
Valorization of Agricultural Side-Streams for the Rearing of Larvae of the Lesser Mealworm, Alphitobius diaperinus (Panzer)




Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Side-Streams
2.3. Bioassay I: Single Side-Streams
2.4. Bioassay II: Side-Streams-Based Isoproteinic Diets
2.5. Statistical Analysis
3. Results
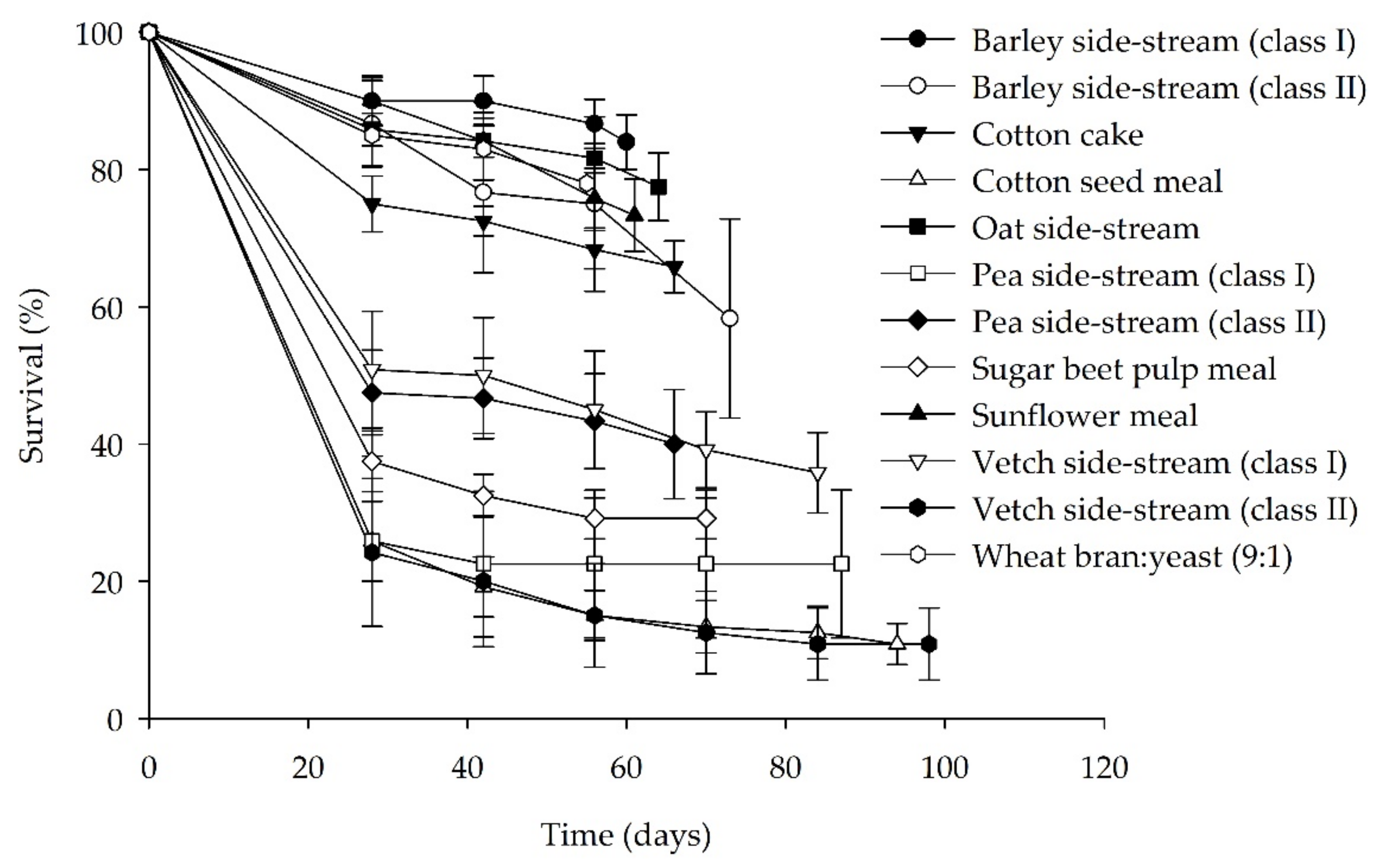
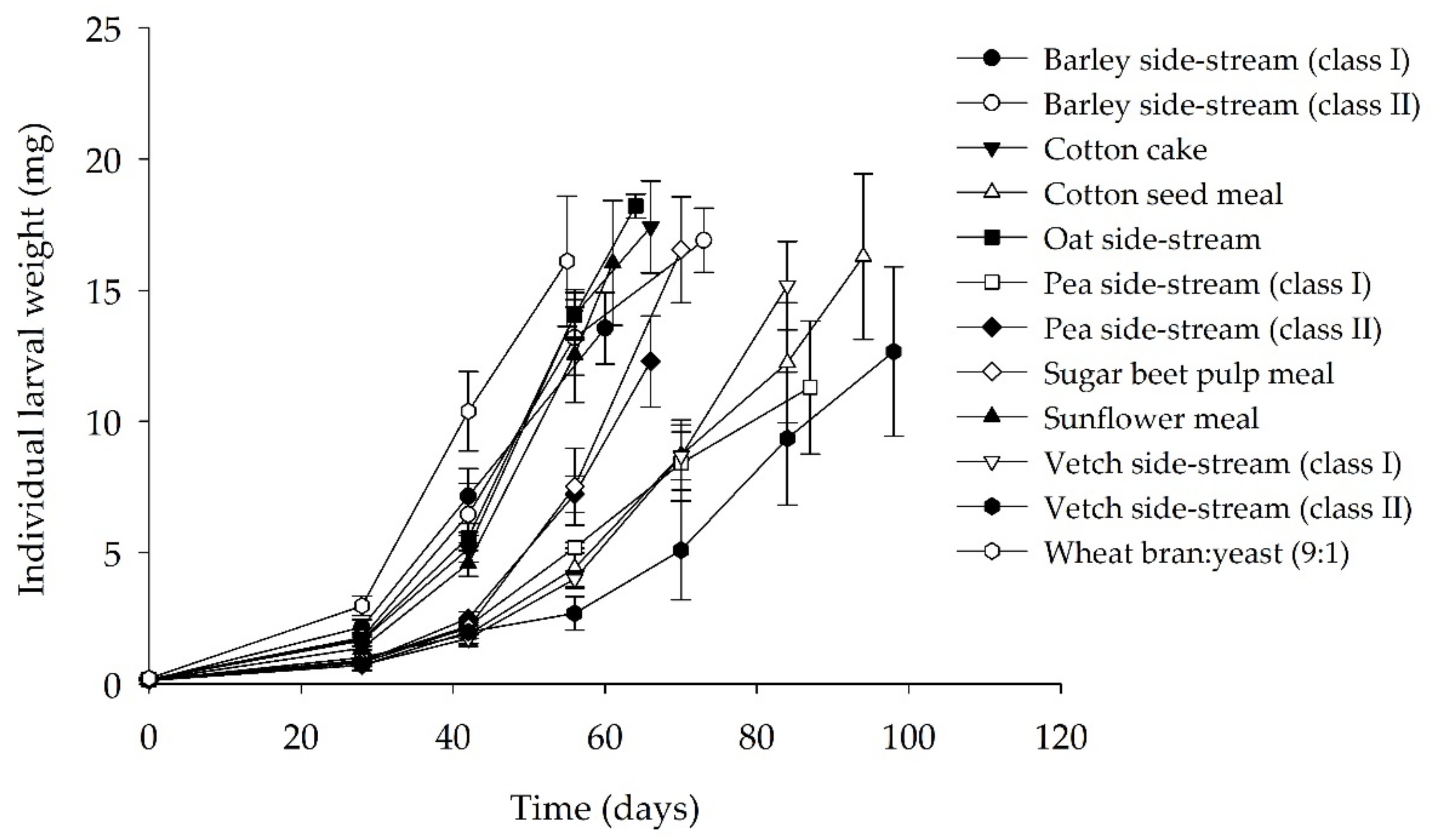
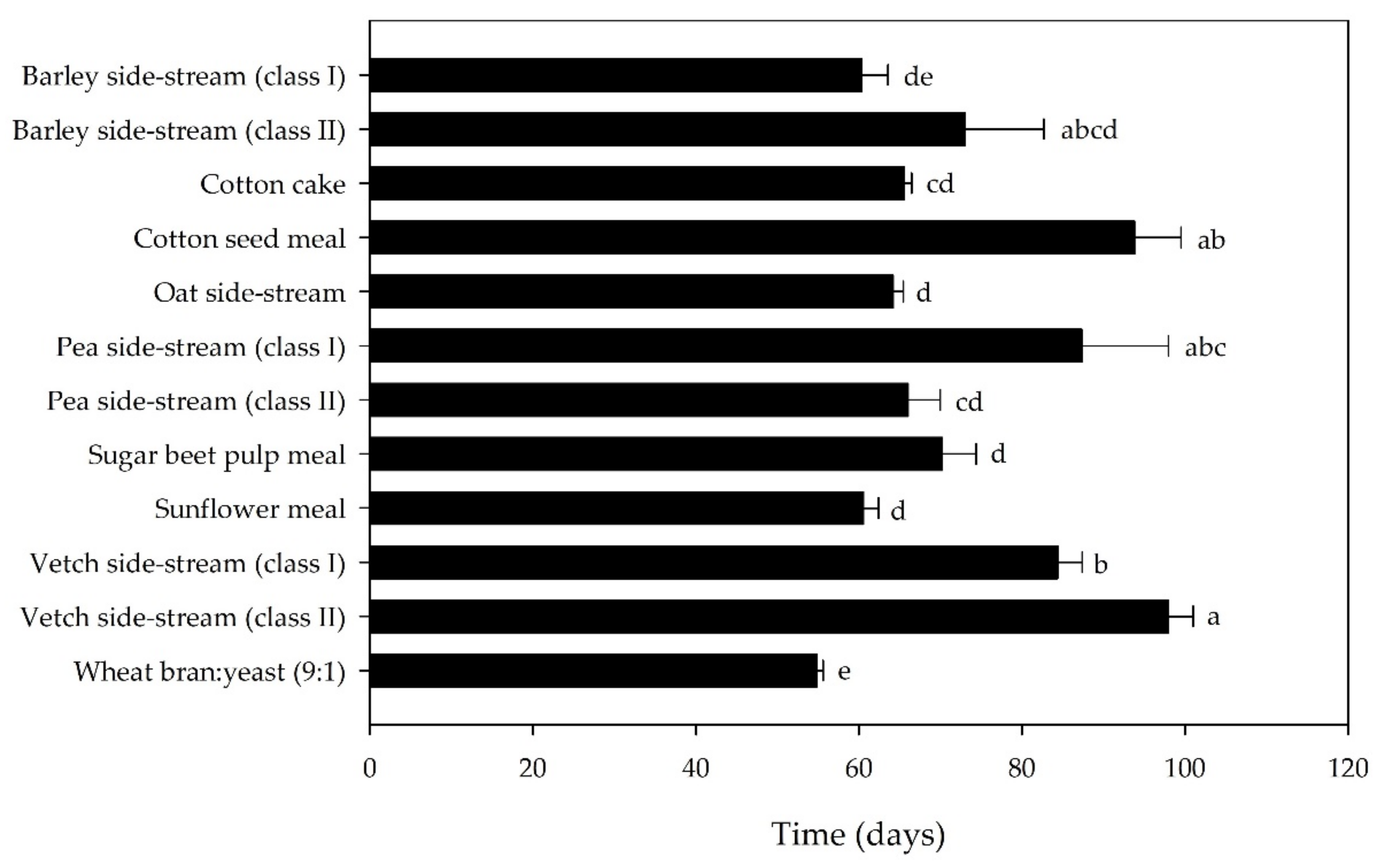
3.1. Bioassay I: Single Side-Streams
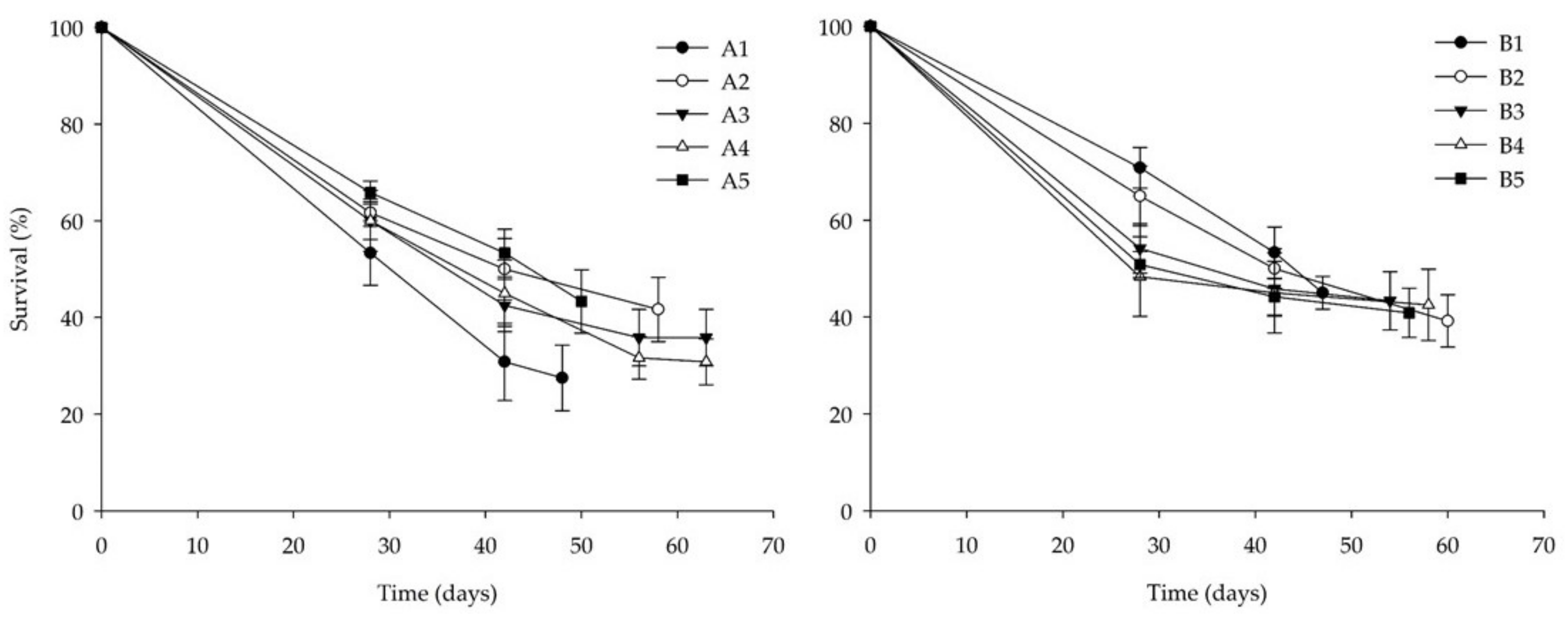
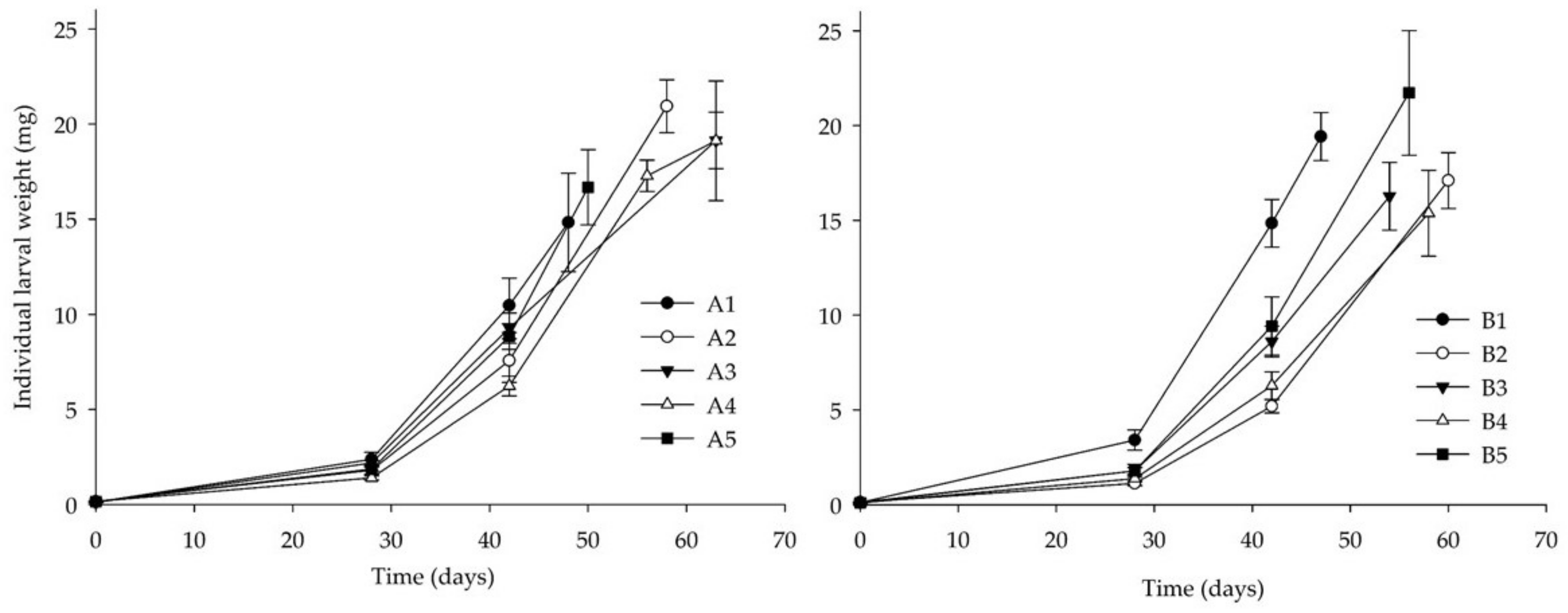
