
Acaulospora as the Dominant Arbuscular Mycorrhizal Fungi in Organic Lowland Rice Paddies Improves Phosphorus Availability in Soils


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Determination of AMF in Rice Roots
2.2. Determination of AMF Diversity in Rice Roots
2.3. Trap Culture
2.4. Identification of Cultured AMF Spores
2.5. Experimental Design for Determination the Role of Acaulospora fungi in Rice Seedling
2.6. Analysis of Soil Properties
2.7. Determination of Plant Growth Performance
2.8. Determination of Pi Content in Rice Seedlings
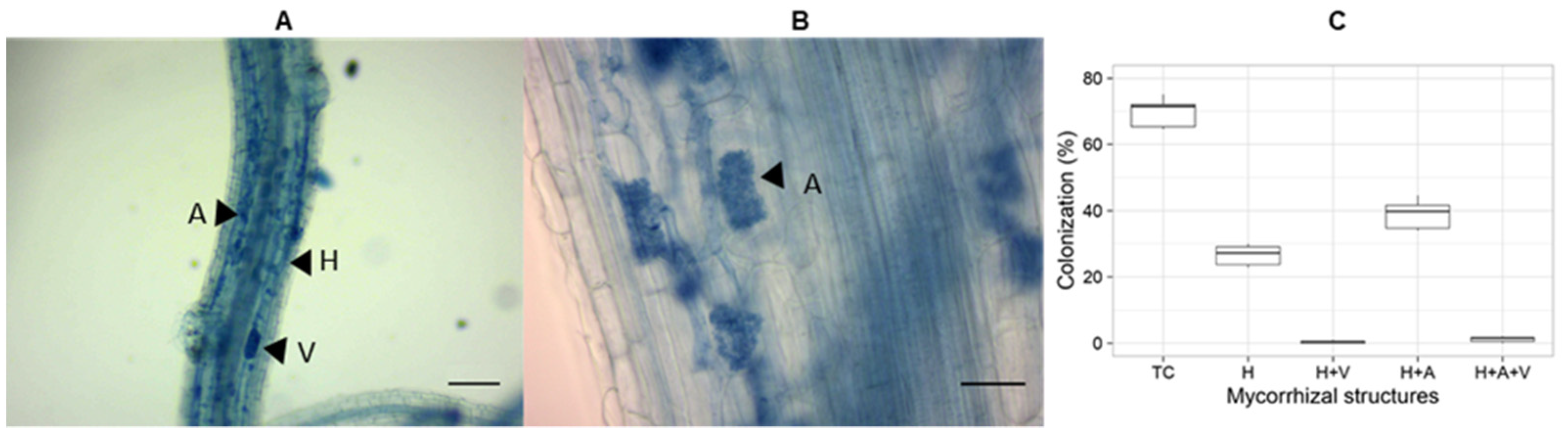
2.9. Determination of AMF Colonization
2.10. Determination of Soil Acid Phosphatase
2.11. Data Analysis
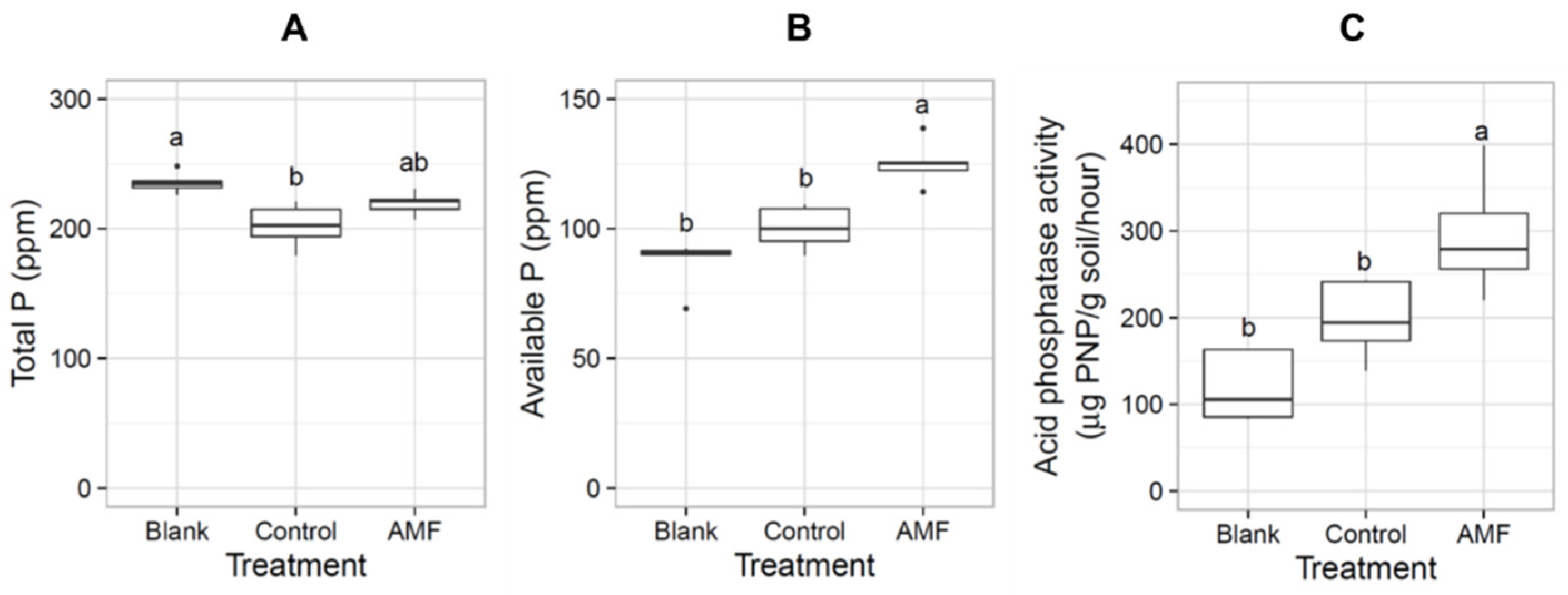
3. Results
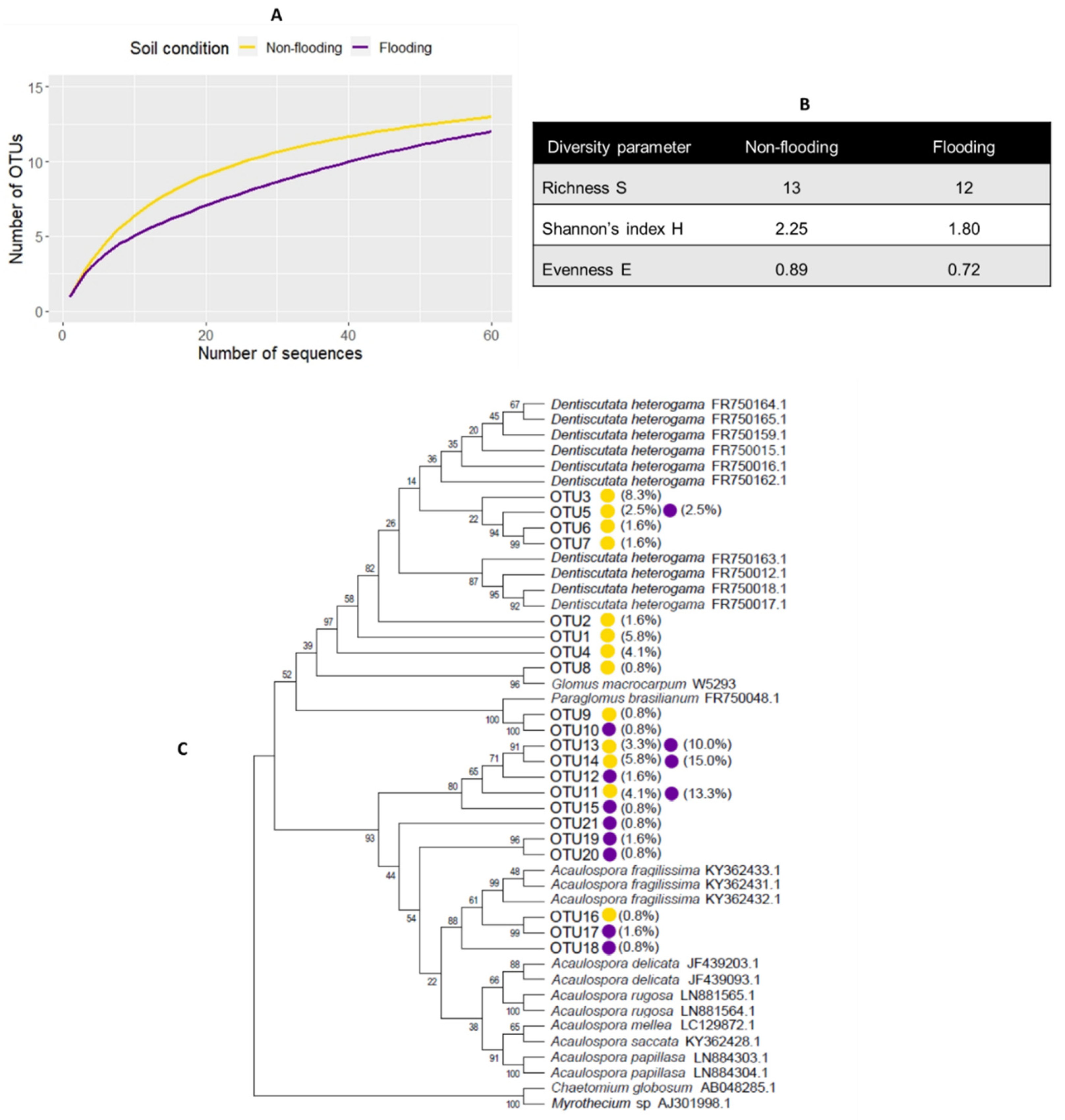
3.1. Acaulospora Is the Dominant AMF in Lowland Rice Roots under Non-Flooded and Flooded Conditions
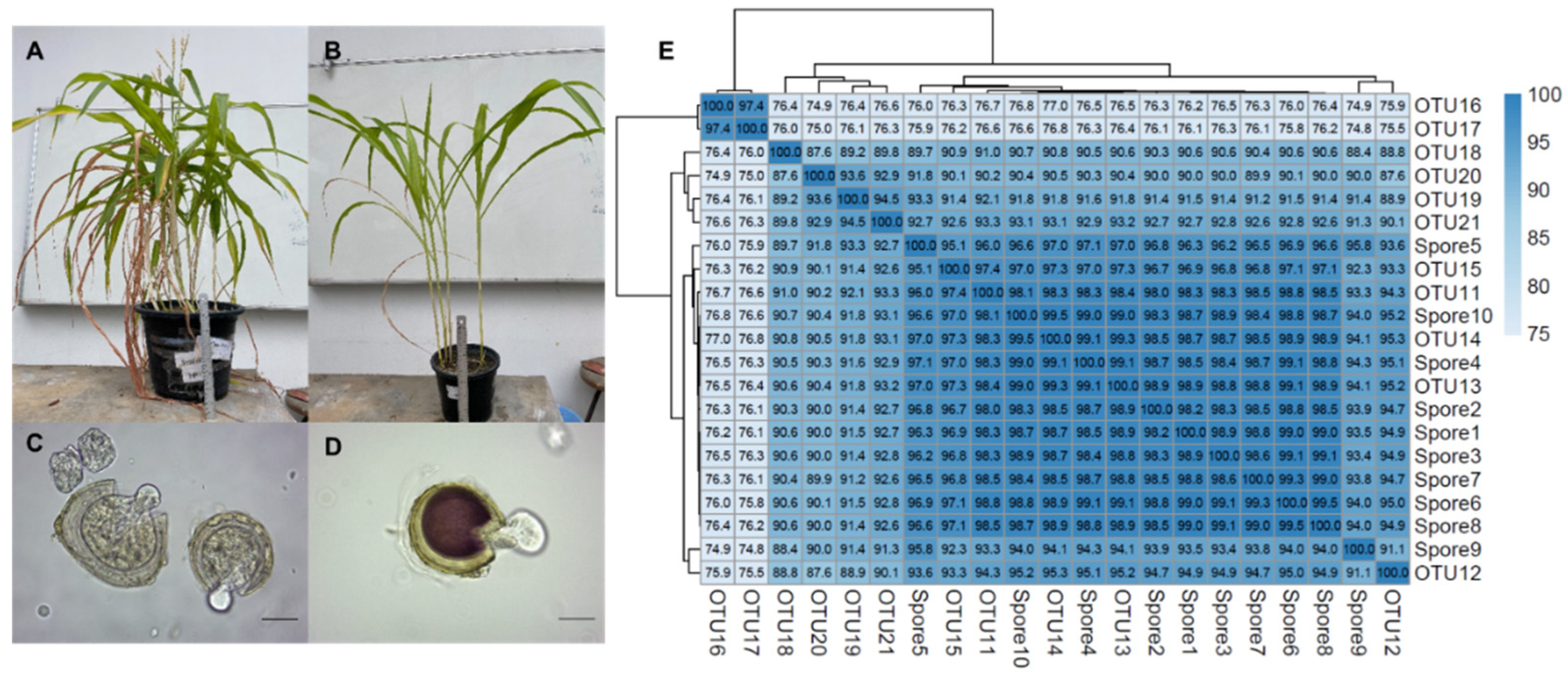
3.2. Acaulospora Spores Are Propagated by Trap Culture
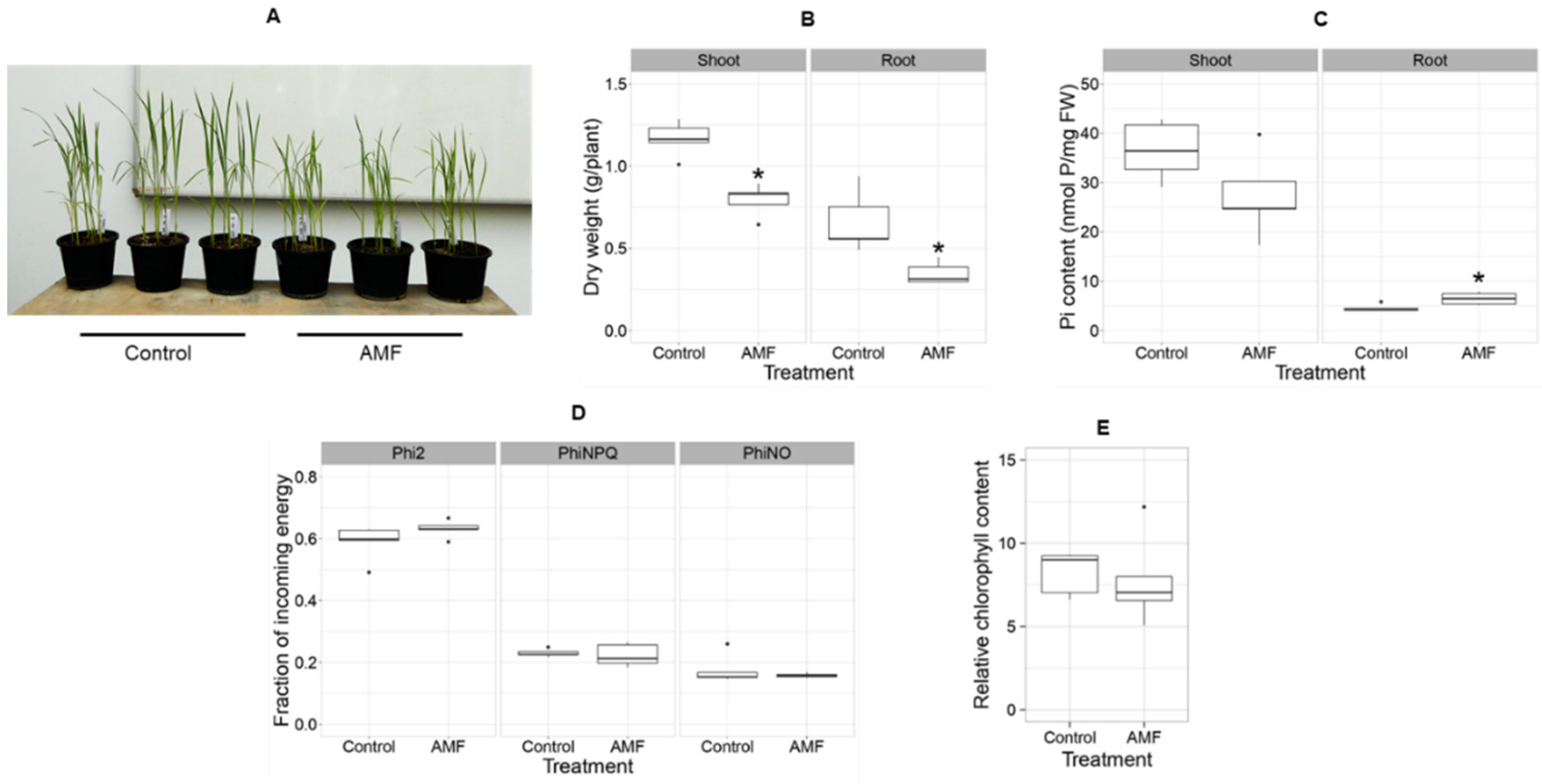
3.3. The Acaulospora Fungi Suppress Rice Seedling Growth but Not Photosynthetic Efficiency
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, S.; Chen, A.; Xie, K.; Yang, X.; Luo, Z.; Chen, J.; Zeng, D.; Ren, Y.; Yang, C.; Wang, L.; et al. Functional analysis of the OsNPF4.5 nitrate transporter reveals a conserved mycorrhizal pathway of nitrogen acquisition in plants. Proc. Natl. Acad. Sci. USA 2020, 117, 16649–16659. [Google Scholar] [CrossRef]
- Kobae, Y.; Ohmori, Y.; Saito, C.; Yano, K.; Ohtomo, R.; Fujiwara, T. Phosphate Treatment Strongly Inhibits New Arbuscule Development But Not the Maintenance of Arbuscule in Mycorrhizal Rice Roots. Plant Physiol. 2016, 171, 566–579. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Rasool, G.; Guo, X.; Sen, L.; Cao, K. Effects of Different Irrigation Methods on Environmental Factors, Rice Production, and Water Use Efficiency. Water 2020, 12, 2239. [Google Scholar] [CrossRef]
