
Tree Species Diversity and Stand Attributes Differently Influence the Ecosystem Functions of Pinus yunnanensis Secondary Forests under the Climate Context



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Forests
2.2. Quantification of Forest Single Functions and Multifunctionality
2.3. Quantification of Forest Diversity Attributes
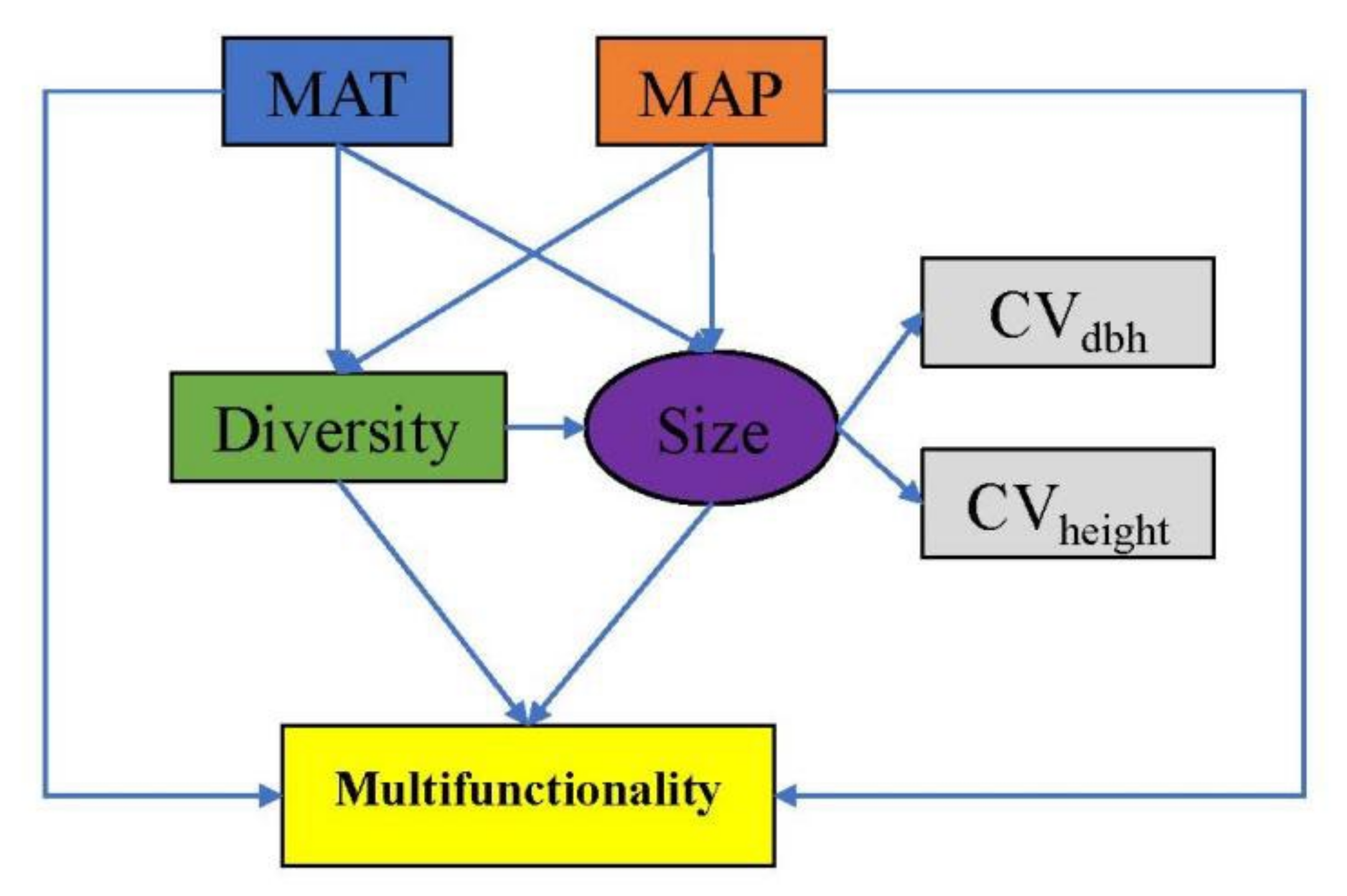
2.4. Statistical Analyses
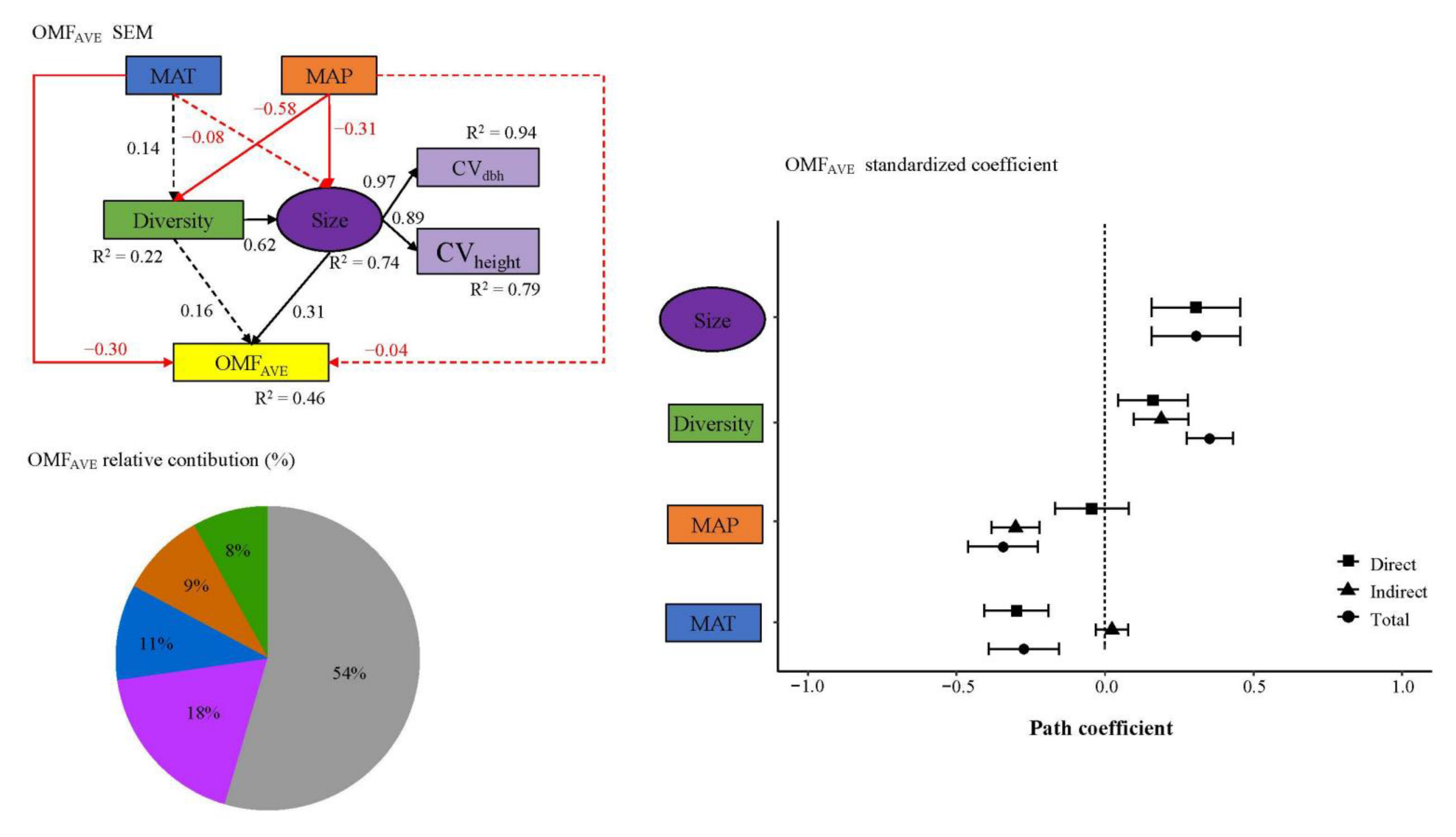
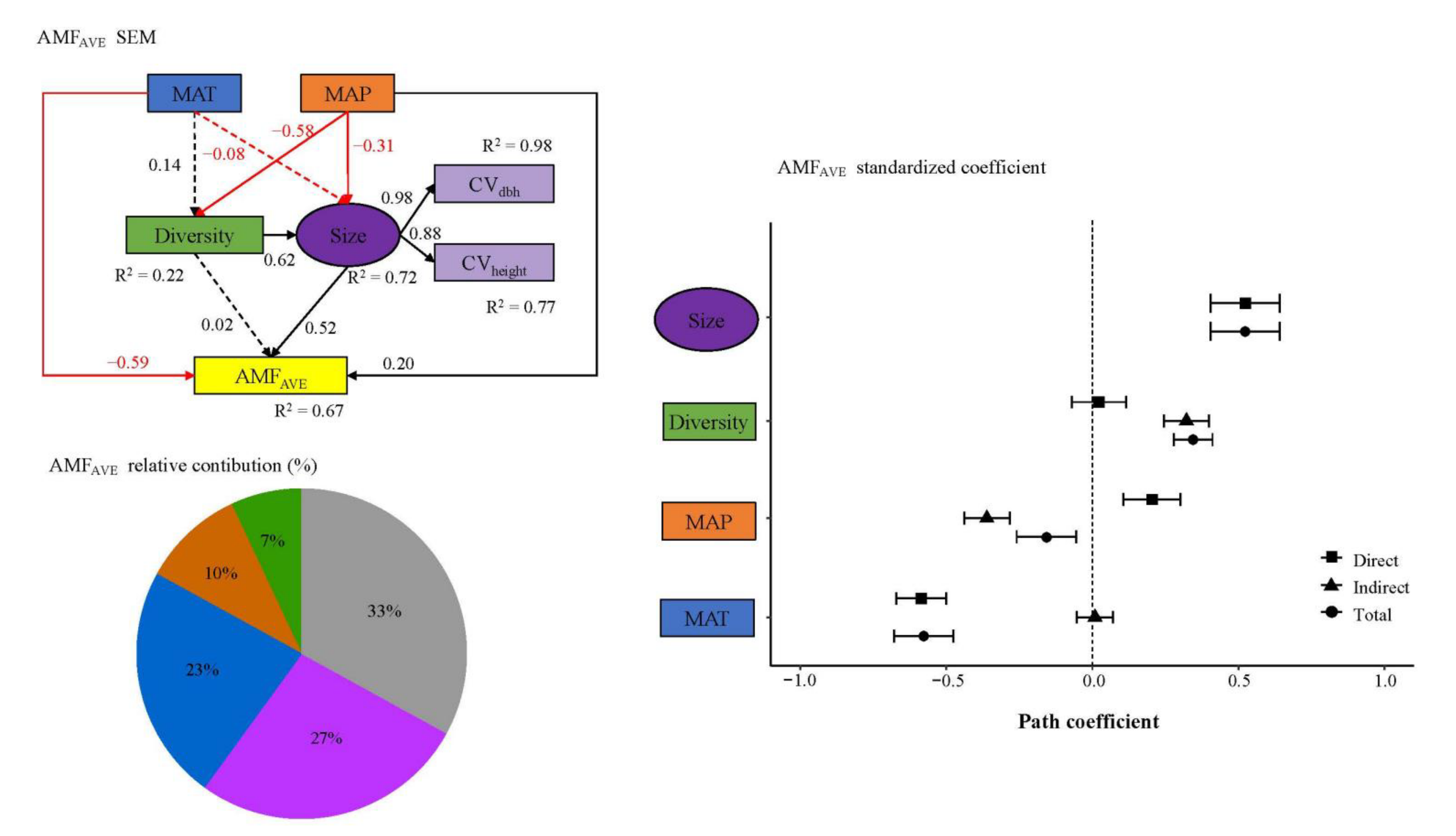
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Gamfeldt, L.; Snall, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Froberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed]
- Newbold, T.; Hudson, L.N.; Arnell, A.P.; Contu, S.; De Palma, A.; Ferrier, S.; Hill, S.L.L.; Hoskins, A.J.; Lysenko, I.; Phillips, H.R.P.; et al. Has land use pushed terrestrial biodiversity beyond the planetary boundary? A global assessment. Science 2016, 353, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Le Provost, G.; Badenhausser, I.; Le Bagousse-Pinguet, Y.; Clough, Y.; Henckel, L.; Violle, C.; Bretagnolle, V.; Roncoroni, M.; Manning, P.; Gross, N. Land-use history impacts functional diversity across multiple trophic groups. Proc. Natl. Acad. Sci. USA 2020, 117, 1573–1579. [Google Scholar] [CrossRef] [PubMed]
- Naeem, S.; Duffy, J.E.; Zavaleta, E. The Functions of Biological Diversity in an Age of Extinction. Science 2012, 336, 1401–1406. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Downing, J.A. Biodiversity and stability in graslands. Nature 1994, 367, 363–365. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Srivastava, D.S.; Duffy, J.E.; Wright, J.P.; Downing, A.L.; Sankaran, M.; Jouseau, C. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 2006, 443, 989–992. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, Y.; Castro-Izaguirre, N.; Baruffol, M.; Brezzi, M.; Lang, A.; Li, Y.; Haerdtle, W.; von Oheimb, G.; Yang, X.; et al. Impacts of species richness on productivity in a large-scale subtropical forest experiment. Science 2018, 362, 80. [Google Scholar] [CrossRef] [Green Version]
- Van der Plas, F. Biodiversity and ecosystem functioning in naturally assembled communities. Biol. Rev. 2019, 94, 1220–1245. [Google Scholar] [CrossRef]
