
Appropriate Irrigation and Fertilization Regime Restrain Indigenous Soil Key Ammonia-Oxidizing Archaeal and Bacterial Consortia to Mitigate Greenhouse Gas Emissions

Abstract
:1. Introduction
2. Materials and Methods
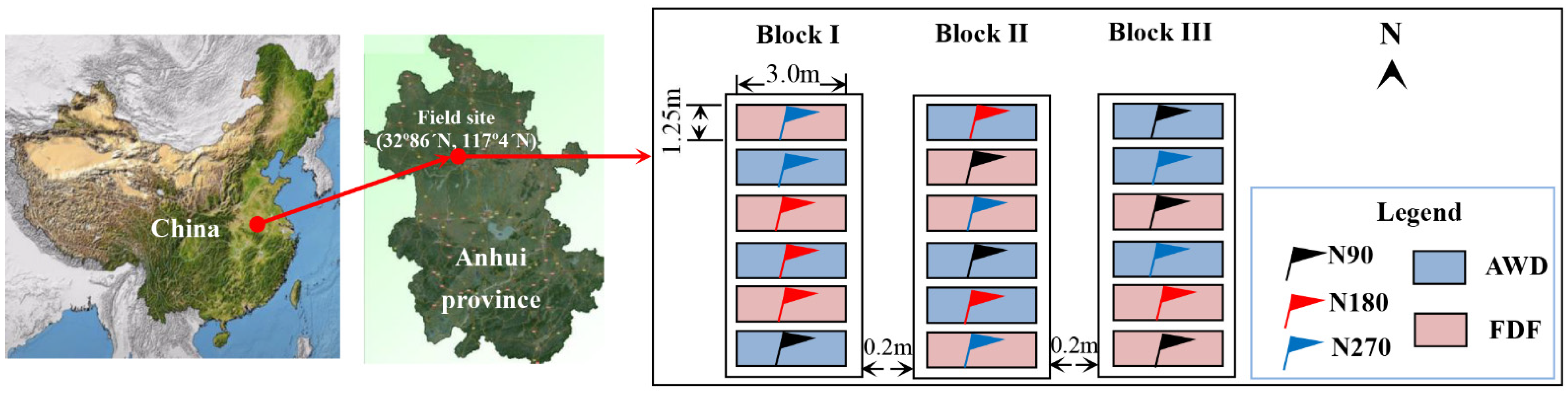
2.1. Study Site and Experimental Design
2.2. N2O and CH4 Fluxes Measurements
2.3. Soil Sampling, Properties Analysis and DNA Extraction
2.4. DNA Extraction and Sequencing
2.5. Sequence Data Processing
2.6. Statistical Analysis
3. Results and Discussion
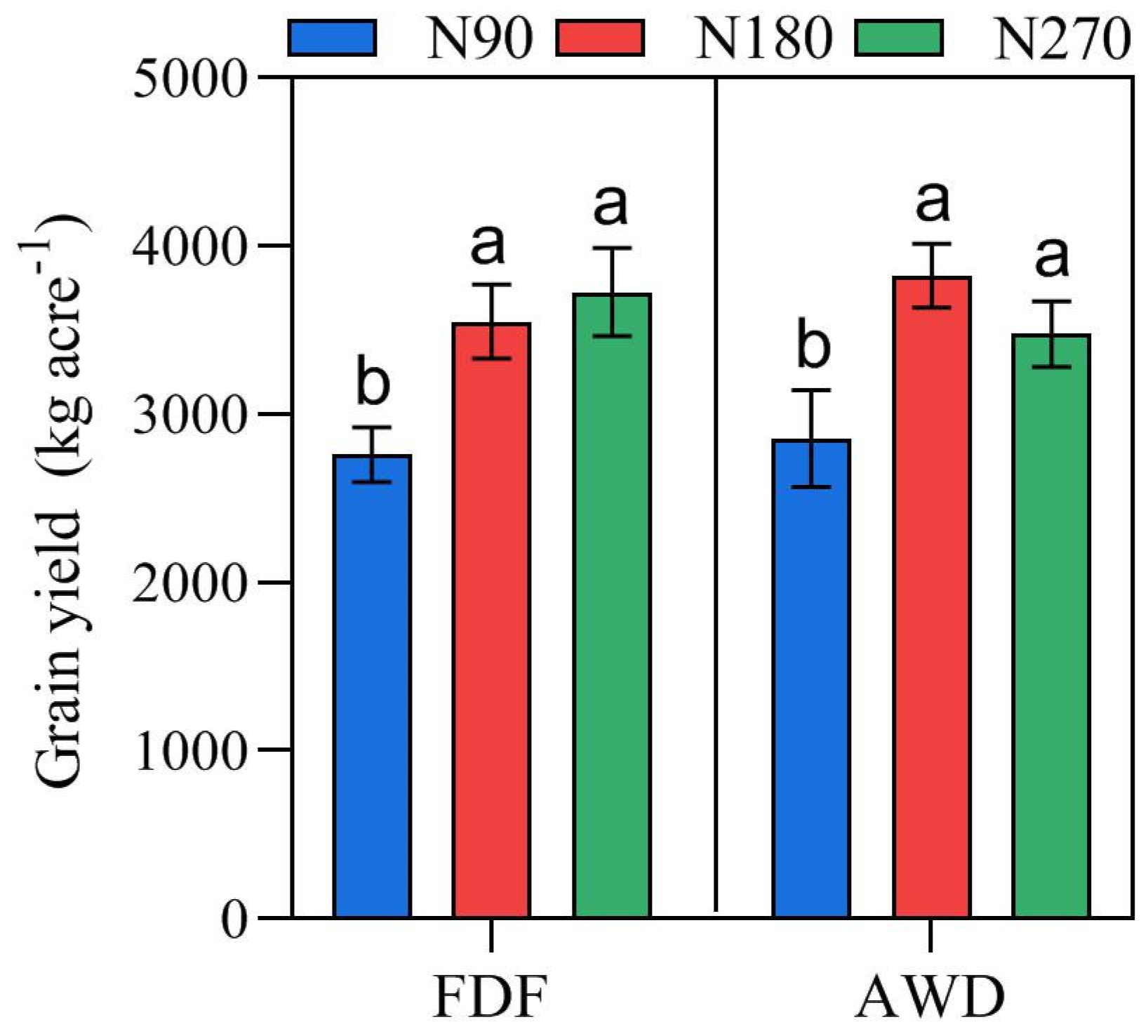
3.1. Variations of Yield
3.2. Variations of Soil Physicochemical Parameters
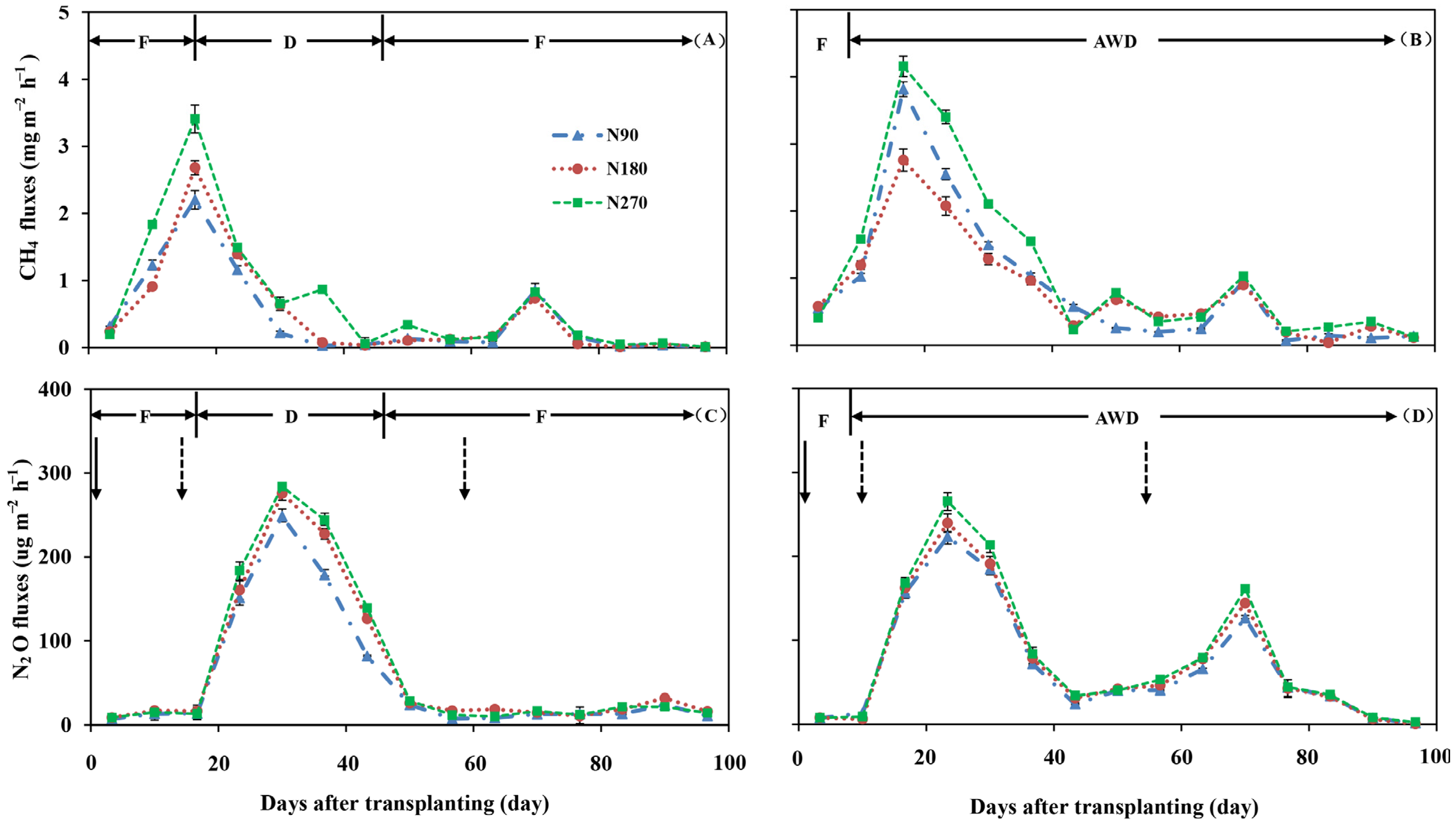
3.3. Variations of GHG Emissions
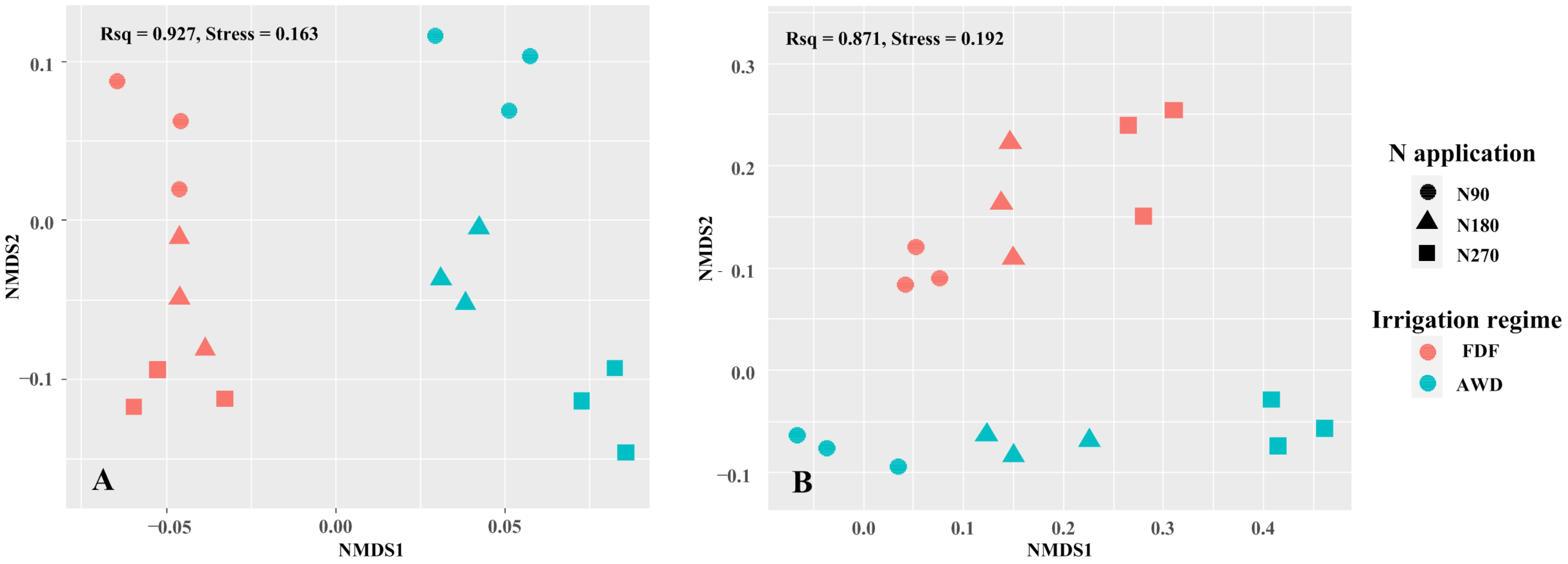
3.4. Succession of Ammonia-Oxidizing Microbiomes
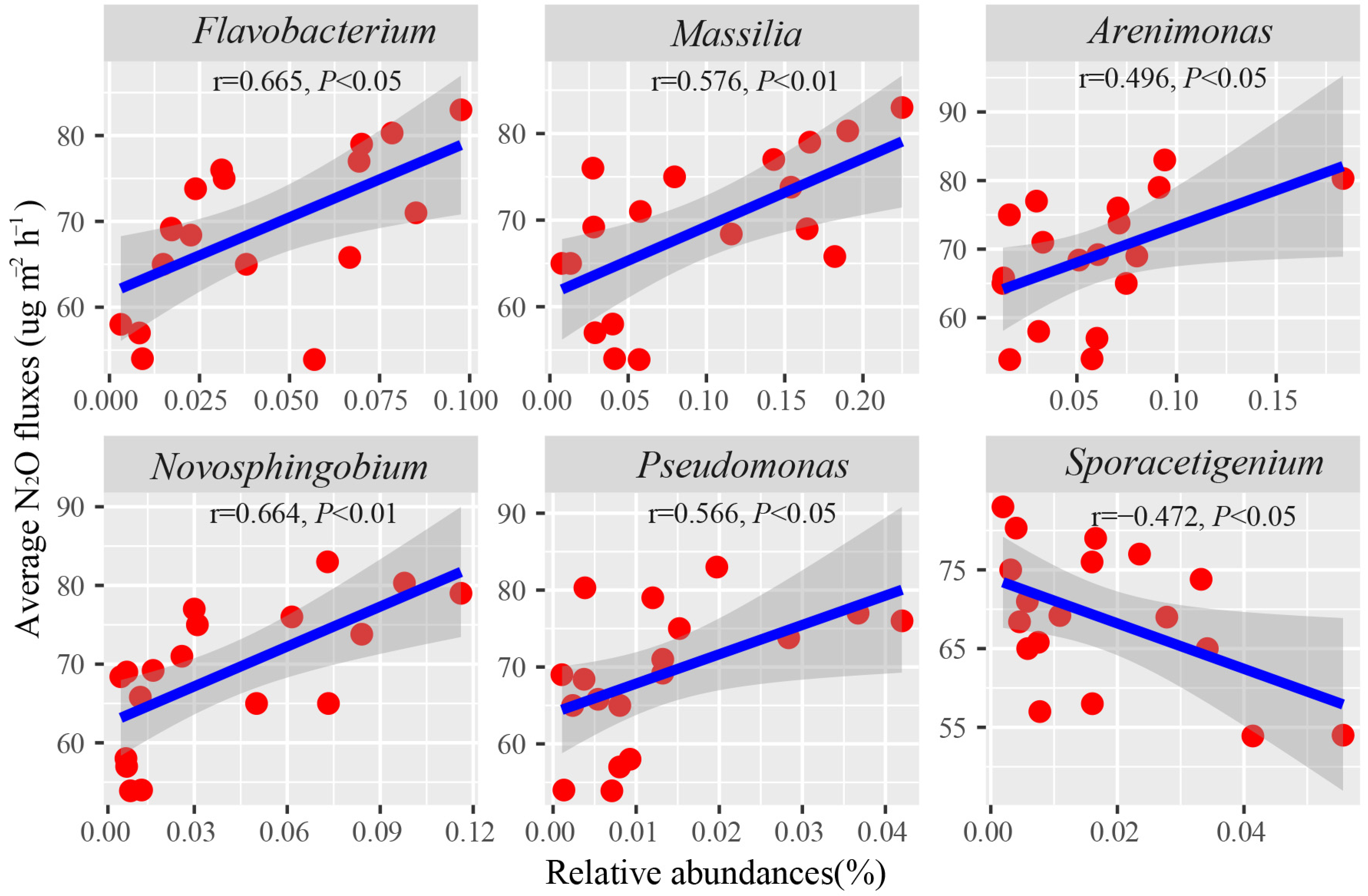
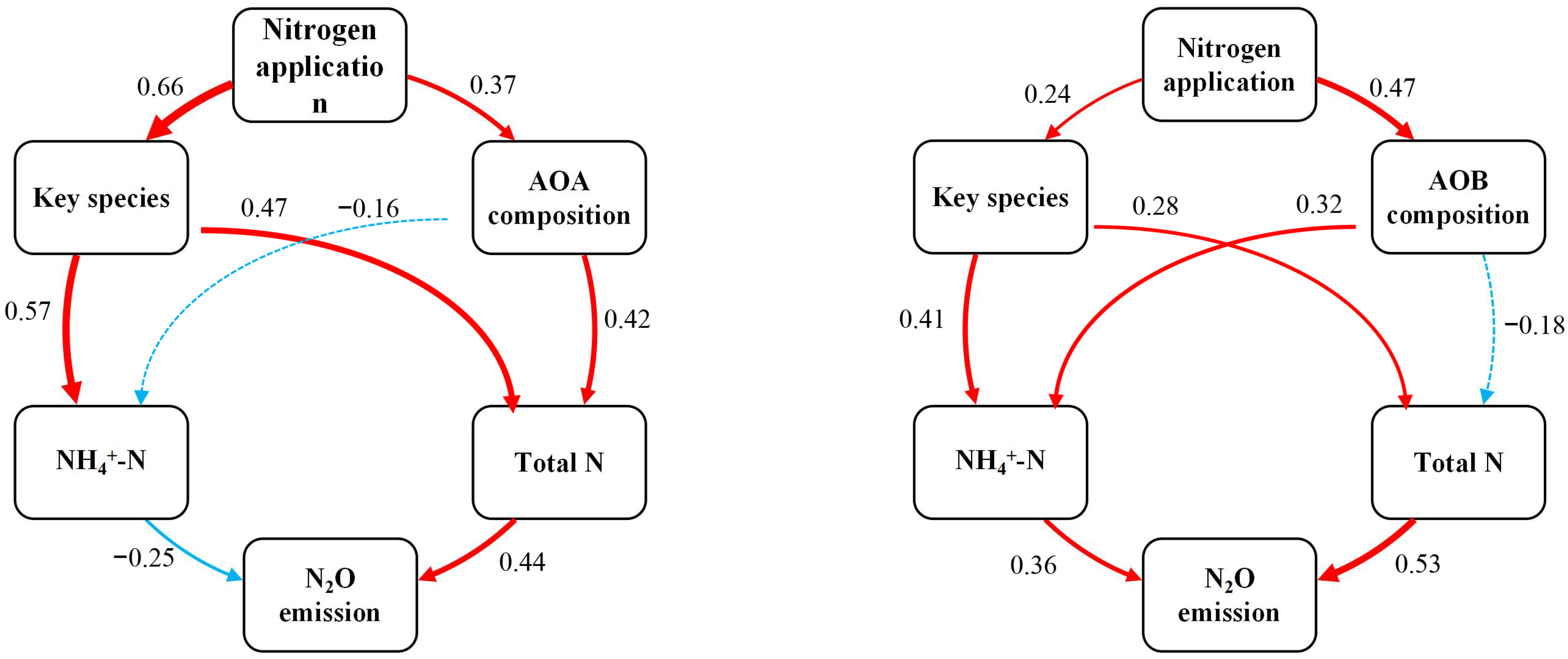
3.5. The Key Taxa Potentially Linked to GHG Emissions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Linquist, B.A.; Anders, M.M.; Adviento-Borbe, M.A.; Chaney, R.L.; Nalley, L.L.; Da, R.E.; van Kessel, C. Reducing greenhouse gas emissions, water use, and grain arsenic levels in rice systems. Glob. Chang. Biol. 2015, 21, 407–417. [Google Scholar] [CrossRef]
- Trivedi, P.; Anderson, I.C.; Singh, B.K. Microbial modulators of soil carbon storage: Integrating genomic and metabolic knowledge for global prediction. Trends Microbiol. 