
Effects of Forest Vegetation Restoration on Soil Organic Carbon and Its Labile Fractions in the Danxia Landform of China
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design and Soil Sampling
2.3. Laboratory Analysis
2.3.1. Soil Physiochemical Properties and Dry Weight of Litterfall
2.3.2. SOC and Soil LOC Fractions Analyses
2.3.3. Soil Enzyme Activities Analysis
2.4. Statistical Methods
3. Results
3.1. Variation of Soil Physicochemical Properties
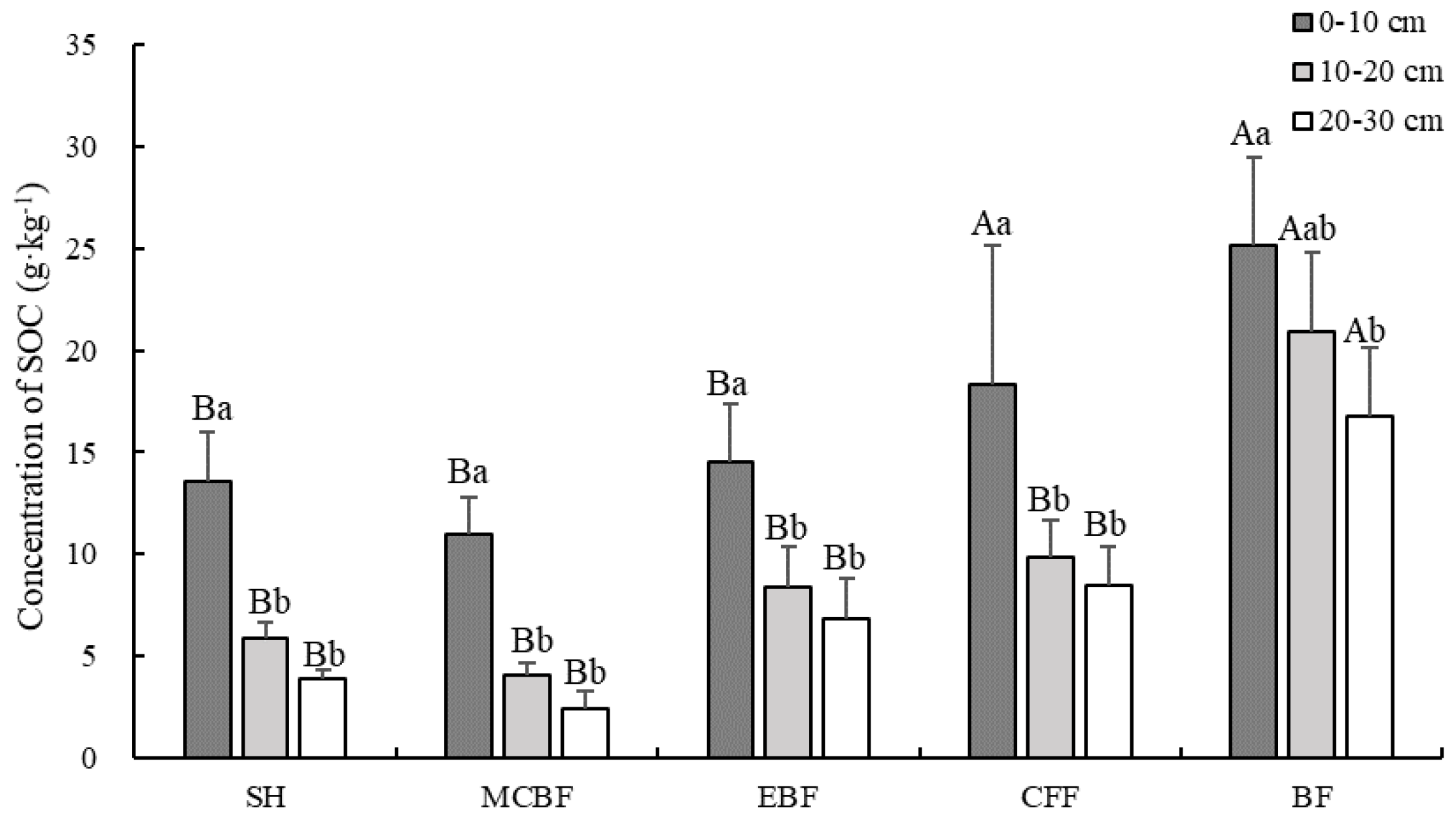
3.2. Changes in SOC Concentrations and SOC Stock
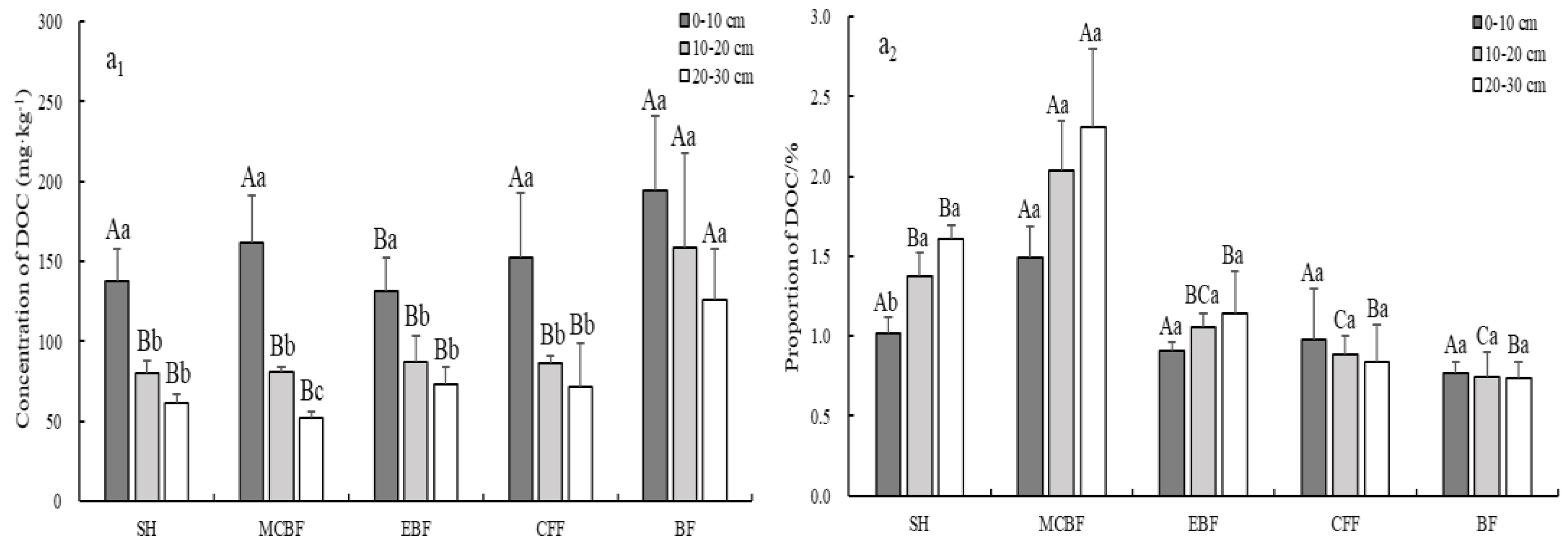
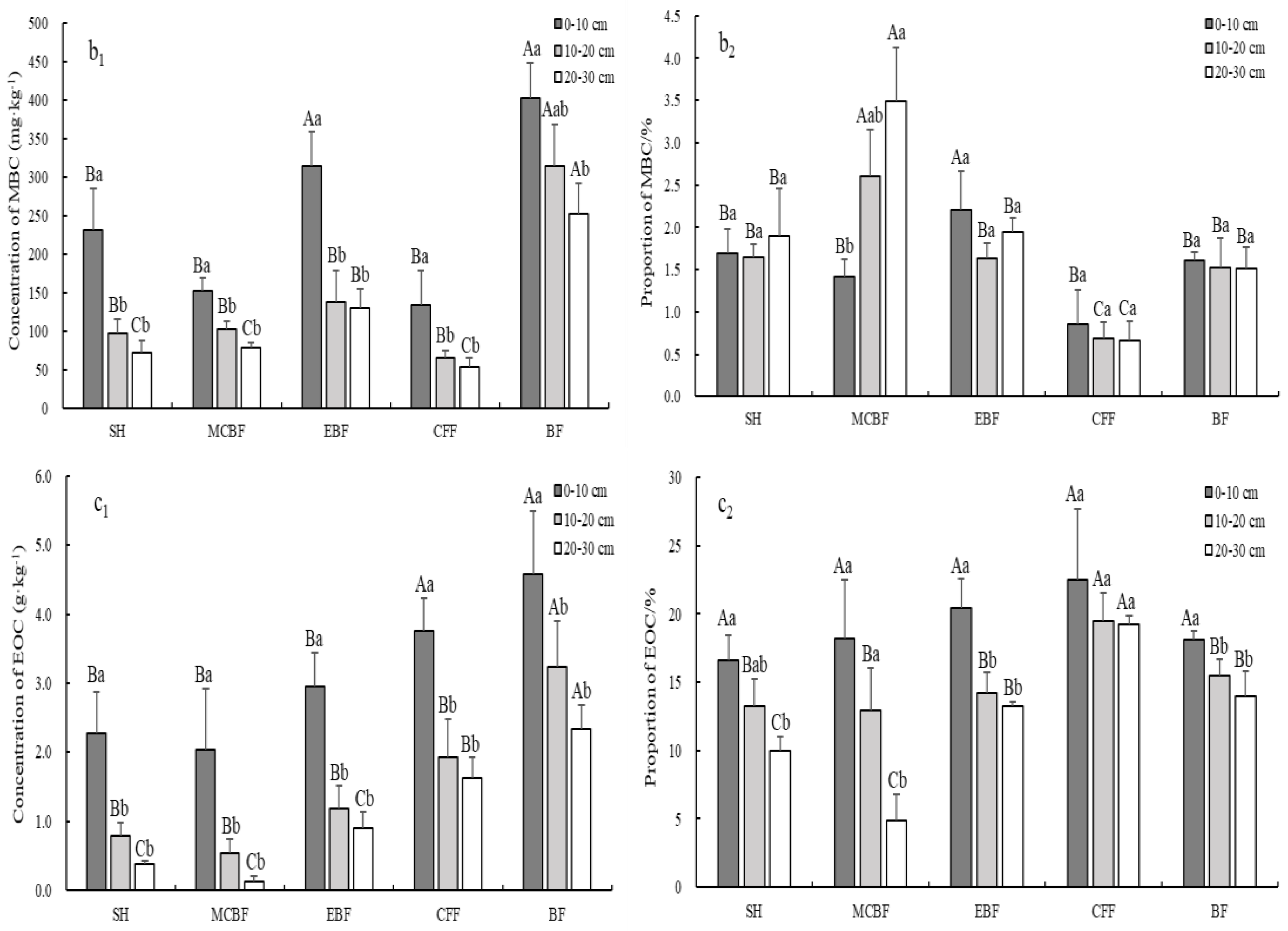
3.3. Variation of Soil LOC Fractions Concentration and Their Stocks
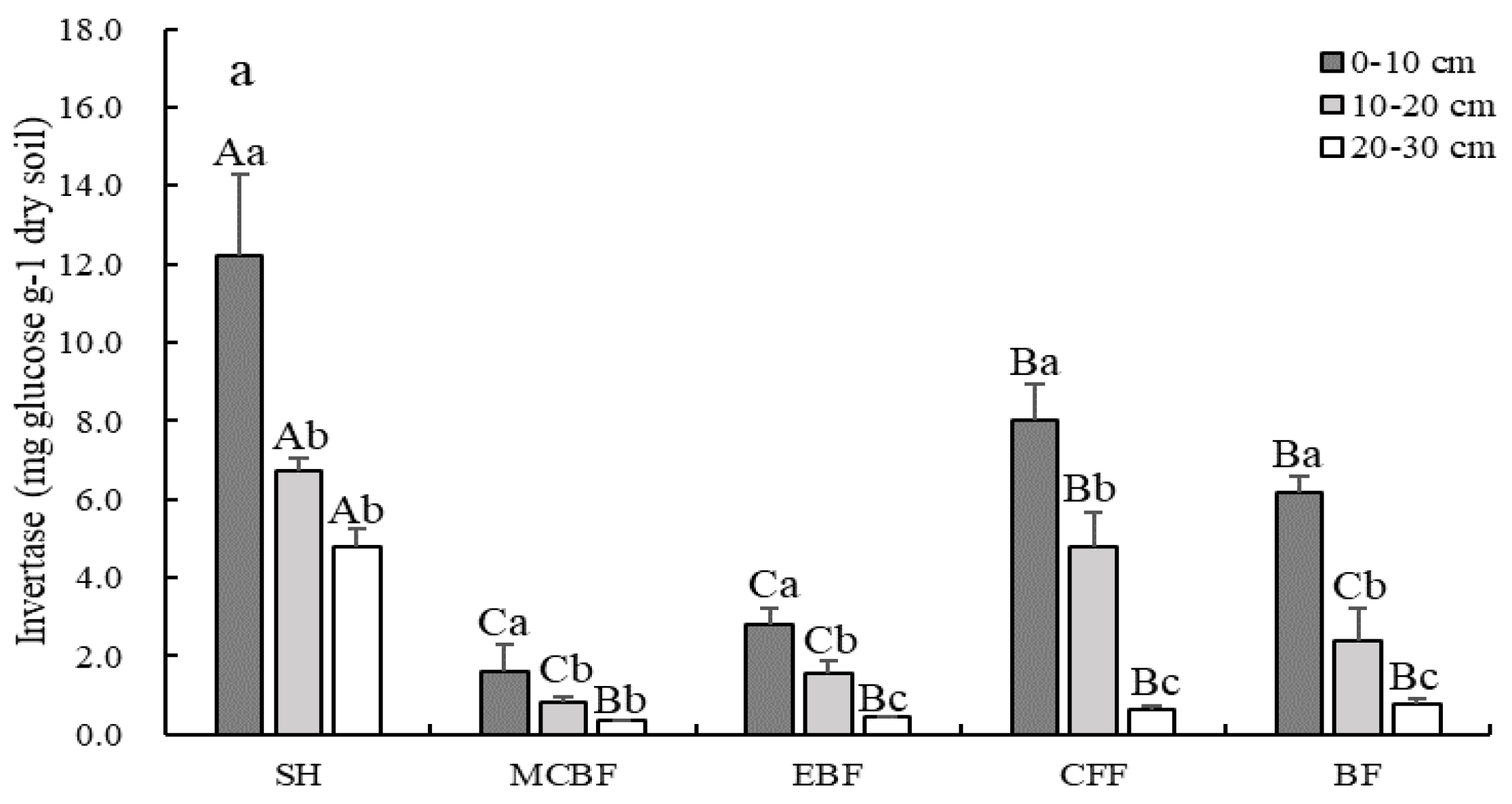
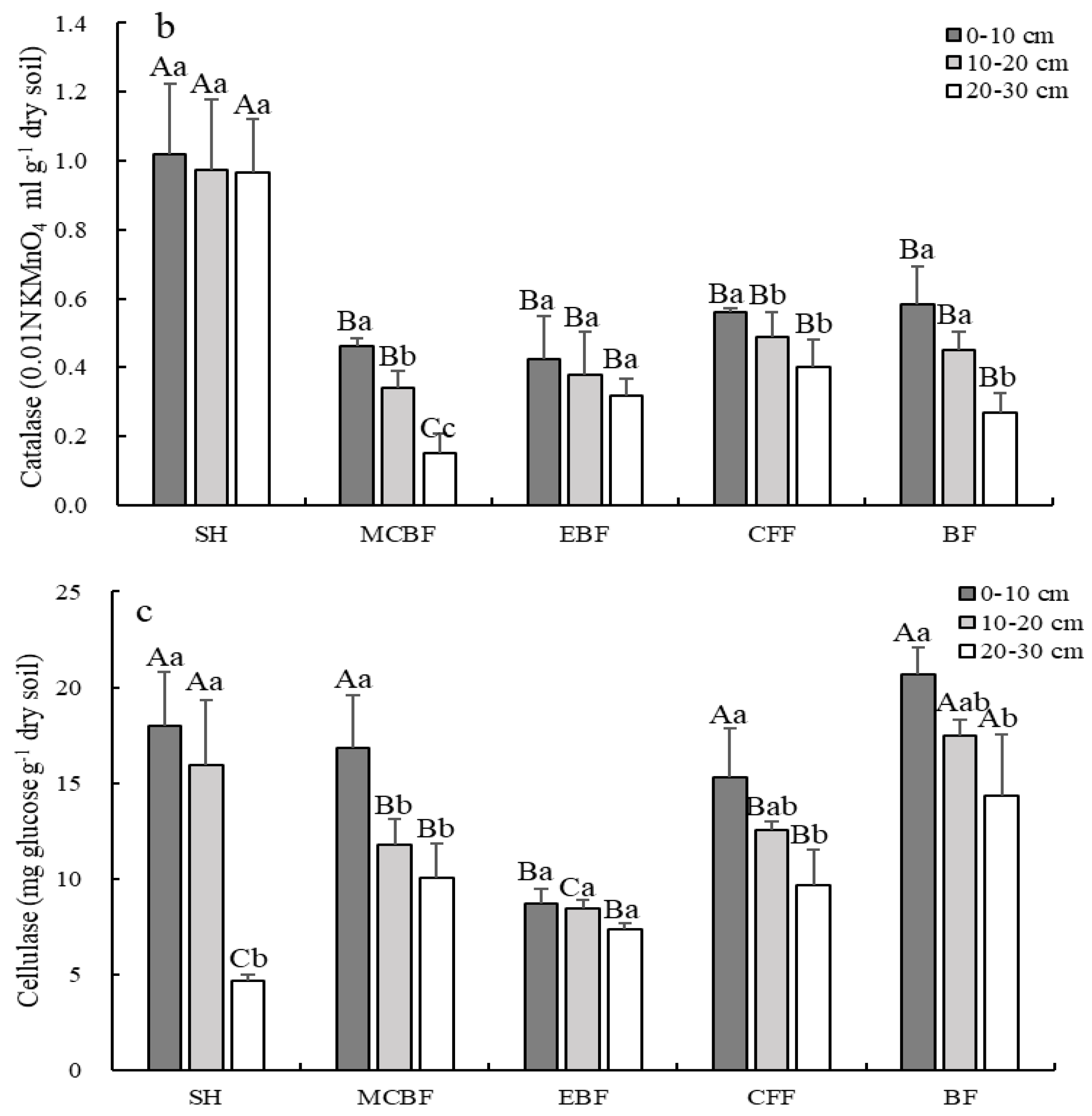
3.4. The Changes of Soil Enzymatic Activities
3.5. Relationship between Soil C Fractions and Soil Physicochemical Properties, Enzymatic Activities
4. Discussion
4.1. Effect of Different Vegetation Restoration Types on SOC Concentrations and Stocks
4.2. Effect of Different Vegetation Restoration Types on Concentrations and Stocks of Soil LOC Fractions
4.3. Contributions of Soil Environmental Factors to Variations in SOC and LOC Fractions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yan, P.; Peng, H.; Yan, L.; Lin, K. Spatial variability of soil physical properties based on GIS and geo-statistical methods in the red beds of the Nanxiong Basin, China. Pol. J. Environ. Stud. 2019, 28, 2961–2972. [Google Scholar] [CrossRef]
- Yang, Q. On the boundary of the loess plateau. J. Natur. Resour. 1988, 3, 9–15. (In Chinese) [Google Scholar]
- Baton, L.; Yang, L. Measurement of carbonate rocks distribution area in China. Carsologica Sin. 1983, 2, 147–150. (In Chinese) [Google Scholar]
- Yan, L.B.; Kasanin-Grubin, M. Land degradation and management of red beds in China: Two case studies. J. Mt. Sci. 2019, 16, 2591–2604. [Google Scholar] [CrossRef]
- Liu, H.; Liu, G.; Zhou, Y.; He, C. Spatial distribution and influence analysis of soil heavy metals in a hilly region of sichuan basin. Pol. J. Environ. Stud. 2017, 26, 725. [Google Scholar] [CrossRef]
- Bai, B.; Nie, Q.; Zhang, Y.; Wang, X.; Hu, W. Cotransport of heavy metals and SiO2 particles at different temperatures by seepage. J. Hydrol. 2021, 597, 125771. [Google Scholar] [CrossRef]
- Li, Y.; Liu, C.; Yuan, X. Spatiotemporal features of soil and water loss in Three Gorges Reservoir Area of Chongqing. J. Geogr. Sci. 2009, 19, 81–94. [Google Scholar] [CrossRef]
- Wang, H.J.; Shi, X.Z.; Yu, D.S.; Weindorf, D.C.; Huang, B.; Sun, W.X.; Ritsema, C.J.; Milne, E. Factors determining soil nutrient distribution in a small-scaled watershed in the purple soil region of Sichuan Province, China. Soil Tillage Res. 2009, 105, 300–306. [Google Scholar] [CrossRef]
- Peng, H.; Ren, F.; Pan, Z. A review of Danxia landforms in China. Z. Für Geomorphol. Suppl. Issues 2015, 59, 19–33. [Google Scholar]
- Li, C.; Li, C.; Zhao, L.; Ma, Y.; Tong, X.; Deng, J.; Ren, C.; Han, X.; Yang, G. Dynamics of storage and relative availability of soil inorganic nitrogen along revegetation chronosequence in the loess hilly region of China. Soil Tillage Res. 2019, 187, 11–20. [Google Scholar] [CrossRef]
