
Application of Zinc Fertilizer and Mycorrhizal Inoculation on Physio-Biochemical Parameters of Wheat Grown under Water-Stressed Environment

 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Site
2.2. Seed Sterilization
2.3. Experimental Design
2.4. Seed Sowing and Application of Treatments
2.5. Plant Harvesting and Growth Attributes
2.6. Chlorophyll Contents
2.7. Electrolyte Leakage
2.8. Evaluation of Protein Contents
2.9. Estimation of Sugars, Proline, and Non-Enzymatic Antioxidants
2.10. Oxidative Stress Indicators
2.11. CAT, POD and SOD Assay
2.12. Na, Ca, P, and K Concentration in Leaves
2.13. Statistical Analysis
3. Results
3.1. Growth Parameters and Chlorophyll Contents
3.2. Antioxidant Contents and Osmolytes
3.3. Oxidative Stress Indicators and Ionic Constituents
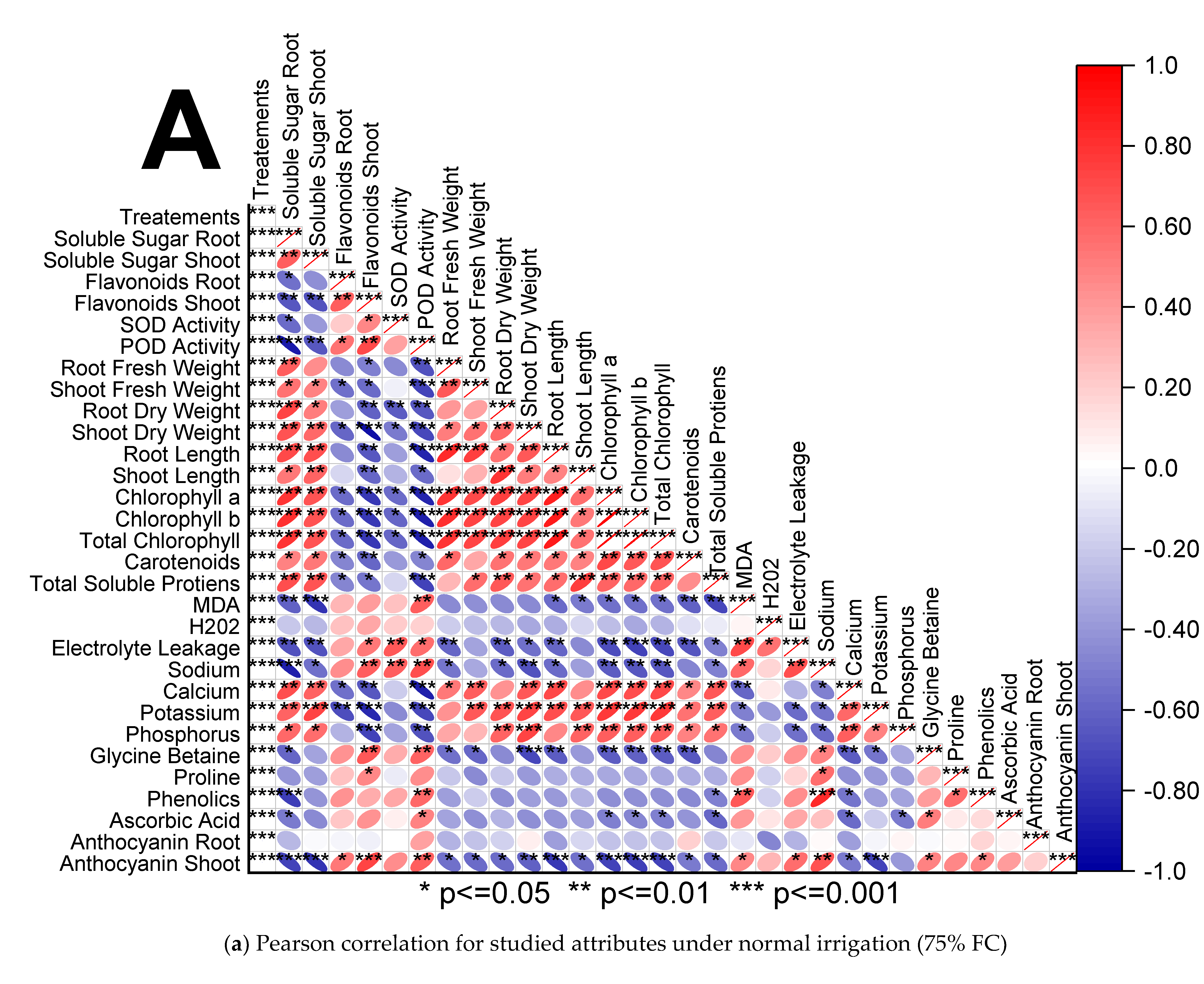
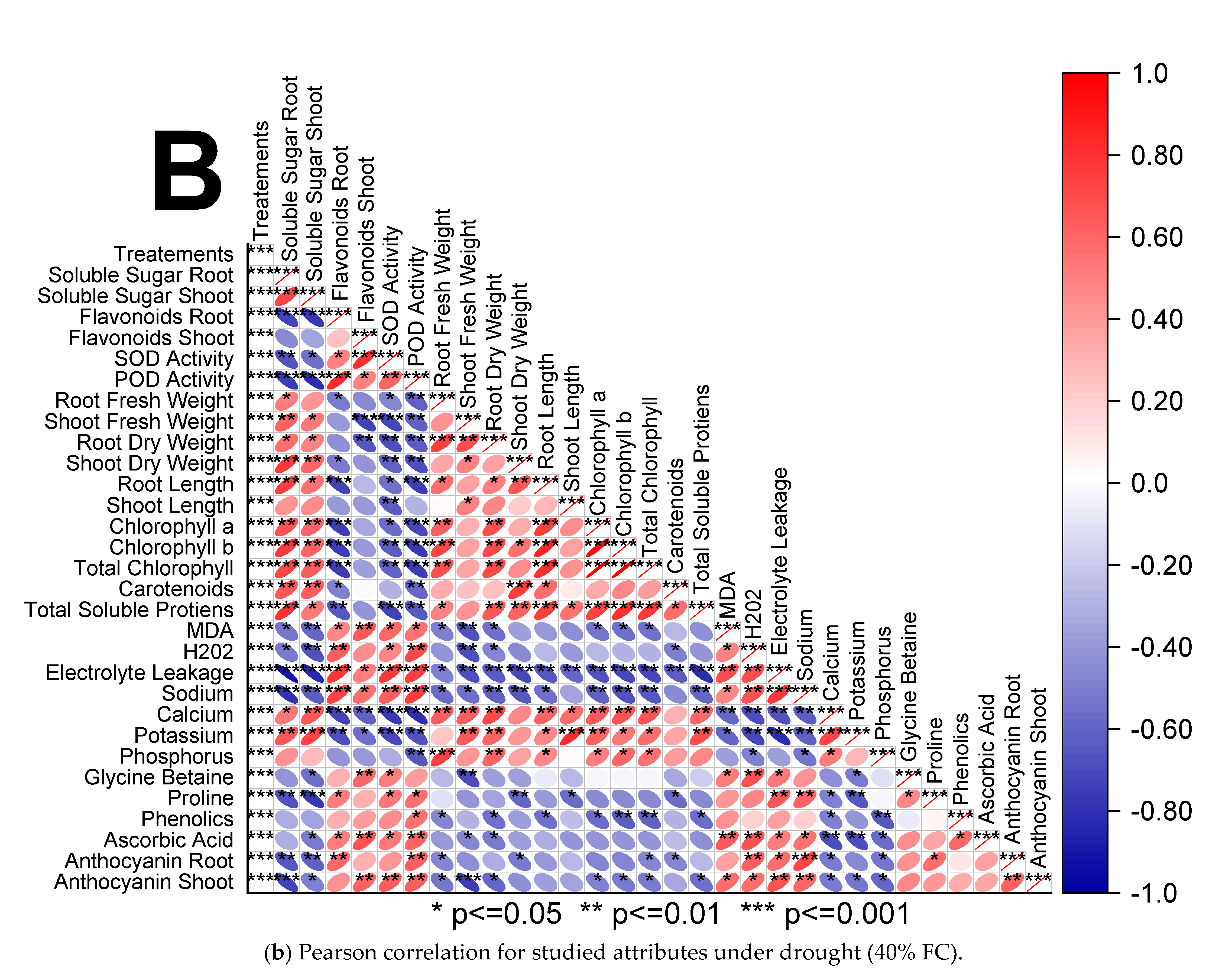
3.4. Pearson Correlation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karim, M.R.; Rahman, M.A. Drought risk management for increased cereal production in Asian least developed countries. Weather Clim. Extrem. 2015, 7, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Danish, S.; Zafar-Ul-Hye, M.; Hussain, S.; Riaz, M.; Qayyum, M.F. Mitigation of drought stress in maize through inoculation with drought tolerant ACC deaminase containing PGPR under axenic conditions. Pak. J. Bot. 2019, 52, 49–60. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-ul-Hye, M.; Mohsin, F.; Hussain, M. ACC-deaminase producing plant growth promoting rhizobacteria and biochar mitigate adverse effects of drought stress on maize growth. PLoS ONE 2020, 15, e0230615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danish, S.; Zafar-ul-Hye, M.; Fahad, S.; Saud, S.; Brtnicky, M.; Hammerschmiedt, T.; Datta, R. Drought stress alleviation by ACC deaminase producing Achromobacter xylosoxidans and Enterobacter cloacae, with and without timber waste biochar in maize. Sustainability 2020, 12, 6286. [Google Scholar] [CrossRef]
- Kheirizadeh Arough, Y.; Seyed Sharifi, R.; Seyed Sharifi, R. Bio fertilizers and zinc effects on some physiological parameters of triticale under water-limitation condition. J. Plant Interact. 2016, 11, 167–177. [Google Scholar] [CrossRef]
- Gusain, Y.S.; Singh, U.S.; Sharma, A.K. Bacterial mediated amelioration of drought stress in drought tolerant and susceptible cultivars of rice (Oryza sativa L.). Afr. J. Biotechnol. 2015, 14, 764–773. [Google Scholar]
- Willis, A.; Rodrigues, B.F.; Harris, P.J.C. The ecology of arbuscular mycorrhizal fungi. Crit. Rev. Plant Sci. 2013, 32, 1–20. [Google Scholar] [CrossRef]
