
Duckweed (Lemna minor L.) Successfully Accumulates Selenium from Selenium-Impacted Water


Abstract
:1. Introduction
2. Materials and Methods
2.1. Common Duckweed Cultivation and Exposure
2.2. Physiological Analyses
2.3. Biochemical Analyses
2.4. Growth Parameters
2.5. Concentrations of Selenium, Calcium and Potassium
2.6. Statistical Analysis
3. Results
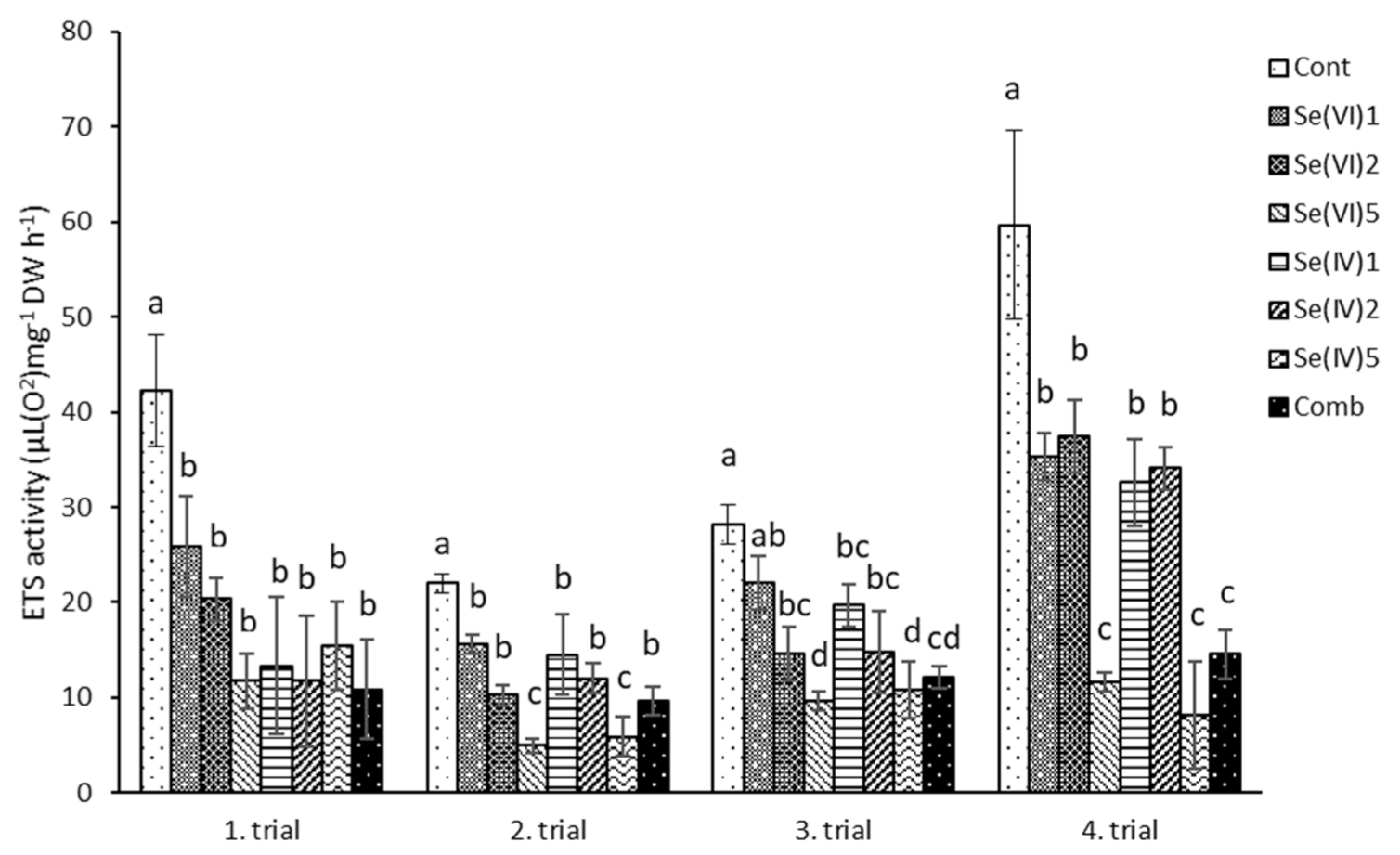
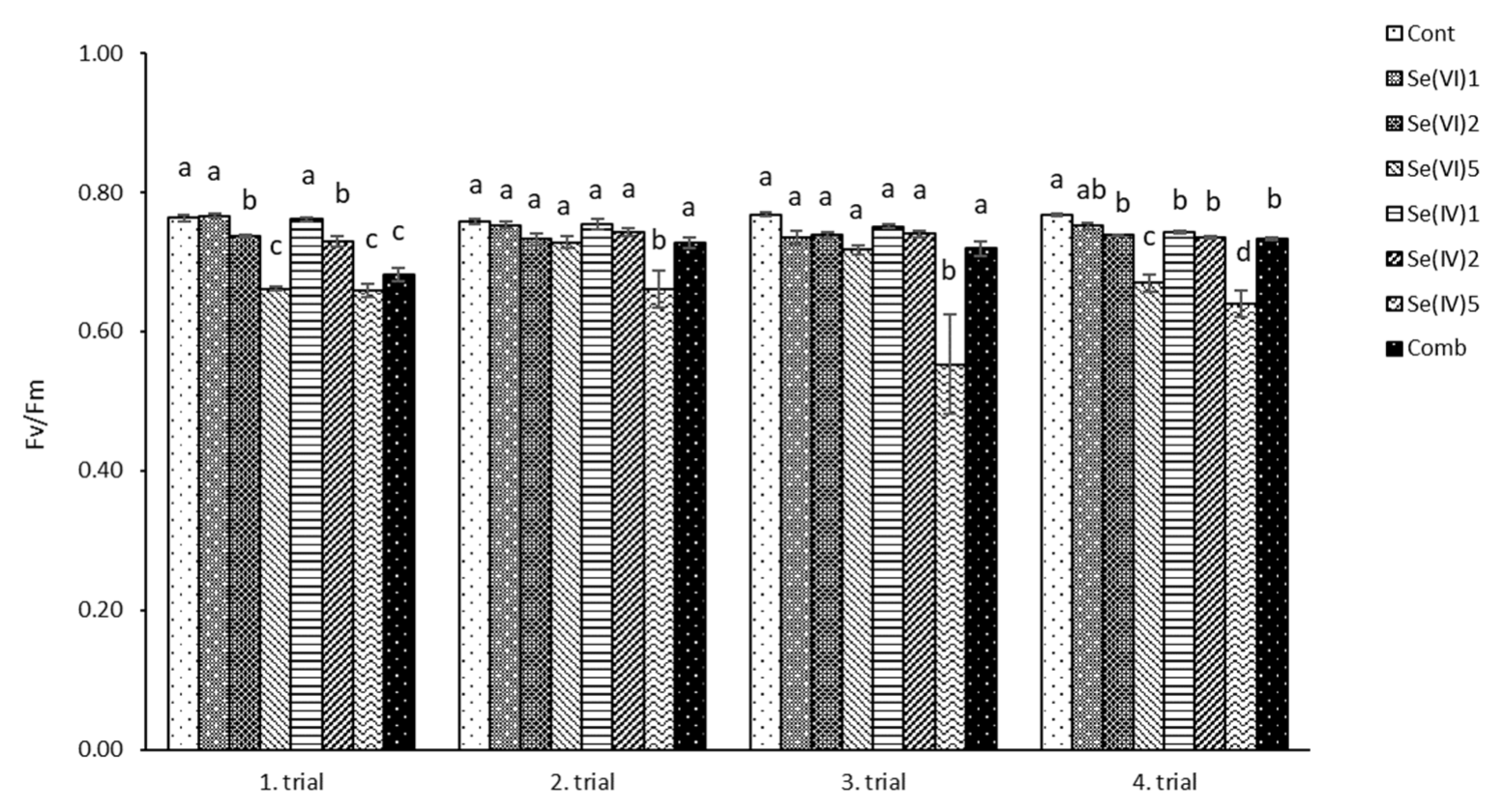
3.1. Physiological Measurements
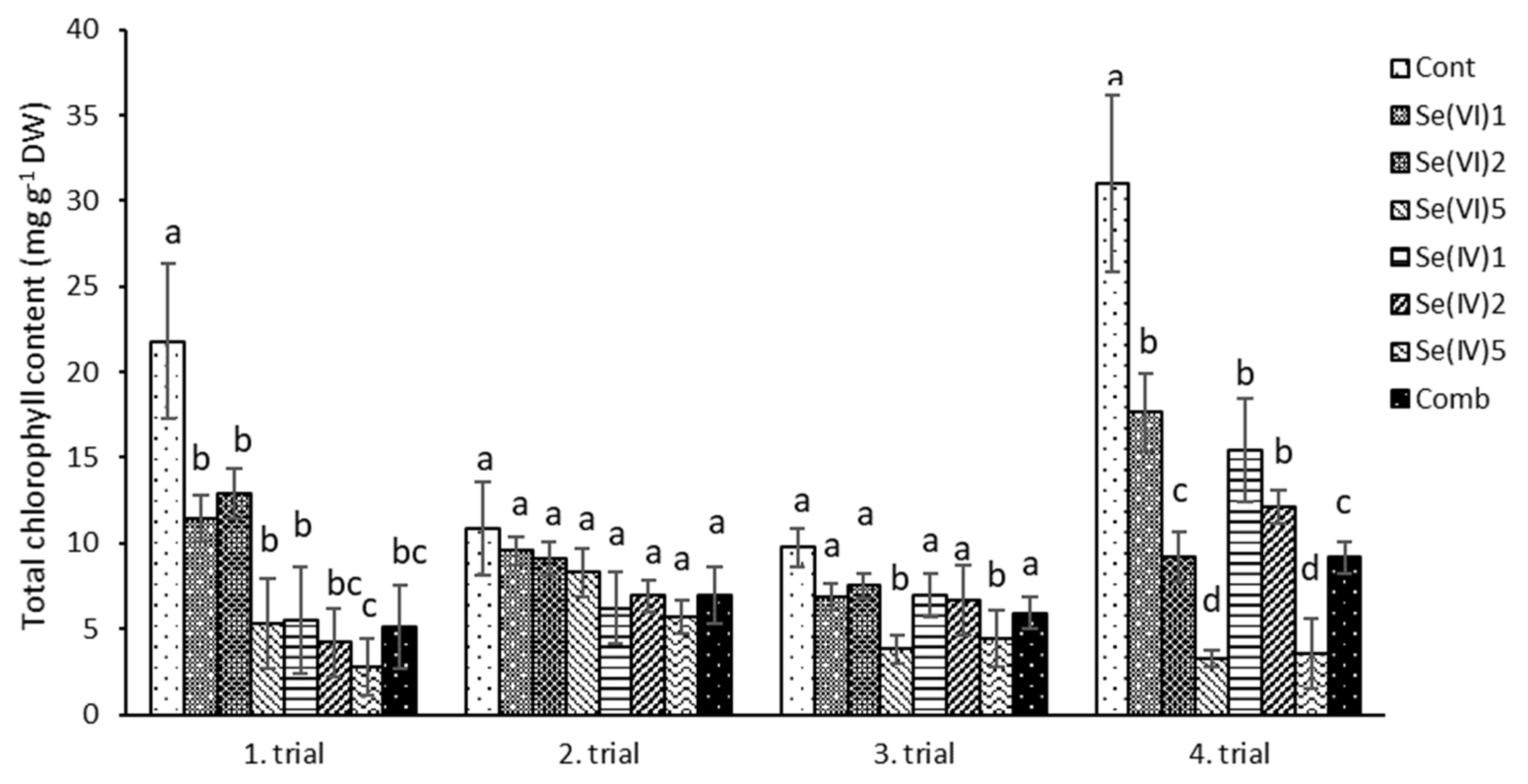
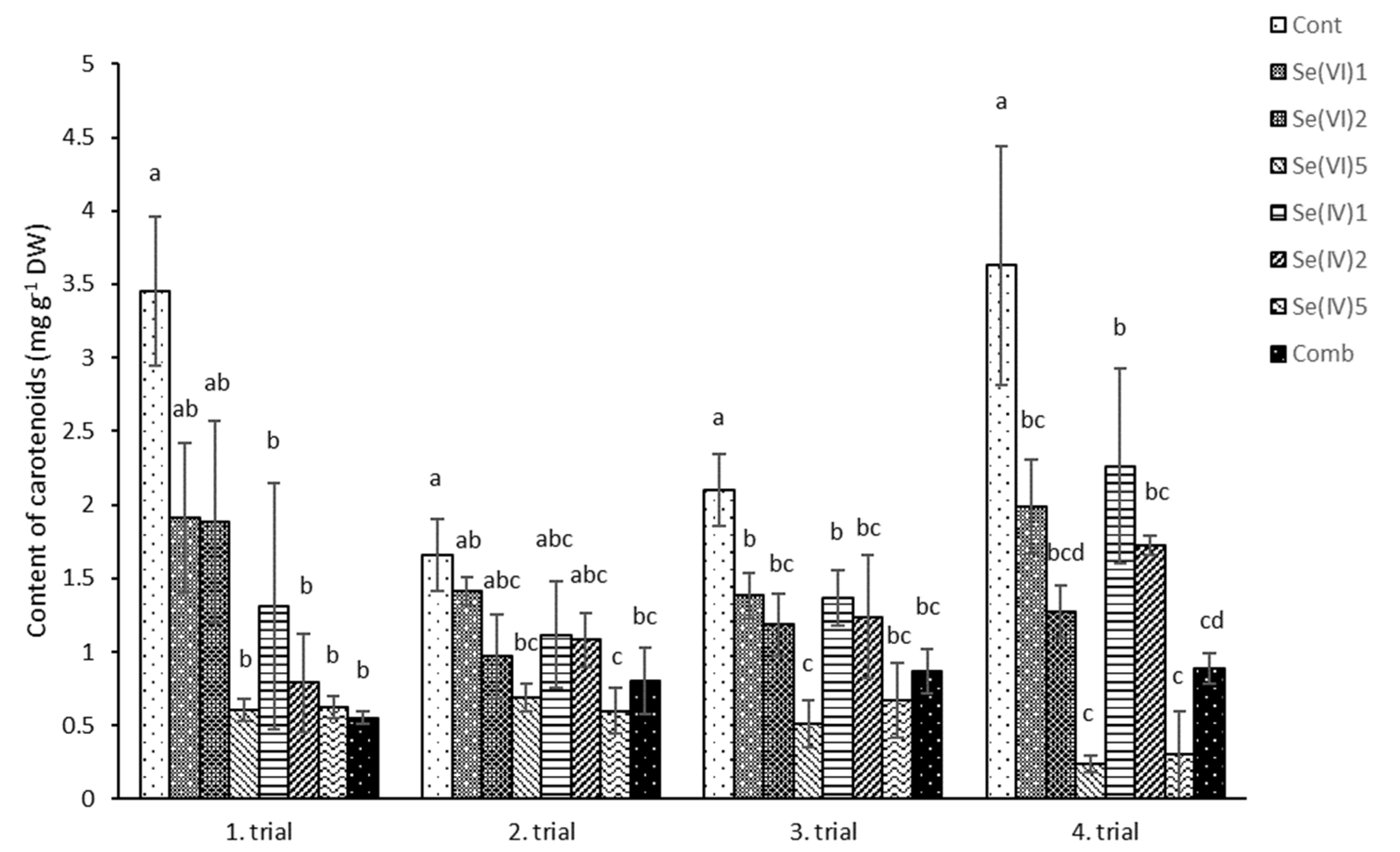
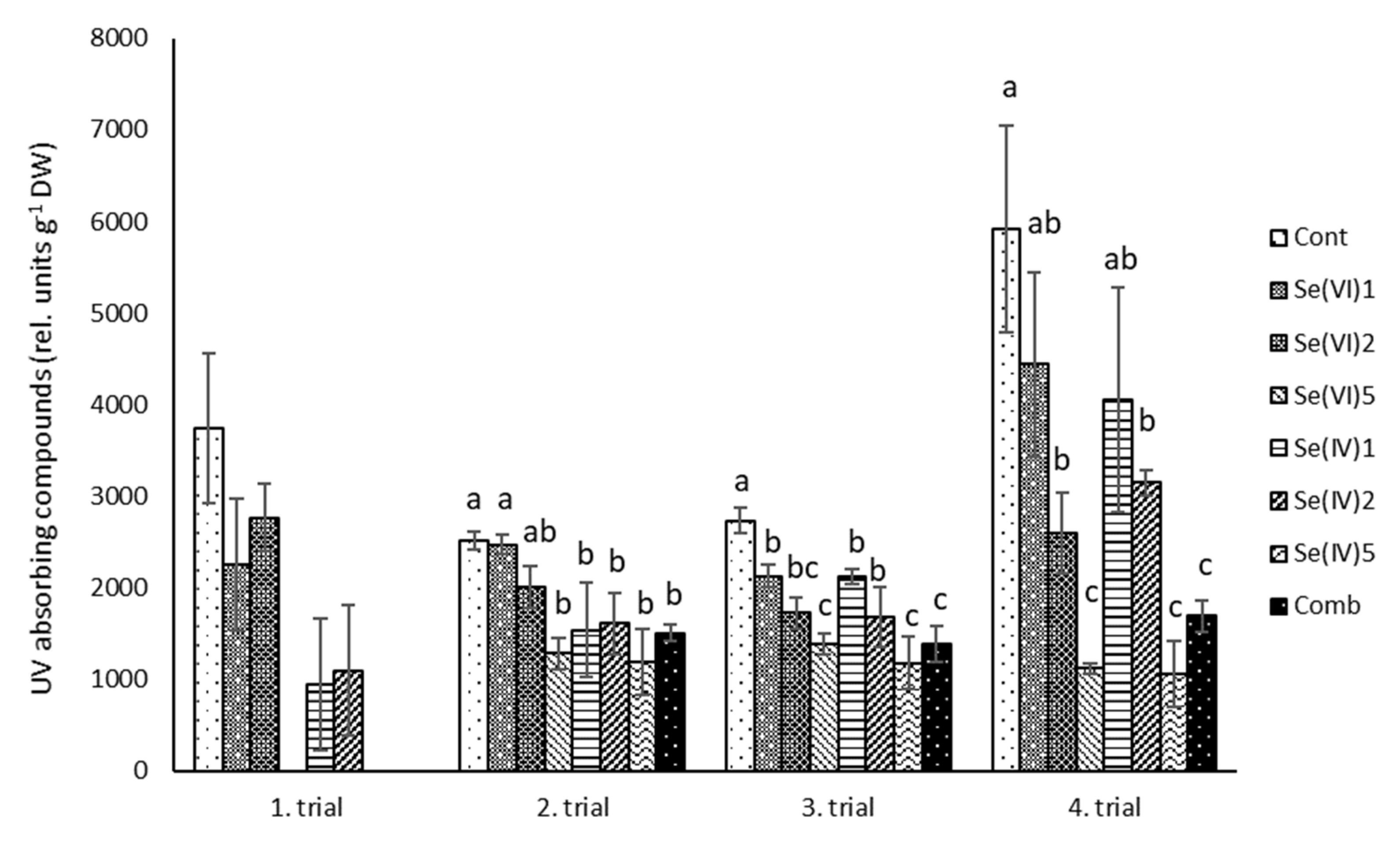