- Liang, J.J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, aaf8957. [Google Scholar] [CrossRef] [Green Version]
- Hector, A.; Bagchi, R. Biodiversity and ecosystem multifunctionality. Nature 2007, 448, 188-U6. [Google Scholar] [CrossRef]
- Vila, M.; Inchausti, P.; Vayreda, J.; Barrantes, O.; Gracia, C.; Ibanez, J.J.; Mata, T. Confounding Factors in the Observational Productivity-Diversity Relationship in Forests. In Proceedings of the LINKECOL Workshop, Weimar, Germany, 13–15 June 2002; pp. 65–86. [Google Scholar]
- Ahlstrom, A.; Raupach, M.R.; Schurgers, G.; Smith, B.; Arneth, A.; Jung, M.; Reichstein, M.; Canadell, J.G.; Friedlingstein, P.; Jain, A.K.; et al. The dominant role of semi-arid ecosystems in the trend and variability of the land CO2 sink. Science 2015, 348, 895–899. [Google Scholar] [CrossRef] [Green Version]
- Byrnes, J.E.K.; Gamfeldt, L.; Isbell, F.; Lefcheck, J.S.; Griffin, J.N.; Hector, A.; Cardinale, B.J.; Hooper, D.U.; Dee, L.E.; Duffy, J.E. Investigating the relationship between biodiversity and ecosystem multifunctionality: Challenges and solutions. Methods Ecol. Evol. 2014, 5, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Felipe-Lucia, M.R.; Soliveres, S.; Penone, C.; Manning, P.; van der Plas, F.; Boch, S.; Prati, D.; Ammer, C.; Schall, P.; Gossner, M.M.; et al. Multiple forest attributes underpin the supply of multiple ecosystem services. Nat. Commun. 2018, 9, 4839. [Google Scholar] [CrossRef] [Green Version]
- De Avila, A.L.; van der Sande, M.T.; Dormann, C.F.; Pena-Claros, M.; Poorter, L.; Mazzei, L.; Ruschel, A.R.; Silva, J.N.M.; de Carvalho, J.O.P.; Bauhus, J. Disturbance intensity is a stronger driver of biomass recovery than remaining tree-community attributes in a managed Amazonian forest. J. Appl. Ecol. 2018, 55, 1647–1657. [Google Scholar] [CrossRef] [Green Version]
- Piponiot, C.; Sist, P.; Mazzei, L.; Pena-Claros, M.; Pute, F.E.; Rutishauser, E.; Shenkin, A.; Ascarrunz, N.; de Azevedo, C.P.; Baraloto, C.; et al. Carbon recovery dynamics following disturbance by selective logging in Amazonian forests. eLife 2016, 5, e21394. [Google Scholar] [CrossRef]
- Diaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Prentice, I.C.; et al. The global spectrum of plant form and function. Nature 2016, 529, 167. [Google Scholar] [CrossRef]
- Poorter, L.; van der Sande, M.T.; Thompson, J.; Arets, E.; Alarcon, A.; Alvarez-Sanchez, J.; Ascarrunz, N.; Balvanera, P.; Barajas-Guzman, G.; Boit, A.; et al. Diversity enhances carbon storage in tropical forests. Glob. Ecol. Biogeogr. 2015, 24, 1314–1328. [Google Scholar] [CrossRef]
- Poorter, L.; van der Sande, M.T.; Arets, E.; Ascarrunz, N.; Enquist, B.; Finegan, B.; Licona, J.C.; Martinez-Ramos, M.; Mazzei, L.; Meave, J.A.; et al. Biodiversity and climate determine the functioning of Neotropical forests. Glob. Ecol. Biogeogr. 2017, 26, 1423–1434. [Google Scholar] [CrossRef]
- Wright, A.J.; Wardle, D.A.; Callaway, R.; Gaxiola, A. The Overlooked Role of Facilitation in Biodiversity Experiments. Trends Ecol. Evol. 2017, 32, 383–390. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Does complementary resource use enhance ecosystem functioning? A model of light competition in plant communities. Ecol. Lett. 2007, 10, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Yan, E.-R.; Chen, H.Y.H.; Chang, S.X.; Zhao, Y.-T.; Yang, X.-D.; Xu, M.-S. Stand structural diversity rather than species diversity enhances aboveground carbon storage in secondary subtropical forests in Eastern China. Biogeosciences 2016, 13, 4627–4635. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.F.; Lei, X.D.; Ma, Z.H.; Kneeshaw, D.D.; Peng, C.H. Positive Relationship between Aboveground Carbon Stocks and Structural Diversity in Spruce-Dominated Forest Stands in New Brunswick, Canada. For. Sci. 2011, 57, 506–515. [Google Scholar]
- Zhang, Y.; Chen, H.Y.H. Individual size inequality links forest diversity and above-ground biomass. J. Ecol. 2015, 103, 1245–1252. [Google Scholar] [CrossRef]
- Yuan, Z.Q.; Wang, S.P.; Ali, A.; Gazol, A.; Ruiz-Benito, P.; Wang, X.G.; Lin, F.; Ye, J.; Hao, Z.Q.; Loreau, M. Aboveground carbon storage is driven by functional trait composition and stand structural attributes rather than biodiversity in temperate mixed forests recovering from disturbances. Ann. For. Sci. 2018, 75, 67. [Google Scholar] [CrossRef] [Green Version]