- Pohl, M.; Sprenger, G.A.; Müller, M. A new perspective on thiamine catalysis. Curr. Opin. Biotechnol. 2004, 15, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Zafar-ul-Hye, M.; Zahra, M.B.; Danish, S.; Abbas, M.; Rehim, A.; Akbar, M.N.; Iftikhar, A.; Gul, M.; Nazir, I.; Abid, M.; et al. Multi-strain inoculation with PGPR producing ACC deaminase is more effective than single-strain inoculation to improve wheat (Triticum aestivum) growth and yield. Phyton-Int. J. Exp. Bot. 2020, 89, 405–413. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.A.; Wahid, A.; Ahmad, N.; Saleem, B.A. Improving the drought tolerance in rice (Oryza sativa L.) by exogenous application of salicylic acid. J. Agron. Crop Sci. 2009, 195, 237–246. [Google Scholar] [CrossRef]
- Tahir, F.A.; Ahamad, N.; Rasheed, M.K.; Danish, S. Effect of various application rate of zinc fertilizer with and without fruit waste biochar on the growth and Zn uptake in maize. Int. J. Biosci. 2018, 13, 159–166. [Google Scholar] [CrossRef]
- Bibi, F.; Saleem, I.; Ehsan, S.; Jamil, S.; Ullah, H.; Mubashir, M.; Kiran, S.; Ahmad, I.; Irshad, I.; Saleem, M.; et al. Effect of various application rates of phosphorus combined with different zinc rates and time of zinc application on phytic acid concentration and zinc bioavailability in wheat. Agric. Nat. Resour. 2020, 54, 265–272. [Google Scholar] [CrossRef]
- Rafiullah; Tariq, M.; Khan, F.; Shah, A.H.; Fahad, S.; Wahid, F.; Ali, J.; Adnan, M.; Ahmad, M.; Irfan, M.; et al. Effect of micronutrients foliar supplementation on the production and eminence of plum. Qual. Assur. Saf. Crops Foods 2020, 12, 32–40. [Google Scholar] [CrossRef]
- Malik, K.M.; Khan, K.S.; Rukh, S.; Khan, A.; Akbar, S.; Billah, M.; Bashir, S.; Danish, S.; Alwahibi, M.S.; Elshikh, M.S.; et al. Immobilization of Cd, Pb and Zn through organic amendments in wastewater irrigated soils. Sustainability 2021, 13, 2392. [Google Scholar] [CrossRef]
- Roach, T.; Krieger-Liszkay, A. Regulation of photosynthetic electron transport and photoinhibition. Curr. Protein Pept. Sci. 2014, 15, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Danish, S.; Zafar-ul-Hye, M. Co-application of ACC-deaminase producing PGPR and timber-waste biochar improves pigments formation, growth and yield of wheat under drought stress. Sci. Rep. 2019, 9, 5999. [Google Scholar] [CrossRef] [Green Version]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lewis, C.E.; Walker, J.R.L.; Lancaster, J.E.; Sutton, K.H. Determination of anthocyanins, flavonoids and phenolic acids in potatoes. I: Coloured cultivars of Solanum tuberosum L. J. Sci. Food Agric. 1998, 77, 45–57. [Google Scholar] [CrossRef]
- Lewis, C.E.; Walker, J.R.L.; Lancaster, J.E.; Sutton, K.H. Determination of anthocyanins, flavonoids and phenolic acids in potatoes. II: Wild, tuberous Solanum species. J. Sci. Food Agric. 1998, 77, 58–63. [Google Scholar] [CrossRef]
