
Soil Microbial and Nematode Community Response to the Field Application of Recycled Bio-Based Fertilisers in Irish Grassland

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Description and Experimental Design
2.2. Soil Sampling, Physiochemical Properties, and Preparation for DNA Extraction and Sequencing
2.3. Sequence Data Analysis
3. Results
3.1. Soil and Plant Properties
3.2. Microbial Analysis
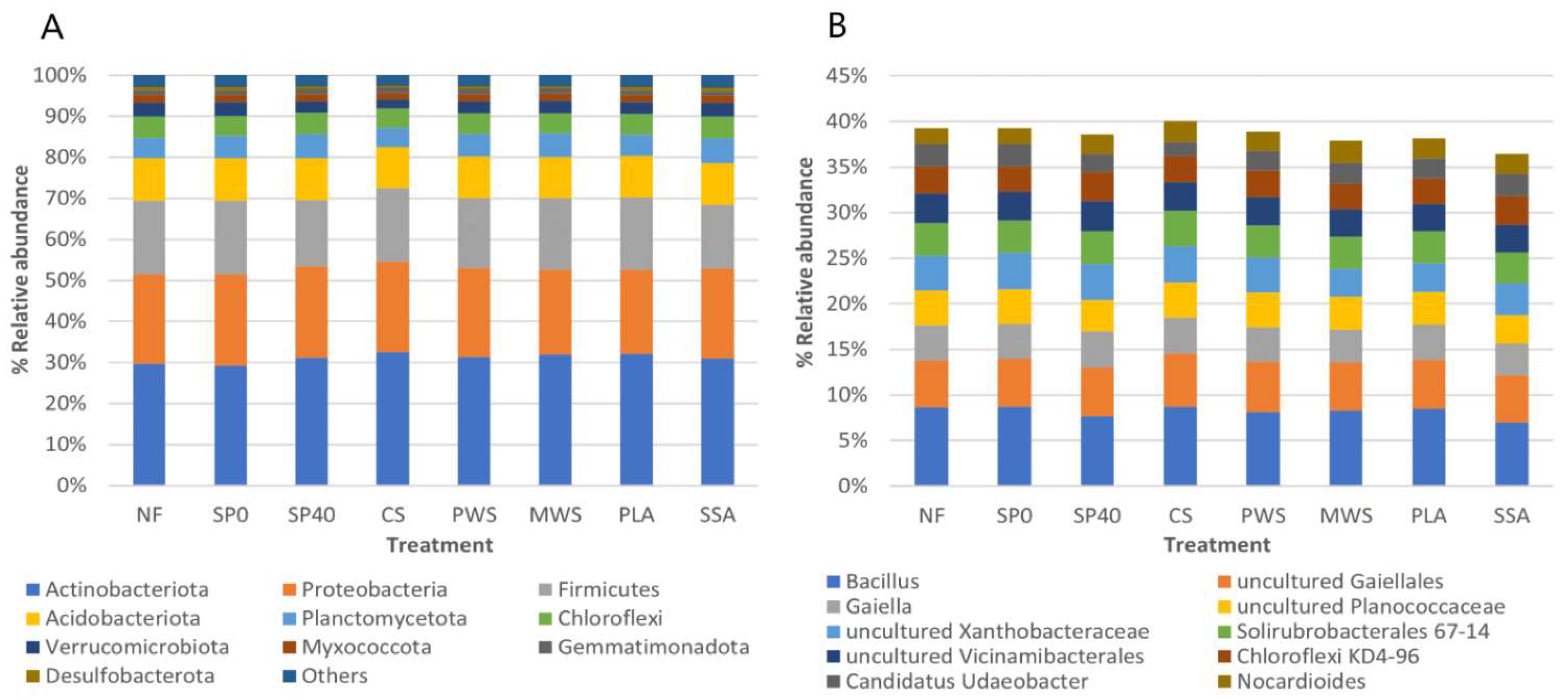
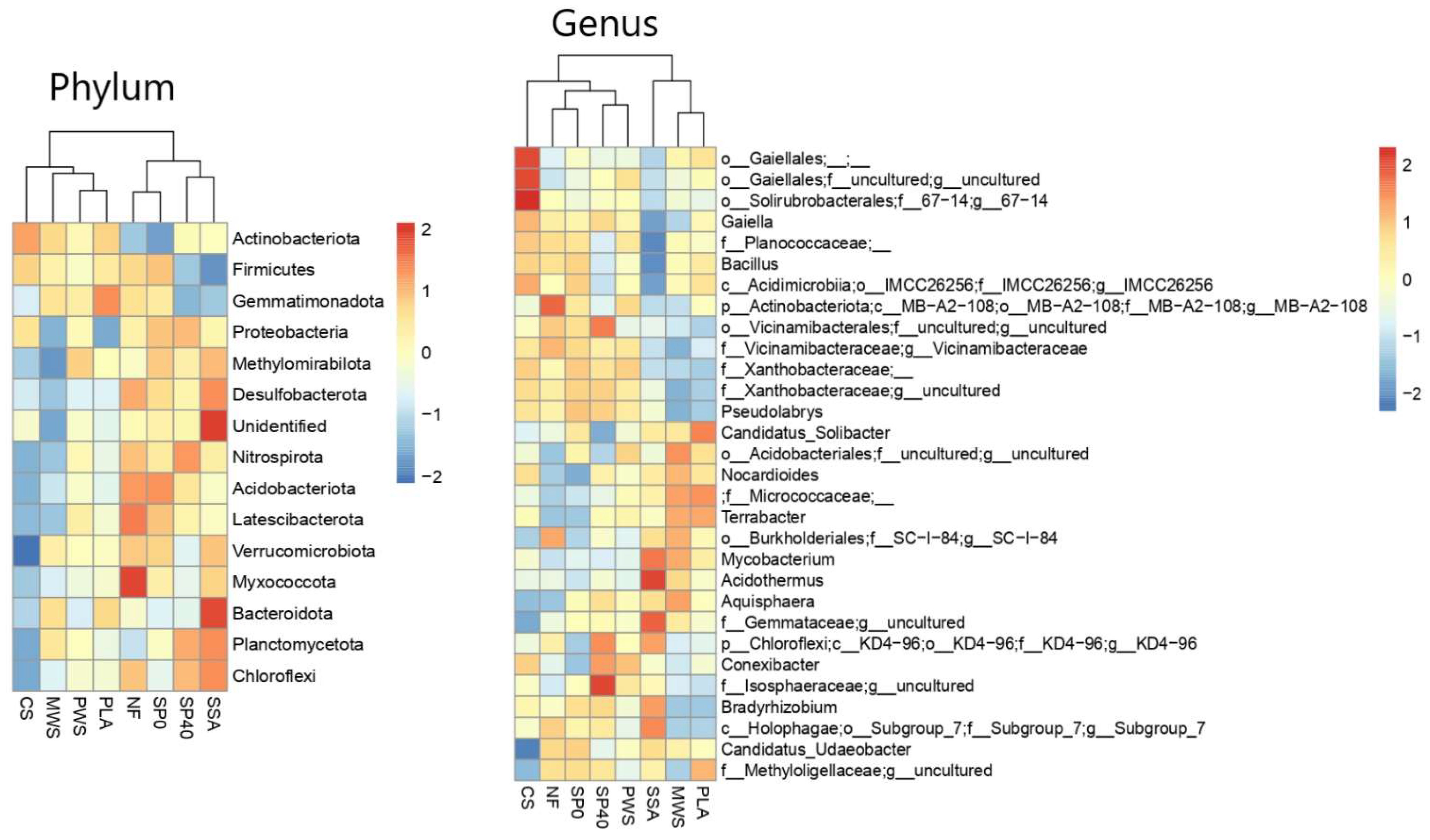
3.2.1. Community Composition
3.2.2. Microbial Alpha Diversity
3.2.3. Microbial Beta Diversity
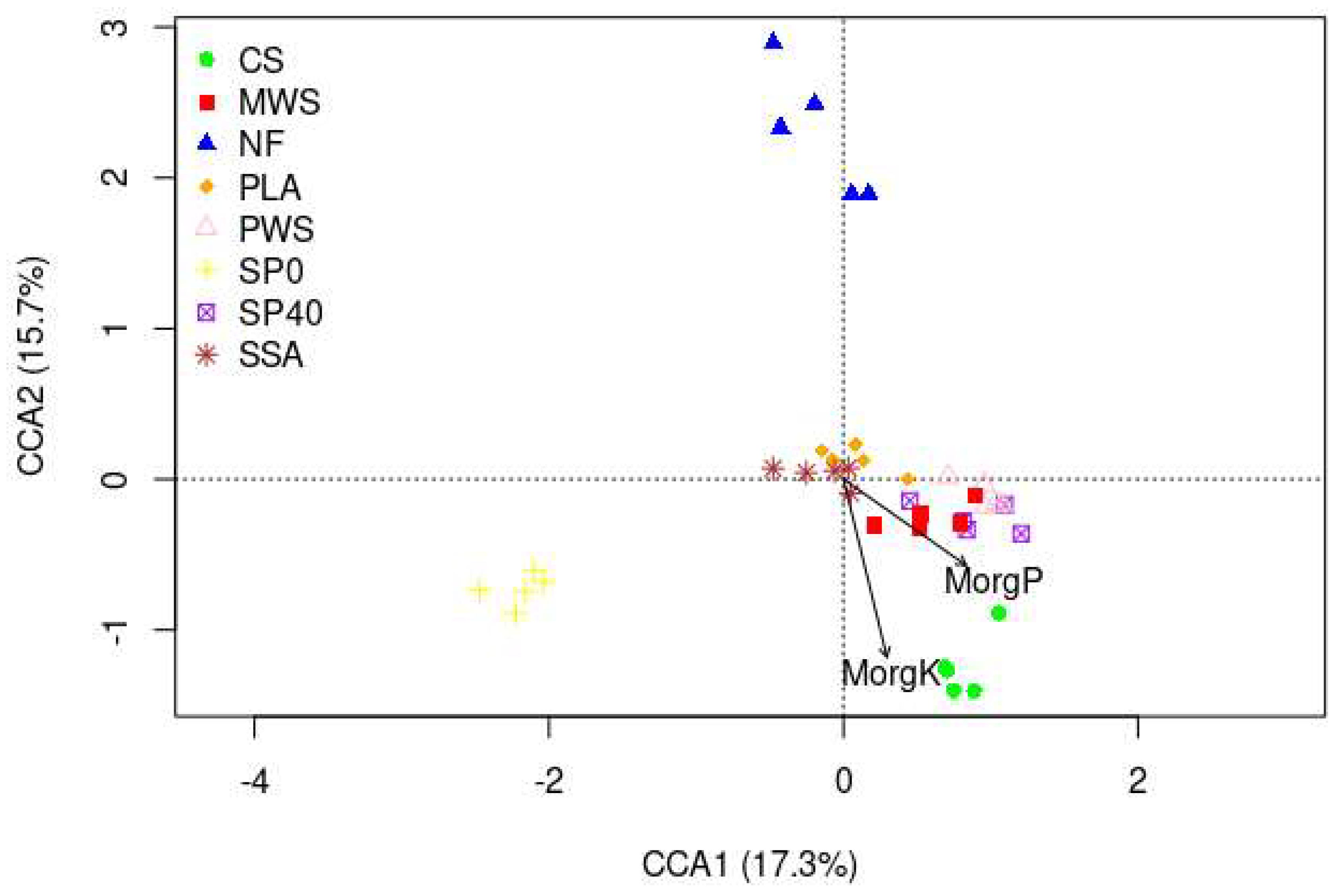
3.2.4. Relationships between Physiochemical Soil Variables and Microbial Communities
3.3. Nematode Analysis
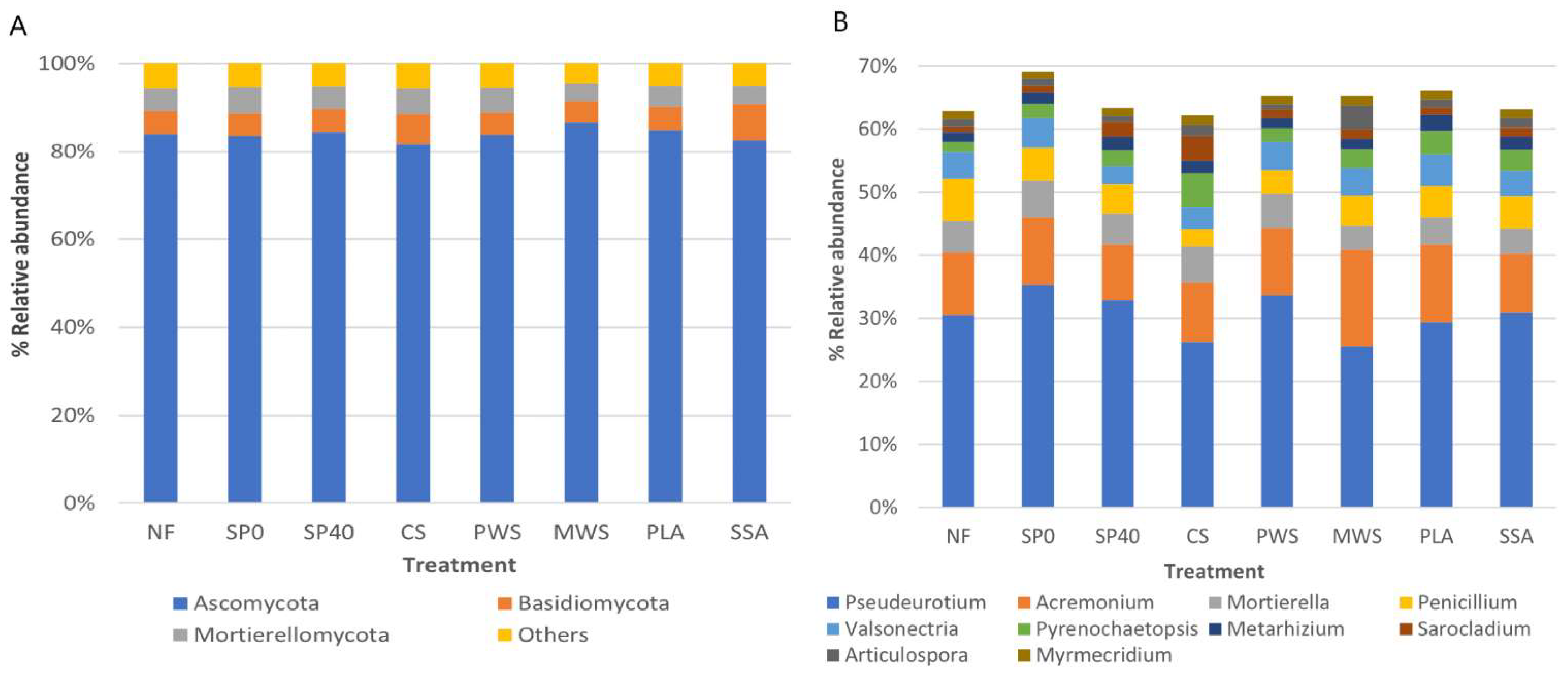
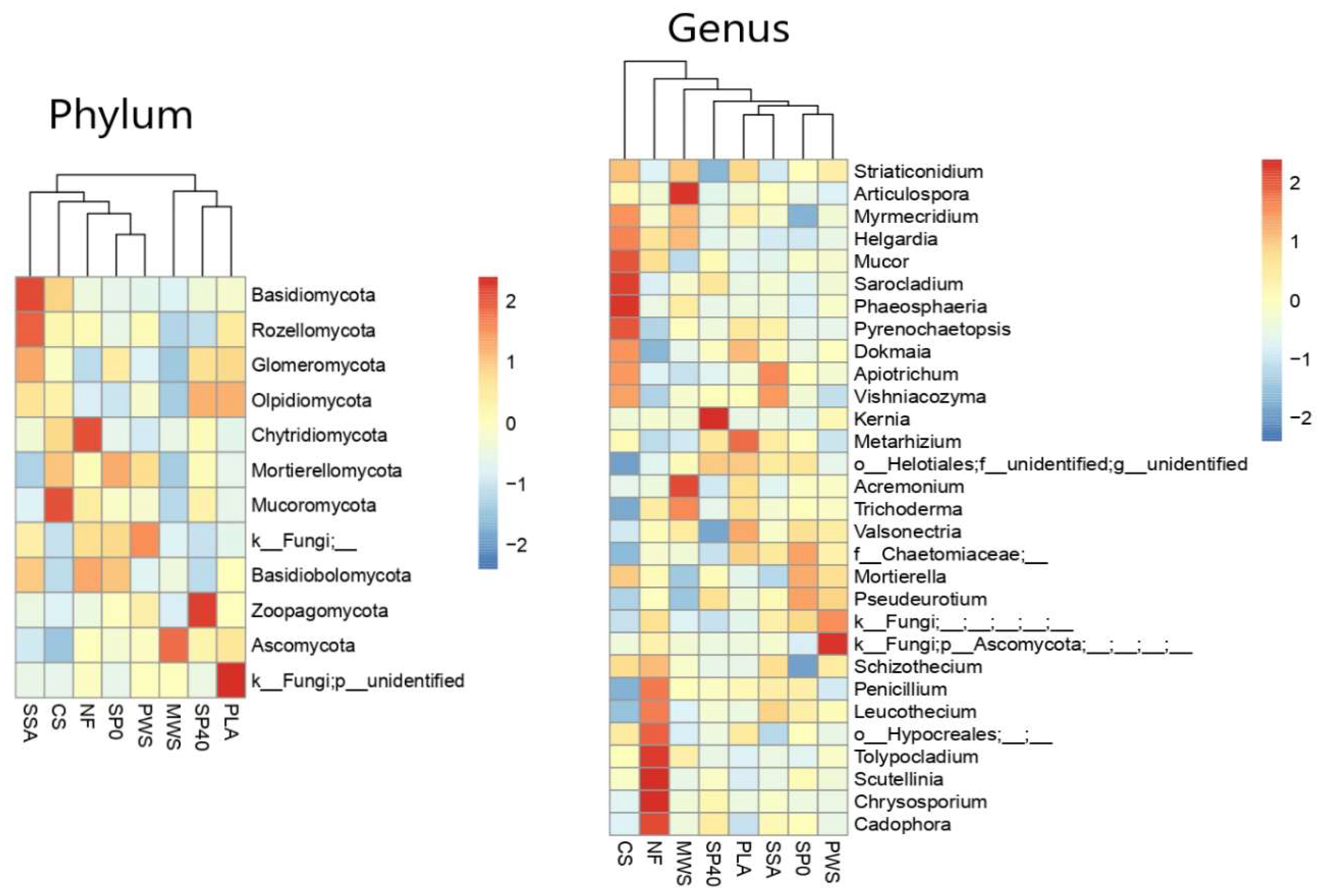
3.3.1. Community Composition
3.3.2. Nematode Alpha and Beta Diversity
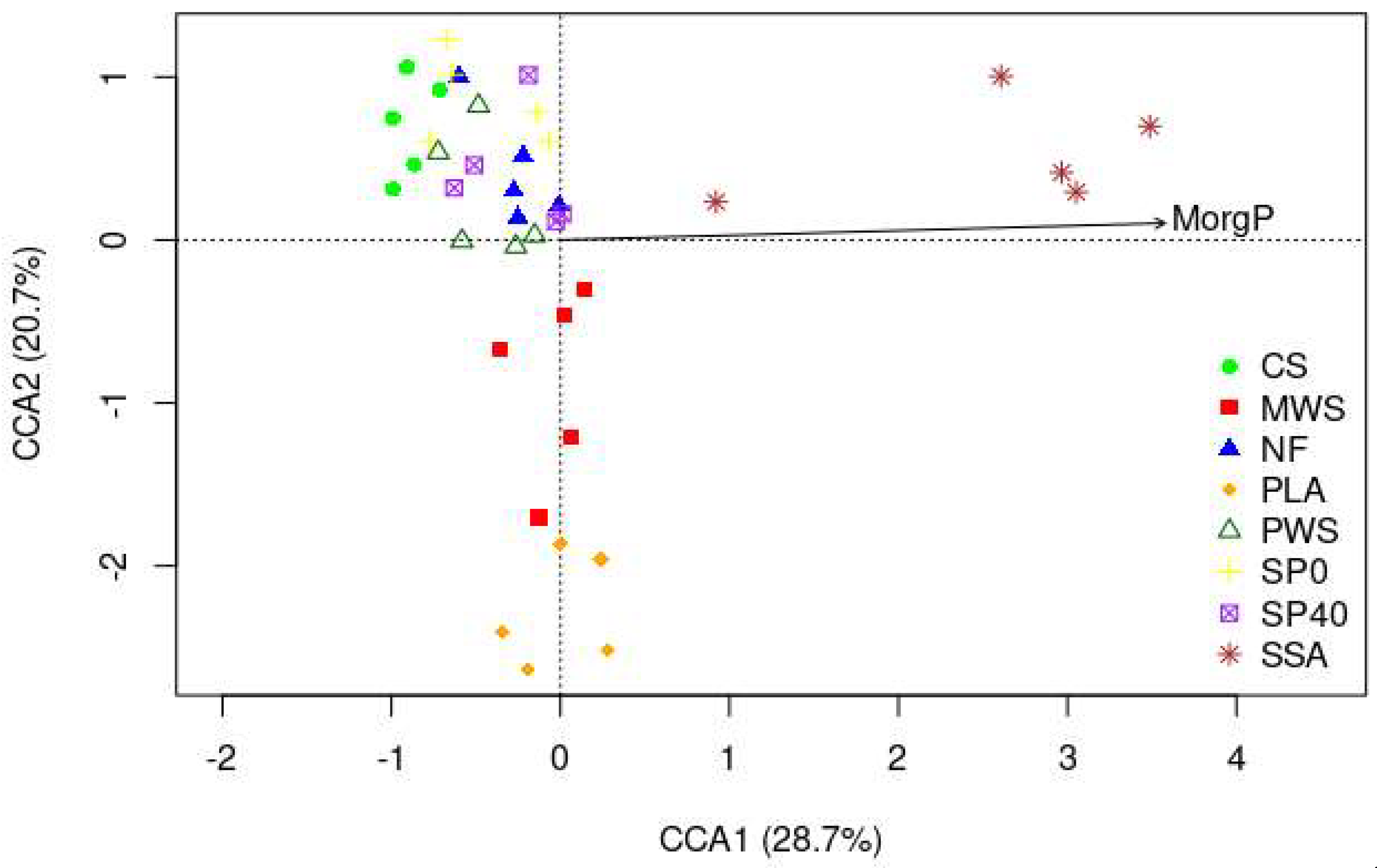
3.3.3. Relationships between Physiochemical Soil Variables and Nematode Communities
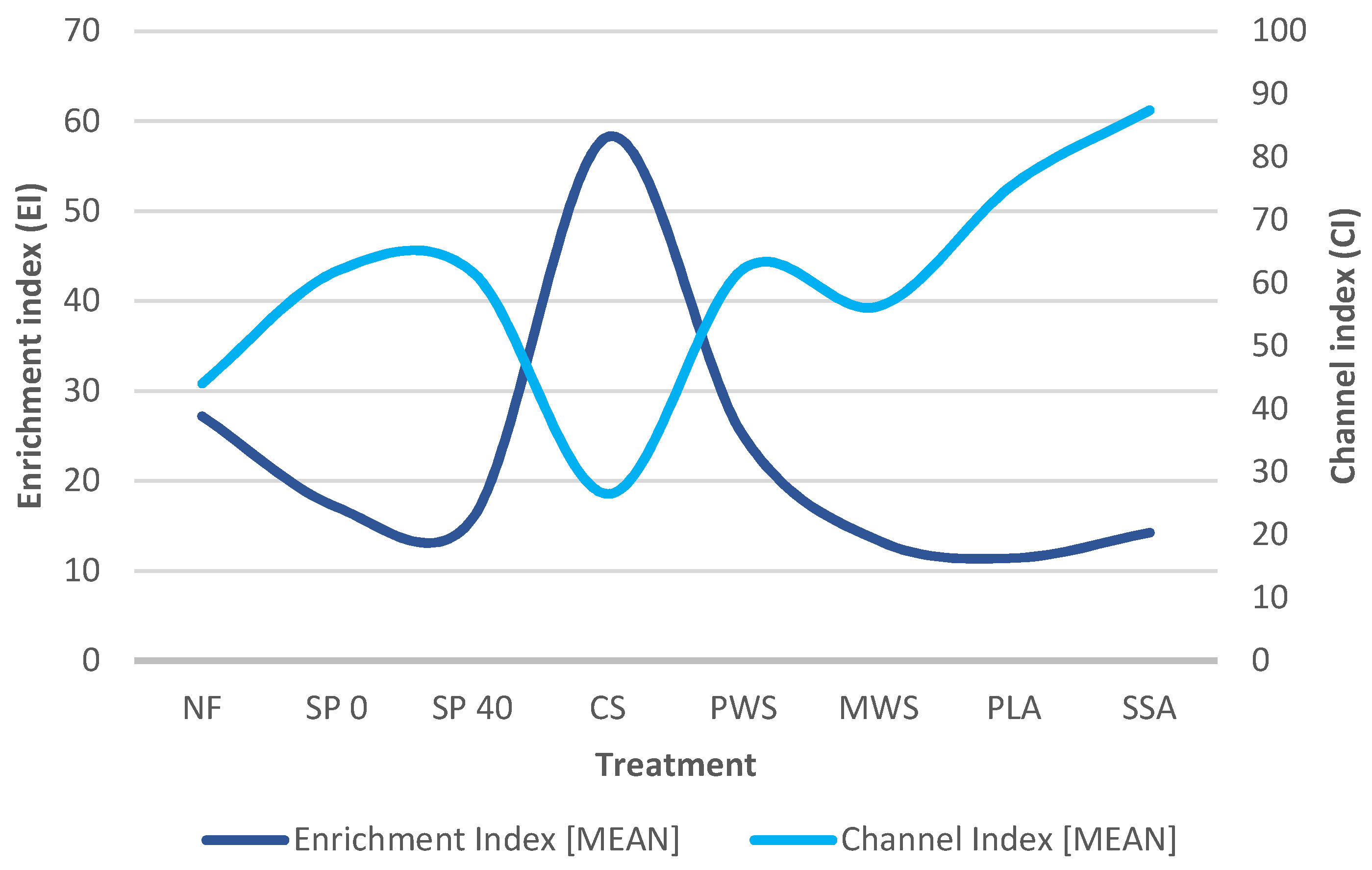
3.3.4. Food Web Diagnostics
4. Discussion
4.1. Microbial Community
4.2. Nematodes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cooper, J.; Lombardi, R.; Boardman, D.; Carliell-Marquet, C. The future distribution and production of global phosphate rock reserves. Resour. Conserv. Recycl. 2011, 57, 78–86. [Google Scholar] [CrossRef]
- Mehta, C.M.; Khunjar, W.O.; Nguyen, V.; Tait, S.; Batstone, D. Technologies to Recover Nutrients from Waste Streams: A Critical Review. Crit. Rev. Environ. Sci. Technol. 2014, 45, 385–427. [Google Scholar] [CrossRef] [Green Version]
- Maron, P.-A.; Mougel, C.; Ranjard, L. Soil microbial diversity: Methodological strategy, spatial overview and functional interest. Comptes Rendus Biol. 2011, 334, 403–411. [Google Scholar] [CrossRef]
- Torsvik, V.; Øvreås, L. Microbial diversity and function in soil: From genes to ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef]
- Verstraete, W.; Mertens, B. Chapter 5 The key role of soil microbes. In Developments in Soil Science; Doelman, P., Eijsackers, H.J.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; Volume 29, pp. 127–157. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Nielsen, M.N.; Winding, A. Microorganisms as Indicators of Soil Health; National Environmental Research Institute: Roskilde, Denmark, 2002. [Google Scholar]
- Singh, B.; Quince, C.; Macdonald, C.; Khachane, A.; Thomas, N.; Abu Al-Soud, W.; Sørensen, S.; He, Z.; White, D.; Sinclair, A.; et al. Loss of microbial diversity in soils is coincident with reductions in some specialized functions. Environ. Microbiol. 2014, 16, 2408–2420. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.-L.; Ding, J.; Zhu, Y.-G.; He, J.-Z.; Hu, H.-W. Soil bacterial taxonomic diversity is critical to maintaining the plant productivity. Environ. Int. 2020, 140, 105766. [Google Scholar] [CrossRef] [PubMed]