- Ali, A. Forest stand structure and functioning: Current knowledge and future challenges. Ecol. Indic. 2019, 98, 665–677. [Google Scholar] [CrossRef]
- Weiher, E.; Keddy, P.A. Ecological Assembly Rules: Perspectives, Advances, Retreats; Cambridge University Press: Cambridge, UK, 1999. [Google Scholar]
- Ali, A.; Wang, L.Q. Big-sized trees and forest functioning: Current knowledge and future perspectives. Ecol. Indic. 2021, 127, 107760. [Google Scholar] [CrossRef]
- Ratcliffe, S.; Wirth, C.; Jucker, T.; van der Plas, F.; Scherer-Lorenzen, M.; Verheyen, K.; Allan, E.; Benavides, R.; Bruelheide, H.; Ohse, B.; et al. Biodiversity and ecosystem functioning relations in European forests depend on environmental context. Ecol. Lett. 2017, 20, 1414–1426. [Google Scholar] [CrossRef]
- Pan, Y.D.; Birdsey, R.A.; Fang, J.Y.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [Green Version]
- Larjavaara, M.; Muller-Landau, H.C. Temperature explains global variation in biomass among humid old-growth forests. Glob. Ecol. Biogeogr. 2012, 21, 998–1006. [Google Scholar] [CrossRef]
- Malhi, Y.; Wood, D.; Baker, T.R.; Wright, J.; Phillips, O.L.; Cochrane, T.; Meir, P.; Chave, J.; Almeida, S.; Arroyo, L.; et al. The regional variation of aboveground live biomass in old-growth Amazonian forests. Glob. Change Biol. 2006, 12, 1107–1138. [Google Scholar] [CrossRef]
- Jin, Z.; Peng, J. Pinus yunnanensis; Yunnan Science and Technology Press: Kunming, China, 2004. [Google Scholar]
- Yuan, C.; Lang, N.; Men, G.; Fang, X.; Li, G.; Wen, S.; Zheng, K. Ecological Benefit of Soil and Water Conservation of Pinus yunnanensis Forests in the Upper Reach of Yangtze River. J. Soil Water Conserv. 2002, 16, 87–90. [Google Scholar]
- Shen, J.; Li, S.; Huang, X.; Wang, S.; Su, J. Ecological Resilience and Growth Degradation of Pinus yunnanensis at Different Altitudes in Jinsha River Basin. Sci. Silvae Sin. 2020, 56, 1–11. [Google Scholar]
- Huang, X.; Su, J.; Li, S.; Liu, W.; Lang, X. Functional diversity drives ecosystem multifunctionality in a Pinus yunnanensis natural secondary forest. Sci. Rep. 2019, 9, 6979. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Ali, A.; Loreau, M.; Ding, F.; Liu, S.; Sanaei, A.; Zhou, W.; Ye, J.; Lin, F.; Fang, S.; et al. Divergent above- and below-ground biodiversity pathways mediate disturbance impacts on temperate forest multifunctionality. Glob. Change Biol. 2021, 27, 2883–2894. [Google Scholar] [CrossRef]
