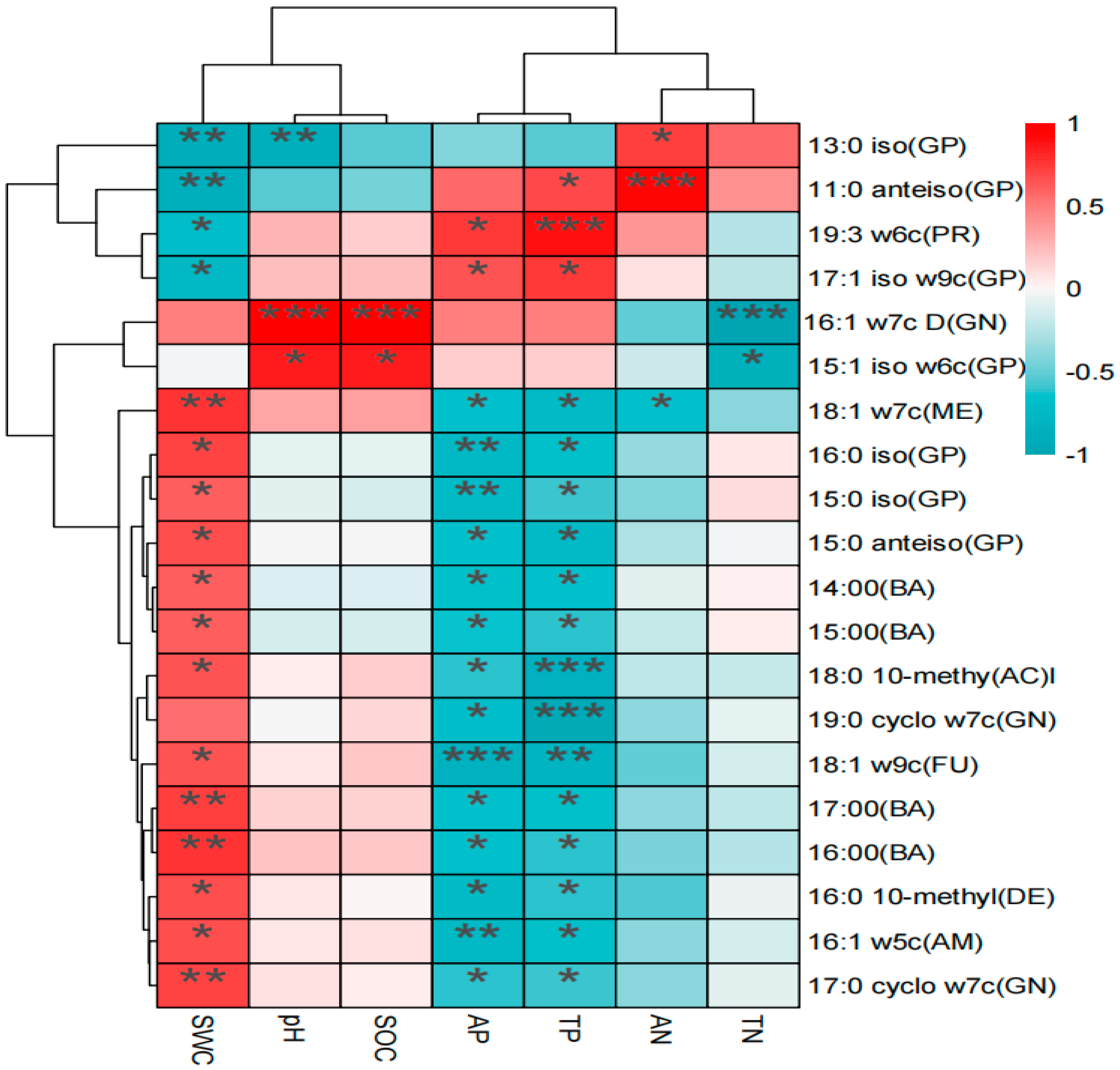
3.4. Relationship between Soil Microbial Structure and Soil Chemical Properties
4. Discussion
4.1. Changes in Soil Physicochemical Properties in Different Wetland Types of Sanjiang Plain
4.2. Effects of Wetland Types on Soil Microbial Composition
4.3. Effects of Wetland Types on Soil Microbial Diversity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, R.T.; Sui, X.; Fu, X.Y.; Liu, Y.N. Effects of elevated CO2 concentration on litter decomposition in wetland ecosystems. Territ. Nat. Resour. Study 2019, 2, 75–78. [Google Scholar] [CrossRef]
- Zhang, R.T.; Sui, X.; Fu, X.Y.; Liu, Y.N.; Ni, H.W. Influence of Simulation Nitrogen Deposition on Soil Microbial Functional Diversity of Calamagrostis Angustifolia Wetland in Sanjiang Plain. Acta Agrestia Sin. 2016, 24, 1226–1233. [Google Scholar] [CrossRef]
- Kotzé, E.; Sandhage-Hofmann, A.; Amelung, W.; Oomen, R.J.; du Preez, C.C. Soil microbial communities in different rangeland management systems of a sandy savanna and clayey grassland ecosystem, South Africa. Nutr. Cycl. Agroecosyst. 2017, 107, 227–245. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, R.; Frey, B.; Yang, L.; Liu, Y.; Ni, H.; Li, M. Soil physicochemical properties drive the variation in soil microbial communities along a forest successional series in a degraded wetland in northeastern China. Ecol. Evol. 2021, 11, 2194–2208. [Google Scholar] [CrossRef]
- Sieben, E.J. Zonal and azonal vegetation revisited: How is wetland vegetation distributed across different zonobiomes. Austral Ecol. 2019, 44, 449–460. [Google Scholar] [CrossRef]
- Chen, X.; Chen, H.; Zhao, J.; Xin, Y.; Li, Y. Bacterial community structure and diversity in the rhizospheric soil of Robinia pseudoacacia and Juniperus sabina planted in iron tailings matrix. Environ. Sci. Pollut. Res. 2022, 1–13. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, Y.; Geng, Y.; Huang, G.; Cui, X.; Hou, M. Effects of Different Land Types on Soil Enzyme Activity in the Qinghai Lake Region. Wetlands 2018, 38, 711–721. [Google Scholar] [CrossRef]
- White, C.; Sayer, J.; Gadd, G. Microbial solubilization and immobilization of toxic metals: Key biogeochemical processes for treatment of contamination. FEMS Microbiol. Rev. 1997, 20, 503–516. [Google Scholar] [CrossRef]
- Kunito, T.; Akagi, Y.; Park, H.-D.; Toda, H. Influences of nitrogen and phosphorus addition on polyphenol oxidase activity in a forested Andisol. Forstwiss. Cent. 2009, 128, 361–366. [Google Scholar] [CrossRef]
- Bell, J.K.; Siciliano, S.; Lamb, E.G. A survey of invasive plants on grassland soil microbial communities and ecosystem services. Sci. Data 2020, 7, 86. [Google Scholar] [CrossRef]