- Bray, H.G.; Thorpe, W.V. Analysis of phenolic compounds of interest in metabolism. Methods Biochem. Anal. 1954, 1, 27–52. [Google Scholar] [CrossRef]
- Azuma, K.; Nakayama, M.; Koshioka, M.; Ippoushi, K.; Yamaguchi, Y.; Kohata, K.; Yamauchi, Y.; Ito, H.; Higashio, H. Phenolic antioxidants from the leaves of Corchorus olitorius L. J. Agric. Food Chem. 1999, 47, 3963–3966. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Holmström, K.-O.; Somersalo, S.; Mandal, A.; Palva, T.E.; Welin, B. Improved tolerance to salinity and low temperature in transgenic tobacco producing glycine betaine. J. Exp. Bot. 2000, 51, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Jana, S.; Choudhuri, M.A. Glycolate metabolism of three submersed aquatic angiosperms during ageing. Aquat. Bot. 1982, 12, 345–354. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, peroxidase, and polyphenoloxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Jones, J.B., Jr.; Wolf, B.; Mills, H.A. Plant Analysis Handbook. A Practical Sampling, Preparation, Analysis, and Interpretation Guide; Micro-Macro Publishing: Athens, GA, USA, 1991. [Google Scholar]
- Nadeem, F.; Ahmad, R.; Rehmani, M.I.A.; Ali, A.; Ahmad, M.; Iqbal, J. Qualitative and chemical analysis of rice kernel to time of application of phosphorus in combination with zinc under anaerobic conditions. Asian J. Agric. Biol. 2013, 1, 67–75. [Google Scholar]
- Yoshida, S.; Forno, D.A.; Cock, J.H. Laboratory Manual for Physiological Studies of Rice; CAB International: Los Banos, Philippines, 1971. [Google Scholar]
- Amjad, S.F.; Mansoora, N.; Yaseen, S.; Kamal, A.; Butt, B.; Matloob, H.; Alamri, S.A.M.; Alrumman, S.A.; Eid, E.M.; Shahbaz, M. Combined use of endophytic bacteria and pre-sowing treatment of thiamine mitigates the adverse effects of drought stress in wheat (Triticum aestivum L.) Cultivars. Sustainability 2021, 13, 6582. [Google Scholar] [CrossRef]
- Carvalho, T.L.G.; Ballesteros, H.G.F.; Thiebaut, F.; Ferreira, P.C.G.; Hemerly, A.S. Nice to meet you: Genetic, epigenetic and metabolic controls of plant perception of beneficial associative and endophytic diazotrophic bacteria in non-leguminous plants. Plant Mol. Biol. 2016, 90, 561–574. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H.M. Drought stress in plants: An overview. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer: New York, NY, USA, 2012. [Google Scholar]
- Sedghi, M.; Hadi, M.; Toluie, S.G. Effect of nano zinc oxide on the germination parameters of soybean seeds under drought stress. Ann. West Univ. Timisoara Ser. Biol. 2013, 16, 73. [Google Scholar]