2013, 21, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Liu, Y.; Zhong, X.; Lampayan, R.; Singleton, G.; Huang, N.; Liang, K.; Peng, B.; Tian, K. Grain yield, water productivity and nitrogen use efficiency of rice under different water management and fertilizer-N inputs in South China. Agric. Water Manag. 2017, 184, 191–200. [Google Scholar] [CrossRef]
- Zhang, J.; Ouyang, Y.; Huang, Z.X.; Quan, G.M. Dynamic Emission of CH4 from a Rice-Duck Farming Ecosystem. J. Environ. Prot. Ecol. 2011, 2, 537–544. [Google Scholar] [CrossRef]
- Kritee, K.; Nair, D.; Zavala-Araiza, D.; Proville, J.; Rudek, J.; Adhya, T.K.; Loecke, T.; Esteves, T.; Balireddygari, S.; Dava, O.; et al. High nitrous oxide fluxes from rice indicate the need to manage water for both long- and short-term climate impacts. Proc. Natl. Acad. Sci. USA 2018, 115, 9720–9725. [Google Scholar] [CrossRef] [Green Version]
- Berger, S.; Jang, I.; Seo, J.; Kang, H.; Gebauer, G. A record of N2O and CH4 emissions and underlying soil processes of Korean rice paddies as affected by different water management practices. Biogeochemistry 2013, 115, 317–332. [Google Scholar] [CrossRef]
- Wang, L.L.; Palta, J.A.; Chen, W.; Chen, Y.L.; Deng, X.P. Nitrogen fertilization improved water-use efficiency of winter wheat through increasing water use during vegetative rather than grain filling. Agric. Water Manag. 2018, 197, 41–53. [Google Scholar] [CrossRef]