- Schlatter, D.C.; Bakker, M.; Bradeen, J.; Kinkel, L.L. Plant community richness and microbial interactions structure bacterial communities in soil. Ecology 2015, 96, 134–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, A.J.; Simpson, M.J.; Smith, E.; Kelleher, B.P. Microbially Derived Inputs to Soil Organic Matter: Are Current Estimates Too Low? Environ. Sci. Technol. 2007, 41, 8070–8076. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, Z.; Tian, L.; Li, X.; Tian, C. Microbial communities in peatlands along a chronosequence on the Sanjiang Plain, China. Sci. Rep. 2017, 7, 9567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Zhou, X.; Tian, L.; Ma, L.; Luo, S.; Zhang, J.; Li, X.; Tian, C. Fungal communities in ancient peatlands developed from different periods in the Sanjiang Plain, China. PLoS ONE 2017, 12, e0187575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Xu, N.; Wang, H.; Li, J.; Zhong, H.; Dong, H.; Zeng, Z.; Zong, C. Variations in the diversity of the soil microbial community and structure under various categories of degraded wetland in Sanjiang Plain, northeastern China. Land Degrad. Dev. 2021, 32, 2143–2156. [Google Scholar] [CrossRef]
- Ji, Y.; Wang, J.H.; Yuan, Z.N.; Guan, J.F.; Liu, C.Y.; Zhu, Y.N. Impact of different restoration methods on fungal community diversity in Qixing River wetland. J. Eng. Heilongjiang Univ. 2017, 8, 60–66. [Google Scholar] [CrossRef]
- Wang, J.F.; Han, D.Y.; Wang, J.B.; Fu, X.L. Variations in plant species composition and diversity of Calamagrostis angustifolia community along soil water level gradient in the Sanjiang Plain. Acta Ecol. Sin. 2017, 37, 3515–3524. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.B.; Wang, Z.Y.; Wu, L.H.; Yang, D.Q.; Ding, C.H.; Wang, M.; Wang, R.; Fu, X.; Wang, Z.B. Conversion of wetlands to farmland and forests reduces soil microbial functional diversity and carbon use intensity. Appl. Ecol. Environ. Res. 2022, 20, 4553–4564. [Google Scholar] [CrossRef]
- Blair, G.; Lefroy, R.; Lisle, L. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems. Aust. J. Agric. Res. 1995, 46, 1459–1466. [Google Scholar] [CrossRef]
- Zeng, Q.T. The Influence Factors and Elimination Methods of the Determination of Nitrogen in Geochemical Samples by Sei-micro Kjeldahl Method. Guangdong Chem. Ind. 2018, 45, 225–226+220. [Google Scholar]
- Pang, Y.Q.; Zhang, Q.Q. Improvement of alkali fusion-molybdenum antimony anti-spectrophotometric method for the determination of soil phosphorus indicators. Acta Geol. Sichuan 2021, 41, 167–168. [Google Scholar] [CrossRef]
- De Silva, C.S.; Koralage, I.S.A.; Weerasinghe, P.; Silva, N.R.N. The Determination of Available Phosphorus in Soil: A Quick and Simple Method. OUSL J. 2015, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.P.; Pang, D.B.; Chen, L.; Wan, H.Y.; Chen, G.L.; Li, X.B. Phospholipid fatty acid analysis of soil microbes in typical vegetation types at different elevation on the east slope of Helan Mountain. Acta Ecol. Sin. 2022, 42, 5045–5058. [Google Scholar] [CrossRef]
- Yang, G.S.; Song, C.C.; Song, Y.Y.; Hou, C.C. Characteristics of microbial activities in soil profiles of Calamagrostis angustifolia wetlands in Sanjiang Plain. Acta Ecol. Sin. 2010, 30, 6146–6153. [Google Scholar]