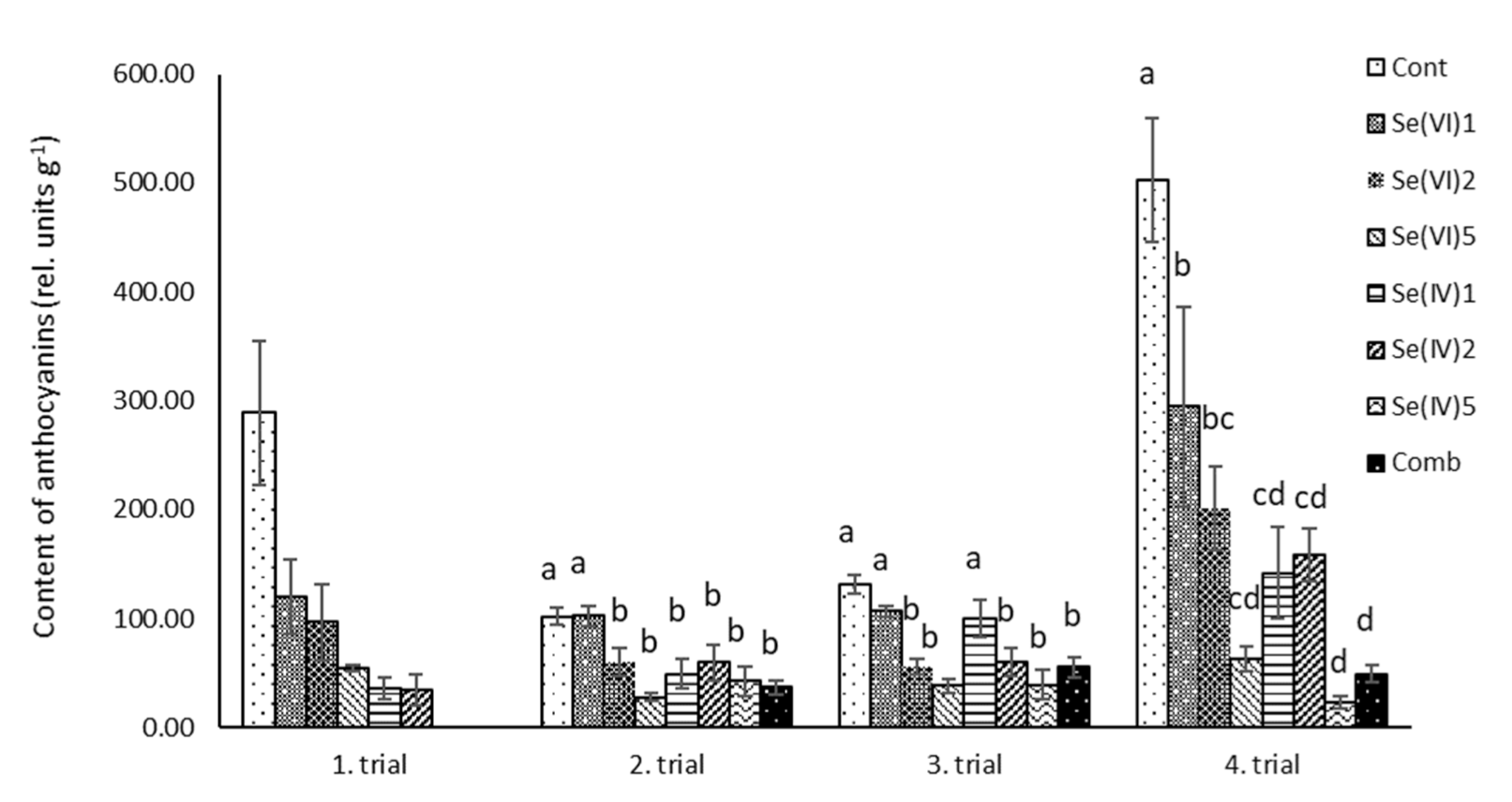
3.2. Biochemical Measurements
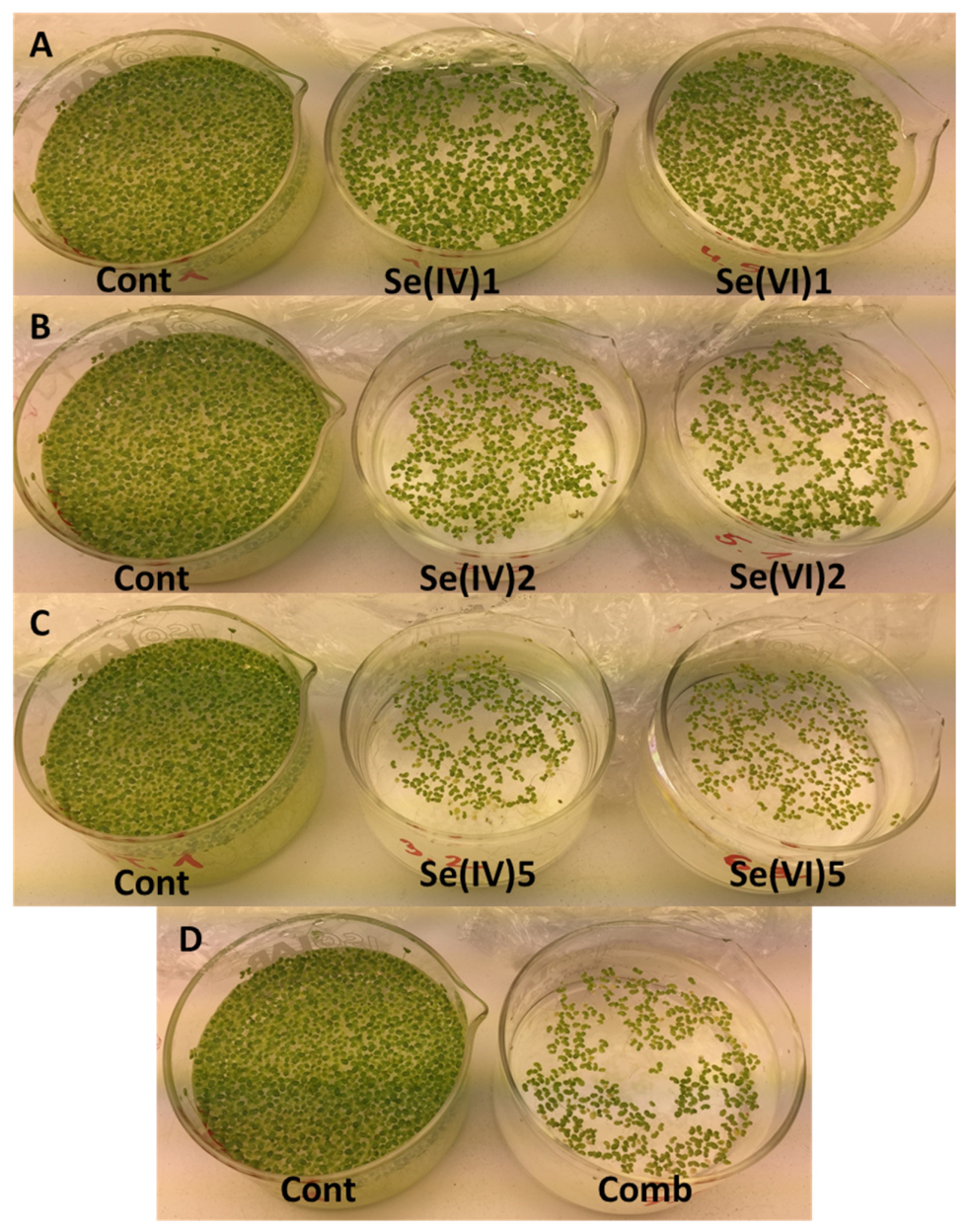
3.3. Growth Parameters
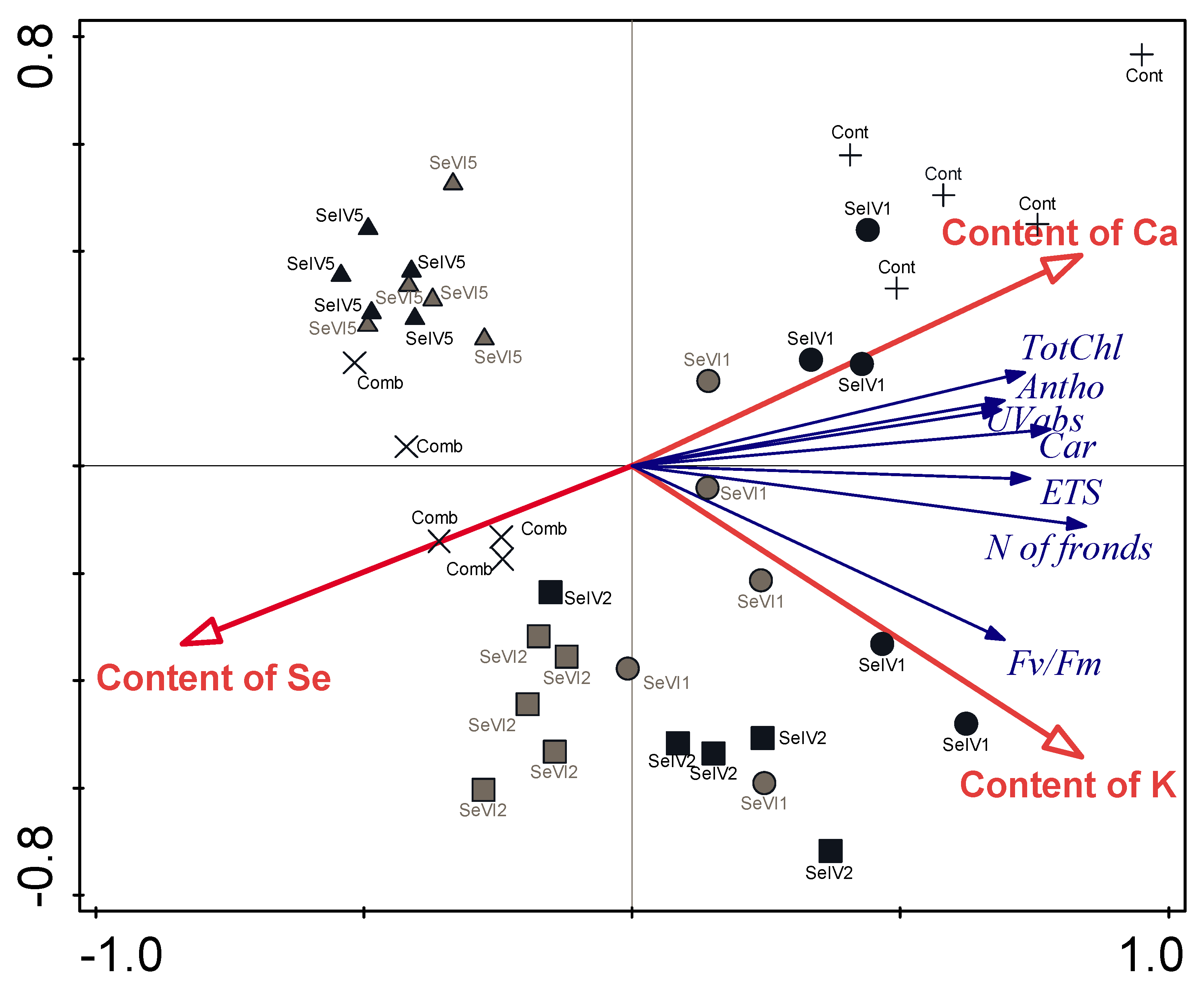
3.4. Concentrations of Elements
4. Discussion
4.1. Physiological Characteristics
4.2. Biochemical Characteristics
4.3. Growth Characteristics
4.4. Elemental Composition of L. Minor
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ellis, D.R.; Salt, D.E. Plants, selenium and human health. Curr. Opin. Plant Biol. 2003, 6, 273–279. