
Nitrogen Fertilizer Driven Bacterial Community Structure in a Semi-Arid Region of Northeast China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Description and Soil Sample Collection
2.2. Soil Chemical Properties Measurements
2.3. Soil DNA Extraction and Quantitative Real-Time PCR (Q-PCR)
2.4. Illumina MiSeq Sequencing
2.5. Processing of Sequencing Data
2.6. Statistical Analysis
3. Results
3.1. Soil Chemical Properties
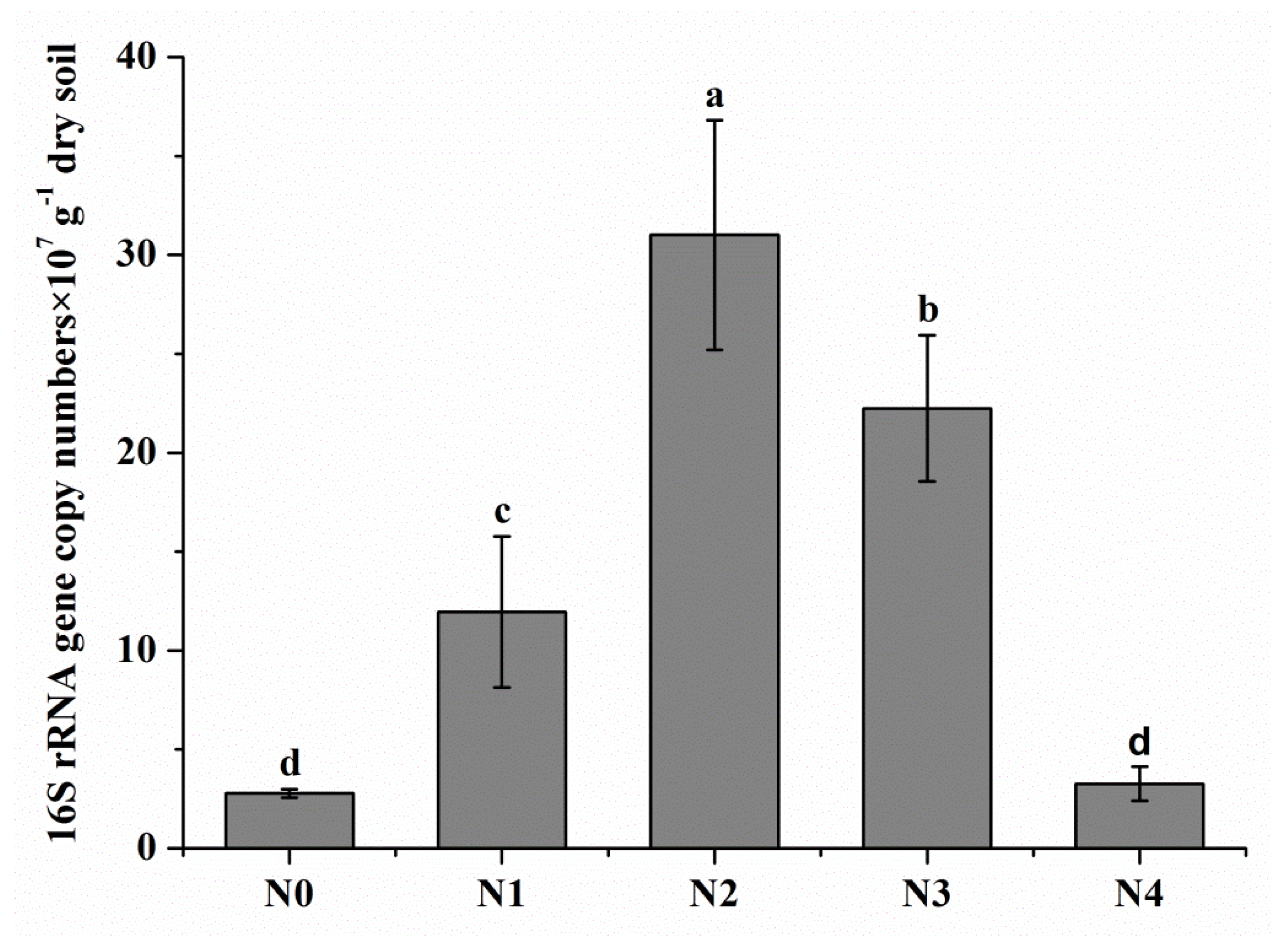
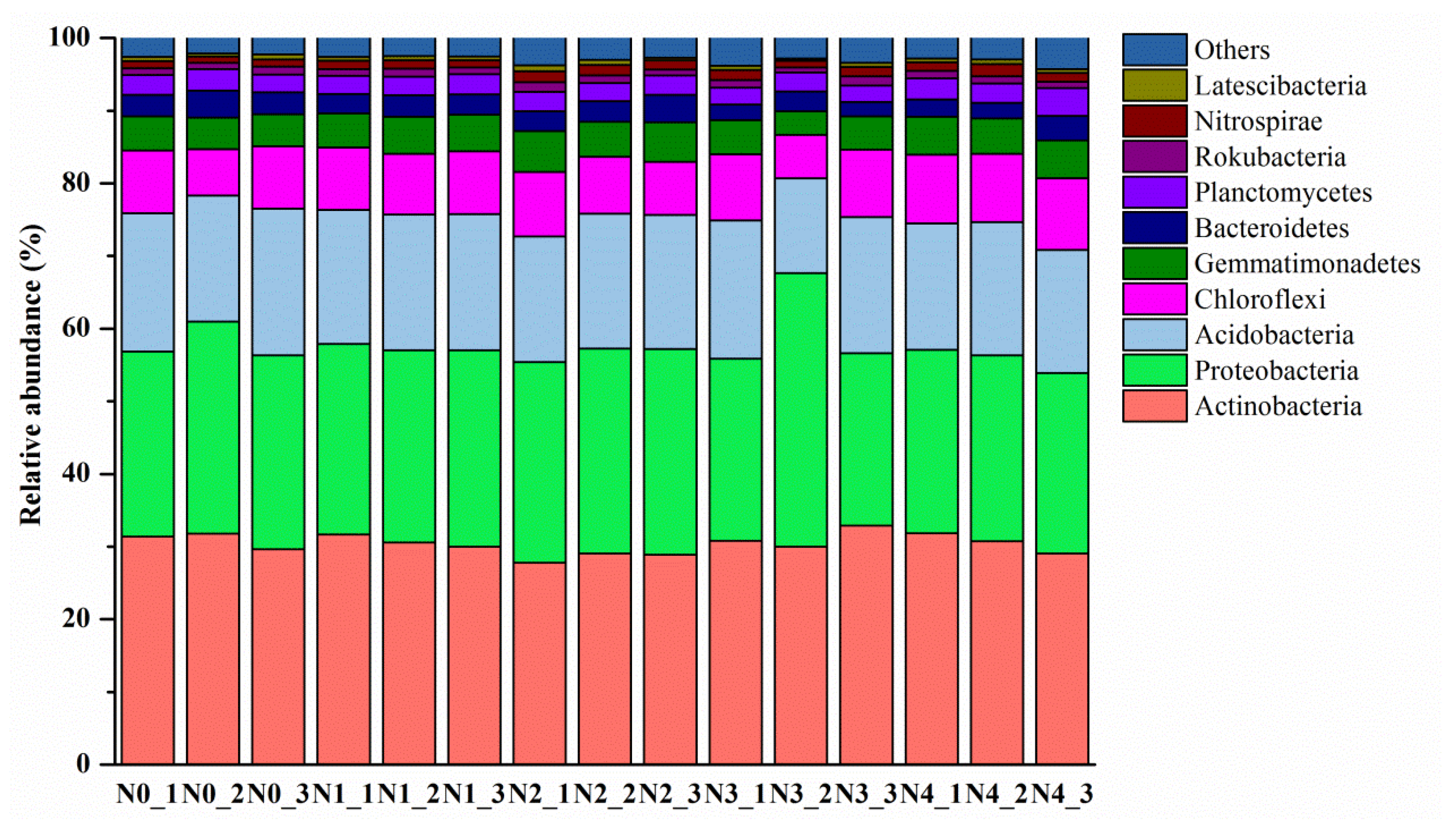
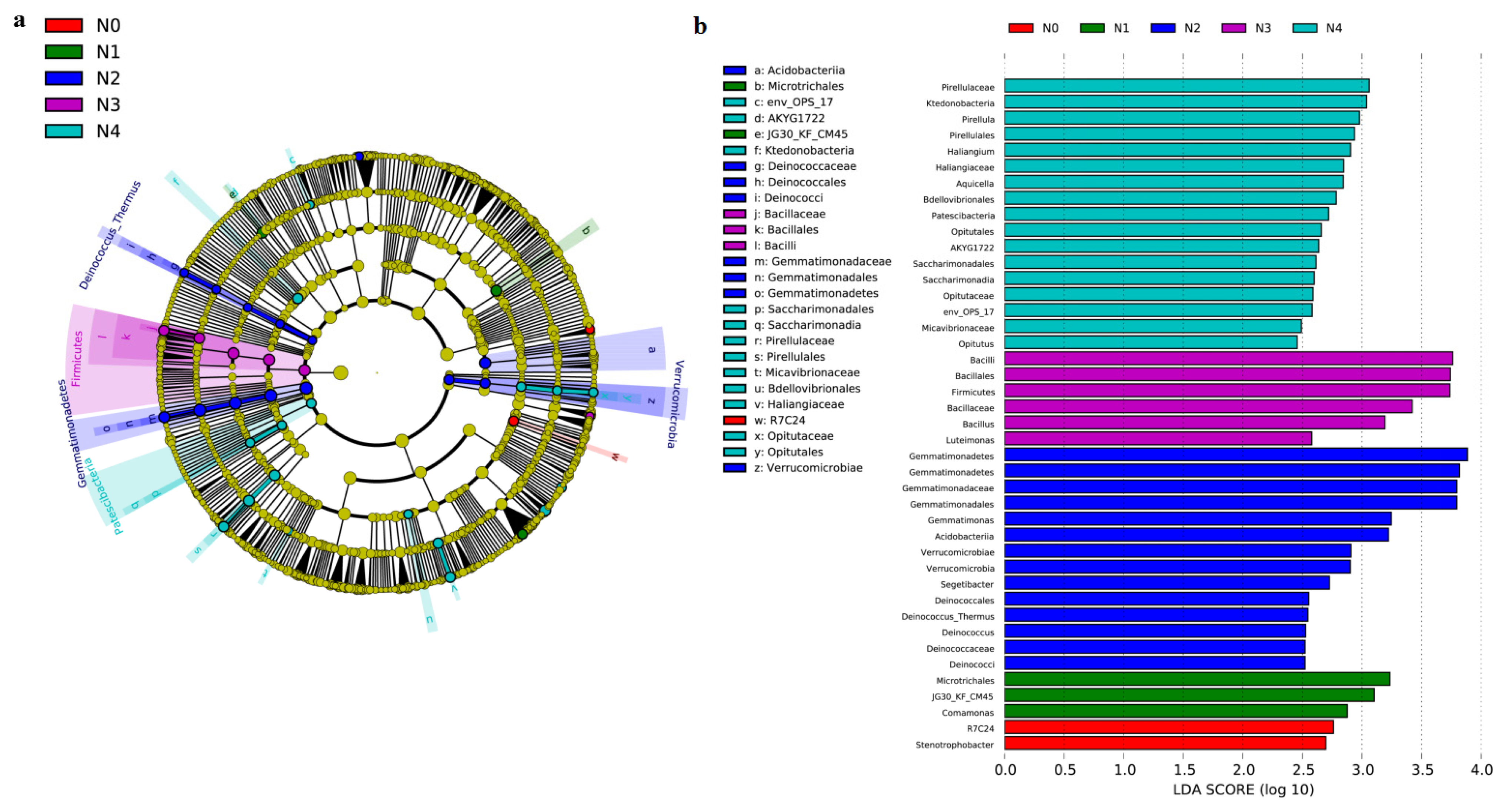
3.2. Soil Bacterial Abundance and Bacterial Community Composition
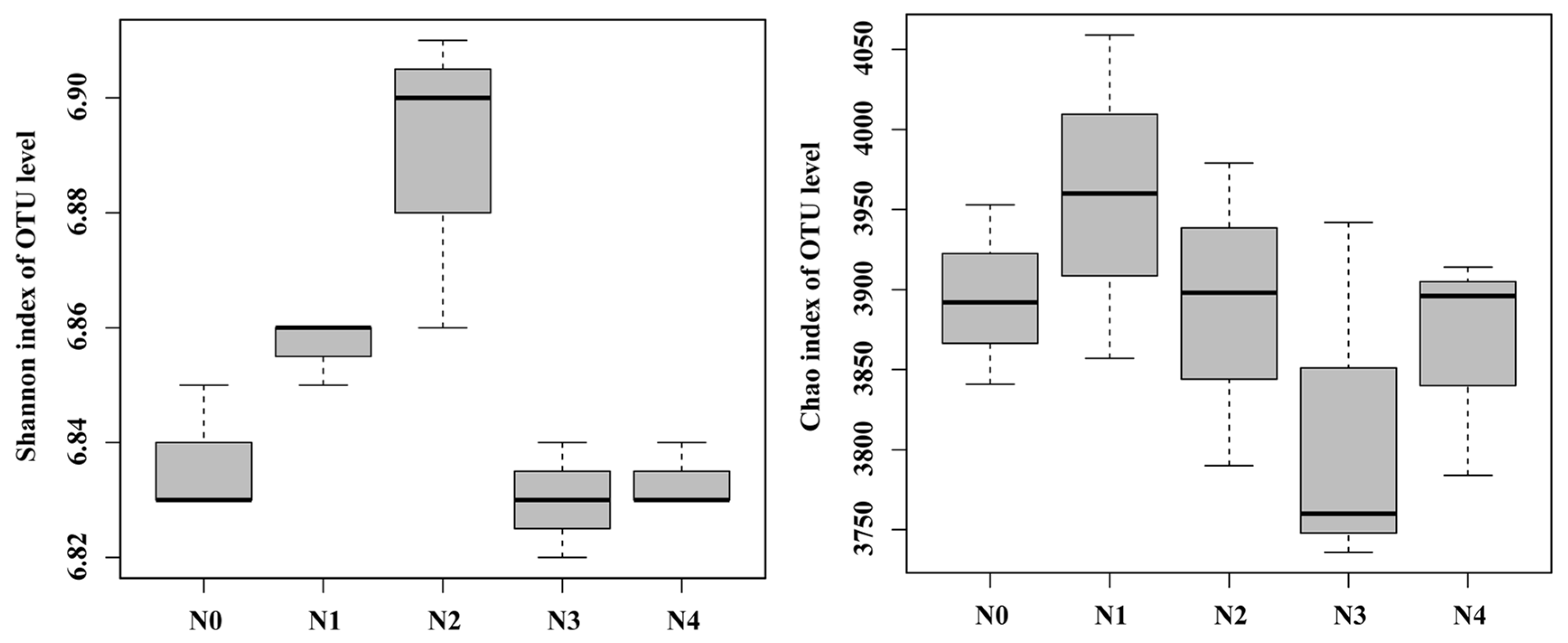
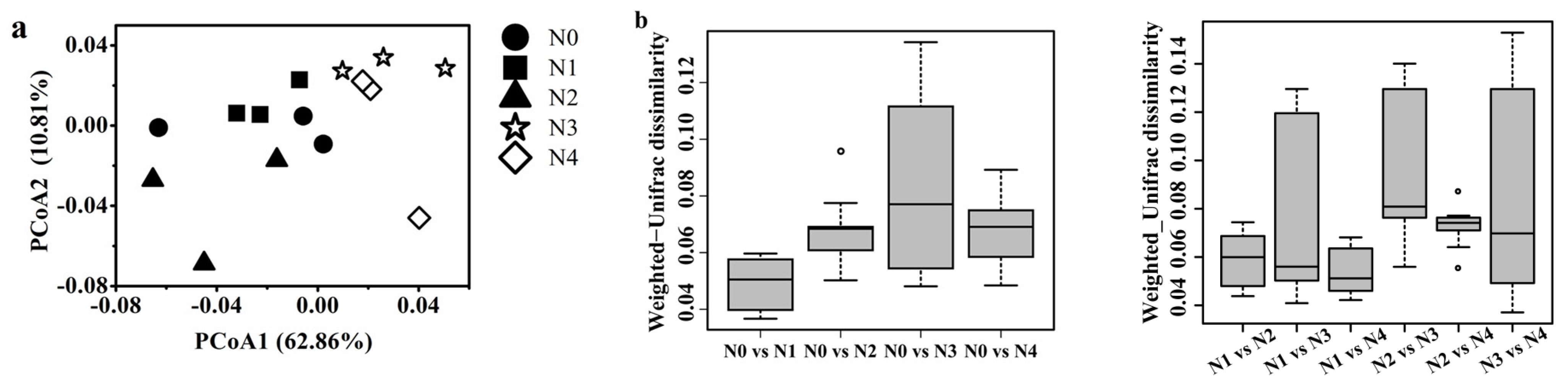
3.3. Soil Bacterial Community Diversity
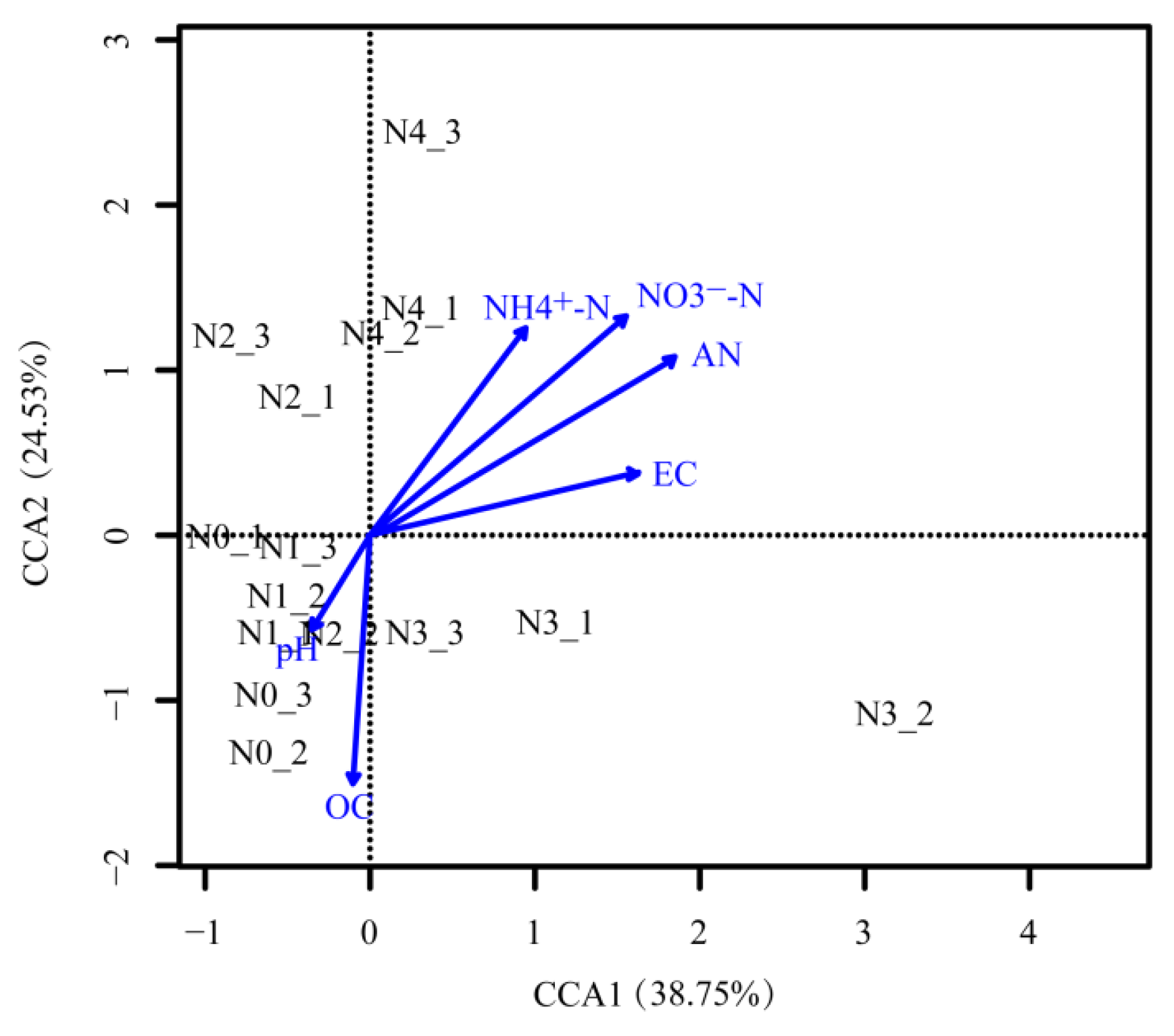
3.4. Relationships between Soil Bacterial Communities and Chemical Properties
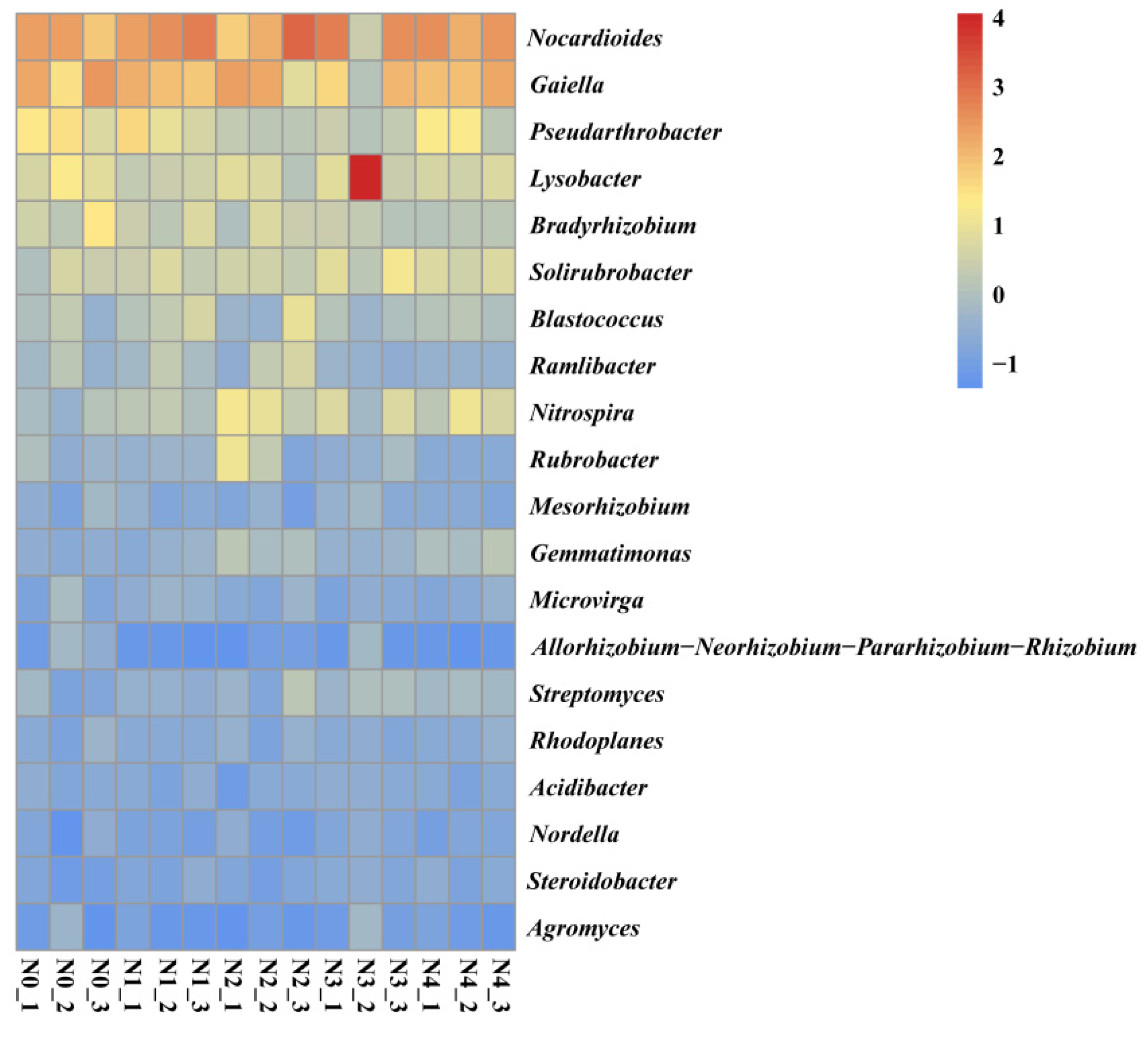
3.5. Soil Bacteria Functional Groups