- Xu, C.; Xiang, W.; Gou, M.; Chen, L.; Lei, P.; Fang, X.; Deng, X.; Ouyang, S. Effects of forest restoration on soil carbon, nitrogen, phosphorus, and their stoichiometry in Hunan, southern China. Sustainability 2018, 10, 1874. [Google Scholar] [CrossRef]
- Han, C.L.; Sun, Z.X.; Shao, S.; Wang, Q.B.; Libohova, Z.; Owens, P.R. Changes of Soil Organic Carbon after Wildfire in a Boreal Forest, Northeast CHINA. Agronomy 2021, 11, 1925. [Google Scholar] [CrossRef]
- Yan, L.; Peng, H.; Hu, Z.; Kašanin-Grubin, M.; Greif, V.; Chen, Z.; Simonson, S. Stone pillar rockfall in Danxia landform area, Mt. Langshan, Hunan Province, China. Physi. Geogr. 2016, 37, 327–343. [Google Scholar] [CrossRef]
- Moreno-Mateos, D.; Barbier, E.B.; Jones, P.C.; Jones, H.P.; Aronson, J.; Lopez-Lopez, J.A.; McCrackin, M.L.; Meli, P.; Montoya, D.; Rey Benayas, J.M. Anthropogenic ecosystem disturbance and the recovery debt. Nat. Commun. 2017, 8, 14163. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, X.; Gebremikael, M.T.; Wu, H.; Cai, D.; Wang, B.; Li, B.; Zhang, J.; Li, Y.; Xi, J. Response of soil organic carbon fractions, microbial community composition and carbon mineralization to high-input fertilizer practices under an intensive agricultural system. PLoS ONE 2018, 13, e0195144. [Google Scholar] [CrossRef]
- Soucémarianadin, L.N.; Cécillon, L.; Guenet, B.; Chenu, C.; Baudin, F.; Nicolas, M.; Girardin, C.; Barré, P. Environmental factors controlling soil organic carbon stability in French forest soils. Plant Soil 2018, 426, 267–286. [Google Scholar] [CrossRef]
- Xu, G.; Chen, J.; Berninger, F.; Pumpanen, J.; Bai, J.; Yu, L.; Duan, B. Labile, recalcitrant, microbial carbon and nitrogen and the microbial community composition at two Abies faxoniana forest elevations under elevated temperatures. Soil Biol. Biochem. 2015, 91, 1–13. [Google Scholar] [CrossRef]
- Blanco-Moure, N.; Gracia, R.; Bielsa, A.C.; López, M.V. Soil organic matter fractions as affected by tillage and soil texture under semiarid Mediterranean conditions. Soil Tillage Res. 2016, 155, 381–389. [Google Scholar] [CrossRef]
- Ruirui, C.; Mehmet, S.; Sergey, B.; Olga, M.; Klaus, D.; Xiangui, L.; Evgenia, B.; Yakov, K. Soil C and N availability determine the priming effect: Microbial N mining and stoichiometric decomposition theories. Glob. Chang. Biol. 2014, 20, 2356–2367. [Google Scholar]
- Huang, W.L.; Tu, Y.L.; Yang, L. Vegetation of Guizhou; Guizhou People’s Publishing House: Guiyang, China, 1988. [Google Scholar]
- Peng, H. China Danxia; Springer Nature: Berlin, Germany, 2020. [Google Scholar]
- Zhao, J.; Tang, J.B.; Huang, S.S. Study on the categories of soil erosion of Chishui River Basin and corresponding measures for soil and water conservation. Hubei Agric. Sci. 2015, 54, 3369–3371. (In Chinese) [Google Scholar]
- Luo, Y.; Liu, L.; Yang, R.Y. Scientific Investigation and Study on National Nature Protection Area of Subtropical Evergreen Broadleaved Forest in Xishui, Guizhou Province; Science and Technology Press: Guizhou, China, 2011. (In Chinese) [Google Scholar]
- Lu, R.K. Soil Agrochemistry and Analytical Methods; Chinese Agricultural Science and Technology Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Xie, Y.J.; Xie, Y.H.; Xiao, H.Y.; Chen, X.S.; Li, F. Controls on Litter Decomposition of Emergent Macrophyte in Dongting Lake Wetlands. Ecosystems 2017, 20, 1383–1389. [Google Scholar]