- Shang, Q.; Yang, X.; Gao, C.; Wu, P.; Liu, J.; Xu, Y.; Shen, Q.; Zou, J.; Guo, S. Net annual global warming potential and greenhouse gas intensity in Chinese double rice-cropping systems: A 3-year field measurement in long-term fertilizer experiments. Glob. Chang. Biol. 2011, 17, 2196–2210. [Google Scholar] [CrossRef]
- Li, P.P.; Han, Y.L.; He, J.Z.; Zhang, S.Q.; Zhang, L.M. Soil aggregate size and long-term fertilization effects on the function and community of ammonia oxidizers. Geoderma 2019, 338, 107–117. [Google Scholar] [CrossRef]
- Álvaro-Fuentes, J.; Arrúe, J.L.; Cantero-Martínez, C.; Isla, R.; Plaza-Bonilla, D.; Quílez, D. Fertilization Scenarios in Sprinkler-Irrigated Corn under Mediterranean Conditions: Effects on Greenhouse Gas Emissions. Soil Sci. Soc. Am. J. 2016, 80, 662–671. [Google Scholar] [CrossRef] [Green Version]
- Ning, D.; Qin, A.; Duan, A.; Xiao, J.; Zhang, J.; Liu, Z.; Liu, Z.; Zhao, B.; Liu, Z. Deficit irrigation combined with reduced N-fertilizer rate can mitigate the high nitrous oxide emissions from Chinese drip-fertigated maize field. Glob. Ecol. Conserv. 2019, 20, e803. [Google Scholar] [CrossRef]
- Trost, B.; Prochnow, A.; Meyer-Aurich, A.; Drastig, K.; Baumecker, M.; Ellmer, F. Effects of irrigation and nitrogen fertilization on the greenhouse gas emissions of a cropping system on a sandy soil in northeast Germany. Eur. J. Agron. 2016, 81, 117–128. [Google Scholar] [CrossRef]