- Vallino, M.; Fiorilli, V.; Bonfante, P. Rice flooding negatively impacts root branching and arbuscular mycorrhizal colonization, but not fungal viability. Plant Cell Environ. 2014, 37, 557–572. [Google Scholar] [CrossRef]
- Klinnawee, L.; Noirungsee, N.; Nopphakat, K.; Runsaeng, P.; Chantarachot, T. Flooding overshadows phosphorus availability in controlling the intensity of arbuscular mycorrhizal colonization in Sangyod Muang Phatthalung lowland indica rice. ScienceAsia 2021, 47, 1–9. [Google Scholar] [CrossRef]
- Chialva, M.; Ghignone, S.; Cozzi, P.; Lazzari, B.; Bonfante, P.; Abbruscato, P.; Lumini, E. Water management and phenology influence the root-associated rice field microbiota. FEMS Microbiol. Ecol. 2020, 96, 96. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, T.; Li, Y.; Björn, L.O.; Rosendahl, S.; Olsson, P.A.; Li, S.; Fu, X. Community dynamics of arbuscular mycorrhizal fungi in high-input and intensively irrigated rice cultivation systems. Appl. Environ. Microbiol. 2015, 81, 2958–2965. [Google Scholar] [CrossRef] [Green Version]
- Watanarojanaporn, N.; Boonkerd, N.; Tittabutr, P.; Longtonglang, A.; Young, J.P.W.; Teaumroong, N. Effect of rice cultivation systems on indigenous arbuscular mycorrhizal fungal community structure. Microbes Environ. 2013, 28, 316–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.; Sun, Y.; Tian, L.; Ji, L.; Luo, S.; Nasir, F.; Kuramae, E.E.; Tian, C. The Structure of Rhizosphere Fungal Communities of Wild and Domesticated Rice: Changes in Diversity and Co-occurrence Patterns. Front. Microbiol. 2021, 12, 45. [Google Scholar] [CrossRef]