- Rahman, F.; Rahman, M.M.; Rahman, G.K.M.M.; Saleque, M.A.; Hossain, A.T.M.S.; Miah, M.G. Effect of organic and inorganic fertilizers and rice straw on carbon sequestration and soil fertility under rice-rice cropping pattern. Carbon Manag. 2016, 7, 41–53. [Google Scholar] [CrossRef]
- Wu, J.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation extraction—an automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Jones, D.L.; Willett, V.B. Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biol. Biochem. 2006, 38, 991–999. [Google Scholar] [CrossRef]
- Blair, G.J.; Lefory, R.D.B.; Lise, L. Soil carbon fractions based on their degree of oxidation and the development of a carbon management index for agricultural system. Aust. J. Agric. Res. 1995, 46, 1459–1466. [Google Scholar] [CrossRef]
- Guan, S.Y. Soil Enzymes and Their Research Methods; Agriculture Press: Beijing, China, 1986. (In Chinese) [Google Scholar]
- Li, Q.; Liang, J.H.; He, Y.Y.; Hu, Q.J.; Yu, S. Effect of land use on soil enzyme activities at karst area in Nanchuan, Chongqing, Southwest China. Plant Soil Environ. 2014, 60, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Carver, R.; Nash, J. Doing Data Analysis with IBM-SPSS: Version 18.0; Cengage Learning: Boston, MA, USA, 2011. [Google Scholar]
- Dabral, P.P.; Baithuri, N.; Pandey, A. Soil Erosion assessment in a hilly catchment of North Eastern India using USLE, GIS and remote sensing. Water Resour. Manag. 2008, 22, 1783–1798. [Google Scholar] [CrossRef]
- Cao, J.; He, X.; Chen, Y.; Chen, Y.; Zhang, Y.; Yu, S.; Zhou, L.; Liu, Z.; Zhang, C.; Fu, S. Leaf litter contributes more to soil organic carbon than fine roots in two 10-year-old subtropical plantations. Sci. Total Environ. 2020, 704, 135341. [Google Scholar] [CrossRef]
- Jia, X.X.; Yang, Y.; Zhang, C.C.; Shao, M.A.; Huang, L.M. A state-space analysis of soil organic carbon in China’s loess plateau. Land Degrad. Dev. 2017, 28, 983–993. [Google Scholar] [CrossRef]
- Yu, H.; Zha, T.; Zhang, X.; Ma, L. Vertical distribution and influencing factors of soil organic carbon in the Loess Plateau, China. Sci. Total Environ. 2019, 693, 133632. [Google Scholar] [CrossRef]
- King, A.E.; Hofmockel, K.S. Diversified cropping systems support greater microbial cycling and retention of carbon and nitrogen. Agric. Ecosyst. Environ. 2017, 240, 66–76. [Google Scholar] [CrossRef]
- Ravenek, J.M.; Bessler, H.; Engels, C.; Scherer-Lorenzen, M.; Gessler, A.; Gockele, A.; Luca, E.D.; Temperton, V.M.; Ebeling, A.; Roscher, C.; et al. Long-term study of root biomass in a biodiversity experiment reveals shifts in diversity effects over time. Oikos 2014, 123, 1528–1536. [Google Scholar] [CrossRef]
- Wiesmeier, M.; Urbanski, L.; Hobley, E.; Lang, B.; Lützow, M.; Marin-Spiotta, E.; Wesemael, B.; Rabot, E.; Ließ, M.; Garcia-Franco, N.; et al. Soil organic carbon storage as a key function of soils-A review of drivers and indicators at various scales. Geoderma 2019, 333, 149–162. [Google Scholar] [CrossRef]
- Fukuzawa, K.; Shibata, H.; Takagi, K.; Satoh, F.; Koike, T.; Sasa, K. Roles of dominant understory Sasa bamboo in carbon and nitrogen dynamics following canopy tree removal in a cool-temperate forest in northern Japan. Plant Species Biol. 2015, 30, 104–115. [Google Scholar] [CrossRef]