- Baggs, E.M. Soil microbial sources of nitrous oxide: Recent advances in knowledge, emerging challenges and future direction. Curr. Opin. Environ. Sustain. 2011, 3, 321–327. [Google Scholar] [CrossRef]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.L.; Xu, Z.W.; Hu, H.W.; Hu, Y.J.; Hao, Z.P.; Jiang, Y. Responses of ammonia-oxidizing bacteria and archaea to nitrogen fertilization and precipitation increment in a typical temperate steppe in Inner Mongolia. Appl. Soil Ecol. 2013, 68, 36–45. [Google Scholar] [CrossRef]
- Cordovez, V.; Dini-Andreote, F.; Carrión, V.J.; Raaijmakers, J.M. Ecology and Evolution of Plant Microbiomes. Annu. Rev. Microbiol. 2019, 73, 69–88. [Google Scholar] [CrossRef] [PubMed]
- Reay, D.S.; Davidson, E.A.; Smith, K.A.; Smith, P.; Melillo, J.M.; Dentener, F. Global agriculture and nitrous oxide emissions. Nat. Clim. Chang. 2012, 2, 410–416. [Google Scholar] [CrossRef]
- Chen, H.Y.; Sanjeev, K.A.; Liu, T.; Duan, Y.; Ren, X.; Zhang, Z.; Ashok, P.; Mukesh, K.A. Effects of microbial culture and chicken manure biochar on compost maturity and greenhouse gas emissions during chicken manure composting. J. Hazard. Mater. 2020, 389, 121908. [Google Scholar] [CrossRef]
- Yao, Z.; Wei, Y.; Liu, C.; Zheng, X.; Xie, B. Organically fertilized tea plantation restrains N2O emissions and lowers NO fuxes in subtropical China. Biogeosciences 2015, 12, 5915–5928. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Hu, Z.; Wu, S.; Li, S.; Li, Z.; Zou, J. Methane and Nitrous Oxide Emissions Reduced Following Conversion of Rice Paddies to Inland Crab-Fish Aquaculture in Southeast China. Environ. Sci. Technol. 2016, 50, 633–642. [Google Scholar] [CrossRef]