- Rao, D. Microbial Diversity, Soil Health and Sustainability. J. Indian. Soc. Soil Sci. 2007, 55, 392–403. [Google Scholar]
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Genet. 2017, 15, 579–590. [Google Scholar] [CrossRef]
- Zhen, Z.; Liu, H.; Wang, N.; Guo, L.; Meng, J.; Ding, N.; Wu, G.; Jiang, G. Effects of Manure Compost Application on Soil Microbial Community Diversity and Soil Microenvironments in a Temperate Cropland in China. PLoS ONE 2014, 9, e108555. [Google Scholar] [CrossRef]
- Wilson, M.; Kakouli-Duarte, T. Nematodes as Environmental Indicators; CABI: Wallingford, UK, 2009; pp. 1–17. [Google Scholar]
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Bongers, T.; Ferris, H. Nematode community structure as a bioindicator in environmental monitoring. Trends Ecol. Evol. 1999, 14, 224–228. [Google Scholar] [CrossRef]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef]
- Odum, E.P. Trends Expected in Stressed Ecosystems. BioScience 1985, 35, 419–422. [Google Scholar] [CrossRef]
- Wardle, D.A.; Yeates, G.W. The dual importance of competition and predation as regulatory forces in terrestrial ecosystems: Evidence from decomposer food-webs. Oecologia 1993, 93, 303–306. [Google Scholar] [CrossRef]
- Ferris, H.; Bongers, T.; Goede, R. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Bang-Andreasen, T.; Nielsen, J.T.; Voriskova, J.; Heise, J.; Rønn, R.; Kjøller, R.; Hansen, H.C.B.; Jacobsen, C.S. Wood Ash Induced pH Changes Strongly Affect Soil Bacterial Numbers and Community Composition. Front. Microbiol. 2017, 8, 1400. [Google Scholar] [CrossRef]
- Robles-Aguilar, A.A.; Grunert, O.; Hernandez-Sanabria, E.; Mysara, M.; Meers, E.; Boon, N.; Jablonowski, N.D. Effect of Applying Struvite and Organic N as Recovered Fertilizers on the Rhizosphere Dynamics and Cultivation of Lupine (Lupinus angustifolius). Front. Plant Sci. 2020, 11, 572741. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2010, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Armitage, D.W.; Gallagher, K.L.; Youngblut, N.D.; Buckley, D.H.; Zinder, S.H. Millimeter-scale patterns of phylogenetic and trait diversity in a salt marsh microbial mat. Front. Microbiol. 2012, 3, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Nuclear ribosomal in-ternal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhadury, P.; Austen, M.; Bilton, D.; Lambshead, P.; Rogers, A.; Smerdon, G. Development and evaluation of a DNA-barcoding approach for the rapid identification of nematodes. Mar. Ecol. Prog. Ser. 2006, 320, 1–9. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Nilsson, H.; Larsson, K.-H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: Handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2018, 47, D259–D264. [Google Scholar] [CrossRef]