- Kramer, C.; Trumbore, S.; Fröberg, M.; Dozal, L.M.C.; Zhang, D.; Xu, X.; Santos, G.M.; Hanson, P.J. Recent (<4 year old) leaf litter is not a major source of microbial carbon in a temperate forest mineral soil. Soil Biol. Biochem. 2010, 42, 1028–1037. [Google Scholar] [CrossRef]
- Lange, M.; Eisenhauer, N.; Sierra, C.A.; Bessler, H.; Engels, C.; Griffiths, R.I.; Mellado-Vázquez, P.G.; Malik, A.A.; Roy, J.; Scheu, S.; et al. Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 2015, 6, 6707. [Google Scholar] [CrossRef]
- Zinn, Y.L.; Andrade, A.B.; Araujo, M.A.; Lal, R. Soil organic carbon retention more affected by altitude than texture in a forested mountain range in Brazil. Soil Res. 2018, 56, 284–295. [Google Scholar] [CrossRef]
- Zech, M.; Hörold, C.; Leiber-Sauheitl, K.; Kühnel, A.; Hemp, A.; Zech, W. Buried black soils on the slopes of Mt. Kilimanjaro as a regional carbon storage hotspot. Catena 2014, 112, 125–130. [Google Scholar] [CrossRef]
- Badía, D.; Ruiz, A.; Girona, A.; Martí, C.; Casanova, J.; Ibarra, P.; Zufiaurre, R. The influence of elevation on soil properties and forest litter in the siliceous Moncayo Massif, SW Europe. J. Mount. Sci. 2016, 13, 2155–2169. [Google Scholar] [CrossRef]
- Lukac, M.; Godbold, D.L. Soil Ecology in Northern Forests: A below Ground View of a Changing World; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Guo, S.L.; Ma, Y.H.; Che, S.G.; Sun, W.Y. Effects of artificial and natural vegetations on litter production and soil organic carbon change in loess hilly areas. Sci. Silvae Sin. 2009, 45, 14–18. [Google Scholar]
- Zhou, Y.; Watts, S.E.; Boutton, T.W.; Archer, S.R. Root density distribution and biomass allocation of co-occurring woody plants on contrasting soils in a subtropical savanna parkland. Plant Soil 2019, 438, 263–279. [Google Scholar] [CrossRef]
- Wang, M.; Wang, S.; Cao, Y.; Jiang, M.; Wang, G.; Dong, Y. The effects of hummock-hollow microtopography on soil organic carbon stocks and soil labile organic carbon fractions in a sedge peatland in Changbai Mountain, China. Catena 2021, 201, 105204. [Google Scholar] [CrossRef]
- Wang, H.; Wu, J.; Li, G.; Yan, L. Changes in soil carbon fractions and enzyme activities under different vegetation types of the northern Loess Plateau. Ecol. Evol. 2020, 10, 12211–12223. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Huang, Z.; Lu, X. Changes of soil labile organic carbon fractions and their relation to soil microbial characteristics in four typical wetlands of Sanjiang Plain, Northeast China. Ecol. Eng. 2015, 82, 381–389. [Google Scholar] [CrossRef]
- Luo, Y.; Li, Q.; Shen, J.; Wang, C.; Li, B.; Yuan, S.; Zhao, B.; Li, H.; Zhao, J.W.; Guo, L.K.; et al. Effects of agricultural land use change on organic carbon and its labile fractions in the soil profile in an urban agricultural area. Land Degrad. Dev. 2019, 30, 1875–1885. [Google Scholar] [CrossRef]
- Li, M.; Zhang, A.; Wu, H.; Liu, H.; Lv, J. Predicting potential release of dissolved organic matter from biochars derived from agricultural residues using fluorescence and ultraviolet absorbance. J. Hazard Mater. 2017, 334, 86–92. [Google Scholar] [CrossRef]
- Kalbitz, K.; Solinger, S.; Park, J.H.; Michalzik, B.; Matzner, E. Controls on the dynamics of dissolved organic matter in soil: A review. Soil Sci. 2000, 165, 277–304. [Google Scholar] [CrossRef]