- Wang, Y.; Bao, X.; Li, S. Effects of Arbuscular Mycorrhizal Fungi on Rice Growth Under Different Flooding and Shading Regimes. Front. Microbiol. 2021, 12, 3276. [Google Scholar] [CrossRef]
- Andrino, A.; Guggenberger, G.; Sauheitl, L.; Burkart, S.; Boy, J. Carbon investment into mobilization of mineral and organic phosphorus by arbuscular mycorrhiza. Biol. Fertil. Soils 2021, 57, 47–64. [Google Scholar] [CrossRef]
- Sato, T.; Hachiya, S.; Inamura, N.; Ezawa, T.; Cheng, W.; Tawaraya, K. Secretion of acid phosphatase from extraradical hyphae of the arbuscular mycorrhizal fungus Rhizophagus clarus is regulated in response to phosphate availability. Mycorrhiza 2019, 29, 599–605. [Google Scholar] [CrossRef]
- Wang, X.-X.; Hoffland, E.; Feng, G.; Kuyper, T.W. Phosphate Uptake from Phytate Due to Hyphae-Mediated Phytase Activity by Arbuscular Mycorrhizal Maize. Front. Plant Sci. 2017, 8, 684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Guo, X.; Yun, W.; Xia, Y.; You, Z.; Rillig, M.C. Arbuscular mycorrhiza contributes to the control of phosphorus loss in paddy fields. Plant Soil 2020, 447, 623–636. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, L.; Ma, F.; Zhang, X.; Fu, D. Reducing nitrogen runoff from paddy fields with arbuscular mycorrhizal fungi under different fertilizer regimes. J. Environ. Sci. 2016, 46, 92–100. [Google Scholar] [CrossRef]
- Chareesri, A.; De Deyn, G.B.; Sergeeva, L.; Polthanee, A.; Kuyper, T.W. Increased arbuscular mycorrhizal fungal colonization reduces yield loss of rice (Oryza sativa L.) under drought. Mycorrhiza 2020, 30, 315–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvin, S.; Van Geel, M.; Yeasmin, T.; Lievens, B.; Honnay, O. Variation in arbuscular mycorrhizal fungal communities associated with lowland rice (Oryza sativa) along a gradient of soil salinity and arsenic contamination in Bangladesh. Sci. Total Environ. 2019, 686, 546–554. [Google Scholar] [CrossRef]