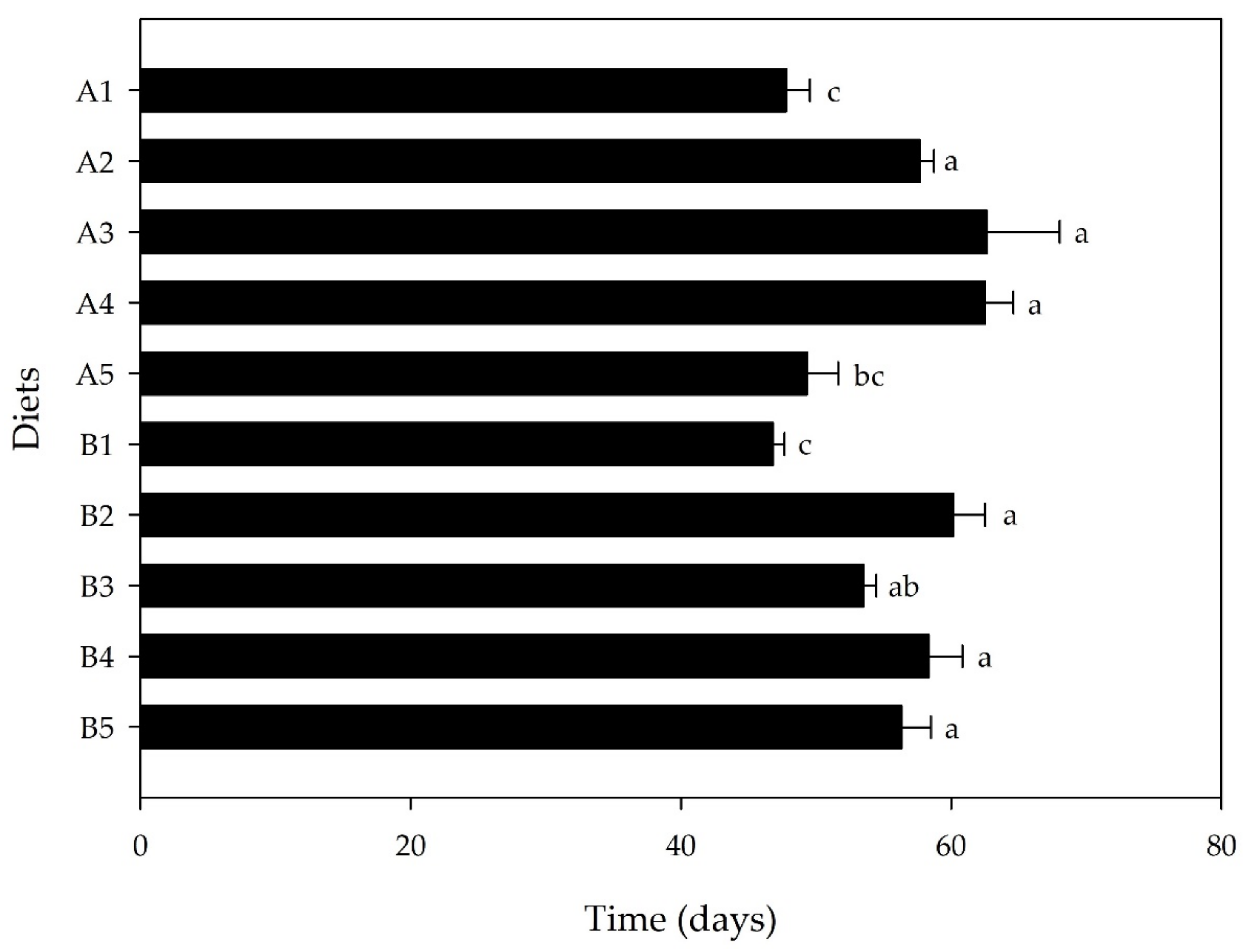
3.2. Bioassay II: Side-Streams-Based Isoproteinic Diets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sogari, G.; Amato, M.; Biasato, I.; Chiesa, S.; Gasco, L. The potential role of insects as feed: A multi-perspective review. Animals 2019, 9, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkey, K.J.; Lopez-Viso, C.; Brameld, J.M.; Parr, T.; Salter, A.M. Insects: A potential source of protein and other nutrients for feed and food. Annu. Rev. Anim. Biosci. 2021, 9, 333–354. [Google Scholar] [CrossRef] [PubMed]