- Sieriebriennikov, B.; Ferris, H.; Goede, R. NINJA: An automated calculation system for nematode-based biological monitoring. Eur. J. Soil Biol. 2014, 61, 90–93. [Google Scholar] [CrossRef]
- Zhang, B.; Wu, X.; Tai, X.; Sun, L.; Wu, M.; Zhang, W.; Chen, X.; Zhang, G.; Chen, T.; Liu, G.; et al. Variation in Actinobacterial Community Composition and Potential Function in Different Soil Ecosystems Belonging to the Arid Heihe River Basin of Northwest China. Front. Microbiol. 2019, 10, 2209. [Google Scholar] [CrossRef]
- Solans, M.; Messuti, M.I.; Reiner, G.; Boenel, M.; Vobis, G.; Wall, L.G.; Scervino, J.M. Exploring the response of Actinobacteria to the presence of phosphorus salts sources: Metabolic and co-metabolic processes. J. Basic Microbiol. 2019, 59, 487–495. [Google Scholar] [CrossRef]
- Zeng, Y.; Nupur, N.W.; Madsen, A.M.; Chen, X.; Gardiner, A.T.; Koblížek, M. Gemmatimonas groenlandica sp. nov. Is an Aerobic Anoxygenic Phototroph in the Phylum Gemmatimonadetes. Front. Microbiol. 2021, 11. [Google Scholar] [CrossRef]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global Biogeography and Quantitative Seasonal Dynamics of Gemmatimonadetes in Soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.; Jiang, N.; Wu, Y.; Yang, Z.; Bello, A.; Yang, W. Disentangling the role of salinity-sodicity in shaping soil microbiome along a natural saline-sodic gradient. Sci. Total Environ. 2021, 765, 142738. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.L.; Biggerstaff, J.; Eichhorn, A.; Ewing, E.; Shahan, R.; Soriano, D.; Stewart, S.; VanMol, K.; Walker, R.; Walters, P.; et al. Genomic characterization of three novel Desulfobacterota classes expand the metabolic and phylogenetic diversity of the phylum. Environ. Microbiol. 2021, 23, 4326–4343. [Google Scholar] [CrossRef] [PubMed]
- Wolińska, A.; Kuźniar, A.; Zielenkiewicz, U.; Izak, D.; Szafranek-Nakonieczna, A.; Banach, A.; Błaszczyk, M. Bacteroidetes as a sensitive biological indicator of agricultural soil usage revealed by a culture-independent approach. Appl. Soil Ecol. 2017, 119, 128–137. [Google Scholar] [CrossRef]
- Oren, A. The Family Xanthobacteraceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Rosenberg, E., De Long, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 709–726. [Google Scholar]
- Oliverio, A.M.; Bissett, A.; McGuire, K.; Saltonstall, K.; Turner, B.L.; Fierer, N.; Moran, M.A. The Role of Phosphorus Limitation in Shaping Soil Bacterial Communities and Their Metabolic Capabilities. Mbio 2020, 11, e01718-20. [Google Scholar] [CrossRef]
- Anderson, C.R.; Condron, L.M.; Clough, T.; Fiers, M.; Stewart, A.; Hill, R.; Sherlock, R.R. Biochar induced soil microbial community change: Implications for biogeochemical cycling of carbon, nitrogen and phosphorus. Pedobiologia 2011, 54, 309–320. [Google Scholar] [CrossRef]