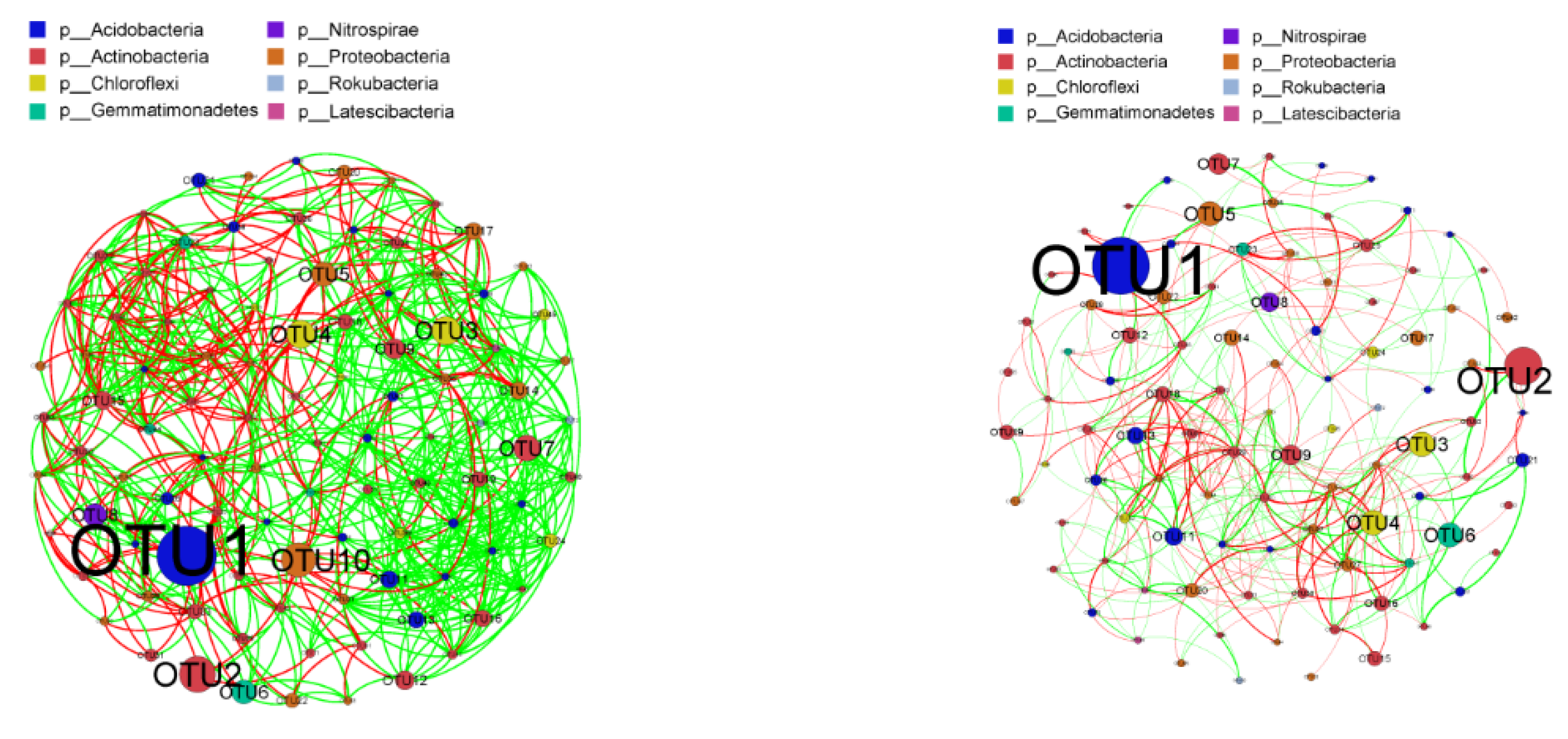
3.6. Bacterial Co-Occurrence Networks
4. Discussion
4.1. Variation in Soil Physicochemical Properties under Different N Gradients
4.2. Shifts in Bacterial Abundance and Community Composition in Response to Different N Gradients
4.3. Effects of Different N Gradients on Bacterial Diversity
4.4. The Main Factors Affecting Bacterial Community Structure Variation
4.5. Effect of N Fertilizer on Soil Functional Bacteria
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Gruber, N.; Galloway, J.N. An Earth-system perspective of the global nitrogen cycle. Nature 2008, 451, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xu, R.; Su, F.; Roelcke, M.; Ju, X. Effects of enhanced efficiency nitrogen fertilizers on NH3 losses in a calcareous fluvo-aquic soil: A laboratory study. J. Soils Sediments 2020, 20, 1887–1896. [Google Scholar] [CrossRef]