- Rafeeq, H.; Arshad, M.A.; Amjad, S.F.; Ullah, M.H.; Imran, H.M.; Khalid, R.; Yaseen, M.; Ajmal, H. Effect of nickel on different physiological parameters of Raphanus sativus. Int. J. Sci. Res. Publ. 2020, 10, 9702. [Google Scholar] [CrossRef]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Rivas-Ubach, A.; Oravec, M.; Vecerova, K.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; et al. Opposite metabolic responses of shoots and roots to drought. Sci. Rep. 2014, 4, 6829. [Google Scholar] [CrossRef] [Green Version]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development; Sinauer Associates: Sunderland, MA, USA, 2015. [Google Scholar]
- Paul, S.; Aggarwal, C.; Manjunatha, B.S.; Rathi, M.S. Characterization of osmotolerant rhizobacteria for plant growth promoting activities in vitro and during plant-microbe association under osmotic stress. Indian J. Exp. Biol. 2018, 56, 582–589. [Google Scholar]
- Gong, M.; Tang, M.; Chen, H.; Zhang, Q.; Feng, X. Effects of two Glomus species on the growth and physiological performance of Sophora davidii seedlings under water stress. New For. 2013, 44, 399–408. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Ding, H.; Qin, H.; Hou, J.; Huang, X.; Xie, Y.; Guo, T. Physiological responses and yield of wheat plants in zinc-mediated alleviation of drought stress. Front. Plant Sci. 2017, 8, 860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, X.; He, Y.; Wang, Z.; Li, X.; Zhang, K.; Zeng, H. Effect of foliar spray of zinc on chloroplast β-carbonic anhydrase expression and enzyme activity in rice (Oryza sativa L.) leaves. Acta Physiol. Plant. 2014, 36, 263–272. [Google Scholar] [CrossRef]
- Cherif, J.; Mediouni, C.; Ammar, W.B.; Jemal, F. Interactions of zinc and cadmium toxicity in their effects on growth and in antioxidative systems in tomato plants (Solarium lycopersicum). J. Environ. Sci. 2011, 23, 837–844. [Google Scholar] [CrossRef]
- Noreen, S.; Sultan, M.; Akhter, M.S.; Shah, K.H.; Ummara, U.; Manzoor, H.; Ulfat, M.; Alyemeni, M.N.; Ahmad, P. Foliar fertigation of ascorbic acid and zinc improves growth, antioxidant enzyme activity and harvest index in barley (Hordeum vulgare L.) grown under salt stress. Plant Physiol. Biochem. 2021, 158, 244–254. [Google Scholar] [CrossRef]
- Tamás, L.; Mistrík, I.; Alemayehu, A.; Zelinová, V.; Bočová, B.; Huttová, J. Salicylic acid alleviates cadmium-induced stress responses through the inhibition of Cd-induced auxin-mediated reactive oxygen species production in barley root tips. J. Plant Physiol. 2015, 173, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Song, C.Z.; Liu, M.Y.; Meng, J.F.; Chi, M.; Xi, Z.M.; Zhang, Z.