- Anand, K.; Kumari, B.; Mallick, M. Phosphate solubilizing microbes: An effective and alternative approach as biofertilizers. Int. J. Pharm. Pharm. Sci. 2016, 8, 37–40. [Google Scholar]
- Zhang, M.-M.; Wang, N.; Hu, Y.-B.; Sun, G.-Y. Changes in soil physicochemical properties and soil bacterial community in mulberry (Morus alba L.)/alfalfa (Medicago sativa L.) intercropping system. Microbiol. Open 2018, 7, e00555. [Google Scholar] [CrossRef]
- Navarrete, A.; Soares, T.; Rossetto, R.; Van Veen, J.A.; Tsai, S.M.; Kuramae, E.E. Verrucomicrobial community structure and abundance as indicators for changes in chemical factors linked to soil fertility. Antonie Leeuwenhoek 2015, 108, 741–752. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Ashworth, A.J.; DeBruyn, J.; Willett, C.; Durso, L.M.; Cook, K.; Moore, J.P.A.; Owens, P.R. Soil bacterial biodiversity is driven by long-term pasture management, poultry litter, and cattle manure inputs. PeerJ 2019, 7, e7839. [Google Scholar] [CrossRef] [Green Version]
- Francioli, D.; Schulz, E.; Lentendu, G.; Wubet, T.; Buscot, F.; Reitz, T. Mineral vs. Organic Amendments: Microbial Community Structure, Activity and Abundance of Agriculturally Relevant Microbes Are Driven by Long-Term Fertilization Strategies. Front. Microbiol. 2016, 7, 1446. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Li, C.; Xiao, X.; Shi, L.; Cheng, K.; Wen, L.; Li, W. Effects of short-term manure nitrogen input on soil microbial community structure and diversity in a double-cropping paddy field of southern China. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Pathan, S.; Scibetta, S.; Grassi, C.; Pietramellara, G.; Orlandini, S.; Ceccherini, M.; Napoli, M. Response of Soil Bacterial Community to Application of Organic and Inorganic Phosphate Based Fertilizers under Vicia faba L. Cultivation at Two Different Phenological Stages. Sustainability 2020, 12, 9706. [Google Scholar] [CrossRef]
- Celestina, C.; Wood, J.L.; Manson, J.B.; Wang, X.; Sale, P.W.G.; Tang, C.; Franks, A.E. Microbial communities in top- and subsoil of repacked soil columns respond differently to amendments but their diversity is negatively correlated with plant productivity. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Li, H.; Xu, Z.; Yang, S.; Li, X.; Top, E.M.; Wang, R.; Zhang, Y.; Cai, J.; Yao, F.; Han, X.; et al. Responses of Soil Bacterial Communities to Nitrogen Deposition and Precipitation Increment Are Closely Linked with Aboveground Community Variation. Microb. Ecol. 2016, 71, 974–989. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and Qualitative β Diversity Measures Lead to Different Insights into Factors That Structure Microbial Communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [Green Version]