- Govorushko, S. Global status of insects as food and feed source: A review. Trends Food Sci. Technol. 2019, 91, 436–445. [Google Scholar] [CrossRef]
- Tang, C.; Yang, D.; Liao, H.; Sun, H.; Liu, C.; Wei, L.; Li, F. Edible insects as a food source: A review. Food Prod. Processing Nutr. 2019, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Franco, A.; Salvia, R.; Scieuzo, C.; Schmitt, E.; Russo, A.; Falabella, P. Lipids from insects in cosmetics and for personal care products. Insects 2022, 13, 41. [Google Scholar] [CrossRef]
- Verheyen, G.R.; Ooms, T.; Vogels, L.; Vreysen, S.; Bovy, A.; Van Miert, S.; Meersman, F. Insects as an alternative source for the production of fats for cosmetics. J. Cosmet. Sci. 2018, 69, 187–202. [Google Scholar]
- Alipour, N.; Vinnerås, B.; Gouanvé, F.; Espuche, E.; Hedenqvist, M.S. A Protein-based material from a new approach using whole defatted larvae, and its interaction with moisture. Polymers 2019, 11, 287. [Google Scholar] [CrossRef] [Green Version]
- Azmiera, N.; Krasilnikova, A.; Sahudin, S.; Al-Talib, H.; Heo, C.C. Antimicrobial peptides isolated from insects and their potential applications. J. Asia-Pac. Entomol. 2022, 25, 101892. [Google Scholar] [CrossRef]
- Rumbos, C.I.; Karapanagiotidis, I.T.; Mente, E.; Athanassiou, C.G. The lesser mealworm Alphitobius diaperinus: A noxious pest or a promising nutrient source? Rev. Aquac. 2019, 11, 1418–1437. [Google Scholar] [CrossRef]
- EU Commission Regulation 2017/893 of 24 May 2017 amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Commission Regulation (EU) No 142/2011 as Regards the Provisions on Processed Animal Protein. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32017R0893 (accessed on 15 April 2022).