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.B.; Raza, A.; Hawrylak-Nowak, B.; Matraszek-Gawron, R.; Al Mahmud, J.; Nahar, K.; Fujita, M. Selenium in plants: Boon or bane? Environ. Exp. Bot. 2020, 178, 104170. [Google Scholar] [CrossRef]
- Naseem, M.; Anwar-Ul-Haq, M.; Wang, X.; Farooq, N.; Awais, M.; Sattar, H.; Malik, H.A.; Mustafa, A.; Ahmad, J.; El-Esawi, M.A. Influence of Selenium on Growth, Physiology, and Antioxidant Responses in Maize Varies in a Dose-Dependent Manner. J. Food Qual. 2021, 2021, 1–9. [Google Scholar] [CrossRef]
- Seppänen, M.; Turakainen, M.; Hartikainen, H. Selenium effects on oxidative stress in potato. Plant Sci. 2003, 165, 311–319. [Google Scholar] [CrossRef]
- Abbas, S.M. Effects of low temperature and selenium application on growth and the physiological changes in sorghum seedlings. J. Stress Physiol. Biochem. 2012, 8, 268–286. [Google Scholar]
- Hasanuzzaman, M.; Hossain, M.A.; Fujita, M. Selenium-Induced Up-Regulation of the Antioxidant Defense and Methylglyoxal Detoxification System Reduces Salinity-Induced Damage in Rapeseed Seedlings. Biol. Trace Element Res. 2011, 143, 1704–1721. [Google Scholar] [CrossRef] [PubMed]
- Hawrylak-Nowak, B.; Dresler, S.; Wójcik, M. Selenium affects physiological parameters and phytochelatins accumulation in cucumber (Cucumis sativus L.) plants grown under cadmium exposure. Sci. Hortic. 2014, 172, 10–18. [Google Scholar] [CrossRef]
- Nawaz, F.; Ashraf, M.; Ahmad, R.; Waraich, E.; Shabbir, R.; Bukhari, M. Supplemental selenium improves wheat grain yield and quality through alterations in biochemical processes under normal and water deficit conditions. Food Chem. 2015, 175, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; Taylor & Francis Group: London, UK, 2010; p. 548. Available online: https://doi.org/10.1201/b10158 (accessed on 18 October 2010).