- Randall, K.; Brennan, F.; Clipson, N.; Creamer, R.; Griffiths, B.; Storey, S.; Doyle, E. Soil bacterial community structure and functional responses across a long-term mineral phosphorus (Pi) fertilisation gradient differ in grazed and cut grasslands. Appl. Soil Ecol. 2019, 138, 134–143. [Google Scholar] [CrossRef]
- Lazcano, C.; Gómez-Brandón, M.; Revilla, P.; Domínguez, J. Short-term effects of organic and inorganic fertilizers on soil microbial community structure and function. Biol. Fertil. Soils 2012, 49, 723–733. [Google Scholar] [CrossRef]
- Tayyab, M.; Islam, W.; Lee, C.G.; Pang, Z.; Khalil, F.; Lin, S.; Lin, W.; Zhang, H. Short-Term Effects of Different Organic Amendments on Soil Fungal Composition. Sustainability 2019, 11, 198. [Google Scholar] [CrossRef] [Green Version]
- Samuel, A.D.; Bungau, S.; Tit, D.M.; Melinte, C.E.; Purza, L.; Badea, G.E. Effects of Long Term Application of Organic and Mineral Fertilizers on Soil Enzymes. Rev. Chim. 2018, 69, 2608–2612. [Google Scholar] [CrossRef]
- Pahalvi, H.N.; Rafiya, L.; Rashid, S.; Nisar, B.; Kamili, A.N. Chemical Fertilizers and Their Impact on Soil Health. In Microbiota and Biofertilizers; Springer: Cham, Switzerland, 2021; pp. 1–20. [Google Scholar] [CrossRef]
- Bungau, S.; Behl, T.; Aleya, L.; Bourgeade, P.; Aloui-Sossé, B.; Purza, A.L.; Abid, A.; Samuel, A.D. Expatiating the impact of anthropogenic aspects and climatic factors on long-term soil monitoring and management. Environ. Sci. Pollut. Res. 2021, 28, 1–23. [Google Scholar] [CrossRef]
- Samuel, A.; Brejea, R.; Domuta, C.; Bungau, S.; Cenusa, N.; Tit, D.M. Enzymatic Indicators of Soil Quality. J. Environ. Prot. Ecol. 2017, 18, 871–878. [Google Scholar]
- Ockenden, M.C.; Hollaway, M.J.; Beven, K.J.; Collins, A.L.; Evans, R.; Falloon, P.D.; Forber, K.J.; Hiscock, K.M.; Kahana, R.; Macleod, C.J.A.; et al. Major agricultural changes re-quired to mitigate phosphorus losses under climate change. Nat. Commun. 2017, 8, 161. [Google Scholar] [CrossRef] [Green Version]
- Korthals, G.W.; Bongers, M.; Kokkema, A.; Dueck, T.A.; Lexmond, T.M. Joint Toxicity of Copper and Zinc to a Terrestrial Nematode Community in an Acid Sandy Soil. Ecotoxicology 2000, 9, 219–228. [Google Scholar] [CrossRef]
- Ferris, H.; Matute, M. Structural and functional succession in the nematode fauna of a soil food web. Appl. Soil Ecol. 2003, 23, 93–110. [Google Scholar] [CrossRef]
- Yeates, G.W.; Wardle, D.A. Nematodes as predators and prey: Relationships to biological control and soil processes. Pedobiologia 1996, 40, 43–50. [Google Scholar]
- Steen, I. Phosphorus availability in the 21st Century: Management of a non-renewable resource. Phosphorus Potassium 1998, 217, 25–31. [Google Scholar]
- Smil, V. Phosphorus in the environment: Natural Flows and Human Interferences. Annu. Rev. Energy Environ. 2000, 25, 53–88. [Google Scholar] [CrossRef] [Green Version]