- Wick, B.; Kuhne, R.F.; Vlek, P.L.G. Soil microbiological parameters as indicators of soil quality under improved fallow management systems in south-western Nigeria. Plant Soil 1998, 202, 97–107. [Google Scholar] [CrossRef]
- Wang, Q.; Xiao, F.; He, T.; Wang, S. Responses of labile soil organic carbon and enzyme activity in mineral soils to forest conversion in the subtropics. Ann. For. Sci. 2013, 70, 579–587. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Meng, X.J.; Gao, L.P.; Sun, X.M.; Fan, J.J.; Xu., L.J. Potential impacts of climate warming on active soil organic carbon contents along natural altitudinal forest transect of Changbai Mountain. Acta Ecol. Sin. 2010, 30, 113–117. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, S.L.; Feng, Z.W.; Gao, L.; Wang, Q.K.; Hu, Y.L.; Yan, S.K. Soil quality assessment of forest stand in different plantation esosystems. Chin. J. Appl. Ecol. 2004, 15, 2199–2205. (In Chinese) [Google Scholar]
- Yang, X.; Lan, Y.; Meng, J.; Chen, W.F.; Huang, Y.W.; Cheng, X.Y.; He, T.Y.; Cao, T.; Liu, Z.Q.; Jiang, L.L.; et al. Effects of maize stover and its derived biochar on greenhouse gases emissions and C-budget of brown earth in Northeast China. Environ. Sci. Pollut. Res. 2017, 24, 8200–8209. [Google Scholar] [CrossRef] [PubMed]
- Duval, M.E.; Galantini, J.A.; Martinez, J.M.; Lopez, F.M.; Wall, L.G. Sensitivity of different soil quality indicators to assess sustainable land management: Influence of site features and seasonality. Soil Tillage Res. 2016, 159, 9–22. [Google Scholar] [CrossRef]
- Zhu, Z.J.; Jiang, P.K.; Xu, Q.F. Study on the active organic carbon in soil under different types of vegetation. Forest Res. 2006, 19, 523–526. (In Chinese) [Google Scholar]
- Xu, H.; Qu, Q.; Lu, B.; Zhang, Y.; Liu, G.; Xue, S. Variation in soil organic carbon stability and driving factors after vegetation restoration in different vegetation zones on the Loess Plateau, China. Soil Tillage Res. 2020, 204, 104727. [Google Scholar] [CrossRef]
- Hu, P.L.; Liu, S.J.; Ye, Y.Y.; Zhang, W.; Wang, K.L.; Su, Y.R. Effects of environmental factors on soil organic carbon under natural or managed vegetation restoration. Land Degrad. Dev. 2018, 29, 387–397. [Google Scholar] [CrossRef]
- Anh, P.T.Q.; Gomi, T.; Macdonald, L.H.; Mizugaki, S.; Khoa, P.V.; Furuichi, T. Linkages among land use, macronutrient levels, and soil erosion in northern Vietnam: A plot-scale study. Geoderma 2014, 232, 352–362. [Google Scholar] [CrossRef]
- Mcdowell, W.H.; Currie, W.S.; Aber, J.D.; Yano, Y. Effects of chronic nitrogen amendments on production of dissolved organic carbon and nitrogen in forest soils. Water Air Soil Pollut. 1998, 105, 175–182. [Google Scholar] [CrossRef]
- Wen, L.; Li, D.; Yang, L.; Luo, P.; Chen, H.; Xiao, K.; Song, T.; Zhang, W.; He, X.; Chen, H.; et al. Rapid recuperation of soil nitrogen following agricultural abandonment in a karst area, southwest China. Biogeochemistry 2016, 129, 341–354. [Google Scholar] [CrossRef]
- Reay, D.S.; Dentener, F.; Smith, P.; Grace, J.; Feely, R.A. Global nitrogen deposition and carbon sinks. Nat. Geosci. 2008, 1, 430–437. [Google Scholar] [CrossRef]
- Li, S.; Zhang, S.R.; Pu, Y.L.; Li, T.; Xu, X.X.; Jia, Y.X.; Deng, O.P.; Gong, G.S. Dynamics of soil labile organic carbon fractions and C-cycle enzyme activities under straw mulch in Chengdu Plain. Soil Tillage Res. 2016, 155, 289–297. [Google Scholar] [CrossRef]
