
Optimal Method for Biomass Estimation in a Cladoceran Species, Daphnia Magna (Straus, 1820): Evaluating Length–Weight Regression Equations and Deriving Estimation Equations Using Body Length, Width and Lateral Area

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Daphnia Magna as the Subject Species
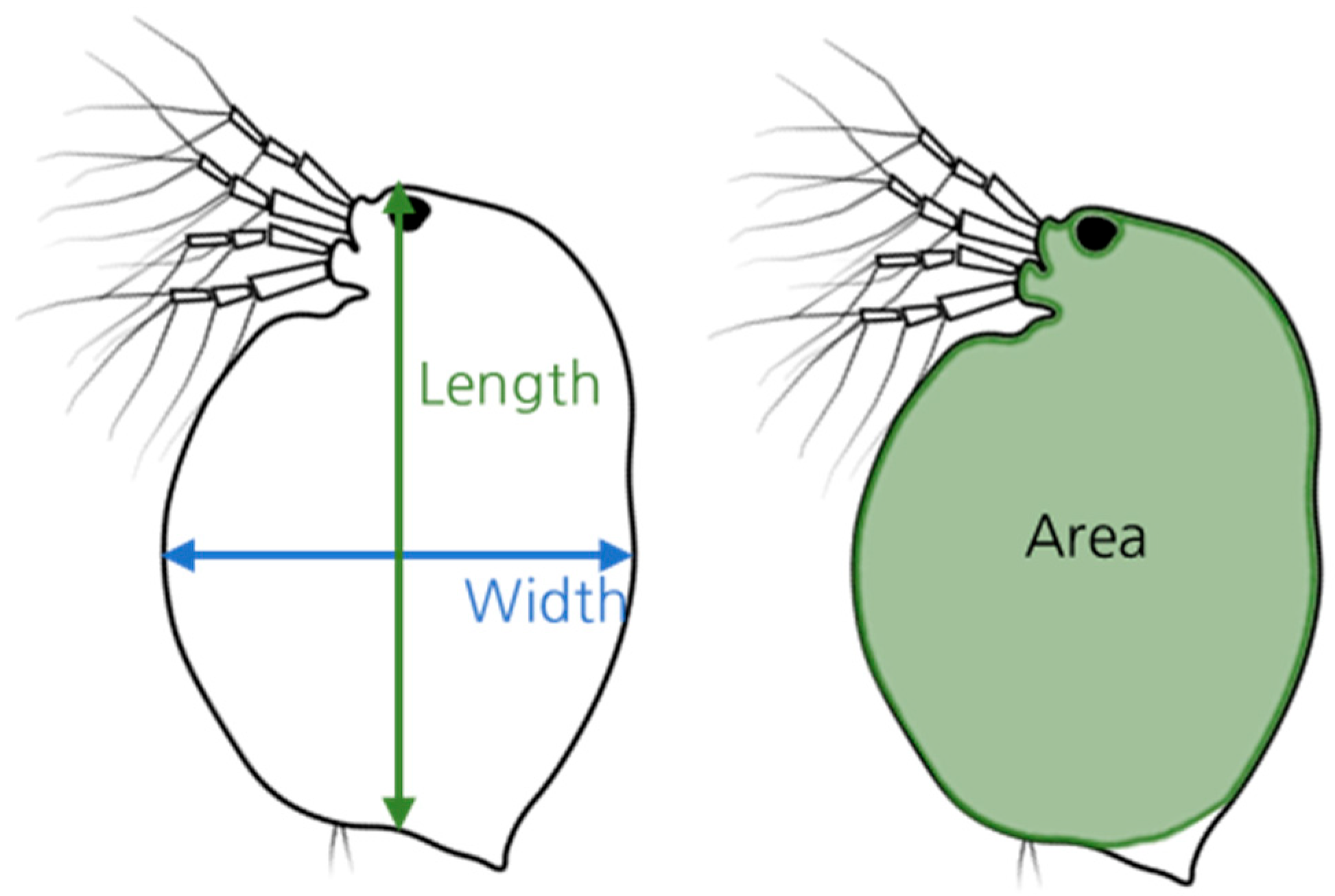
2.2. Estimation and Measurement of D. Magna Biomass
2.3. Statistical Analyses
3. Results
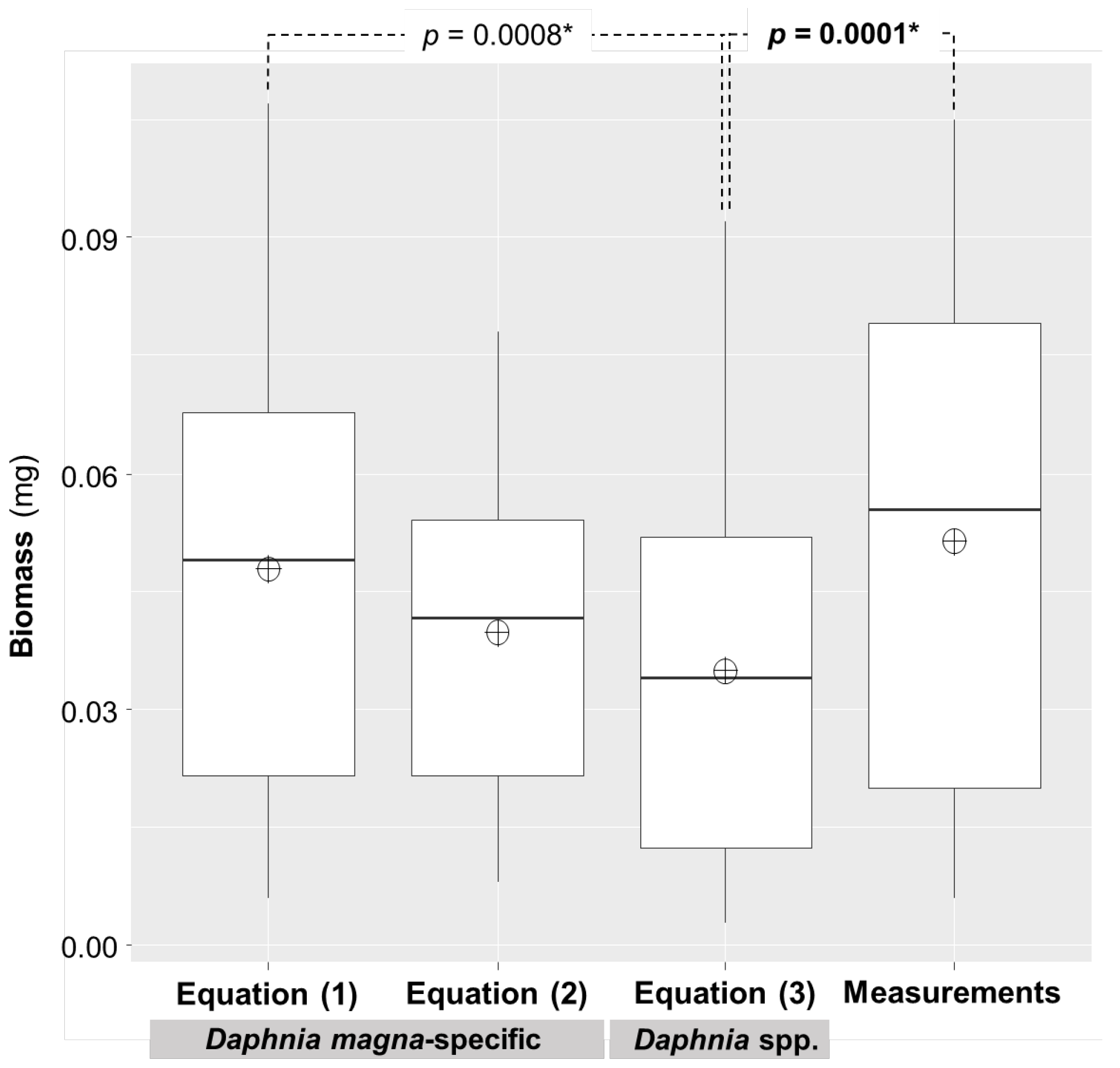