- Ashley, K.; Mavinic, D.; Koch, F. International Conference on Nutrient Recovery from Wastewater Streams; IWA Publishing: Vancouver, BC, Canada, 2009; pp. 20–40. [Google Scholar] [CrossRef]
- Sarkar, S.; Singh, S.R.; Singh, R.P. The effect of organic and inorganic fertilizers on soil physical condition and the productivity of a rice–lentil cropping sequence in India. J. Agric. Sci. 2003, 140, 419–425. [Google Scholar] [CrossRef]
- Hilton, J.; Johnston, A.E.; Dawson, C.J. The Phosphate Life-Cycle: Rethinking the Options for a Finite Resource; International Fertiliser Society: London, UK, 2010; pp. 1–13. [Google Scholar]
- DeSmidt, E.; Ghyselbrecht, K.; Zhang, Y.; Pinoy, L.; Van Der Bruggen, B.; Verstraete, W.; Rabaey, K.; Meesschaert, B. Global Phosphorus Scarcity and Full-Scale P-Recovery Techniques: A Review. Crit. Rev. Environ. Sci. Technol. 2014, 45, 336–384. [Google Scholar] [CrossRef]
- Briar, S.S.; Miller, S.A.; Stinner, D.; Kleinhenz, M.; Grewal, P.S. Effects of organic transition strategies for peri-urban vegetable production on soil properties, nematode community, and tomato yield. Appl. Soil Ecol. 2011, 47, 84–91. [Google Scholar] [CrossRef]
- Ugarte, C.M.; Zaborski, E.R.; Wander, M.M. Nematode indicators as integrative measures of soil condition in organic cropping systems. Soil Biol. Biochem. 2013, 64, 103–113. [Google Scholar] [CrossRef]
- Leroy, B.L.; Reheul, D.; Moens, M.; Ferris, H.; De Sutter, N. Short-term nematode population dynamics as influenced by the quality of exogenous organic matter. Nematology 2009, 11, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Talgre, L.; Lauringson, E.; Roostalu, H.; Astover, A.; Makke, A. Green manure as a nutrient source for succeeding crops. Plant Soil Environ. 2012, 58, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Ya, T.; Js, Z.; Pc, G.; Jn, C. Effects of various organic materials on soil aggregate stability and soil microbiological properties on the Loess Plateau of China. Plant Soil Environ. 2013, 59, 162–168. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Ding, W.; He, X.; Yu, H.; Fan, J.; Liu, D. Influence of 20–Year Organic and Inorganic Fertilization on Organic Carbon Accumulation and Microbial Community Structure of Aggregates in an Intensively Cultivated Sandy Loam Soil. PLoS ONE 2014, 9, e92733. [Google Scholar] [CrossRef]
- Gruzdeva, L.I.; Matveeva, E.M.; Kovalenko, T.E. Changes in soil nematode communities under the impact of fertilizers. Eurasian Soil Sci. 2007, 40, 681–693. [Google Scholar] [CrossRef]
- Withers, P.J.A.; Neal, C.; Jarvie, H.P.; Doody, D.G. Agriculture and Eutrophication: Where Do We Go from Here? Sustainability 2014, 6, 5853–5875. [Google Scholar] [CrossRef] [Green Version]
- Hoogen, J.V.D.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.; De Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.; et al. Soil nematode abundance and functional group composition at a global scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Bardgett, R. The Biology of Soil, a Community and Ecosystem Approach; Oxford University Press: Oxford, UK, 2005; p. 256. [Google Scholar]