- Cassman, K.G.; Dobermann, A.; Walters, D.T. Agroecosystems, nitrogen-use efficiency, and nitrogen management. Ambio 2002, 31, 132–140. [Google Scholar] [CrossRef]
- Cui, Z.; Zhang, F.; Chen, X.; Dou, Z.; Li, J. In-season nitrogen management strategy for winter wheat: Maximizing yields, minimizing environmental impact in an over-fertilization context. Field Crop. Res. 2010, 116, 140–146. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Naylor, R.; Crews, T.; David, M.B.; Drinkwater, L.E.; Holland, E.; Johnes, P.J.; Katzenberger, J.; Martinelli, L.A.; Matson, P.A.; et al. Nutrient imbalances in agricultural development. Science 2009, 324, 1519–1520. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Xing, G.; Chen, X.; Zhang, S.; Zhang, L.; Liu, X.; Cui, Z.; Yin, B.; Christie, P.; Zhu, Z.; et al. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2011, 418, 671–677. [Google Scholar] [CrossRef] [PubMed]
- FAO. FAOSTAT Database—Agriculture Production. 2012. Available online: http://www.fao.org/home/en/ (accessed on 23 August 2019).
- Guo, J.H.; Liu, X.J.; Zhang, Y.; Shen, J.L.; Han, W.X.; Zhang, W.F.; Christie, P.; Goulding, K.W.T.; Vitousek, P.M.; Zhang, F.S. Significant acidification in major chinese croplands. Science 2010, 327, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, C.; Zha, Y.; Li, Y.; Sun, D.; Lu, H.; Yin, B. Eutrophication of lake waters in China: Cost, causes, and control. Environ. Manag. 2010, 45, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Wang, G.; Yue, S.; Wu, L.; Zhang, W.; Zhang, F.; Chen, X. Closing the N-use efficiency gap to achieve food and environmental security. Environ. Sci. Technol. 2014, 48, 5780–5787. [Google Scholar] [CrossRef] [PubMed]