-W. Promoting effect of foliage sprayed zinc sulfate on accumulation of sugar and phenolics in berries of Vitis vinifera cv. Merlot growing on zinc deficient soil. Molecules 2015, 20, 2536–2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.A.; Eweda, W.E.E.; Heggo, A.M.; Hassan, E.A. Effect of dual inoculation with arbuscular mycorrhizal fungi and sulphur-oxidising bacteria on onion (Allium cepa L.) and maize (Zea mays L.) grown in sandy soil under green house conditions. Ann. Agric. Sci. 2014, 59, 109–118. [Google Scholar] [CrossRef]
- Evelin, H.; Giri, B.; Kapoor, R. Contribution of Glomus intraradices inoculation to nutrient acquisition and mitigation of ionic imbalance in NaCl-stressed Trigonella foenum-graecum. Mycorrhiza 2012, 22, 203–217. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatment (T) | RFW (g) | RL (cm) | RDW (g) | ||||||
| T × FC | Mean (T) | T × FC | Mean (T) | T × FC | Mean (T) | ||||
| 75 FC | 40 FC | 75 FC | 40 FC | 75 FC | 40 FC | ||||
| C | 2.41 cd | 2.05 e | 2.18 C | 0.51 fg | 0.40 g | 0.46 B | 0.35 | 0.29 | 0.28 C |
| Zn 1 | 2.63 c | 2.19 de | 2.42 BC | 0.73 ef | 0.6 ef | 0.67 B | 0.43 | 0.41 | 0.43 BC |
| Zn 2 | 3.03 b | 2.25 de | 2.65 AB | 1.43 bc | 0.9 de | 1.17 A | 0.48 | 0.43 | 0.46 A-C |
| AM | 3.33 ab | 2.26 cd | 2.80 AB | 1.63 ab | 0.83 ef | 1.23 A | 0.46 | 0.44 | 0.45 A-C |
| Zn1+AM | 3.24 ab | 2.39 cd | 2.93 A | 1.70 ab | 0.9 de | 1.30 A | 0.52 | 0.54 | 0.53 AB |
| Zn2+AM | 3.46 a | 2.51 cd | 2.88 A | 1.8 a | 1.16 cd | 1.48 A | 0.61 | 0.55 | 0.58 A |
| Mean (FC) | 3.02 A | 2.26 B | 1.30 A | 0.80 B | 0.48 | 0.43 | |||
| p-value | Treatment | 4.33124 × 10−5 | 9.0904 × 10−9 | 2.8426 × 10−4 | |||||
| FC | 4.63411 × 10−10 | 5.75823 × 10−8 | 0.2667 ns | ||||||
| Interaction | 0.03579 | 0.0054 | 0.95431 ns | ||||||
| Treatment (T) | SDW (g) | SFW (g) | SL (cm) | ||||||
| C | 0.51 | 0.30 | 0.41 C | 0.64 | 0.46 | 0.51 C | 2.16 | 1.77 | 1.97 B |
| Zn 1 | 0.59 | 0.43 | 0.52 BC | 0.92 | 0.71 | 0.82 BC | 2.36 | 1.73 | 2.05 AB |
| Zn 2 | 0.62 | 0.44 | 0.53 BC | 1.32 | 0.94 | 1.14 A-C | 2.43 | 2.03 | 2.23 AB |
| AM | 0.55 | 0.39 | 0.47 C | 1.36 | 1.14 | 1.25 AB | 2.26 | 1.66 | 1.97 B |
| Zn1+AM | 0.92 | 0.66 | 0.79 AB | 1.52 | 1.34 | 1.43 AB | 2.50 | 1.96 | 2.23 AB |
| Zn2+AM | 1.06 | 0.86 | 0.97 A | 1.57 | 1.51 | 1.54 A | 2.66 | 2.19 | 2.43 A |
| Mean (FC) | 0.71 A | 0.52 B | 1.23 | 1.00 | 2.40 A | 1.89 B | |||
| p-value | Treatment | 9.45218 × 10−6 | 3.72445 × 10−4 | 0.01609 | |||||
| FC | 9.90009 × 10−4 | 0.0891 ns | 1.99655 × 10−6 | ||||||
| Interaction | 0.99351 ns | 0.98412 ns | 0.96762 ns | ||||||
| Treatment (T) | Chla (mg/g) | Chlb (mg/g) | TChl (mg/g) | ||||||
| C | 0.50 | 0.29 | 0.39 C | 0.33 | 0.11 | 0.22 D | 0.82 | 0.40 | 0.61 D |