- Sarkodee-Addo, E.; Yasuda, M.; Gyu Lee, C.; Kanasugi, M.; Fujii, Y.; Omari, R.A.; Abebrese, S.O.; Bam, R.; Asuming-Brempong, S.; Dastogeer, K.M.G.; et al. Arbuscular Mycorrhizal Fungi Associated with Rice (Oryza sativa L.) in Ghana: Effect of Regional Locations and Soil Factors on Diversity and Community Assembly. Agronomy 2020, 10, 559. [Google Scholar] [CrossRef]
- Baki, M.Z.I.; Suzuki, K.; Takahashi, K.; Chowdhury, S.A.; Asiloglu, R.; Harada, N. Molecular genetic characterization of arbuscular mycorrhizal fungi associated with upland rice in Bangladesh. Rhizosphere 2021, 18, 100357. [Google Scholar] [CrossRef]
- Martins, W.F.X.; Rodrigues, B.F. Identification of dominant arbuscular mycorrhizal fungi in different rice ecosystems. Agric. Res. 2020, 9, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Kruger, M.; Stockinger, H.; Kruger, C.; Schussler, A. DNA-based species level detection of Glomeromycota: One PCR primer set for all arbuscular mycorrhizal fungi. New Phytol. 2009, 183, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jombart, T. Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, S. Kmer: An R Package for Fast Alignment-Free Clustering of Biological Sequences; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.B.; Simpson, G.; Solymos, P.; Stevenes, M.H.H.; Wagner, H. Vegan: Community Ecology Package; R Package Version 2.4-6; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Gerdemann, J.W.; Nicolson, T.H. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- Charif, D.; Lobry, J.R. SeqinR 1.0-2: A Contributed Package to the R Project for Statistical Computing Devoted to Biological Sequences Retrieval and Analysis BT. In Structural Approaches to Sequence Evolution: Molecules, Networks, Populations; Bastolla, U., Porto, M., Roman, H.E., Vendruscolo, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 207–232. ISBN 978-3-540-35306-5. [Google Scholar]
- Kolde, R. Pheatmap: Pretty Heatmaps, R Package Version 1.2; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- AOAC International. AOAC Official Method 958.01. In Official Methods of Analysis of AOAC International; Kane, P.F., Ed.; AOAC International: Rockville, MD, USA, 2016; p. 6. [Google Scholar]
- AOAC International. AOAC Official Method 970.01. In Official Methods of Analysis of AOAC International; Kane, P.F., Ed.; AOAC International: Rockville, MD, USA, 2016; p. 9. [Google Scholar]
- AOAC International. AOAC Official Method 983.02. In Official Methods of Analysis of AOAC International; Kane, P.F., Ed.; AOAC International: Rockville, MD, USA, 2016; pp. 23–26. [Google Scholar]
- Kuhlgert, S.; Austic, G.; Zegarac, R.; Osei-Bonsu, I.; Hoh, D.; Chilvers, M.I.; Roth, M.G.; Bi, K.; TerAvest, D.; Weebadde, P.; et al. MultispeQ Beta: A tool for large-scale plant phenotyping connected to the open PhotosynQ network. R. Soc. Open Sci. 2016, 3, 160592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurry, V.; Strand, A.; Furbank, R.; Stitt, M. The role of inorganic phosphate in the development of freezing tolerance and the acclimatization of photosynthesis to low temperature is revealed by the pho mutants of Arabidopsis thaliana. Plant J. 2000, 24, 383–396. [Google Scholar] [CrossRef]