- Maucieri, C.; Zhang, Y.; McDaniel, M.; Borin, M.; Adams, M. Short-term effects of biochar and salinity on soil greenhouse gas emissions from a semi-arid Australian soil after re-wetting. Geoderma 2017, 307, 267–276. [Google Scholar] [CrossRef]
- Qiao, C.C.; Christopher, R.P.; Xiong, W.; Liu, C.; Wang, R.; Liu, Z.; Xu, X.; Li, R.; Shen, Q.R. Reshaping the rhizosphere microbiome by bio-organic amendment to enhance crop yield in a maize-cabbage rotation system. Appl. Soil. Ecol. 2019, 142, 136–146. [Google Scholar] [CrossRef]
- Ju, X.T.; Xing, G.X.; Chen, X.P.; Zhang, S.L.; Zhang, L.J.; Liu, X.J.; Cui, Z.L.; Yin, B.; Christie, P.; Zhu, Z.L.; et al. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef] [Green Version]
- Chu, G.; Wang, Z.; Zhang, H.; Liu, L.; Yang, J.; Zhang, J. Alternate wetting and moderate drying increases rice yield and reduces methane emission in paddy field with wheat straw residue incorporation. Food Energy Secur. 2015, 4, 238–254. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Zhang, Y.; McDaniel, M.D.; Sun, L.; Su, W.; Fan, X.; Liu, S.; Xiao, X. Water-saving irrigation is a ‘win-win’ management strategy in rice paddies—With both reduced greenhouse gas emissions and enhanced water use efficiency. Agric. Water Manag. 2020, 228, 105889. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Z.; Hu, Y.; Zeng, Z. Irrigation frequency alters the abundance and community structure of ammonia-oxidizing archaea and bacteria in a northern Chinese upland soil. Eur. J. Soil. Biol. 2017, 83, 34–42. [Google Scholar] [CrossRef]