- Abujabhah, I.S.; Doyle, R.B.; Bound, S.A.; Bowman, J.P. Assessment of bacterial community composition, methanotrophic and nitrogen-cycling bacteria in three soils with different biochar application rates. J. Soils Sediments 2018, 18, 148–158. [Google Scholar] [CrossRef]
- Hu, H.; He, J. Comammox—A newly discovered nitrification process in the terrestrial nitrogen cycle. J. Soils Sediments 2017, 17, 2709–2717. [Google Scholar] [CrossRef]
- Bowles, T.M.; Acosta-Martínez, V.; Calderón, F.; Jackson, L.E. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Sun, R.; Guo, X.; Wang, D.; Chu, H. Effects of long-term application of chemical and organic fertilizers on the abundance of microbial communities involved in the nitrogen cycle. Appl. Soil Ecol. 2015, 95, 171–178. [Google Scholar] [CrossRef]
- Chaudhry, V.; Rehman, A.; Mishra, A.; Chauhan, P.S.; Nautiyal, C.S. Changes in bacterial community structure of agricultural land due to long-term organic and chemical amendments. Microb. Ecol. 2012, 64, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, L.; Cui, Z.; Chen, X.; Xie, J.; Hou, Y. Closing the yield gap and achieving high N use efficiency and low apparent N losses. Field Crop. Res. 2017, 209, 39–46. [Google Scholar] [CrossRef]
- Wang, M.; Wang, L.; Cui, Z.; Chen, X.; Hou, Y. Optimizing management of mulched fertigation systems to improve maize production efficiency in northeast China. Agron. J. 2019, 3149, 3140–3149. [Google Scholar] [CrossRef]
- Yusoff, M.Z.M.; Hu, A.; Feng, C.; Shirai, Y.; Hassan, M.A.; Yu, C.P. Influence of pretreated activated sludge for electricity generation in microbial fuel cell application. Bioresour. Technol. 2013, 145, 90–96. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Glo, F.O.; Yarza, P. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- R-Project. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2010; ISBN 3-900051-07-0. Available online: http://www.r-project.org (accessed on 30 March 2020).