- Severi, A. Toxicity of selenium to Lemna minor in relation to sulfate concentration. Physiol. Plant. 2001, 113, 523–532. [Google Scholar] [CrossRef]
- Landrigan, P.J.; Fuller, R.; Acosta, N.J.R.; Adeyi, O.; Arnold, R.; Basu, N.N.; Baldé, A.B.; Bertollini, R.; Bose-O’Reilly, S.; Boufford, J.I.; et al. The Lancet Commission on pollution and health. Lancet 2018, 391, 462–512. [Google Scholar] [CrossRef] [Green Version]
- Haug, A.; Graham, R.D.; Christophersen, O.A.; Lyons, G.H. How to use the world’s scarce selenium resources efficiently to increase the selenium concentration in food. Microb. Ecol. Health Dis. 2007, 19, 209–228. [Google Scholar] [CrossRef] [Green Version]
- Winkel, L.H.E.; Johnson, C.A.; Lenz, M.; Grundl, T.; Leupin, O.X.; Amini, M.; Charlet, L. Environmental Selenium Research: From Microscopic Processes to Global Understanding. Environ. Sci. Technol. 2011, 46, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Alarcon, M.; Cabrera-Vique, C. Selenium in food and the human body: A review. Sci. Total. Environ. 2008, 400, 115–141. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.; Ungureanu, G.; Boaventura, R.; Botelho, C. Selenium contaminated waters: An overview of analytical methods, treatment options and recent advances in sorption methods. Sci. Total. Environ. 2015, 521-522, 246–260. [Google Scholar] [CrossRef]
- Zhou, C.; Huang, J.-C.; Liu, F.; He, S.; Zhou, W. Selenium removal and biotransformation in a floating-leaved macrophyte system. Environ. Pollut. 2019, 245, 941–949. [Google Scholar] [CrossRef]
- Dhote, S.; Dixit, S. Water quality improvement through macrophytes—a review. Environ. Monit. Assess. 2009, 152, 149–153. [Google Scholar] [CrossRef]
- Pistone, G.; Allinson, G.; Stagnitti, F.; Colville, S. Effect of selenium on the growth ofLemna minor. Toxicol. Environ. Chem. 1999, 71, 271–277. [Google Scholar] [CrossRef]
- Ziegler, P.; Sree, K.; Appenroth, K.-J. Duckweeds for water remediation and toxicity testing. Toxicol. Environ. Chem. 2016, 98, 1127–1154. [Google Scholar] [CrossRef]
- Ghanem, H.; Haddad, A.; Baydoun, S.; Hamdan, H.A.; Korfali, S.; Chalak, L. In vitro proliferation of Lebanese Lemna minor and Lemna gibba on different nutrient media. J. Taibah Univ. Sci. 2019, 13, 497–503. [Google Scholar] [CrossRef]
- Forni, C.; Tommasi, F. Duckweed: A Tool for Ecotoxicology and a Candidate for Phytoremediation. Curr. Biotechnol. 2016, 5, 2–10. [Google Scholar] [CrossRef]
- Carvalho, K.; Martin, D. Removal of Aqueous Selenium by Four Aquatic Plants. J. Aquat. Plant Manag. 2001, 39, 33–36. [Google Scholar]
- Nattrass, M.; McGrew, N.R.; Morrison, J.I.; Baldwin, B.S. Phytoremediation of selenium-impacted water by aquatic macrophytes. J. Am. Soc. Min. Reclam. 2019, 8, 69–79. [Google Scholar] [CrossRef]
- Vladimirova, I.N.; Georgiyants, V.A. Biologically Active Compounds from Lemna Minor S.F. Gray. Pharm. Chem. J. 2014, 47, 599–601. [Google Scholar] [CrossRef]
- Kastratović, V.; Jaćimović, Ž.; Đurović, D.; Bigović, M.; Krivokapić, S. Lemna minor L. As Bioindicator Of Heavy Metal Pollution In Skadar Lake: Montenegro. Kragujev. J. Sci. 2015, 37, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Mo, F.; Li, Y.; Wang, M.; Li, Z.; Hu, H.; Deng, W.; Zhang, R. Effects of silver(I) toxicity on microstructure, biochemical activities, and genic material of Lemna minor L. with special reference to application of bioindicator. Environ. Sci. Pollut. Res. 2020, 27, 22735–22748. [Google Scholar] [CrossRef]
- Pollard, J.; Cizdziel, J.; Stave, K.; Reid, M. Selenium concentrations in water and plant tissues of a newly formed arid wetland in Las Vegas, Nevada. Environ. Monit. Assess. 2007, 135, 447–457. [Google Scholar] [CrossRef]
- Delmail, D.; Labrousse, P.; Hourdin, P.; Larcher, L.; Moesch, C.; Botineau, M. Physiological, anatomical and phenotypical effects of a cadmium stress in different-aged chlorophyllian organs of Myriophyllum alterniflorum DC (Haloragaceae). Environ. Exp. Bot. 2011, 72, 174–181. [Google Scholar] [CrossRef]
- Mechora, Š; Stibilj, V.; Germ, M. Response of duckweed to various concentrations of selenite. Environ. Sci. Pollut. Res. 2014, 22, 2416–2422. [Google Scholar] [CrossRef]
- Mechora, S.; Stibilj, V.; Germ, M. The uptake and distribution of selenium in three aquatic plants grown in Se(IV) solution. Aquat. Toxicol. 2013, 128-129, 53–59. [Google Scholar] [CrossRef]