- Yan, D.; Yan, D.; Song, X.; Yu, Z.; Peng, D.; Ting, X.; Weng, B. Community structure of soil nematodes under different drought conditions. Geoderma 2018, 325, 110–116. [Google Scholar] [CrossRef]
- Simmons, B.; Wall, D.; Adams, B.; Ayres, E.; Barrett, J.; Virginia, R. Long-term experimental warming reduces soil nematode populations in the McMurdo Dry Valleys, Antarctica. Soil Biol. Biochem. 2009, 41, 2052–2060. [Google Scholar] [CrossRef]
- Li, Q.; Bai, H.; Liang, W.; Xia, J.; Wan, S.; Van Der Putten, W.H. Nitrogen Addition and Warming Independently Influence the Belowground Micro-Food Web in a Temperate Steppe. PLoS ONE 2013, 8, e60441. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Hou, R.; Chen, Q.; Ouyang, Z.; Ge, F. Response of soil nematodes to elevated temperature in conventional and no-tillage cropland systems. Plant Soil 2013, 373, 907–918. [Google Scholar] [CrossRef]
- Ruess, L.; Michelsen, A.; Schmidt, I.K.; Jonasson, S. Simulated climate change affecting microorganisms, nematode density and biodiversity in subarctic soils. Plant Soil 1999, 212, 63–73. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Available online: https://www.ipcc.ch/site/assets/uploads/2017/09/WG1AR5_Frontmatter_FINAL.pdf (accessed on 23 October 2021).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recycling-Derived Fertiliser (RDF) | pH | N | P | K | S | Ca | Al | Fe |
|---|---|---|---|---|---|---|---|---|
| % | ||||||||
| Potato waste struvite (PWS) | 8.3 | 3.0 | 6.2 | 0.7 | 0 | 0.03 | 0.002 | 0.004 |
| Municipal waste struvite (MWS) | 7.5 | 2.6 | 5.2 | 0.03 | 0 | 0.02 | 0.002 | 0.01 |
| Poultry litter ash (PLA) | 12.3 | 0.02 | 5.5 | 10.7 | 3.1 | 15.6 | 0.75 | 0.46 |
| Sewage sludge ash (SSA) | 11.4 | 0.03 | 8.4 | 1.3 | 3.0 | 10.3 | 5.3 | 6.0 |
| Treatment | Tot C (%) | Org C (%) | OM (% of DM) | pH | N (% of DM) |
|---|---|---|---|---|---|
| NF | 2.23 ± 0.27 | 1.65 ± 0.15 | 5.54 ± 0.66 | 5.91 ± 0.16 | 0.24 ± 0.03 |
| SP0 | 2.53 ± 0.06 | 1.86 ± 0.11 | 5.98 ± 0.05 | 5.89 ± 0.09 | 0.26 ± 0.00 |
| SP40 | 2.14 ± 0.07 | 1.58 ± 0.03 | 5.46 ± 0.05 | 5.95 ± 0.00 | 0.23 ± 0.00 |
| CS | 2.40 ± 0.07 | 1.81 ± 0.10 | 5.84 ± 0.00 | 6.01 ± 0.02 | 0.25 ± 0.00 |
| PWS | 2.56 ± 0.23 | 1.93 ± 0.16 | 6.26 ± 0.50 | 5.83 ± 0.24 | 0.27 ± 0.02 |
| MWS | 2.24 ± 0.46 | 1.65 ± 0.21 | 5.72 ± 0.70 | 5.94 ± 0.04 | 0.24 ± 0.03 |
| PLA | 2.46 ± 0.15 | 1.86 ± 0.04 | 6.02 ± 0.50 | 5.95 ± 0.27 | 0.26 ± 0.01 |
| SSA | 2.28 ± 0.21 | 1.69 ± 0.14 | 5.44 ± 0.15 | 6.05 ± 0.16 | 0.24 ± 0.02 |
| Treatment | Morg P (mg/L) | Morg K (mg/L) | Morg Mg (mg/L) | DM Yield (ton/ha) | Uptake P (kg/ha) |
| NF | 2.38 ± 0.51 a | 61.14 ± 18.14 a | 158.60 ± 10.90 | 1.39 ± 0.36 | 2.86 ± 0.71 a |