- Ho-Plágaro, T.; Tamayo-Navarrete, M.I.; García-Garrido, J.M. Histochemical Staining and Quantification of Arbuscular Mycorrhizal Fungal Colonization BT. In Arbuscular Mycorrhizal Fungi: Methods and Protocols; Ferrol, N., Lanfranco, L., Eds.; Springer: New York, NY, USA, 2020; pp. 43–52. ISBN 978-1-0716-0603-2. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer International Publishing: Cham, Switzerland, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Mendiburu, F.D. Agricolae: Statistical Procedures for Agricultural Research; R Package Version 1.2-3; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Miller, S.P.; Bever, J.D. Distribution of Arbuscular Mycorrhizal Fungi in Stands of the Wetland Grass Panicum hemitomon along a Wide Hydrologic Gradient. Oecologia 1999, 119, 586–592. [Google Scholar] [CrossRef]
- Fabián, D.; Guadarrama, P.; Hernadez-Cuevas, L.; Ramos-Zapata, J.A. Arbuscular mycorrhizal fungi in a coastal wetland in Yucatan, Mexico. Bot. Sci. 2018, 96, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Asano, K.; Kagong, W.V.; Mohammad, S.M.; Sakazaki, K.; Talip, M.S.; Sahmat, S.S.; Chan, M.K.; Isoi, T.; Kano-Nakata, M.; Ehara, H. Arbuscular Mycorrhizal Communities in the Roots of Sago Palm in Mineral and Shallow Peat Soils. Agriculture 2021, 11, 1161. [Google Scholar] [CrossRef]
- Yang, Z.; Shi, Y.; Wang, J.; Wang, L.; Li, X.; Zhang, D. Unique functional responses of fungal communities to various environments in the mangroves of the Maowei Sea in Guangxi, China. Mar. Pollut. Bull. 2021, 173, 113091. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A Place for DNA-DNA Reassociation and 16S rRNA Sequence Analysis in the Present Species Definition in Bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- House, G.L.; Ekanayake, S.; Ruan, Y.; Schütte, U.M.E.; Kaonongbua, W.; Fox, G.; Ye, Y.; Bever, J.D. Phylogenetically Structured Differences in rRNA Gene Sequence Variation among Species of Arbuscular Mycorrhizal Fungi and Their Implications for Sequence Clustering. Appl. Environ. Microbiol. 2016, 82, 4921–4930. [Google Scholar] [CrossRef] [Green Version]