- Mechora, Š; Cuderman, P.; Stibilj, V.; Germ, M. Distribution of Se and its species in Myriophyllum spicatum and Ceratophyllum demersum growing in water containing se (VI). Chemosphere 2011, 84, 1636–1641. [Google Scholar] [CrossRef] [PubMed]
- Packard, T.T. The Measurement of Respiratory Electron-transport Activity tn Marine Phytoplankton. J. Mar. Res. 1971, 29, 235–243. [Google Scholar]
- Kenner, R.A.; Ahmed, S.I. Measurements of electron transport activities in marine phytoplankton. Mar. Biol. 1975, 33, 119–127. [Google Scholar] [CrossRef]
- Schreiber, U.; Bilger, W.; Neubauer, C. Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. In Ecophysiology of Photosynthesis; Springer: Berlin/Heidelberg, Germany, 1995; pp. 49–70. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Drumm, H.; Mohr, H. The mode of interaction between blue (uv) light photoreceptor and phytochrome in anthocyanin formation of the sorghum seedling. Photochem. Photobiol. 1978, 27, 241–248. [Google Scholar] [CrossRef]
- Caldwell, M.M. Solar Ultraviolet Radiation as an Ecological Factor for Alpine Plants. Ecol. Monogr. 1968, 38, 243–268. [Google Scholar] [CrossRef]
- Kump, P.; Nečemer, M.; Rupnik, Z.; Pelicon, P.; Ponikvar, D.; Vogel-Mikuš, K.; Regvar, M.; Pongrac, P. Improvement of the XRF Quantification and Enhancement of the Combined Applications by EDXRF and Micro PIXE. 2011. Available online: https://inis.iaea.org/search/search.aspx?orig_q=RN:43021135 (accessed on 15 October 2021).
- Nečemer, M.; Kump, P.; Ščančar, J.; Jaćimović, R.; Simčič, J.; Pelicon, P.; Budnar, M.; Jeran, Z.; Pongrac, P.; Regvar, M.; et al. Application of X-ray fluorescence analytical techniques in phytoremediation and plant biology studies. Spectrochim. Acta Part B At. Spectrosc. 2008, 63, 1240–1247. [Google Scholar] [CrossRef]
- Zayed, A.; Gowthaman, S.; Terry, N. Phytoaccumulation of Trace Elements by Wetland Plants: I. Duckweed. J. Environ. Qual. 1998, 27, 715–721. [Google Scholar] [CrossRef]
- Ter Braak, C.; Šmilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (version 4.5). 2002. Available online: https://research.wur.nl/en/publications/canoco-reference-manual-and-canodraw-for-windows-users-guide-soft (accessed on 11 October 2021).
- Bartoli, C.G.; Gomez, F.; Gergoff, G.; Guiamét, J.J.; Puntarulo, S. Up-regulation of the mitochondrial alternative oxidase pathway enhances photosynthetic electron transport under drought conditions. J. Exp. Bot. 2005, 56, 1269–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germ, M.; Kreft, I.; Osvald, J. Influence of UV-B exclusion and selenium treatment on photochemical efficiency of photosystem II, yield and respiratory potential in pumpkins (Cucurbita pepo L.). Plant Physiol. Biochem. 2005, 43, 445–448. [Google Scholar] [CrossRef]
- Smrkolj, P.; Germ, M.; Kreft, I.; Stibilj, V. Respiratory potential and Se compounds in pea (Pisum sativum L.) plants grown from Se-enriched seeds. J. Exp. Bot. 2006, 57, 3595–3600. [Google Scholar] [CrossRef] [Green Version]
- Terry, N.; Zayed, A.M.; de Souza, M.P.; Tarun, A.S. Selenium in higher plants. Annu. Rev. Plant Biol. 2000, 51, 401–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechora, Š.; Germ, M. Selenium induced lower respiratory potential in Glycine max (L.) Merr. Acta Agric. Slov. 2010, 95, 29–34. [Google Scholar] [CrossRef]
- White, P.J. Selenium metabolism in plants. Biochim. et Biophys. Acta (BBA) Gen. Subj. 2018, 1862, 2333–2342. [Google Scholar] [CrossRef] [PubMed]
- Tarrahi, R.; Khataee, A.; Movafeghi, A.; Rezanejad, F.; Gohari, G. Toxicological implications of selenium nanoparticles with different coatings along with Se 4+ on Lemna minor. Chemosphere 2017, 181, 655–665. [Google Scholar] [CrossRef]
- Akbulut, M.; Çakır, S. The effects of Se phytotoxicity on the antioxidant systems of leaf tissues in barley (Hordeum vulgare L.) seedlings. Plant Physiol. Biochem. 2010, 48, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Vítová, M.; Bišová, K.; Hlavová, M.; Zachleder, V.; Rucki, M.; Čížková, M. Glutathione peroxidase activity in the selenium-treated alga Scenedesmus quadricauda. Aquat. Toxicol. 2011, 102, 87–94. [Google Scholar] [CrossRef]
- Šircelj, H. Karotenoidi v Fotosinteznem Aparatu in Odziv na Stres. 2008. Available online: https://repozitorij.uni-lj.si/IzpisGradiva.php?id=17642&lang=eng (accessed on 15 October 2021).
