
Effects of Soil pH on Gaseous Nitrogen Loss Pathway via Feammox Process

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isotopic Tracer Incubation
2.2. Anaerobic Chemodenitrification Experiment
2.3. Physicochemical Analysis
2.4. Microbial Community Analysis
2.5. Statistical Analyses
3. Results and Discussion
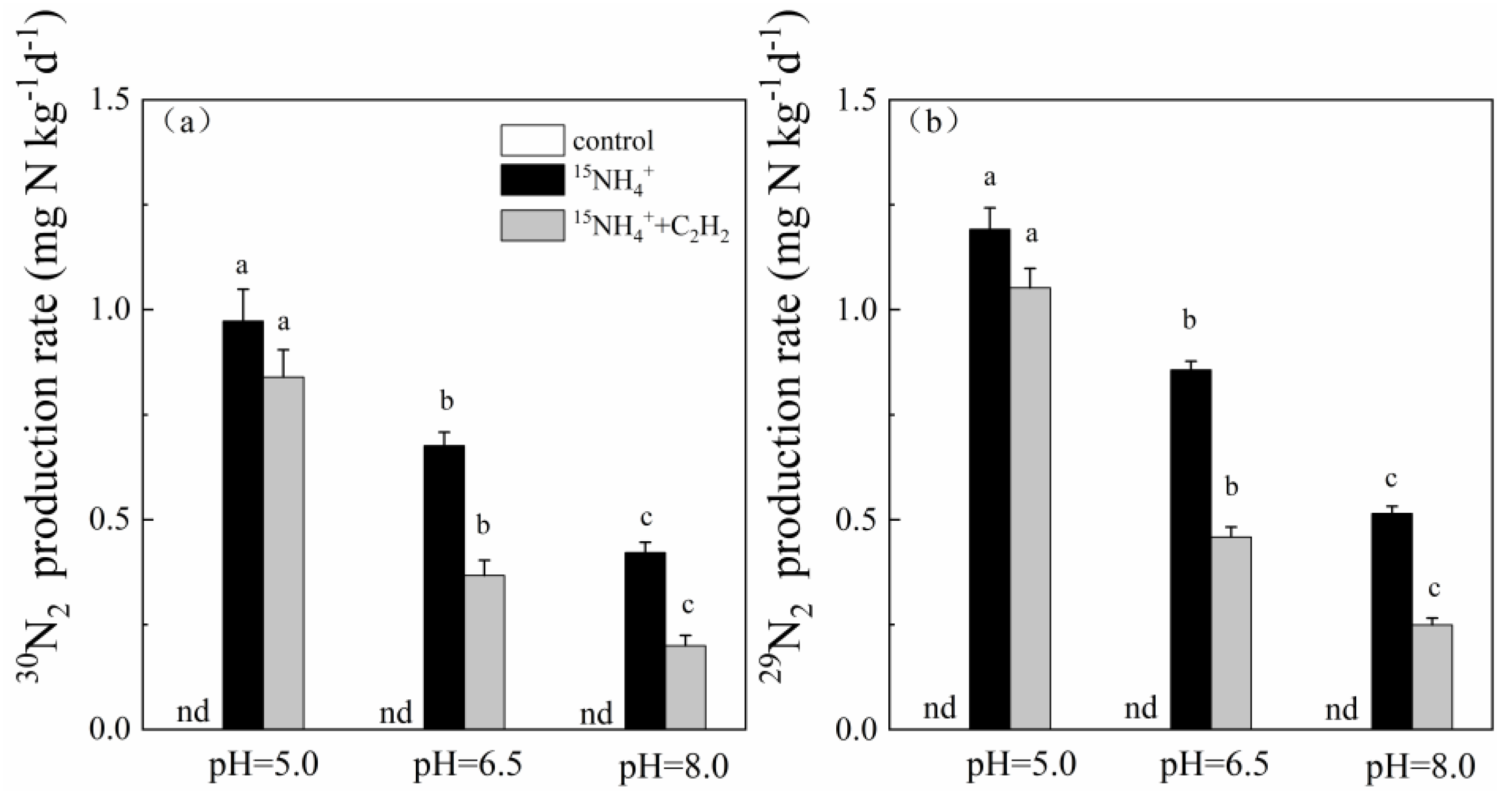