- Song, Y.N.; Lin, Z.M. Abundance and Community Composition of Ammonia-Oxidizers in Paddy Soil at Different Nitrogen Fertilizer Rates. J. Integr. Agric. 2014, 13, 870–880. [Google Scholar] [CrossRef] [Green Version]
- He, L.L.; Bi, Y.C.; Zhao, J.; Pittelkow, C.M.; Zhao, X.; Wang, S.Q.; Xing, G.X. Population and community structure shifts of ammonia oxidizers after four-year successive biochar application to agricultural acidic and alkaline soils. Sci. Total Environ. 2018, 619, 1105–1115. [Google Scholar] [CrossRef]
- Pishgar, R.; Dominic, J.A.; Sheng, Z.; Tay, J.H. Denitrification performance and microbial versatility in response to different selection pressures. Bioresour. Technol. 2019, 281, 72–83. [Google Scholar] [CrossRef]
- Yu, S.; He, R.; Song, A.; Huang, Y.; Jin, Z.; Liang, Y.; Li, Q.; Wang, X.; Müller, W.E.G.; Cao, J. Spatial and temporal dynamics of bacterioplankton community composition in a subtropical dammed karst river of southwestern China. Microbiologyopen 2019, 8, e00849. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, J.T.; Hu, H.W.; Cai, Z.J.; Lei, Y.R.; Li, W. Changes of the denitrifying communities in a multi-stage free water surface constructed wetland. Sci. Total Environ. 2019, 650, 1419–1425. [Google Scholar] [CrossRef] [PubMed]