- Yang, G.S.; Song, C.C.; Wang, L.; Wang, L.L.; Li, Y.C. Influence of water level gradient on marsh soil microbial activity of Calamagrostis angustifolia. Environ. Sci. 2010, 31, 444–449. [Google Scholar] [CrossRef]
- Ji, Y.H.; Lu, X.G.; Yang, Q.; Zhao, K.Y. Recent thirty years evolvement trend of Calamagrostis angustifolia community in Sanjiang Plain. Chin. J. Ecol. 2006, 25, 1328–1332. [Google Scholar] [CrossRef]
- Tu, C.; He, T.; Lu, X.; Luo, Y.; Smith, P. Extent to which pH and topographic factors control soil organic carbon level in dry farming cropland soils of the mountainous region of Southwest China. CATENA 2018, 163, 204–209. [Google Scholar] [CrossRef]
- Lou, H.; Yang, S.; Zhao, C.; Zhou, Q.; Bai, J.; Hao, F.; Wu, L. Phosphorus risk in an intensive agricultural area in a mid-high latitude region of China. CATENA 2015, 127, 46–55. [Google Scholar] [CrossRef]
- Brockett, B.F.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Yu, S.; Ehrenfeld, J.G. The effects of changes in soil moisture on nitrogen cycling in acid wetland types of the New Jersey Pinelands (USA). Soil Biol. Biochem. 2009, 41, 2394–2405. [Google Scholar] [CrossRef]
- Gartzia-Bengoetxea, N.; Kandeler, E.; de Arano, I.M.; Arias-González, A. Soil microbial functional activity is governed by a combination of tree species composition and soil properties in temperate forests. Appl. Soil Ecol. 2016, 100, 57–64. [Google Scholar] [CrossRef]
- Wang, J.F.; Li, M.S.; Zhang, R.T.; Sui, X.; Ma, W.C.; Ni, H.W. Assessment of microbial diversity of Deyeuxia angustifolia wetland through phospholipid fatty acids (PLFA) in Sanjiang plain. Int. J. Agric. Biol. 2018, 20, 1463–1470. [Google Scholar] [CrossRef]
- Wang, Q.L.; Cao, G.M.; Wang, C.T. Quantitative characters of soil microbes and microbial biomass under different vegetations in alpine meadow. Chin. J. Ecol. 2007, 26, 1002–1008. [Google Scholar] [CrossRef]
- Xu, S.; Wang, Y.; Guo, C.; Zhang, Z.; Shang, Y.; Chen, Q.; Wang, Z.-L. Comparison of Microbial Community Composition and Diversity in Native Coastal Wetlands and Wetlands that Have Undergone Long-Term Agricultural Reclamation. Wetlands 2017, 37, 99–108. [Google Scholar] [CrossRef]
- Bossio, D.A.; Scow, K.M. Impact of carbon and flooding on the metabolic diversity of microbial communities in soils. Appl. Environ. Microbiol. 1995, 61, 4043–4050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drenovsky, R.; Vo, D.; Graham, K.; Scow, K. Soil Water Content and Organic Carbon Availability Are Major Determinants of Soil Microbial Community Composition. Microb. Ecol. 2004, 48, 424–430. [Google Scholar] [CrossRef]
- Xu, F.; Zhang, T.; Huai, B.D.; Sui, W.Z.; Yang, X. Effects of Land Use Changes on Soil Fungal Community Structure and Function in the Riparian Wetland Along the Downstream of the Songhua River. Environ. Sci. 2021, 42, 2531–2540. [Google Scholar] [CrossRef]