- Feng, J.; Zhu, B. A global meta-analysis of soil respiration and its components in response to phosphorus addition. Soil Biol. Biochem. 2019, 135, 3–47. [Google Scholar] [CrossRef]
- Shen, C.C.; Xiong, J.B.; Zhang, H.Y.; Feng, Y.Z.; Li, X.G.; Li, X.Y.; Liang, W.J.; Chu, H.Y. Soil pH drives the spatial distribution of bacterial communities along elevation on Chang Bai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Yu, X.L. Interactions between soil microbial biomass carbon and soil enzyme activities in Momoge National Nature Reserve. J. Northeast Forest Univer. 2020, 48, 59–63. (In Chinese) [Google Scholar]
- Mayor, Á.G.; Goirán, S.B.; Vallejo, V.R.; Bautista, S. Variation in soil enzyme activity as a function of vegetation amount, type, and spatial structure in fire-prone mediterranean shrublands. Sci. Total Environ. 2016, 573, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.A.; Scelza, R.; Gianfreda, L. Soil Enzymes. Enzymes in Agricultural Sciences; OMICS Group eBooks: Foster City, CA, USA, 2014; pp. 10–43. [Google Scholar]
- Chen, Q.M.; Wang, Z.X.; Liu, Y.; Zheng, H.F.; Li, H.J.; Wang, L.F.; Chen, Y.M.; Shen, X.; Tang, S.Y. Response of soil enzyme activities and stoichiometric ratio to simulated nitrogen deposition in subalpine coniferous forests of western Sichuan. Chin. J. Appl. Environ. Biol. 2019, 25, 791–800. [Google Scholar]
- Ma, W.W.; Li, G.; Wu, J.H.; Xu, G.R.; Wu, J.Q. Response of soil labile organic carbon fractions and carbon-cycle enzyme activities to vegetation degradation in a wet meadow on the Qinghai-Tibet Plateau. Geoderma 2020, 377, 114565. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L. Phenol oxidase, peroxidase and organic matter dynamics of soil. Soil Biol. Biochem. 2010, 42, 391–404. [Google Scholar] [CrossRef]
- Raju, M.N.; Golla, N.; Vengatampalli, R. Soil cellulase. In Soil Enzymes; Springer: Berlin, Germany, 2017; pp. 25–30. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Types | Elevation (m) | Slope (°) | Aspect | Soil Type | Tree Height (m) | Dominant Species |
|---|---|---|---|---|---|---|
| SH | 928 | 24 | West by north 26° | Yellow soil | 1.4–2.0 | Coriariasinica + Vitex negundo + Zanthoxylum planispinum + Lonicera ligustrina |
| MCBF | 955 | 20 | East by south 63° | Yellow soil | 9–18 | Fokieniahodginsis + Pinus massoniana + Castanopsischunii + Elaeocarpus japonicus |
| EBF | 991 | 32 | West by north 73° | Yellow soil | 12–16 | Lithocarpus glabra + Phoebe zhennan + Clethra pinfaensis |
| CFF | 1025 | 41 | West by north 77° | Yellow soil | 17–20 | Cunninghamia lanceolata |
| BF | 1028 | 11 | West by north 9° | Yellow soil | 10–12 | Phyllostachys pubescens |
| Vegetation Type | Soil Depth (cm) | pH | Moisture (g·100 g−1) | BD (g·cm−3) | TN (g·kg−1) | TP (g·kg−1) | Dry Weight of Litterfall (g·m−2) |
|---|---|---|---|---|---|---|---|
| SH | 0–10 | 8.03 ± 0.05 Aa | 16.38 ± 1.36 Ba | 1.50 ± 0.14 Aa | 1.33 ± 0.16 Ba | 0.43 ± 0.01 Ba | 299.04 ± 51.03 B |
| 10–20 | 8.23 ± 0.13 Aa | 14.18 ± 1.74 Ba | 1.58 ± 0.11 ABa | 0.72 ± 0.13 Bb | 0.38 ± 0.00 Bb | ||
| 20–30 | 8.20 ± 0.30 Aa | 13.17 ± 2.23 Ba | 1.74 ± 0.12 Aa | 0.59 ± 0.02 Bb | 0.37 ± 0.01 Bb | ||