- Xing, W.; Li, J.; Li, D.; Hu, J.; Deng, S.; Cui, Y.; Yao, H. Stable-Isotope Probing Reveals the Activity and Function of Autotrophic and Heterotrophic Denitrifiers in Nitrate Removal from Organic-Limited Wastewater. Environ. Sci. Technol. 2018, 52, 7867–7875. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Panicles (×104·ha−1) | Grains per Panicle | Filled Grains (%) | 1000-Grain Weight (g) | Yield (kg.acre−1) |
|---|---|---|---|---|---|
| Nitrogen application (NA) | |||||
| N90 | 12.17 ± 1.89 a | 176.09 ± 3.29 b | 91.76 ± 1.20 a | 28.40 ± 0.66 a | 2808.04 ± 68.59 b |
| N180 | 11.91 ± 0.15 a | 202.27 ± 5.75 a | 94.03 ± 1.57 a | 28.53 ± 0.81 a | 3688.37 ± 191.75 a |
| N270 | 12.59 ± 1.29 a | 197.67 ± 4.51 a | 94.30 ± 1.77 a | 28.35 ± 1.24 a | 3603.03 ± 177.18 a |
| Irrigation regime (IR) | |||||
| FDF | 11.16 ± 1.00 a | 191.65 ± 13.73 a | 93.09 ± 3.59 a | 28.70 ± 0.67 a | 3346.87 ± 516.19 a |
| AWD | 12.11 ± 1.34 a | 192.37 ± 15.13 a | 93.63 ± 5.36 a | 28.18 ± 0.82 a | 3386.08 ± 490.21 a |
| ANOVA p-values | |||||
| NA | NS | NS | NS | NS | <0.01 |
| IR | NS | NS | NS | NS | NS |
| NA*IR | NS | <0.05 | NS | NS | <0.05 |
| Items | pH | TOC (g kg−1) | NH4+-N (mg kg−1) | NO3−-N (mg kg−1) | Total N (g kg−1) |
|---|---|---|---|---|---|
| Nitrogen application (NA) | |||||
| N90 | 7.17 ± 0.04 a | 13.79 ± 1.54 a | 69.50 ± 4.61 c | 3.81 ± 0.14 b | 1.18 ± 0.08 c |
| N180 | 7.04 ± 0.09 b | 13.44 ± 1.71 a | 75.76 ± 2.64 b | 4.21 ± 0.54 b | 1.35 ± 0.06 b |