- Anderson, I.C.; Cairney, J.W.G. Diversity and ecology of soil fungal communities: Increased understanding through the application of molecular techniques. Environ. Microbiol. 2010, 6, 769–779. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen–phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Gundersen, P.; Zhang, T.; Mo, J. Effects of phosphorus addition on soil microbial biomass and community composition in three forest types in tropical China. Soil Biol. Biochem. 2012, 44, 31–38. [Google Scholar] [CrossRef]
- Li, J.; Li, Z.; Wang, F.; Zou, B.; Chen, Y.; Zhao, J.; Mo, Q.; Li, Y.; Li, X.; Xia, H. Effects of nitrogen and phosphorus addition on soil microbial community in a secondary tropical forest of China. Biol. Fertil. Soils 2015, 51, 207–215. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, T.; Gilliam, F.S.; Gundersen, P.; Zhang, W.; Chen, H.; Mo, J. Interactive Effects of Nitrogen and Phosphorus on Soil Microbial Communities in a Tropical Forest. PLoS ONE 2013, 8, e61188. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Hu, B.; Qi, K.; Chen, W.; Pang, X.; Bao, W.; Tian, G. Effects of phosphorus addition on soil microbial biomass and community composition in a subalpine spruce plantation. Eur. J. Soil Biol. 2016, 72, 35–41. [Google Scholar] [CrossRef]
- Teng, Z.; Zhu, Y.; Li, M.; Whelan, M.J. Microbial community composition and activity controls phosphorus transformation in rhizosphere soils of the Yeyahu Wetland in Beijing, China. Sci. Total Environ. 2018, 628–629, 1266–1277. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.P.; Song, Q.; Liu, Z.K.; Zhao, K.; Liu, Y.L.; Xiao, H.; Cao, J.; Rong, Y.P. Effects of grassland utilization on microbial community structure in Hulunbuir grassland. Pratacul. Sci. 2022, 39, 421–430. [Google Scholar] [CrossRef]
- Cai, X.B.; Zhang, Y.Q.; Shao, W. Characteristics of soil fertility in alpine steppes at different degradation grades. Acta Ecol. Sin. 2008, 28, 1034–1044. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wetland Types | Longitude | Latitude | Vegetations |
|---|---|---|---|
| PF | 133°37′43″ | 47°47′16″ | Calamagrostis Angustifolia, Phragmites australis, Carex miyabei var. maopengensis, Stachys baicalensis, Glyceria spiculosa |
| NF | 133°37′51″ | 47°47′21″ | Calamagrostis Angustifolia, Anemone rivularis, Stachys lanata, Sanguisorba tenuifolia, Carex korshinskyi, Lathyrus quinquenervius |
| SF | 133°37′04″ | 47°45′39″ | Calamagrostis Angustifolia, Carex korshinskyi, Trollius paluster, Sanguisorba tenuifolia, Geranium linearilobum |
| Microbial Group * | PLFA Biomarkers |
|---|---|
| Unspecified bacteria (BA) | 12:0, 13:0, 14:0, 15:0, 16:0, 17:0, 18:0, 22:0, 24:0 |
| Gram-positive bacteria (GP) | a11:0, i13:0, a13:0, i14:0, a14:0, i15:0, a15:0, i15:1ω6c, a16:0, i16:0, a17:0, i17:0, i17:1ω9c, i18:0, i19:0 |