- EU Commission Regulation 2021/1372 of 17 August 2021 amending Annex IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council as Regards the Prohibition to Feed Non-Ruminant Farmed Animals, Other Than Fur Animals, with Protein Derived from Animals. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32021R1372 (accessed on 15 April 2022).
- Kurečka, M.; Kulma, M.; Petříčková, D.; Plachý, V.; Kouřimská, L. Larvae and pupae of Alphitobius diaperinus as promising protein alternatives. Eur. Food Res. Technol. 2021, 247, 2527–2532. [Google Scholar] [CrossRef]
- Yi, L.; Lakemond, C.M.M.; Sagis, L.M.C.; Eisner-Schadler, V.; van Huis, A.; van Boekel, M.A.J.S. Extraction and characterisation of protein fractions from five insect species. Food Chem. 2013, 141, 3341–3348. [Google Scholar] [CrossRef]
- Roncolini, A.; Milanović, V.; Aquilanti, L.; Cardinali, F.; Garofalo, C.; Sabbatini, R.; Clementi, F.; Belleggia, L.; Pasquini, M.; Mozzon, M.; et al. Lesser mealworm (Alphitobius diaperinus) powder as a novel baking ingredient for manufacturing high-protein, mineral-dense snacks. Food Res. Int. 2020, 131, 109031. [Google Scholar] [CrossRef]
- Hermans, W.J.H.; Senden, J.M.; Churchward-Venne, T.A.; Paulussen, K.J.M.; Fuchs, C.J.; Smeets, J.S.J.; van Loon, J.J.A.; Verdijk, L.B.; van Loon, L.J.C. Insects are a viable protein source for human consumption: From insect protein digestion to postprandial muscle protein synthesis in vivo in humans: A double-blind randomized trial. Am. J. Clin. Nutr. 2021, 114, 934–944. [Google Scholar] [CrossRef]
- Kotsou, K.; Rumbos, C.I.; Baliota, G.V.; Gourgouta, M.; Athanassiou, C.G. Influence of temperature, relative humidity and protein content on the growth and development of larvae of the lesser mealworm, Alphitobius diaperinus (Panzer). Sustainability 2021, 13, 11087. [Google Scholar] [CrossRef]
- Bjørge, J.D.; Overgaard, J.; Maltea, H.; Gianotten, N.; Heckmann, L. Role of temperature on growth and metabolic rate in the tenebrionid beetles Alphitobius diaperinus and Tenebrio molitor. J. Insect Physiol. 2018, 107, 89–96. [Google Scholar] [CrossRef]
- Kim, S.H.; Chung, T.H.; Park, H.C.; Shin, M.J.; Park, I.G.; Choi, I.H. Effects of diet composition on growth performance and feed conversion efficiency in Alphitobius diaperinus larvae. J. Entomol. Acarol. Res. 2012, 51, 771. [Google Scholar] [CrossRef]
- Ricciardi, C.; Baviera, C. Role of carbohydrates and proteins in maximizing productivity in Alphitobius diaperinus (Coleoptera: Tenebrionidae). J. Zool. 2016, 99, 97–105. [Google Scholar]
- Hosen, M.; Khan, A.R.; Hossain, M. Growth and development of the lesser mealworm, Alphitobius diaperinus (Panzer) (Coleoptera: Tenebrionidae) on cereal flours. Pak. J. Biol. Sci. 2004, 7, 1505–1508. [Google Scholar]
- Varelas, V. Food wastes as a potential new source for edible insect mass production for food and feed: A review. Fermentation 2019, 5, 81. [Google Scholar] [CrossRef] [Green Version]
- Gasco, L.; Biancarosa, I.; Liland, N.S. From waste to feed: A review of recent knowledge on insects as producers of protein and fat for animal feeds. Curr. Opin. Green Sustain. Chem. 2020, 23, 67–79. [Google Scholar] [CrossRef]
- EU Commission. Towards a Circular Economy: A ZeroWaste Programme for Europe. 2014. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A52014DC0398 (accessed on 11 April 2022).