3.1. Comparison of D. Magna Biomass Estimates and Measurements
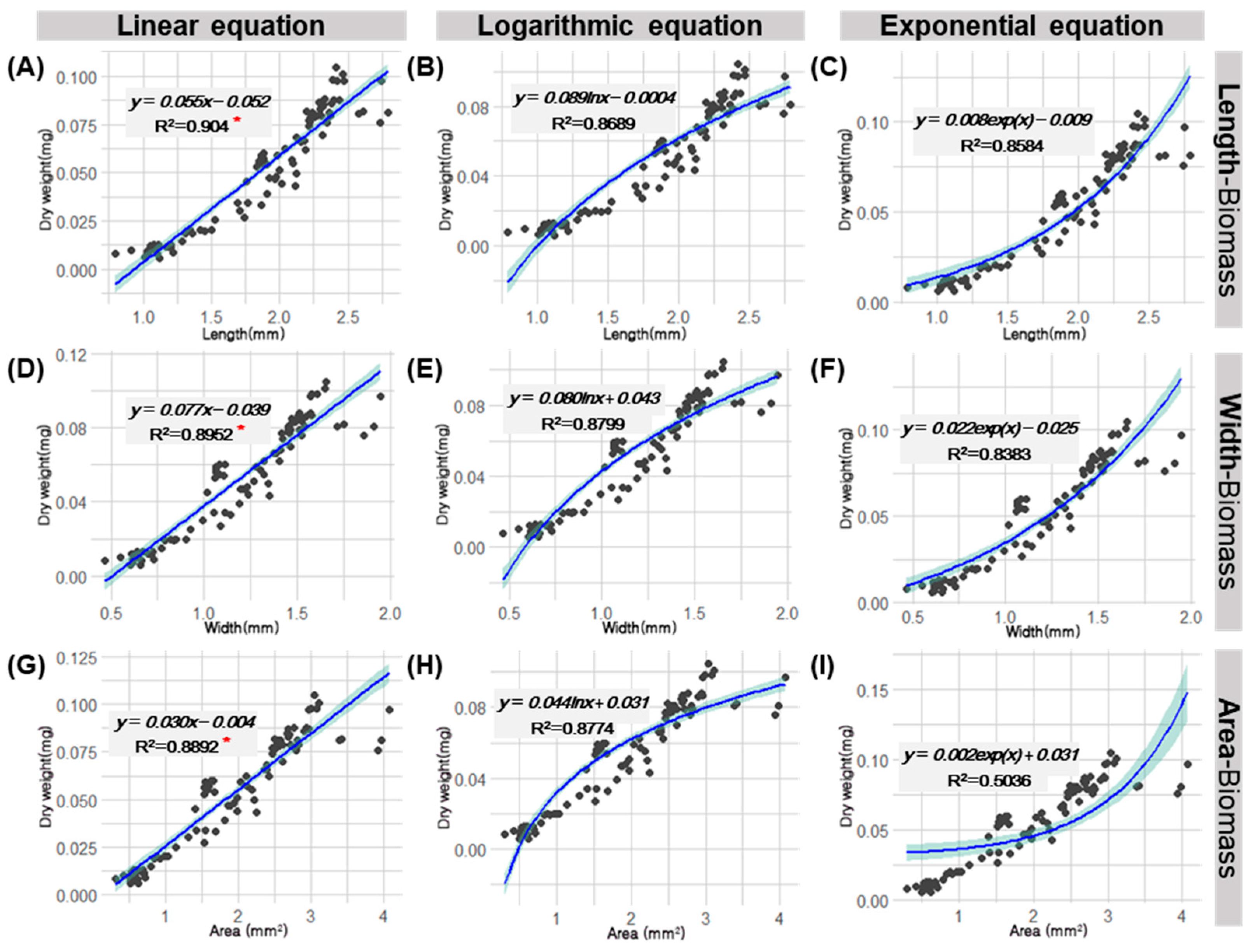
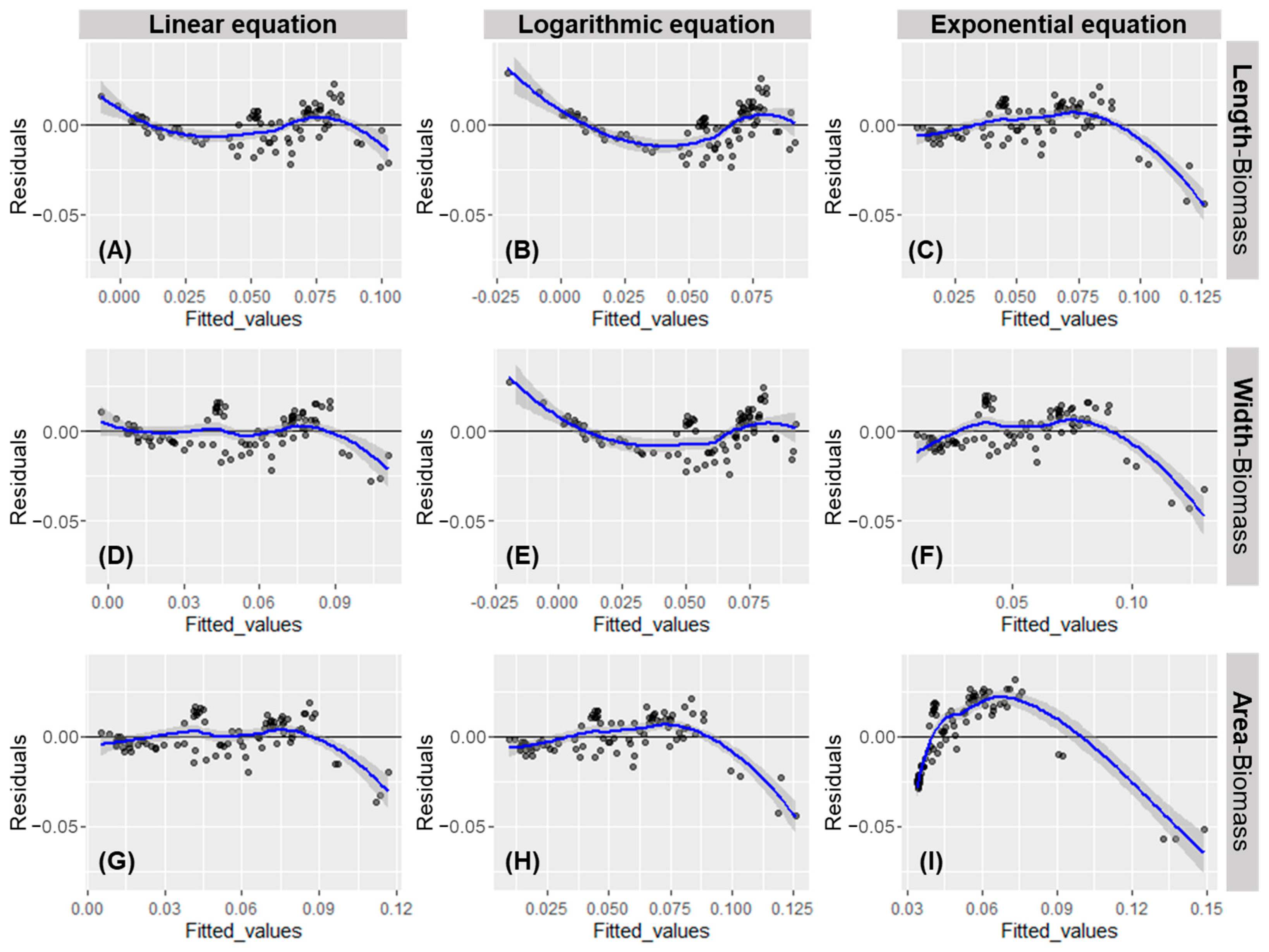
3.2. Derivation of Regression Equations Considering Body Specifications and Biomass Measurements of D. Magna, and a Comparison of the Suitability of the Derived Equations
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Choi, K.H.; Lee, C.R.; Kang, H.K.; Kang, K.A. Characteristics and variation of size-fractionated zooplankton biomass in the Northern East China sea. Ocean Polar Res. 2011, 33, 135–147, [Korean literature]. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Cui, J.; Song, T.; Liu, Y. Application of an AQUATOX model for direct toxic effects and indirect ecological effects assessment of Polycyclic aromatic hydrocarbons (PAHs) in a plateau eutrophication lake, China. Ecol. Model. 2018, 388, 31–44. [Google Scholar] [CrossRef]
- Oh, H.J.; Seo, D.H.; Choi, M.J.; Jeong, H.G.; Kim, H.W.; Oh, J.M.; Chang, K.H. Calculation and applicability of rotifers biomass (Polyarthra spp.) based on length-weight relationship and spatial distribution of body length. KJEE 2018, 51, 205–211, [Korean literature]. [Google Scholar] [CrossRef]
- US Environmental Protection Agency, USEPA. Modeling Environmental Fate and Ecological Effects in Aquatic Ecosystems; United States Environmental Protection Agency Office of Water: Washington, DC, USA, 2014; Volume 1, User’s manual, EPA-820-R-14-005. Available online: nepis.epa.gov (accessed on 17 May 2022).