| Gram-negative bacteria (GN) | 10:0-2OH, 14:1ω5c, 16:1ω7c, 16:0-2OH, 16:1ω7cDMA, 17:0DMA, 17:1ω8c, 17:0cycloω7c, 19:0cycloω7c, 20:1ω9c, 22:1ω9c |
| Actinomycetes (AC) | 10Me17:0, 10Me17:1ω7c, 10Me18:0, 10Me18:1ω7c, 10Me19:1ω7c |
| Anaerobic Bacteria (AB) | * 16:2DMA, 18:2DMA |
| Aerobic bacteria (AE) | 15:1ω6c |
| Desulfobacteria (DE) | 16:0-methyl |
| Methanotrophs (ME) | 18:1ω5c, 18:1ω6c, 18:1ω7c |
| Unspecified Fungi (FU) | 18:1ω9c, 23:0 |
| Arbuscular Mycorrhiza (AM) | 16:1ω5c |
| Saprophytic Fungi (SF) | 18:2ω6c |
| Protozoa (PR) | 19:3ω6c, 20:3ω6c, 20:4ω6c, 20:5ω3c |
| Wetland Type | pH | AN (mg/kg) | AP (mg/kg) | TP (g/kg) | TN (g/kg) | SOC (g/kg) | SWC (%) |
|---|---|---|---|---|---|---|---|
| Permanently flooded (PF) | 6.00 ± 0.09 a | 496.77 ± 89.02 a | 33.12 ± 1.82 b | 6.36 ± 2.02 b | 4.29 ± 0.08 c | 52.96 ± 2.86 a | 82.67 ± 2.05 a |
| Seasonally flooded (SF) | 5.48 ± 0.05 c | 594.4 ± 66.98 a | 30.43 ± 3.61 b | 6.03 ± 1.82 b | 7.62 ± 0.2 a | 35.05 ± 1 c | 52.33 ± 2.05 b |
| Non-flooded (NF) | 5.65 ± 0.05 b | 613.56 ± 17.74 a | 43.58 ± 2.67 a | 21.79 ± 1.34 a | 5.62 ± 0.69 b | 46.57 ± 4.46 b | 35.00 ± 2.45 c |
| Diversity Index | Permanently Flooded Wetlands | Non-Flooded Wetlands | Seasonally Flooded Wetlands |
|---|---|---|---|
| Shannon-Wiener (H) | 3.05 ± 0.04 a | 2.12 ± 0.09 b | 3.02 ± 0.07 a |
| Simpson (D) | 0.93 ± 0.00 a | 0.71 ± 0.03 b | 0.93 ± 0.01 a |
| Margalef (M) | 4.79 ± 0.27 a | 3.38 ± 0.23 b | 4.94 ± 0.11 a |
| Menhinick (E) | 0.39 ± 0.02 a | 0.27 ± 0.04 b | 0.42 ± 0.02 a |
| Brillouin (B) | 3.02 ± 0.04 a | 2.10 ± 0.09 b | 2.99 ± 0.07 a |
| Diversity Index | pH | TP | AP | TN | AN | SOC | SWC |
|---|---|---|---|---|---|---|---|
| Shannon-Wiener (H) | 0.212 | −0.959 ** | −0.913 ** | 0.072 | −0.432 | −0.118 | 0.864 ** |
| Simpson (D) | 0.188 | −0.964 ** | −0.901 ** | 0.080 | −0.427 | −0.107 | 0.855 ** |
| Margalef (M) | 0.134 | −0.935 ** | −0.940 ** | 0.157 | −0.320 | −0.228 | 0.791 * |
| Menhinick (E) | −0.063 | −0.871 ** | −0.790 * | 0.298 | −0.271 | −0.282 | 0.682 * |
| Brillouin (B) | 0.215 | −0.959 ** | −0.914 ** | 0.069 | −0.433 | −0.116 | 0.866 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Weng, X.; Zhang, R.; Yang, L.; Liu, Y.; Sui, X. The Diversity and Composition of Soil Microbial Community Differ in Three Typical Wetland Types of the Sanjiang Plain, Northeastern China. Sustainability 2022, 14, 14394. https://doi.org/10.3390/su142114394
Wang M, Weng X, Zhang R, Yang L, Liu Y, Sui X. The Diversity and Composition of Soil Microbial Community Differ in Three Typical Wetland Types of the Sanjiang Plain, Northeastern China. Sustainability. 2022; 14(21):14394. https://doi.org/10.3390/su142114394
Chicago/Turabian StyleWang, Mingyu, Xiaohong Weng, Rongtao Zhang, Libin Yang, Yingnan Liu, and Xin Sui. 2022. "The Diversity and Composition of Soil Microbial Community Differ in Three Typical Wetland Types of the Sanjiang Plain, Northeastern China" Sustainability 14, no. 21: 14394. https://doi.org/10.3390/su142114394