- EU Commission. A New Circular Economy Action Plan: For a Cleaner and More Competitive Europe. 2020. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?qid=1583933814386&uri=COM:2020:98:FIN (accessed on 11 April 2022).
- Van Broekhoven, S.; Oonincx, D.G.A.B.; Van Huis, A.; Van Loon, J.J.A. Growth performance and feed conversion efficiency of three edible mealworm species (Coleoptera: Tenebrionidae) on diets composed of organic byproducts. J. Insect Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef]
- Gianotten, N.; Soetemans, L.; Bastiaens, L. Agri–food side–stream inclusions in the diet of Alphitobius diaperinus Part 1: Impact on larvae growth performance parameters. Insects 2020, 11, 79. [Google Scholar] [CrossRef] [Green Version]
- Soetemans, L.; Gianotten, N.; Bastiaens, L. Agri–food side–stream inclusion in the diet of Alphitobius diaperinus. Part 2: Impact on larvae composition. Insects 2020, 11, 190. [Google Scholar] [CrossRef] [Green Version]
- Rumbos, C.I.; Bliamplias, D.; Gourgouta, M.; Michail, V.; Athanassiou, C.G. Rearing Tenebrio molitor and Alphitobius diaperinus larvae on seed cleaning process byproducts. Insects 2021, 12, 293. [Google Scholar] [CrossRef]
- Rumbos, C.I.; Oonincx, D.G.A.B.; Karapanagiotidis, I.T.; Vrontaki, M.; Gourgouta, M.; Asimaki, A.; Mente, E.; Athanassiou, C.G. Agricultural byproducts from Greece as feed for yellow mealworm larvae: Circular economy at local level. J. Insects Food Feed 2021, 8, 9–22. [Google Scholar] [CrossRef]
- Hagstrum, D.W.; Subramanyam, B. Stored-Product Insect Resource; AACC International Inc.: St. Paul, MN, USA, 2009; p. 506. [Google Scholar]
- Hagstrum, D.W.; Klejdysz, T.; Subramanyam, B.; Nawrot, J. Atlas of Stored-Product Insects and Mites; AACC International Inc.: St. Paul, MN, USA, 2013; p. 589. [Google Scholar]
- Rumbos, C.I.; Pantazis, I.; Athanassiou, C.G. Population growth of Alphitobius diaperinus (Coleoptera: Tenebrionidae) on various commodities. J. Econ. Entomol. 2020, 113, 1001–1007. [Google Scholar] [CrossRef]
- Cohen, A.C. Insect Diets: Science and Technology, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2020; p. 474. [Google Scholar]
- Morales-Ramos, J.A.; Rojas, G.M.; Kay, S.; Shapiro-Ilan, D.I.; Tedders, L.W. Impact of adult weight, density, and age on reproduction of Tenebrio molitor (Coleoptera: Tenebrionidae). J. Entomol. Sci. 2012, 47, 208–220. [Google Scholar] [CrossRef]
- Howe, R.W. The influence of age of the parents on some characteristics of the offspring of insects bred in the laboratory. J. Stored Prod. Res. 1967, 3, 371–385. [Google Scholar] [CrossRef]
- Ludwig, D.; Fiore, C. Further studies on the relationship between parental age and the life cycle of the mealworm, Tenebrio molitor. Ann. Entomol. Soc. Am. 1960, 53, 595–600. [Google Scholar] [CrossRef]
- Tracey, K.M.; Jakowska, S.; Nolan, S. Parental age and free amino acids in larvae of Tenebrio molitor. Anat. Rec. 1958, 132, 513–514. [Google Scholar]