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louca, S.; Parfrey, L.W.; Doebeli1, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, L.; Guo, J.; Ray, J.L.; He, J. Impact of long-term fertilization practices on the abundance and composition of soil bacterial communities in Northeast China. Appl. Soil Ecol. 2010, 46, 119–124. [Google Scholar] [CrossRef]
- Luo, P.; Han, X.; Wang, Y.; Han, M.; Shi, H.; Liu, N.; Bai, H. Influence of long-term fertilization on soil microbial biomass, dehydrogenase activity, and bacterial and fungal community structure in a brown soil of northeast China. Ann. Microbiol. 2015, 65, 533–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamer, M.; Chida, Y. Physical activity and risk of neurodegenerative disease: A systematic review of prospective evidence. Psychol. Med. 2009, 39, 3–11. [Google Scholar] [CrossRef]
- Chu, H.; Fujii, T.; Morimoto, S.; Lin, X.; Yagi, K.; Hu, J.; Zhang, J. Community structure of ammonia-oxidizing bacteria under long-term application of mineral fertilizer and organic manure in a sandy loam soil. Appl. Environ. Microbiol. 2007, 73, 485–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Guan, D.; Zhou, B.; Zhao, B.; Ma, M.; Qin, J. Influence of 34-years of fertilization on bacterial communities in an intensively cultivated black soil in northeast China. Soil Biol. Biochem. 2015, 90, 42–51. [Google Scholar] [CrossRef]
- Zhao, J.; Ni, T.; Li, Y.; Xiong, W.; Ran, W.; Shen, B.; Shen, Q.; Zhang, R. Responses of bacterial communities in arable soils in a rice-wheat cropping system to different fertilizer regimes and sampling times. PLoS ONE 2014, 9, e85301. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Liu, X.; Song, L.; Lin, X.; Zhang, H.H.; Shen, C.; Chu, H. Nitrogen fertilization directly affects soil bacterial diversity and indirectly affects bacterial community composition. Soil Biol. Biochem. 2016, 92, 41–49. [Google Scholar] [CrossRef]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Nemergut, D.R.; Townsend, A.R.; Sattin, S.R.; Freeman, K.R.; Fierer, N.; Neff, J.C.; Bowman, W.D.; Schadt, C.W.; Weintraub, M.N.; Schmidt, S.K. The effects of chronic nitrogen fertilization on alpine tundra soil microbial communities: Implications for carbon and nitrogen cycling. Environ. Microbiol. 2008, 10, 3093–3105. [Google Scholar] [CrossRef] [PubMed]
- Lipson, D.A.; Schmidt, S.K. Seasonal changes in an alpine soil bacterial community in the Colorado Rocky mountains. Appl. Environ. Microbiol. 2004, 70, 2867–2879. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Jiang, X.; Wei, D.; Zhao, B.; Ma, M.; Chen, S. Consistent effects of nitrogen fertilization on soil bacterial communities in black soils for two crop seasons in China. Sci. Rep. 2017, 7, 3267. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Sun, H.; Yang, R.; Li, S.; Zhou, M.; Gao, T.; An, L.; Chen, X.; Dyson, P. Complete genome sequence of a psychotrophic Pseudarthrobacter sulfonivorans strain Ar51 (CGMCC 4.7316), a novel crude oil and multi benzene compounds degradation strain. J. Biotechnol. 2016, 231, 81–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Zheng, S.; Cao, C.; Li, C. Tillage practices and straw-returning methods affect topsoil bacterial community and organic C under a rice-wheat cropping system in central China. Sci. Rep. 2016, 6, 33155. [Google Scholar] [CrossRef] [Green Version]
- De Vries, F.T.; Hoffland, E.; Eekeren, N. Van Fungal/bacterial ratios in grasslands with contrasting nitrogen management. Soil Biol. Biochem. 2006, 38, 2092–2103. [Google Scholar] [CrossRef] [Green Version]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Cinderby, S.; Davidson, E.; Dentener, F.; Emmett, B.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis stable. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, K.S.; Lauber, C.L.; Knight, R.; Bradford, M.A.; Fierer, N. Consistent effects of nitrogen fertilization on soil bacterial communities in contrasting systems. Ecology 2010, 91, 3463–3470. [Google Scholar] [CrossRef]
- Bascompte, J. Networks in ecology. Basic Appl. Ecol. 2007, 8, 485–490. [Google Scholar] [CrossRef]