- Kane, D.D.; Gordon, S.I.; Munawar, M.; Charlton, M.N.; Culver, D.A. The planktonic index of biotic integrity (P-IBI): An approach for assessing lake ecosystem health. Ecol. Indic. 2009, 9, 1234–1247. [Google Scholar] [CrossRef]
- Beaver, J.R.; Tausz, C.E.; Black, K.M.; Bolam, B.A. Cladoceran body size distributions along temperature and trophic gradients in the conterminous USA. J. Plankton Res. 2020, 42, 613–629. [Google Scholar] [CrossRef]
- Brucet, S.; Boix, D.; Quintana, X.D.; Jensen, E.; Nathansen, L.W.; Trochine, C.; Meerhoff, M.; Gasco´n, S.; Jeppesena, E. Factors influencing zooplankton size structure at contrasting temperatures in coastal shallow lakes: Implications for effects of climate change. Limnol. Oceanogr. 2020, 55, 1697–1711. [Google Scholar] [CrossRef] [Green Version]
- Stamou, G.; Katsiapi, M.; Moustaka-Gouni, M.; Michaloudi, E. Grazing potential—A functional plankton food web metric for ecological water quality assessment in Mediterranean lakes. Water 2019, 11, 1274. [Google Scholar] [CrossRef] [Green Version]
- Zhan, H.; Duan, Z.; Wang, Z.; Zhong, M.; Tian, W.; Wang, H.; Huang, H. Freshwater lake ecosystem health assessment and its response to pollution stresses based on planktonic index of biotic integrity. Environ. Sci. Pollut. Res. 2019, 26, 35240–35252. [Google Scholar] [CrossRef]
- Yao, N.; Feng, B.; Zhang, M.; He, L.; Zhang, H.; Liu, Z. Impact of industrial production, dam construction, and agriculture on the Z-IBI in river ecosystems: A case study of the Wanan River Basin in China. Water 2021, 12, 123. [Google Scholar] [CrossRef]
- Stamou, G.; Mazaris, A.D.; Moustaka-Gouni, M.; Spoljar, M.; Ternjej, I.; Drazina, T.; Dorak, Z.; Michaloudi, E. Introducing a zooplanktonic index for assessing water quality of natural lakes in the Mediterranean region. Ecol. Inform. 2022, 69, 101616. [Google Scholar] [CrossRef]
- Lombardo, A.; Franco, A.; Pivato, A.; Barauss, A. Food web modeling of a river ecosystem for risk assessment of down-the-drain chemicals: A case study with AQUATOX. Sci. Total Environ. 2015, 508, 214–227. [Google Scholar] [CrossRef]
- He, H.; Jeppesen, E.; Bruhn, D.; Yde, M.; Hansen, J.K.; Spanggaard, L.; Madsen, N.; Liu, W.; Søndergaard, M.; Lauridsen, T.L. Decadal changes in zooplankton biomass, composition, and body mass in four shallow brackish lakes in Denmark subjected to varying degrees of eutrophication. Inland Waters 2020, 10, 186–196. [Google Scholar] [CrossRef]
- Taipale, S.J.; Kuoppamäki, K.; Strandberg, U.; Peltomaa, E.; Vuorio, K. Lake restoration influences nutritional quality of algae and consequently Daphnia biomass. Hydrobiologia 2020, 847, 4539–4557. [Google Scholar] [CrossRef]
- Irigoien, X.; Huisman, J.; Harris, R.P. Global biodiversity patterns of marine phytoplankton and zooplankton. Nature 2004, 429, 863–867. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.J.; Chang, K.H.; Jeong, H.G.; Go, S.M.; La, G.H.; Kim, H.W. Quantitative zooplankton collection methods for various freshwater ecosystems and their applications. KJEE 2019, 52, 231–244, [Korean literature]. [Google Scholar] [CrossRef]
- Dumont, H.J.; Velde, I.V.D.; Dumont, S. The dry weight estimate of biomass in a selection of cladocera, Copepoda and Rotifera from the plankton, periphyton and benthos of continental waters. Oecologia 1975, 19, 75–97. [Google Scholar] [CrossRef]
- Park, R.A.; Clough, J.S.; Wellman, M.C. AQUATOX: Modeling environmental fate and ecological effects in aquatic ecosystems. Ecol. Model. 2008, 213, 1–15. [Google Scholar] [CrossRef]
- Cremona, F.; Blank, K.; Haberman, J. Effects of environmental stressors and their interactions on zooplankton biomass and abundance in a large eutrophic lake. Hydrobiologia 2021, 848, 4401–4418. [Google Scholar] [CrossRef]