- Candia, F.; Manzanares, A. Insect market outlook: Are insect producers ready to deliver? In The Basics of Edible Insect Rearing: Handbook for the Production Chain; Veldkamp, T., Claeys, J., Haenen, O.L.M., van Loon, J.J.A., Spranghers, T., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2022; pp. 257–269. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | FCR | SGR (%) | Final Survival Rate (%) | Final Individual Larval Weight (mg) |
|---|---|---|---|---|
| Barley side-stream (class I) | 5.2 ± 0.6 d | 7.4 ± 0.7 a | 84.0 ± 4.0 a | 13.6 ± 1.6 |
| Barley side-stream (class II) | 5.5 ± 18 d | 5.6 ± 0.7 abcd | 58.3 ± 14.5 ab | 16.9 ± 1.2 |
| Cotton cake | 5.5 ± 0.7 d | 6.3 ± 0.2 abcd | 65.8 ± 3.7 ab | 17.4 ± 1.8 |
| Cotton seed meal | 22.9 ± 2.1 a | 2.5 ± 0.4 e | 10.8 ± 3.0 c | 16.3 ± 3.2 |
| Oat side-stream | 4.1 ± 0.3 d | 7.0 ± 0.4 abc | 77.5 ± 5.0 a | 18.2 ± 0.5 |
| Pea side-stream (class I) | 15.7 ± 4.6 b | 4.0 ± 0.4 de | 22.5 ± 10.8 c | 11.3 ± 2.5 |
| Pea side-stream (class II) | 12.3 ± 2.4 bc | 4.8 ± 0.6 bcde | 40.0 ± 8.0 bc | 12.3 ± 1.7 |
| Sugar beet pulp meal | 14.3 ± 1.3 b | 4.6 ± 0.3 cde | 29.2 ± 3.0 c | 16.6 ± 2.0 |
| Sunflower meal | 5.7 ± 1.2 d | 7.2 ± 0.4 ab | 73.3 ± 5.3 a | 16.0 ± 2.4 |
| Vetch side-stream (class I) | 12.7 ± 2.3 bc | 4.3 ± 0.4 de | 35.8 ± 6.0 bc | 15.2 ± 1.7 |
| Vetch side-stream (class II) | 8.0 ± 2.9 bc | 3.8 ± 0.7 de | 10.8 ± 5.2 c | 12.7 ± 3.2 |
| Wheat bran/yeast (9:1) (control) | 3.9 ± 0.1 d | 7.8 ± 0.6 a | 78.0 ± 4.1 a | 16.1 ± 2.5 |
| Diet | FCR | SGR (%) | Final Survival Rate (%) | Final Individual Larval Weight (mg) |
|---|---|---|---|---|
| A1 | 12.0 ± 1.8 | 8.0 ± 0.6 ab | 33.0 ± 7.5 | 14.8 ± 5.1 |
| A2 | 8.6 ± 2.8 | 6.6 ± 0.5 ab | 41.7 ± 6.8 | 20.9 ± 1.4 |
| A3 | 9.4 ± 1.2 | 6.4 ± 0.6 ab | 35.8 ± 5.8 | 19.1 ± 1.5 |
| A4 | 12.9 ± 2.7 | 5.7 ± 0.5 b | 30.8 ± 4.7 | 19.1 ± 7.7 |
| A5 | 12.8 ± 3.9 | 6.5 ± 0.9 ab | 43.3 ± 6.5 | 16.7 ± 2.0 |
| B1 | 7.0 ± 0.5 | 8.9 ± 0.4 a | 45.0 ± 3.4 | 19.4 ± 1.3 |
| B2 | 10.4 ± 0.9 | 6.3 ± 0.6 b | 39.2 ± 5.4 | 17.1 ± 1.5 |
| B3 | 10.4 ± 1.8 | 7.0 ± 0.3 ab | 43.3 ± 6.0 | 16.3 ± 1.8 |
| B4 | 11.4 ± 2.6 | 6.9 ± 0.4 ab | 42.5 ± 7.4 | 16.2 ± 2.3 |
| B5 | 8.1 ± 0.7 | 7.4 ± 0.3 ab | 40.8 ± 5.1 | 21.5 ± 1.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gourgouta, M.; Rumbos, C.I.; Michail, V.; Athanassiou, C.G. Valorization of Agricultural Side-Streams for the Rearing of Larvae of the Lesser Mealworm, Alphitobius diaperinus (Panzer). Sustainability 2022, 14, 7680. https://doi.org/10.3390/su14137680
Gourgouta M, Rumbos CI, Michail V, Athanassiou CG. Valorization of Agricultural Side-Streams for the Rearing of Larvae of the Lesser Mealworm, Alphitobius diaperinus (Panzer). Sustainability. 2022; 14(13):7680. https://doi.org/10.3390/su14137680
Chicago/Turabian StyleGourgouta, Marina, Christos I. Rumbos, Vasilios Michail, and Christos G. Athanassiou. 2022. "Valorization of Agricultural Side-Streams for the Rearing of Larvae of the Lesser Mealworm, Alphitobius diaperinus (Panzer)" Sustainability 14, no. 13: 7680. https://doi.org/10.3390/su14137680