| Zn 1 | 0.84 | 0.68 | 0.76 B | 0.69 | 0.55 | 0.62 C | 1.52 | 1.23 | 1.38 C |
| Zn 2 | 1.66 | 1.25 | 1.46 A | 0.83 | 0.63 | 0.73 C | 2.49 | 1.88 | 2.19 B |
| AM | 1.57 | 1.27 | 1.42 A | 1.07 | 0.76 | 0.91 B | 2.63 | 2.03 | 2.33 B |
| Zn1+AM | 1.59 | 1.45 | 1.52 A | 1.22 | 0.99 | 1.10 A | 2.81 | 2.43 | 2.62 A |
| Zn2+AM | 1.63 | 1.53 | 1.58 A | 1.23 | 1.01 | 1.12 A | 2.87 | 2.54 | 2.70 A |
| Mean (FC) | 1.30 A | 1.08 B | 0.89 A | 0.67 B | 2.19 A | 1.75 B | |||
| p-value | Treatment | 6.93916 × 10−17 | 6.31474 × 10−16 | 3.13157 × 10−20 | |||||
| FC | 2.2081 × 10−6 | 1.14979 × 10−8 | 1.48905 × 10−10 | ||||||
| Interaction | 0.06971 ns | 0.59239 ns | 0.0858 ns | ||||||
| Treatment (T) | Car (mg/g) | Pro (μmol/g) | Phen (mg/g) | ||||||
| T × FC | Mean (T) | T × FC | Mean (T) | T × FC | Mean (T) | ||||
| 75 FC | 40 FC | 75 FC | 40 FC | 75 FC | 40 FC | ||||
| C | 0.06 | 0.05 | 0.06 B | 2.96 | 2.84 | 2.90 | 2.18 | 4.92 | 3.55 |
| Zn 1 | 0.07 | 0.06 | 0.07 AB | 2.72 | 3.03 | 2.87 | 1.85 | 2.61 | 2.23 |
| Zn 2 | 0.08 | 0.08 | 0.08 AB | 2.56 | 2.85 | 2.71 | 1.68 | 2.37 | 2.02 |
| AM | 0.08 | 0.06 | 0.07 AB | 3.04 | 3.35 | 3.19 | 1.93 | 2.71 | 2.32 |
| Zn1+AM | 0.10 | 0.07 | 0.08 AB | 2.17 | 2.77 | 2.47 | 1.67 | 2.38 | 2.02 |
| Zn2+AM | 0.13 | 0.11 | 0.12 A | 1.93 | 2.23 | 2.08 | 1.43 | 2.14 | 1.78 |
| Mean (FC) | 0.09 | 0.07 | 2.56 | 2.85 | 1.79 B | 2.85 A | |||
| p-value | Treatment | 0.01486 | 0.14849 ns | 0.12036 ns | |||||
| FC | 0.06848 ns | 0.1351 ns | 4.01679 × 10−4 | ||||||
| Interaction | 0.97112 ns | 0.98515 ns | 0.98447 ns | ||||||
| Treatment (T) | Asa (mg/g) | AnthR (μg/g) | AnthS (μg/g) | ||||||
| C | 23.67 | 31.00 | 27.33 A | 0.81 | 1.21 | 1.01 ns | 1.25 | 1.71 | 1.48 A |
| Zn 1 | 22.33 | 25.33 | 23.83 AB | 0.64 | 1.41 | 1.02 ns | 1.04 | 1.41 | 1.22 AB |
| Zn 2 | 19.67 | 23.00 | 21.33 AB | 0.44 | 1.35 | 0.89 ns | 0.69 | 1.20 | 0.94 A-C |
| AM | 22.00 | 26.67 | 24.33 AB | 0.33 | 1.42 | 0.88 ns | 0.76 | 1.33 | 1.05 A-C |
| Zn1+AM | 21.00 | 23.67 | 22.33 AB | 0.51 | 1.11 | 0.81 ns | 0.50 | 0.95 | 0.73 BC |
| Zn2+AM | 18.30 | 22.33 | 20.32 B | 0.74 | 0.95 | 0.85 ns | 0.49 | 0.73 | 0.61 C |
| Mean (FC) | 21.16B | 25.33 A | 0.58 B | 1.24 A | 0.79 B | 1.22 A | |||
| p-value | Treatment | 0.02465 | 0.25072 ns | 0.00101 | |||||
| FC | 7.65328 × 10−4 | 8.18068 × 10−9 | 4.62156 × 10−4 | ||||||
| Interaction | 0.97757 ns | 0.07905 ns | 0.96471 ns | ||||||
| Treatment (T) | SSR (mg/g) | SSS (mg/g) | FlavR (μg/g) | ||||||
| C | 1.12 | 1.09 | 1.11 D | 1.38 | 1.03 | 1.21 D | 0.83 | 0.81 | 0.82 A |
| Zn 1 | 1.36 | 1.04 | 1.20 CD | 1.65 | 1.31 | 1.48 CD | 0.55 | 0.92 | 0.74 AB |
| Zn 2 | 1.47 | 1.20 | 1.33 A-C | 2.00 | 1.48 | 1.74 A-C | 0.49 | 0.73 | 0.61 AB |