- Long, S.X.; Hamilton, P.B.; Yang, Y.; Wang, S.; Chen, C.; Tao, R. Differential bioaccumulation of mercury by zooplankton taxa in a mercury-contaminated reservoir Guizhou China. Environ. Pollut. 2018, 239, 147–160. [Google Scholar] [CrossRef]
- Scheffer, M.; Rinaldi, S.; Kuznetsov, Y.A.; Nes, E.H.V. Seasonal dynamics of Daphnia and algae explained as a periodically forced predator-prey system. Oikos 1997, 80, 519–532. [Google Scholar] [CrossRef]
- Lampert, W.; Fleckner, W.; Hakumat, R.; Taylor, B.E. Phytoplankton control by grazing zooplankton: A study on the spring clear-water phase. Limnol. Oceanogr. 1986, 31, 478–490. [Google Scholar] [CrossRef]
- Wilson, S.M.; Corsi, M.P.; Brandt, D.H.; Stark, E.J. The response of Daphnia to nutrient additions and kokanee abundance in Dworshak Reservoir, Idaho. Can. J. Fish. Aquat. Sci. 2021, 78, 1677–1688. [Google Scholar] [CrossRef]
- Kawabata, K.; Urabe, J. Length-weight relationships of eight freshwater planktonic crustacean species in Japan. Freshw. Biol. 1998, 39, 199–205. [Google Scholar] [CrossRef]
- Færøvig, P.J.; Andersen, T.; Hessen, D.O. Image analysis of Daphnia populations: Non-destructive determination of demography and biomass in cultures. Freshw. Biol. 2002, 47, 1956–1962. [Google Scholar] [CrossRef]
- Pearre, S., Jr. The copepod width–weight relation and its utility in food chain research. Can. J. Zool. 1980, 58, 1884–1891. [Google Scholar] [CrossRef]
- Sperfeld, E.; Nilssen, J.P.; Rinehart, S.; Schwenk, K.; Hessen, D.O. Ecology of predator-induced morphological defense traits in Daphnia longispina (Cladocera, Arthropoda). Oecologia 2020, 192, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Burgess, S.; Jackson, E.W.; Schwarzman, L.; Gezon, N.; Lehman, J.T. Improved estimates of calanoid copepod biomass in the St. Lawrence Great Lakes. J. Great Lakes Res. 2015, 41, 484–491. [Google Scholar] [CrossRef]
- Maia-Barbosa, P.M.; Bozelli, R.L. Length-weight relationships for five cladoceran species in an Amazonian lake. Braz. Arch. Biol. Technol. 2005, 48, 303–308. [Google Scholar] [CrossRef]
- Vakkilainen, K.; Kairesalo, T.; Hietala, J.; Balayla, D.M.; Bécares, E.; Van de Bund, W.J.; Van Donk, E.; Fernández-Aláez, M.; Gyllström, M.; Hansson, L.-A.; et al. Response of zooplankton to nutrient enrichment and fish in shallow lakes: A pan-European mesocosm experiment. Freshw. Biol. 2004, 49, 1619–1632. [Google Scholar] [CrossRef] [Green Version]
- Alcaraz, M.; Saiz, E.; Calbet, A.; Trepat, I.; Broglio, E. Estimating zooplankton biomass through image analysis. Mar. Biol. 2003, 143, 307–315. [Google Scholar] [CrossRef]
- OECD. Daphnia magna reproduction test. In OECD Guidelines for Testing of Chemicals, 211; OECD: Paris, France, 1998. [Google Scholar] [CrossRef] [Green Version]
- McCauley, E. The estimation of the abundance and biomass of zooplankton in samples. In A Manual on Methods for the Assessment of Secondary Productivity in Fresh Waters, 2nd ed.; Downing, J.A., Rigler, F.H., Eds.; Blackwell: Oxford, UK, 1984; pp. 228–265. [Google Scholar]
- Lawrence, S.G.; Malley, D.F.; Findlay, W.J.; MacIver, M.A.; Delbaere, I.L. Method for estimating dry weight of freshwater planktonic crustaceans from measures of length and shape. Can. J. Fish. Aquat. Sci. 1987, 44, 264–274. [Google Scholar] [CrossRef]
- USEPA. National Lakes Assessment 2017. In Laboratory Operations Manual; EPA-841-B11-004; US Environmental Protection Agency: Washington, DC, USA, 2017. Available online: www.epa.gov/national-aquatic-resource-surveys/national-lakes-assessment-2017-laboratory-operations-manual (accessed on 17 May 2022).