- Maeda, T.; Kobayashi, Y.; Kameoka, H.; Okuma, N.; Takeda, N.; Yamaguchi, K.; Bino, T.; Shigenobu, S.; Kawaguchi, M. Evidence of non-tandemly repeated rDNAs and their intragenomic heterogeneity in Rhizophagus irregularis. Commun. Biol. 2018, 1, 87. [Google Scholar] [CrossRef] [Green Version]
- Gutjahr, C.; Paszkowski, U. Multiple Control Levels of Root System Remodeling in Arbuscular Mycorrhizal Symbiosis. Front. Plant Sci. 2013, 4, 204. [Google Scholar] [CrossRef] [Green Version]
- Pinit, S.; Chadchawan, S.; Chaiwanon, J. A simple high-throughput protocol for the extraction and quantification of inorganic phosphate in rice leaves. Appl. Plant Sci. 2020, 8, e11395. [Google Scholar] [CrossRef]
- Uwasawa, M.; Sangtong, P.; Cholitkul, W. Behavior of phosphorus in paddy soils of Thailand. Soil Sci. Plant Nutr. 1988, 34, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Phairat, P.; Toru, M. The status of phosphorus in Thai soils and P evaluation using EDTA-NaF extraction method. Songklanakarin J. Sci. Technol. 2003, 25, 423–434. [Google Scholar]
- Graham, J.H.; Abbott, L.K. Wheat responses to aggressive and non-aggressive arbuscular mycorrhizal fungi. Plant Soil 2000, 220, 207–218. [Google Scholar] [CrossRef]
- Friede, M.; Unger, S.; Hellmann, C.; Beyschlag, W. Conditions Promoting Mycorrhizal Parasitism Are of Minor Importance for Competitive Interactions in Two Differentially Mycotrophic Species. Front. Plant Sci. 2016, 7, 1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, S.F.; Yu, J.Q.; Peng, Y.H.; Zheng, J.H.; Zou, L.Y. Incidence of Fusarium wilt in Cucumis sativus L. is promoted by cinnamic acid, an autotoxin in root exudates. Plant Soil 2004, 263, 143–150. [Google Scholar] [CrossRef]