- Hawrylak-Nowak, B. Changes in Anthocyanin Content as Indicator of Maize Sensitivity to Selenium. J. Plant Nutr. 2008, 31, 1232–1242. [Google Scholar] [CrossRef]
- Charron, C.S.; A Kopsell, D.; Randle, W.M.; E Sams, C. Sodium selenate fertilisation increases selenium accumulation and decreases glucosinolate concentration in rapid-cyclingBrassica oleracea. J. Sci. Food Agric. 2001, 81, 962–966. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef] [Green Version]
- Besser, J.M.; Huckins, J.N.; Little, E.E.; La Point, T.W. Distribution and bioaccumulation of selenium in aquatic microcosms. Environ. Pollut. 1989, 62, 1–12. [Google Scholar] [CrossRef]
- Kacjan Maršič, N.; Golob, A.; Šircelj, H.; Mihorič, M.; Kroflič, A.; Stibilj, V.; Germ, M. Effects of Exogenous Selenium in Different Concentrations and Forms on Selenium Accumulation and Growth of Spinach (Spinacia oleracea L.). J. Agric. Sci. Technol 2019, 21, 1905–1917. [Google Scholar]
- Thor, K. Calcium—Nutrient and Messenger. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Du, X.; Wang, F.; Sha, J.; Chen, Q.; Tian, G.; Zhu, Z.; Ge, S.; Jiang, Y. Effects of Potassium Levels on Plant Growth, Accumulation and Distribution of Carbon, and Nitrate Metabolism in Apple Dwarf Rootstock Seedlings. Front. Plant Sci. 2020, 11, 904. [Google Scholar] [CrossRef] [PubMed]
- Keppeler, E.C. Toxicity of sodium chloride and methyl parathion on the macrophyte Lemna minor (Linnaeus, 1753) with respect to frond number and chlorophyll. Biotemas 2011, 22, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Forni, C.; Braglia, R.; Harren, F.; Cristescu, S. Stress responses of duckweed (Lemna minor L.) and water velvet (Azolla filiculoides Lam.) to anionic surfactant sodium-dodecyl-sulphate (SDS). Aquat. Toxicol. 2012, 110-111, 107–113. [Google Scholar] [CrossRef]
- Liu, C.; Gu, W.; Dai, Z.; Li, J.; Jiang, H.; Zhang, Q. Boron accumulation by Lemna minor L. under salt stress. Sci. Rep. 2018, 8, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Xiang, Y.; Zhou, Y.; Yang, Y.; Zhang, J.; Huang, H.; Shang, C.; Luo, L.; Gao, J.; Tang, L. Selenium contamination, consequences and remediation techniques in water and soils: A review. Environ. Res. 2018, 164, 288–301. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macroelements | mg L−1 |
| KNO3 | 350.0 |
| Ca(NO3)2·4H2O | 295.0 |
| KH2PO4 | 90.0 |
| K2HPO4 | 12.6 |
| MgSO4·7H2O | 100.0 |
| Macroelements | µg L−1 |
| H3BO3 | 120.0 |
| ZnSO4·7H2O | 180.0 |
| Na2MoO4·2H2O | 44.0 |
| MnCl2·4H2O | 180.0 |
| FeCl3·6H2O | 760.0 |
| EDTA | 1500.0 |
| Treatment | 1. Trial | 2. Trial | 3. Trial | 4. Trial | Bio-Concentration Factor | Fronds with Chlorosis (%) * |
|---|---|---|---|---|---|---|
| Cont | 1 ± 0.042 a | 1 ± 0.028 a | 1 ± 0.031 a | 1 ± 0.007 a | 0.36 ± 0.09 d | |
| Se(VI)1 | 1.20 ± 0.042 a | 0.94 ± 0.027 b | 0.93 ± 0.067 b | 0.93 ± 0.011 b | 153 ± 6 a | 0.91 ± 0.11 d |
| Se(VI)2 | 0.99 ± 0.065 a | 0.78 ± 0.065 c | 0.85 ± 0.043 b | 0,79 ± 0.004 c | 166 ± 7 a | 1.36 ± 0.12 c |
| Se(VI)5 | 0.40 ± 0.036 c | 0.50 ± 0.040 d | 0.59 ± 0.073 cd | 0.51 ± 0.004 e | 55 ± 2 c | 2.17 ± 0.35 bc |
| Se(IV)1 | 1.08 ± 0.056 a | 0.94 ± 0.059 b | 0.98 ± 0.081 ab | 0.92 ± 0.023 b | 99 ± 4 b | 0.78 ± 0.13 d |
| Se(IV)2 | 0.68 ± 0.040 b | 0.68 ± 0.039 c | 0.8 ± 0.107 b | 0.72 ± 0.007 d | 80 ± 2 c | 1.90 ± 0.28 c |
| Se(IV)5 | 0.39 ± 0.014 c | 0.48 ± 0.056 d | 0.47 ± 0.047 d | 0.45 ± 0.002 f | 45 ± 1 d | 8.38 ± 1.66 a |
| Comb | 0.49 ± 0.010 c | 0.49 ± 0.048 d | 0.61 ± 0.033 c | 0.51 ± 0.002 e | 66 ± 1 c | 3.65 ± 0.89 b |
| Treatment | Content of Elements | ||
|---|---|---|---|
| Selenium | Calcium | Potassium | |
| (µg g−1 DW) | (mg g−1 DW) | (mg g−1 DW) | |
| Cont | 1.2 ± 0.1 f | 8.3 ± 0.7 a | 22 ± 0.9 a |
| Se(VI)1 | 153 ± 6.0 d | 5.5 ± 0.3 c | 22 ± 0.9 a |
| Se(VI)2 | 332 ± 7.8 a | 5.3 ± 0.2 cd | 21 ± 0.3 a |
| Se(VI)5 | 277 ± 13.9 b | 5.4 ± 0.4 cd | 14 ± 0.5 cb |
| Se(IV)1 | 99 ± 4.0 e | 7.2 ± 0.4 b | 23 ± 1.4 a |
| Se(IV)2 | 159 ± 4.4 d | 4.8 ± 0.4 cd | 23 ± 1.4 a |
| Se(IV)5 | 227 ± 7.4 c | 4.3 ± 0.2 d | 12 ± 0.4 c |
| Comb | 263 ± 2.8 b | 4.4 ± 0.2 cd | 15 ± 1.0 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golob, A.; Vogel-Mikuš, K.; Brudar, N.; Germ, M. Duckweed (Lemna minor L.) Successfully Accumulates Selenium from Selenium-Impacted Water. Sustainability 2021, 13, 13423. https://doi.org/10.3390/su132313423
Golob A, Vogel-Mikuš K, Brudar N, Germ M. Duckweed (Lemna minor L.) Successfully Accumulates Selenium from Selenium-Impacted Water. Sustainability. 2021; 13(23):13423. https://doi.org/10.3390/su132313423
Chicago/Turabian StyleGolob, Aleksandra, Katarina Vogel-Mikuš, Nina Brudar, and Mateja Germ. 2021. "Duckweed (Lemna minor L.) Successfully Accumulates Selenium from Selenium-Impacted Water" Sustainability 13, no. 23: 13423. https://doi.org/10.3390/su132313423