| N270 | 6.96 ± 0.05 b | 13.46 ± 0.87 a | 86.80 ± 3.78 a | 5.30 ± 0.54 a | 1.59 ± 0.13 a |
| Irrigation regime (IR) | |||||
| FDF | 7.03 ± 0.10 a | 13.01 ± 1.16 a | 75.45 ± 8.7 b | 4.24 ± 0.67 b | 1.38 ± 0.02 a |
| AWD | 7.08 ± 0.11 a | 14.12 ± 1.35 a | 79.26 ± 7.54 a | 4.64 ± 0.86 a | 1.36 ± 0.20 a |
| ANOVA p-values | |||||
| NA | <0.01 | NS | NS | <0.01 | <0.001 |
| IR | NS | NS | <0.05 | <0.05 | NS |
| NA*IR | NS | NS | <0.05 | NS | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, L.; Bao, L.; Ren, L.; Xie, Y.; Wang, H.; Wang, X.; Wang, J.; Qiao, C.; Xiao, X. Appropriate Irrigation and Fertilization Regime Restrain Indigenous Soil Key Ammonia-Oxidizing Archaeal and Bacterial Consortia to Mitigate Greenhouse Gas Emissions. Sustainability 2022, 14, 6113. https://doi.org/10.3390/su14106113
Xiao L, Bao L, Ren L, Xie Y, Wang H, Wang X, Wang J, Qiao C, Xiao X. Appropriate Irrigation and Fertilization Regime Restrain Indigenous Soil Key Ammonia-Oxidizing Archaeal and Bacterial Consortia to Mitigate Greenhouse Gas Emissions. Sustainability. 2022; 14(10):6113. https://doi.org/10.3390/su14106113
Chicago/Turabian StyleXiao, Liang, Libin Bao, Lantian Ren, Yiqin Xie, Hong Wang, Xiang Wang, Jianfei Wang, Cece Qiao, and Xin Xiao. 2022. "Appropriate Irrigation and Fertilization Regime Restrain Indigenous Soil Key Ammonia-Oxidizing Archaeal and Bacterial Consortia to Mitigate Greenhouse Gas Emissions" Sustainability 14, no. 10: 6113. https://doi.org/10.3390/su14106113