| Average | 8.15 ± 0.16 A | 14.57 ± 1.78 B | 1.61 ± 0.12 A | 0.88 ± 0.10 B | 0.40 ± 0.09 B | ||
| MCBF | 0–10 | 5.31 ± 0.37 Ca | 15.37 ± 0.41 Ba | 1.42 ± 0.20 ABa | 1.02 ± 0.24 Ba | 0.22 ± 0.02 Ca | 363.80 ± 44.16 B |
| 10–20 | 5.22 ± 0.10 Ca | 13.33 ± 1.22 Ba | 1.53 ± 0.17 ABa | 0.43 ± 0.13 Cb | 0.16 ± 0.03 Cb | ||
| 20–30 | 5.24 ± 0.04 Ba | 11.90 ± 1.31 Ba | 1.58 ± 0.10 ABa | 0.34 ± 0.11 Cb | 0.13 ± 0.01 Cb | ||
| Average | 5.26 ± 0.17 C | 13.54 ± 2.28 B | 1.51 ± 0.16 ABa | 0.60 ± 0.16 C | 0.17 ± 0.02 C | ||
| EBF | 0–10 | 5.19 ± 0.38 Ca | 25.21 ± 1.45 Aa | 1.50 ± 0.17 Aa | 1.40 ± 0.16 Ba | 0.23 ± 0.02 Ca | 311.32 ± 60.96 B |
| 10–20 | 5.30 ± 0.30 Ca | 23.81 ± 1.44 Aa | 1.56 ± 0.25 ABa | 0.78 ± 0.19 Bb | 0.20 ± 0.03 Ca | ||
| 20–30 | 5.61 ± 0.08 Ba | 21.06 ± 1.09 Ab | 1.65 ± 0.17 Aa | 0.64 ± 0.08 Bb | 0.19 ± 0.03 Ca | ||
| Average | 5.37 ± 0.25 C | 23.36 ± 1.33 A | 1.57 ± 0.20 AB | 0.94 ± 0.15 B | 0.21 ± 0.03 C | ||
| CFF | 0–10 | 6.47 ± 0.29 Ba | 23.96 ± 3.24 Aa | 1.37 ± 0.13 ABb | 1.57 ± 0.67 Ba | 0.27 ± 0.03 Ca | 497.12 ± 32.08 A |
| 10–20 | 6.44 ± 0.60 Ba | 21.98 ± 3.48 Aa | 1.83 ± 0.10 Aa | 1.11 ± 0.21 Ba | 0.24 ± 0.03 Ca | ||
| 20–30 | 6.25 ± 0.95 Ba | 21.68 ± 2.96 Aa | 1.95 ± 0.23 Aa | 1.00 ± 0.47 ABa | 0.21 ± 0.06 Ca | ||
| Average | 6.39 ± 0.61 B | 22.54 ± 3.23 A | 1.71 ± 0.16 A | 1.22 ± 0.45 B | 0.25 ± 0.04 C | ||
| BF | 0–10 | 5.66 ± 0.43 Ca | 28.43 ± 2.95 Aa | 0.96 ± 0.28 Ba | 2.62 ± 0.58 Aa | 0.64 ± 0.06 Aa | 141.65 ± 31.78 C |
| 10–20 | 5.88 ± 0.09 BCa | 23.14 ± 4.34 Aa | 1.17 ± 0.28 Ba | 1.83 ± 0.20 Aab | 0.62 ± 0.06 Aa | ||
| 20–30 | 6.17 ± 0.22 Ba | 21.10 ± 1.94 Aa | 1.25 ± 0.08 Ba | 1.57 ± 0.13 Ab | 0.59 ± 0.07 Aa | ||
| Average | 5.90 ± 0.25 BC | 24.22 ± 3.08 A | 1.13 ± 0.21 B | 2.01 ± 0.30 A | 0.63 ± 0.06 A |
| Soil Layer | SH | MCBF | EBF | CFF | BF |
|---|---|---|---|---|---|
| SOC (t ha−1) | |||||
| 0–10 cm | 20.14 ± 1.91 Aa | 15.30 ± 1.55 Aa | 21.56 ± 2.72 Aa | 24.49 ± 6.95 Aa | 26.46 ± 3.88 Aa |
| 10–20 cm | 9.27 ± 1.31 Cb | 6.13 ± 0.62 Db | 11.91 ± 2.16 Cb | 17.91 ± 2.47 Bb | 23.97 ± 0.99 Aa |
| 20–30 cm | 6.68 ± 0.32 Cb | 3.82 ± 1.51 Cb | 11.09 ± 1.96 Bb | 16.40 ± 3.18 Ab | 20.77 ± 3.15 Aa |
| 0–30 cm | 36.09 ± 2.48 C | 25.25 ± 0.30 D | 44.56 ± 4.46 C | 58.79 ± 4.65 B | 71.02 ± 3.82 A |
| DOC (t·ha−1) | |||||
| 0–10 cm | 0.21 ± 0.03 Aa | 0.23 ± 0.01 Aa | 0.19 ± 0.01 Aa | 0.21 ± 0.07 Aa | 0.18 ± 0.05 Aa |
| 10–20 cm | 0.13 ± 0.02 Bb | 0.12 ± 0.01 Bb | 0.13 ± 0.02 Bb | 0.16 ± 0.01 Ba | 0.19 ± 0.02 Aa |
| 20–30 cm | 0.11 ± 0.01 Ab | 0.08 ± 0.01 Ac | 0.12 ± 0.03 Ab | 0.14 ± 0.04 Aa | 0.16 ± 0.04 Aa |
| 0–30 cm | 0.44 ± 0.04 A | 0.43 ± 0.00 A | 0.44 ± 0.03 A | 0.50 ± 0.11 A | 0.53 ± 0.03 A |
| MBC (t·ha−1) | |||||
| 0–10 cm | 0.34 ± 0.08 ABa | 0.21 ± 0.01 BCa | 0.47 ± 0.04 Aa | 0.18 ± 0.07 Ca | 0.39 ± 0.12 ABa |
| 10–20 cm | 0.15 ± 0.03 Bb | 0.16 ± 0.03 Bab | 0.19 ± 0.04 Bb | 0.12 ± 0.02 Ba | 0.40 ± 0.07 Aa |
| 20–30 cm | 0.13 ± 0.03 Cb | 0.12 ± 0.02 Cb | 0.21 ± 0.02 Bb | 0.10 ± 0.01 Ca | 0.31 ± 005 Aa |
| 0–30 cm | 0.62 ± 0.12 BC | 0.50 ± 0.02 C | 0.88 ± 0.06 A | 0.41 ± 0.04 C | 1.10 ± 0.21 A |
| EOC (t·ha−1) | |||||
| 0–10 cm | 3.37 ± 0.66 Aa | 2.77 ± 0.94 Aa | 4.38 ± 0.27 Aa | 5.10 ± 0.48 Aa | 4.40 ± 1.47 Aa |
| 10–20 cm | 1.24 ± 0.35 Bb | 0.80 ± 0.15 Bb | 1.68 ± 0.26 Bb | 3.48 ± 0.84 Ab | 4.02 ± 0.13 Aa |
| 20–30 cm | 0.67 ± 0.08 Bb | 0.20 ± 0.04 BCb | 1.46 ± 0.23 Bb | 3.16 ± 0.55 Ab | 2.90 ± 0.44 Aa |