- GLNPO. LG403, Revision 07; Standard Operating Procedure for Zooplankton Analysis.; U.S. EPA Great Lakes National Program Office: Chicago, IL, USA, 2016. Available online: www.epa.gov/sites/default/files/2017-01/documents/sop-for-zooplankton-analysis-201607-22pp.pdf (accessed on 17 May 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: www.R-project.org (accessed on 17 May 2022).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Rudis, B. Hrbrthemes: Additional Themes, Theme Components and Utilities for ‘ggplot2’. R Package Version 0.8.0. Available online: CRAN.R-project.org/package=hrbrthemes (accessed on 17 May 2022).
- Dodson, S.I. Cyclomorphosis in Daphnia galeata mendotae Birge and D. retrocurva Forbes as a predator-induced response. Freshw. Biol. 1988, 19, 109–114. [Google Scholar] [CrossRef]
- Spaak, P.; Boersma, M. Tail spine length in the Daphnia galeata complex: Costs and benefits of induction by fish. Aquat. Ecol. 1997, 31, 89–98. [Google Scholar] [CrossRef]
- Smakulska, J.; Górniak, A. Morphological variation in Daphnia cucullata Sars with progressive eutrophication of a polymictic lowland reservoir. Hydrobiologia 2004, 526, 119–127. [Google Scholar] [CrossRef]
- Chang, K.H.; Hanazato, T. Seasonal and reciprocal succession and cyclomorphosis of two Bosmina species (Cladocera, Crustacea) co-existing in a lake: Their relationship with invertebrate predators. J. Plankton Res. 2003, 25, 141–150. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Equations | Characteristics | Unit | |
|---|---|---|---|
| (1) | Derived using individuals from Donk Lake | W: μg L: μm | |
| (2) | Derived using individuals from Sambre River | W: μg L: μm | |
| (3) | Derived using pooled Daphnia species | W: μg L: mm | |
| Biomass (dry weight, mg) | Equation (1) | Equation (2) | Equation (3)1 | Measurements |
| D. magna in Donk Lake | D. magna in Sambre River | Pooled Daphnia spp. | ||
| Mean ± Standard deviation | 0.048 ± 0.026 | 0.040 ± 0.018 | 0.035 ± 0.022 | 0.051 ± 0.030 |
| Body Specifications | Length (mm) | Width (mm) | Area (mm2) |
|---|---|---|---|
| Mean ± standard deviation (Minimum–Maximum) | 1.866 ± 0.519 (0.796–2.791) | 1.176 ± 0.372 (0.469–1.948) | 1.872 ± 0.962 (0.314–4.073) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ku, D.; Chae, Y.-J.; Choi, Y.; Ji, C.W.; Park, Y.-S.; Kwak, I.-S.; Kim, Y.-J.; Chang, K.-H.; Oh, H.-J. Optimal Method for Biomass Estimation in a Cladoceran Species, Daphnia Magna (Straus, 1820): Evaluating Length–Weight Regression Equations and Deriving Estimation Equations Using Body Length, Width and Lateral Area. Sustainability 2022, 14, 9216. https://doi.org/10.3390/su14159216
Ku D, Chae Y-J, Choi Y, Ji CW, Park Y-S, Kwak I-S, Kim Y-J, Chang K-H, Oh H-J. Optimal Method for Biomass Estimation in a Cladoceran Species, Daphnia Magna (Straus, 1820): Evaluating Length–Weight Regression Equations and Deriving Estimation Equations Using Body Length, Width and Lateral Area. Sustainability. 2022; 14(15):9216. https://doi.org/10.3390/su14159216
Chicago/Turabian StyleKu, Doyeong, Yeon-Ji Chae, Yerim Choi, Chang Woo Ji, Young-Seuk Park, Ihn-Sil Kwak, Yong-Jae Kim, Kwang-Hyeon Chang, and Hye-Ji Oh. 2022. "Optimal Method for Biomass Estimation in a Cladoceran Species, Daphnia Magna (Straus, 1820): Evaluating Length–Weight Regression Equations and Deriving Estimation Equations Using Body Length, Width and Lateral Area" Sustainability 14, no. 15: 9216. https://doi.org/10.3390/su14159216