- Fry, E.L.; Savage, J.; Hall, A.L.; Oakley, S.; Pritchard, W.J.; Ostle, N.J.; Pywell, R.F.; Bullock, J.M.; Bardgett, R.D. Soil multifunctionality and drought resistance are determined by plant structural traits in restoring grassland. Ecology 2018, 99, 2260–2271. [Google Scholar] [CrossRef] [Green Version]
- Maestre, F.T.; Quero, J.L.; Gotelli, N.J.; Escudero, A.; Ochoa, V.; Delgado-Baquerizo, M.; Garcia-Gomez, M.; Bowker, M.A.; Soliveres, S.; Escolar, C.; et al. Plant Species Richness and Ecosystem Multifunctionality in Global Drylands. Science 2012, 335, 214–218. [Google Scholar] [CrossRef] [Green Version]
- Isbell, F.; Calcagno, V.; Hector, A.; Connolly, J.; Harpole, W.S.; Reich, P.B.; Scherer-Lorenzen, M.; Schmid, B.; Tilman, D.; van Ruijven, J.; et al. High plant diversity is needed to maintain ecosystem services. Nature 2011, 477, 199–202. [Google Scholar] [CrossRef]
- Yuan, Z.Q.; Ali, A.; Ruiz-Benito, P.; Jucker, T.; Mori, A.S.; Wang, S.P.; Zhang, X.K.; Li, H.; Hao, Z.Q.; Wang, X.G.; et al. Above-and below-ground biodiversity jointly regulate temperate forest multifunctionality along a local-scale environmental gradient. J. Ecol. 2020, 108, 2012–2024. [Google Scholar] [CrossRef]
- Chave, J.; Rejou-Mechain, M.; Burquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Change Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Wang, G.Y.; Wang, T.L.; Kang, H.J.; Mang, S.; Riehl, B.; Seely, B.; Liu, S.R.; Guo, F.T.; Li, Q.L.; Innes, J.L. Adaptation of Asia-Pacific forests to climate change. J. For. Res. 2016, 27, 469–488. [Google Scholar] [CrossRef]
- Rosseel, Y. lavaan: An R Package for Structural Equation Modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Malaeb, Z.A.; Summers, J.K.; Pugesek, B.H. Using structural equation modeling to investigate relationships among ecological variables. Environ. Ecol. Stat. 2000, 7, 93–111. [Google Scholar] [CrossRef]
- Chisholm, R.A.; Muller-Landau, H.C.; Abdul Rahman, K.; Bebber, D.P.; Bin, Y.; Bohlman, S.A.; Bourg, N.A.; Brinks, J.; Bunyavejchewin, S.; Butt, N.; et al. Scale-dependent relationships between tree species richness and ecosystem function in forests. J. Ecol. 2013, 101, 1214–1224. [Google Scholar] [CrossRef]