| 0–30 cm | 5.28 ± 0.97 BC | 3.77 ± 0.95 C | 7.52 ± 0.64 B | 11.74 ± 1.69 A | 11.32 ± 1.49 A |
| Vegetation Types | Indexes | DOC | MBC | EOC | BD | Moisture | pH | TN | TP | Invertase | Catalase | Cellulase |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SH | SOC | 0.981 ** | 0.962 ** | 0.993 ** | −0.729 * | 0.735 * | −0.447 | 0.990 ** | 0.845 ** | 0.989 ** | 0.090 | 0.017 |
| DOC | 1 | 0.971 ** | 0.989 ** | −0.670 * | 0.706 * | −0.398 | 0.984 ** | 0.849 ** | 0.982 ** | 0.168 | 0.067 | |
| MBC | 1 | 0.970 ** | −0.654 | 0.752 * | −0.509 | 0.972 ** | 0.907 ** | 0.947 ** | 0.281 | 0.048 | ||
| EOC | 1 | −0.697 * | 0.755 * | −0.455 | 0.989 ** | 0.831 ** | 0.988 ** | 0.119 | 0.015 | |||
| MCBF | SOC | 0.971 ** | 0.930 ** | 0.953 ** | −0.567 | 0.485 | −0.026 | 0.951 ** | 0.894 ** | 0.765 * | 0.842 ** | 0.063 |
| DOC | 1 | 0.977 ** | 0.950 ** | −0.565 | 0.476 | 0.080 | 0.955 ** | 0.925 ** | 0.873 ** | 0.881 ** | 0.116 | |
| MBC | 1 | 0.926 ** | −0.473 | 0.548 | 0.100 | 0.930 ** | 0.941 ** | 0.879 ** | 0.864 ** | 0.142 | ||
| EOC | 1 | −0.716 * | 0.467 | −0.155 | 0.968 ** | 0.893 ** | 0.833 ** | 0.796 * | 0.069 | |||
| EBF | SOC | 0.935 ** | 0.901 ** | 0.961 ** | −0.529 | 0.577 | −0.672 * | 0.944 ** | 0.826 ** | 0.725 * | 0.336 | 0.382 |
| DOC | 1 | 0.883 ** | 0.942 ** | −0.417 | 0.642 | −0.765 * | 0.937 ** | 0.729 * | 0.722 * | 0.383 | 0.357 | |
| MBC | 1 | 0.973 ** | −0.410 | 0.703 * | −0.554 | 0.983 ** | 0.832 ** | 0.783 * | 0.272 | 0.446 | ||
| EOC | 1 | −0.440 | 0.644 | −0.624 | 0.990 ** | 0.814 ** | 0.810 ** | 0.264 | 0.454 | |||
| CFF | SOC | 0.447 | 0.490 | 0.837 ** | −0.849 ** | 0.553 | 0.137 | 0.850 ** | 0.479 | 0.746 * | 0.509 | 0.666 |
| DOC | 1 | 0.857 ** | 0.795 * | −0.758 * | 0.359 | 0.509 | 0.345 ** | 0.486 | 0.732 * | 0.633 | 0.500 | |
| MBC | 1 | 0.829 ** | −0.763 * | 0.375 | 0.302 | 0.424 ** | 0.430 | 0.760 * | 0.661 | 0.737 * | ||
| EOC | 1 | −0.904 ** | 0.606 | 0.336 | 0.711 ** | 0.581 | 0.870 ** | 0.557 | 0.730 * | |||
| BF | SOC | 0.929 ** | 0.853 ** | 0.949 ** | −0.598 | 0.961 ** | −0.695 * | 0.918 ** | 0.395 | 0.759 * | 0.866 ** | 0.298 |
| DOC | 1 | 0.835 ** | 0.890 ** | −0.711 * | 0.939 ** | −0.458 | 0.807 ** | 0.116 | 0.667 * | 0.783 * | 0.220 | |
| MBC | 1 | 0.948 ** | −0.543 | 0.874 ** | −0.701 * | 0.884 ** | 0.066 | 0.907 ** | 0.892 ** | 0.387 | ||
| EOC | 1 | −0.599 | 0.956 ** | −0.705 * | 0.968 ** | 0.268 | 0.899 ** | 0.954 ** | 0.449 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, Y.; Huang, Z.; Ling, Y.; Cai, S.; Zeng, B.; Liang, S.; Wang, X. Effects of Forest Vegetation Restoration on Soil Organic Carbon and Its Labile Fractions in the Danxia Landform of China. Sustainability 2022, 14, 12283. https://doi.org/10.3390/su141912283
Xiao Y, Huang Z, Ling Y, Cai S, Zeng B, Liang S, Wang X. Effects of Forest Vegetation Restoration on Soil Organic Carbon and Its Labile Fractions in the Danxia Landform of China. Sustainability. 2022; 14(19):12283. https://doi.org/10.3390/su141912283
Chicago/Turabian StyleXiao, Ye, Zhigang Huang, Yulin Ling, Shenwen Cai, Boping Zeng, Sheng Liang, and Xiao Wang. 2022. "Effects of Forest Vegetation Restoration on Soil Organic Carbon and Its Labile Fractions in the Danxia Landform of China" Sustainability 14, no. 19: 12283. https://doi.org/10.3390/su141912283