- Layeghifard, M.; Hwang, D.M.; Guttman, D.S. Disentangling interactions in the microbiome: A network perspective. Trends Microbiol. 2017, 25, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Yan, W.; Shangguan, Z. Impact of long-term N additions upon coupling between soil microbial community structure and activity, and nutrient-use ef fi ciencies. Soil Biol. Biochem. 2015, 91, 151–159. [Google Scholar] [CrossRef]
- Zul, D.; Denzel, S.; Kotz, A. Effects of plant biomass, plant diversity, and water content on bacterial communities in soil lysimeters: Implications for the determinants of bacterial diversity. Appl. Environ. Microbiol. 2007, 73, 6916–6929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wessén, E.; Hallin, S.; Philippot, L. Differential responses of bacterial and archaeal groups at high taxonomical ranks to soil management. Soil Biol. Biochem. 2010, 42, 1759–1765. [Google Scholar] [CrossRef]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Nitrogen fertilization inhibits soil microbial respiration regardless of the form of nitrogen applied. Soil Biol. Biochem. 2010, 42, 2336–2338. [Google Scholar] [CrossRef]
- Garrity, E.G.; Brenner, D.J.; Krieg, N.R.; Eds, J.R.S. Bergey’s manual of systematic bacteriology one of the most comprehensive and authoritative works in the field of bacterial taxonomy Volume 1; Springer: Berlin, Germany, 2005; Volume 2, p. 24144. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Tourova, T.P.; Braker, G.; Muyzer, G. Thiohalomonas denitrificans gen. nov., sp. nov. and Thiohalomonas nitratireducens sp. nov., novel obligately chemolithoautotrophic, moderately halophilic, thiodenitrifying Gammaproteobacteria from hypersaline habitats. Int. J. Syst. Evol. Microbiol. 2007, 57, 1582–1589. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | OC (mg kg−1) | pH | EC | NH4+-N (mg kg−1) | NO3−-N (mg kg−1) | AN (mg kg−1) |
|---|---|---|---|---|---|---|
| N0 a | 12.56 ± 1.27a b | 6.96 ± 0.09a | 0.05 ± 0.03b | 0.34 ± 0.07b | 3.38 ± 0.27d | 92.65 ± 1.55c |
| N1 | 13.84 ± 0.20a | 6.86 ± 0.26ba | 0.08 ± 0.02b | 0.59 ± 0.09a | 4.36 ± 0.11d | 93.47 ± 0.76c |
| N2 | 14.10 ± 0.32a | 6.71 ± 0.11ba | 0.06 ± 0.03b | 0.71 ± 0.15a | 7.38 ± 0.53c | 94.01 ± 1.31c |
| N3 | 13.86 ± 0.74a | 6.69 ± 0.31ba | 0.18 ± 0.02a | 0.75 ± 0.03a | 9.82 ± 1.82b | 107.70 ± 6.56b |
| N4 | 10.39 ± 1.15b | 6.52 ± 0.30b | 0.14 ± 0.03a | 0.77 ± 0.13a | 12.38 ± 1.94a | 114.86 ± 3.23a |
| Bacterial Functional Group | Genus | N0 a | N1 | N2 | N3 | N4 |
|---|---|---|---|---|---|---|
| Nitrate_reduction | 16 | 2.14 ± 0.11b b | 2.33 ± 0.07ba | 2.42 ± 0.12a | 2.30 ± 0.11ba | 2.14 ± 0.12b |
| Ureolysis | 9 | 2.02 ± 0.02a | 2.03 ± 0.06a | 2.01 ± 0.13a | 1.92 ± 0.21a | 2.01 ± 0.10a |
| Nitrate_respiration | 6 | 1.81 ± 0.25ba | 1.67 ± 0.12b | 2.08 ± 0.06a | 1.97 ± 0.21ba | 1.86 ± 0.25ba |
| Nitrate_denitrification | 3 | 0.26 ± 0.04ba | 0.34 ± 0.11a | 0.35 ± 0.08a | 0.22 ± 0.01b | 0.21 ± 0.03b |
| Nitrogen_fixation | 2 | 0.56 ± 0.12a | 0.33 ± 0.04b | 0.50 ± 0.04a | 0.31 ± 0.03b | 0.29 ± 0.04b |
| Nitrate_ammonification | 2 | 0.56 ± 0.12a | 0.33 ± 0.04b | 0.50 ± 0.04a | 0.31 ± 0.03b | 0.29 ± 0.04b |
| Aerobic_ammonia_oxidation | 1 | 0.93 ± 0.07b | 1.12 ± 0.09ba | 1.40 ± 0.13a | 1.20 ± 0.24ba | 1.35 ± 0.26a |
| Aerobic_nitrite_oxidation | 1 | 0.02 ± 0.00c | 0.04 ± 0.01b | 0.03 ± 0.01c | 0.03 ± 0.01cb | 0.05 ± 0.01a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Wang, L.; Li, Q.; Liu, H.; Lin, Y.; Wang, L. Nitrogen Fertilizer Driven Bacterial Community Structure in a Semi-Arid Region of Northeast China. Sustainability 2021, 13, 11967. https://doi.org/10.3390/su132111967
Wang M, Wang L, Li Q, Liu H, Lin Y, Wang L. Nitrogen Fertilizer Driven Bacterial Community Structure in a Semi-Arid Region of Northeast China. Sustainability. 2021; 13(21):11967. https://doi.org/10.3390/su132111967
Chicago/Turabian StyleWang, Meng, Ling Wang, Qian Li, Hang Liu, Yuan Lin, and Lichun Wang. 2021. "Nitrogen Fertilizer Driven Bacterial Community Structure in a Semi-Arid Region of Northeast China" Sustainability 13, no. 21: 11967. https://doi.org/10.3390/su132111967