- Schuldt, A.; Assmann, T.; Brezzi, M.; Buscot, F.; Eichenberg, D.; Gutknecht, J.; Hardtle, W.; He, J.S.; Klein, A.M.; Kuhn, P.; et al. Biodiversity across trophic levels drives multifunctionality in highly diverse forests. Nat. Commun. 2018, 9, 2989. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.Q.; Wang, S.P.; Gazol, A.; Mellard, J.; Lin, F.; Ye, J.; Hao, Z.Q.; Wang, X.G.; Loreau, M. Multiple metrics of diversity have different effects on temperate forest functioning over succession. Oecologia 2016, 182, 1175–1185. [Google Scholar] [CrossRef] [Green Version]
- Richards, A.E.; Forrester, D.I.; Bauhus, J.; Scherer-Lorenzen, M. The influence of mixed tree plantations on the nutrition of individual species: A review. Tree Physiol. 2010, 30, 1192–1208. [Google Scholar] [CrossRef] [Green Version]
- Wolters, V.; Silver, W.L.; Bignell, D.E.; Coleman, D.C.; Lavelle, P.; Van der Putten, W.H.; De Ruiter, P.; Rusek, J.; Wall, D.H.; Wardle, D.A.; et al. Effects of global changes on above- and belowground biodiversity in terrestrial ecosystems: Implications for ecosystem functioning. Bioscience 2000, 50, 1089–1098. [Google Scholar] [CrossRef] [Green Version]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setala, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef]
- Maestre, F.T.; Delgado-Baquerizo, M.; Jeffries, T.C.; Eldridge, D.J.; Ochoa, V.; Gozalo, B.; Luis Quero, J.; Garcia-Gomez, M.; Gallardo, A.; Ulrich, W.; et al. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proc. Natl. Acad. Sci. USA 2015, 112, 15684–15689. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Wu, J.-X.; Luo, L.-M.; Tu, Y.-L.; Yu, C.-Q.; Zhang, X.-Z.; Miao, Y.-J.; Zhao, Y.; Yang, J.-L. Climatic and geographic factors affect ecosystem multifunctionality through biodiversity in the Tibetan alpine grasslands. J. Mt. Sci. 2017, 14, 1604–1614. [Google Scholar] [CrossRef]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- van der Sande, M.T.; Poorter, L.; Kooistra, L.; Balvanera, P.; Thonicke, K.; Thompson, J.; Arets, E.; Alaniz, N.G.; Jones, L.; Mora, F.; et al. Biodiversity in species, traits, and structure determines carbon stocks and uptake in tropical forests. Biotropica 2017, 49, 593–603. [Google Scholar] [CrossRef]