| AM | 1.39 | 1.12 | 1.26 BC | 1.76 | 1.33 | 1.55 BC | 0.65 | 0.87 | 0.76 AB |
| Zn1+AM | 1.49 | 1.38 | 1.44 AB | 2.13 | 1.79 | 1.96 AB | 0.38 | 0.60 | 0.49 B |
| Zn2+AM | 1.59 | 1.49 | 1.54 A | 2.31 | 1.81 | 2.06 A | 0.26 | 0.47 | 0.36 B |
| Mean (FC) | 1.40 A | 1.22 B | 1.87 A | 1.46 B | 0.53 B | 0.73 A | |||
| p-value | Treatment | 1.77298 × 10−6 | 1.97823 × 10−4 | 0.00377 | |||||
| FC | 2.05111 × 10−5 | 2.15272 × 10−4 | 0.00375 | ||||||
| Interaction | 0.49373 ns | 0.97856 ns | 0.99365 ns | ||||||
| Treatment (T) | FlavS (μg/g) | SOD (U g−1) | POD (U g−1) | ||||||
| C | 0.81 | 1.71 | 1.26 A | 1.16 | 2.12 | 1.64 A | 4.62 | 6.10 | 5.36 A |
| Zn 1 | 0.68 | 1.15 | 0.91 AB | 0.96 | 1.77 | 1.36 A | 3.54 | 4.23 | 3.88 B |
| Zn 2 | 0.56 | 0.96 | 0.76 A-C | 0.71 | 1.62 | 1.17 AB | 2.67 | 3.79 | 3.23 BC |
| AM | 0.76 | 1.09 | 0.92 AB | 0.83 | 1.76 | 1.30 AB | 3.08 | 4.06 | 3.57 B |
| Zn1+AM | 0.39 | 0.62 | 0.50 BD | 0.43 | 1.28 | 0.86 AB | 2.68 | 3.05 | 2.87 BC |
| Zn2+AM | 0.23 | 0.55 | 0.39 C | 0.15 | 0.73 | 0.44 B | 1.97 | 2.47 | 2.22 C |
| Mean (FC) | 0.57 B | 1.01 A | 0.71 B | 1.55 A | 3.09 B | 3.95 A | |||
| p-value | Treatment | 0.00202 | 0.00725 | 6.69258 × 10−8 | |||||
| FC | 4.97548 × 10−4 | 3.74663 × 10−5 | 1.81577 × 10−4 | ||||||
| Interaction | 0.92577 ns | 0.99049 ns | 0.5551 | ||||||
| Treatment (T) | CAT (U g−1 FW) | TSP (mg/g FW) | MDA (μmol/g) | ||||||
| T × FC | Mean (T) | T × FC | Mean (T) | T × FC | Mean (T) | ||||
| 75 FC | 40 FC | 75 FC | 40 FC | 75 FC | 40 FC | ||||
| C | 1.10 | 1.20 | 1.15 ns | 2.80 | 2.10 | 2.45 C | 0.85 | 1.85 | 1.35 A |
| Zn 1 | 0.82 | 1.15 | 0.98 ns | 3.33 | 2.69 | 3.01 BC | 0.79 | 1.22 | 1.01 AB |
| Zn 2 | 0.74 | 1.07 | 0.90 ns | 3.59 | 3.26 | 3.43 A-C | 0.61 | 1.11 | 0.86 AB |
| AM | 0.85 | 1.18 | 1.01 ns | 3.28 | 2.53 | 2.91 BC | 0.70 | 1.13 | 0.91 AB |
| Zn1+AM | 0.52 | 0.85 | 0.68 ns | 3.86 | 3.36 | 3.61 AB | 0.55 | 1.09 | 0.82 B |
| Zn2+AM | 0.45 | 0.79 | 0.62 ns | 4.23 | 3.99 | 4.11 A | 0.36 | 0.78 | 0.57 B |
| Mean (FC) | 0.75 B | 1.04 A | 3.52 A | 2.99 B | 0.64 B | 1.20 A | |||
| p-value | Treatment | 0.15084 ns | 5.4797 × 10−4 | 0.00301 | |||||
| FC | 0.0268 | 0.01031 | 4.46462 × 10−6 | ||||||
| Interaction | 0.99915 ns | 0.96292 ns | 0.45346 ns | ||||||
| Treatment (T) | H2O2 (μmol/g) | EL (%) | Na (μg/g) | ||||||
| C | 2.47 | 3.00 | 2.74 A | 22.67 | 30.00 | 26.33 A | 14.83 | 18.00 | 16.42 A |
| Zn 1 | 2.32 | 3.09 | 2.70 AB | 21.67 | 29.67 | 25.67 AB | 13.33 | 20.83 | 17.08 A |
| Zn 2 | 2.19 | 2.68 ns | 2.44 AB | 19.00 | 27.00 | 23.00 BC | 12.67 | 19.00 | 15.83 AB |
| AM | 2.23 | 2.96 | 2.59 AB | 20.00 | 28.33 | 24.17 A-C | 13.33 | 19.67 | 16.50 A |
| Zn1+AM | 2.14 | 2.53 | 2.34 AB | 17.33 | 22.67 | 20.00 CD | 11.00 | 14.00 | 12.50 BC |