- Sayago, P.; Juncosa, F.; Orio, A.G.A.; Luna, D.F.; Molina, G.; Lafi, J.; Ducasse, D.A. Bacillus subtilis ALBA01 alleviates onion pink root by antagonizing the pathogen Setophoma terrestris and allowing physiological status maintenance. Eur. J. Plant Pathol. 2020, 157, 509–519. [Google Scholar] [CrossRef]
- Deng, S.; Lu, L.; Li, J.; Du, Z.; Liu, T.; Li, W.; Xu, F.; Shi, L.; Shou, H.; Wang, C. Purple acid phosphatase 10c encodes a major acid phosphatase that regulates plant growth under phosphate-deficient conditions in rice. J. Exp. Bot. 2020, 71, 4321–4332. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Ezawa, T.; Cheng, W.; Tawaraya, K. Release of acid phosphatase from extraradical hyphae of arbuscular mycorrhizal fungus Rhizophagus clarus. Soil Sci. Plant Nutr. 2015, 61, 269–274. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nopphakat, K.; Runsaeng, P.; Klinnawee, L. Acaulospora as the Dominant Arbuscular Mycorrhizal Fungi in Organic Lowland Rice Paddies Improves Phosphorus Availability in Soils. Sustainability 2022, 14, 31. https://doi.org/10.3390/su14010031
Nopphakat K, Runsaeng P, Klinnawee L. Acaulospora as the Dominant Arbuscular Mycorrhizal Fungi in Organic Lowland Rice Paddies Improves Phosphorus Availability in Soils. Sustainability. 2022; 14(1):31. https://doi.org/10.3390/su14010031
Chicago/Turabian StyleNopphakat, Khachonphong, Phanthipha Runsaeng, and Lompong Klinnawee. 2022. "Acaulospora as the Dominant Arbuscular Mycorrhizal Fungi in Organic Lowland Rice Paddies Improves Phosphorus Availability in Soils" Sustainability 14, no. 1: 31. https://doi.org/10.3390/su14010031