- Ali, A.; Lin, S.L.; He, J.K.; Kong, F.M.; Yu, J.H.; Jiang, H.S. Climate and soils determine aboveground biomass indirectly via species diversity and stand structural complexity in tropical forests. For. Ecol. Manag. 2019, 432, 823–831. [Google Scholar] [CrossRef]
- Michaletz, S.T.; Kerkhoff, A.J.; Enquist, B.J. Drivers of terrestrial plant production across broad geographical gradients. Glob. Ecol. Biogeogr. 2018, 27, 166–174. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Name (County) | Latitude | Longitude | No. of Plots |
|---|---|---|---|
| Qiubei (QB) | 24°04′41.58″–24°13′45.59″ | 104°05′28.22″–104°29′52.11″ | 30 |
| Shuangbai (SB) | 24°28′03.02″–24°40′15.67″ | 101°32′24.20″–101°35′20.15″ | 36 |
| Shidian (SD) | 24°46′26.08″–24°51′13.01″ | 99°07′55.71″–99°09′08.53″ | 28 |
| Yunlong (YL) | 25°51′15.28″–25°52′38.10″ | 99°15′49.77″–99°17′15.89″ | 28 |
| Variables | Mean | 95% Confidence Intervals |
|---|---|---|
| MAT | 15.7 °C | 15.3 °C–16.1 °C |
| MAP | 930 mm | 904 mm–956 mm |
| CVdbh | 0.69 | 0.65–0.74 |
| CVheight | 0.53 | 0.50–0.57 |
| Diversity | 6.3 | 5.6–7.0 |
| OMF | 0.61 | 0.59–0.62 |
| AMF | 0.72 | 0.70–0.73 |
| BMF | 0.68 | 0.66–0.70 |
| EMF 25% | 6.33 | 6.22–6.43 |
| EMF 50% | 5.30 | 5.17–5.44 |
| EMF 75% | 3.17 | 2.92–2.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Huang, X.; Su, J. Tree Species Diversity and Stand Attributes Differently Influence the Ecosystem Functions of Pinus yunnanensis Secondary Forests under the Climate Context. Sustainability 2022, 14, 8332. https://doi.org/10.3390/su14148332
Wang L, Huang X, Su J. Tree Species Diversity and Stand Attributes Differently Influence the Ecosystem Functions of Pinus yunnanensis Secondary Forests under the Climate Context. Sustainability. 2022; 14(14):8332. https://doi.org/10.3390/su14148332
Chicago/Turabian StyleWang, Lei, Xiaobo Huang, and Jianrong Su. 2022. "Tree Species Diversity and Stand Attributes Differently Influence the Ecosystem Functions of Pinus yunnanensis Secondary Forests under the Climate Context" Sustainability 14, no. 14: 8332. https://doi.org/10.3390/su14148332