| Zn2+AM | 2.01 | 2.27 | 2.14 B | 15.67 | 19.67 | 17.67 D | 10.00 | 13.83 | 11.92 C |
| Mean (FC) | 2.23B | 2.75 A | 19.39 B | 26.22 A | 12.53 B | 17.56 A | |||
| p-value | Treatment | 0.04916 | 2.09285 × 10−6 | 4.91172 × 10−5 | |||||
| FC | 2.31731 × 10−4 | 3.42732 × 10−9 | 2.12199 × 10−8 | ||||||
| Interaction | 0.7903 ns | 0.35393 ns | 0.35391 ns | ||||||
| Treatment (T) | Ca (%) | K (%) | P (%) | ||||||
| C | 1.57 | 1.20 | 1.38 C | 2.83 | 2.00 | 2.42 C | 0.12 | 0.04 | 0.08 B |
| Zn 1 | 2.20 | 1.87 | 2.03 BC | 3.60 | 2.50 | 3.05 BC | 0.13 | 0.12 | 0.12 AB |
| Zn 2 | 3.00 | 2.10 | 2.55 BC | 4.67 | 3.67 | 4.17 AB | 0.14 | 0.13 | 0.13 AB |
| AM | 2.33 | 1.80 | 2.07 AB | 3.50 | 2.57 | 3.03 BC | 0.12 | 0.11 | 0.12 AB |
| Zn1+AM | 2.67 | 2.20 | 2.43 AB | 5.00 | 3.83 | 4.42 A | 0.14 | 0.14 | 0.14 AB |
| Zn2+AM | 3.50 | 2.96 | 3.23 A | 5.67 | 4.33 | 5.00 A | 0.17 | 0.15 | 0.16 A |
| Mean (FC) | 2.54 A | 2.02 B | 4.21 A | 3.15 B | 0.14 ns | 0.11 ns | |||
| p-value | Treatment | 4.61249 × 10−5 | 3.11022 × 10−6 | 0.01961 | |||||
| FC | 0.00407 | 7.41822 × 10−5 | 0.14451 ns | ||||||
| Interaction | 0.92374 ns | 0.9964 ns | 0.9518 ns | ||||||
| Treatment (T) | GB (μmol/g) | ||||||||
| C | 1.20 | 1.69 | 1.45 | ||||||
| Zn 1 | 1.12 | 1.78 | 1.45 | ||||||
| Zn 2 | 0.93 | 1.50 | 1.21 | ||||||
| AM | 0.99 | 1.83 | 1.41 | ||||||
| Zn1+AM | 0.69 | 1.22 | 0.95 | ||||||
| Zn2+AM | 0.33 | 1.10 | 0.71 | ||||||
| Mean (FC) | 0.88 B | 1.52 A | |||||||
| p-value | Treatment | 0.0597 ns | |||||||
| FC | 5.35532 × 10−4 | ||||||||
| Interaction | 0.99143 ns | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amjad, S.F.; Mansoora, N.; Din, I.U.; Khalid Iqbal, R.; Jatoi, G.H.; Murtaza, G.; Yaseen, S.; Naz, M.; Danish, S.; Fahad, S.; et al. Application of Zinc Fertilizer and Mycorrhizal Inoculation on Physio-Biochemical Parameters of Wheat Grown under Water-Stressed Environment. Sustainability 2021, 13, 11007. https://doi.org/10.3390/su131911007
Amjad SF, Mansoora N, Din IU, Khalid Iqbal R, Jatoi GH, Murtaza G, Yaseen S, Naz M, Danish S, Fahad S, et al. Application of Zinc Fertilizer and Mycorrhizal Inoculation on Physio-Biochemical Parameters of Wheat Grown under Water-Stressed Environment. Sustainability. 2021; 13(19):11007. https://doi.org/10.3390/su131911007
Chicago/Turabian StyleAmjad, Syeda Fasiha, Nida Mansoora, Israr Ud Din, Rana Khalid Iqbal, Ghulam Hussain Jatoi, Ghulam Murtaza, Samia Yaseen, Maliha Naz, Subhan Danish, Shah Fahad, and et al. 2021. "Application of Zinc Fertilizer and Mycorrhizal Inoculation on Physio-Biochemical Parameters of Wheat Grown under Water-Stressed Environment" Sustainability 13, no. 19: 11007. https://doi.org/10.3390/su131911007