| SP0 | 2.47 ± 1.07 a | 95.52 ± 1.30 ab | 141.80 ± 7.50 | 3.15 ± 0.38 a | 6.28 ± 1.03 ab |
| SP40 | 2.95 ± 0.31 ab | 70.04 ± 7.30 ab | 144.80 ± 20.50 | 3.22 ± 0.44 a | 7.28 ± 0.87 b |
| CS | 3.50 ± 0.06 ab | 162.60 ± 9.45 | 159.00 ± 19.00 | 3.66 ± 0.23 a | 8.00 ± 0.84 b |
| PWS | 3.93 ± 0.68 ab | 93.32 ± 55.21 ab | 163.40 ± 20.80 | 3.47 ± 0.34 a | 8.04 ± 1.40 b |
| MWS | 4.51 ± 0.58 b | 88.92 ± 5.20 ab | 161.60 ± 9.50 | 3.64 ± 0.70 a | 8.20 ± 1.45 b |
| PLA | 3.86 ± 0.05 ab | 102.48 ± 33.15 b | 148.00 ± 2.50 | 2.83 ± 0.59 a | 7.32 ± 3.91 b |
| SSA | 4.18 ± 1.16 ab | 85.88 ± 11.45 ab | 158.00 ± 8.10 | 3.31 ± 0.21 a | 6.83 ± 0.77 b |
| Bacteria | Fungi | |||||
|---|---|---|---|---|---|---|
| Treatment | Observed OTUs | Simpson | Chao1 | Observed OTUs | Simpson | Chao1 |
| NF | 1887 ± 37 bd | 0.9923 ± 0.001 abc | 2925 ± 176 | 411 ± 21 | 0.888 ± 0.026 | 538 ± 44 |
| SP0 | 1865 ± 36 bcd | 0.9922 ± 0.001 ab | 2853 ± 78 | 435 ± 37 | 0.881 ± 0.027 | 621 ± 65 |
| SP40 | 1810 ± 53 ac | 0.9923 ± 0.001 abc | 2690 ± 154 | 455 ± 47 | 0.903 ± 0.038 | 615 ± 60 |
| CS | 1787 ± 23 a | 0.992 ± 0.000 a | 2674 ± 149 | 426 ± 40 | 0.916 ± 0.027 | 560 ± 49 |
| PWS | 1828 ± 40 abc | 0.9924 ± 0.000 abc | 2755 ± 76 | 445 ± 40 | 0.890 ± 0.035 | 658 ± 60 |
| MWS | 1847 ± 35 bcd | 0.9929 ± 0.001 bcd | 2711 ± 132 | 427 ± 18 | 0.914 ± 0.026 | 581 ± 47 |
| PLA | 1875 ± 31 bcd | 0.9928 ± 0.000 cd | 2838 ± 105 | 443 ± 41 | 0.915 ± 0.015 | 600 ± 72 |
| SSA | 1895 ± 36 d | 0.9933 ± 0.000 d | 2815 ± 99 | 464 ± 30 | 0.912 ± 0.010 | 625 ± 53 |
| Treatment | Observed OTUs ± S.D. | Shannon ± S.D. |
|---|---|---|
| NF | 175 ± 4.40 a | 4.56 ± 0.11 a |
| PLA | 174 ± 6.20 a | 5.1 ± 0.15 a |
| PWS | 173 ± 3.80 a | 4.55 ± 0.26 a |
| SP 40 | 172 ± 9.30 a | 4.47 ± 0.33 a |
| CS | 172 ± 5.13 a | 4.62 ± 0.47 a |
| SP 0 | 171 ± 6.40 ab | 4.72 ± 0.24 a |
| MWS | 167 ± 7.50 a | 4.48 ± 0.6 |
| SSA | 153 ± 15.9 b | 4.3 ± 0.73 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karpinska, A.; Ryan, D.; Germaine, K.; Dowling, D.; Forrestal, P.; Kakouli-Duarte, T. Soil Microbial and Nematode Community Response to the Field Application of Recycled Bio-Based Fertilisers in Irish Grassland. Sustainability 2021, 13, 12342. https://doi.org/10.3390/su132212342
Karpinska A, Ryan D, Germaine K, Dowling D, Forrestal P, Kakouli-Duarte T. Soil Microbial and Nematode Community Response to the Field Application of Recycled Bio-Based Fertilisers in Irish Grassland. Sustainability. 2021; 13(22):12342. https://doi.org/10.3390/su132212342
Chicago/Turabian StyleKarpinska, Anna, Demi Ryan, Kieran Germaine, David Dowling, Patrick Forrestal, and Thomais Kakouli-Duarte. 2021. "Soil Microbial and Nematode Community Response to the Field Application of Recycled Bio-Based Fertilisers in Irish Grassland" Sustainability 13, no. 22: 12342. https://doi.org/10.3390/su132212342