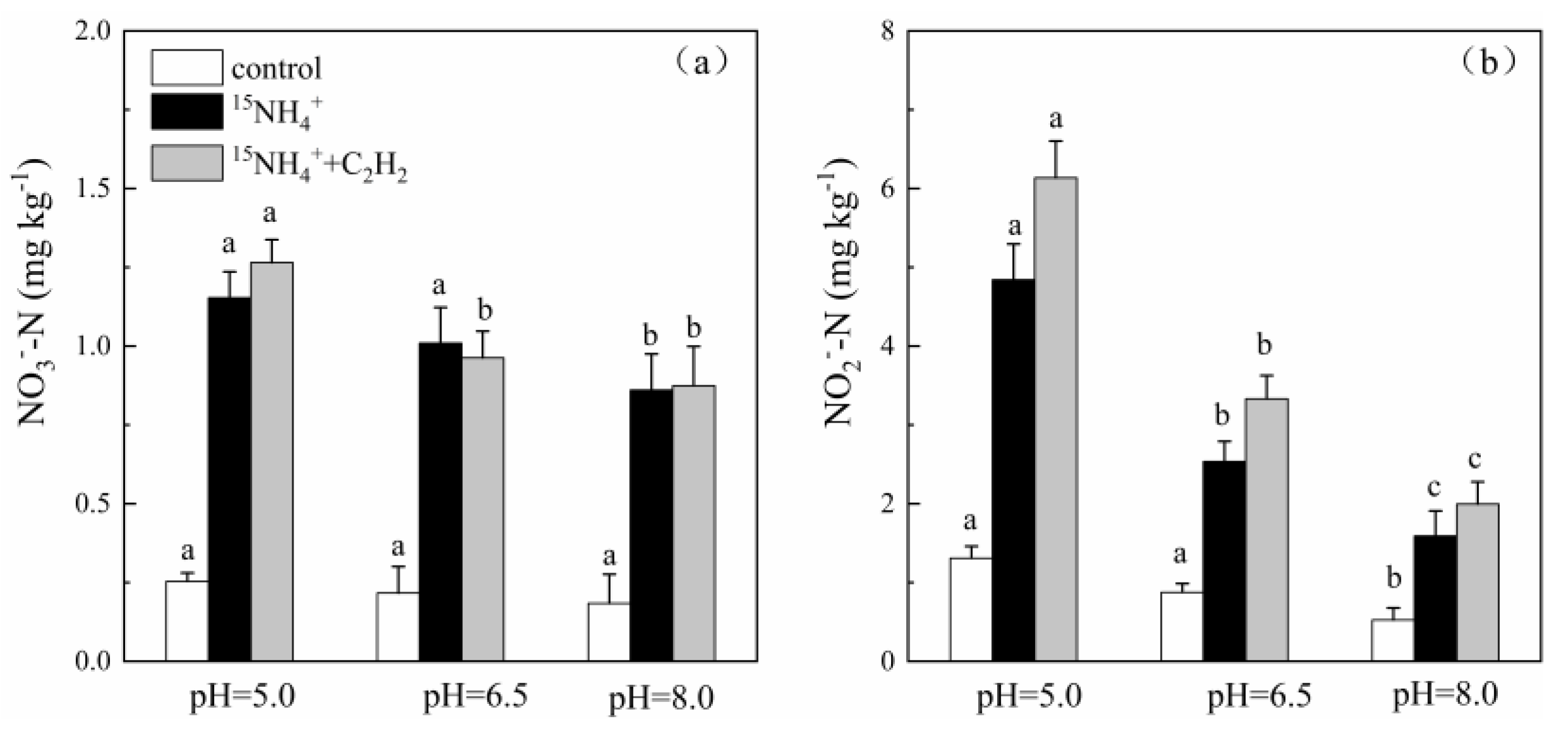
3.1. pH-Dependent Feammox Rates and Pathways
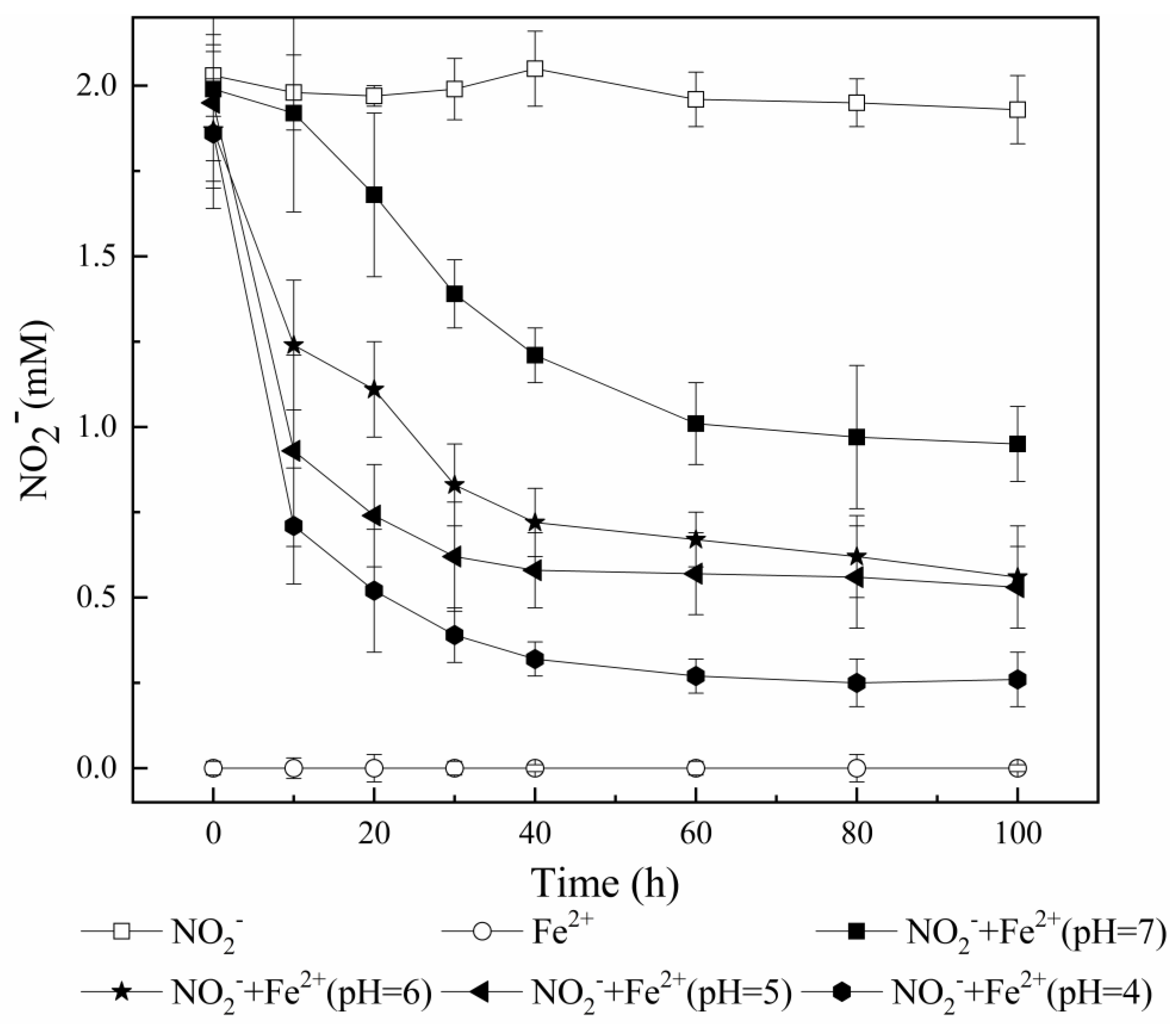
3.2. Potential Transformation Pathways of NO2−
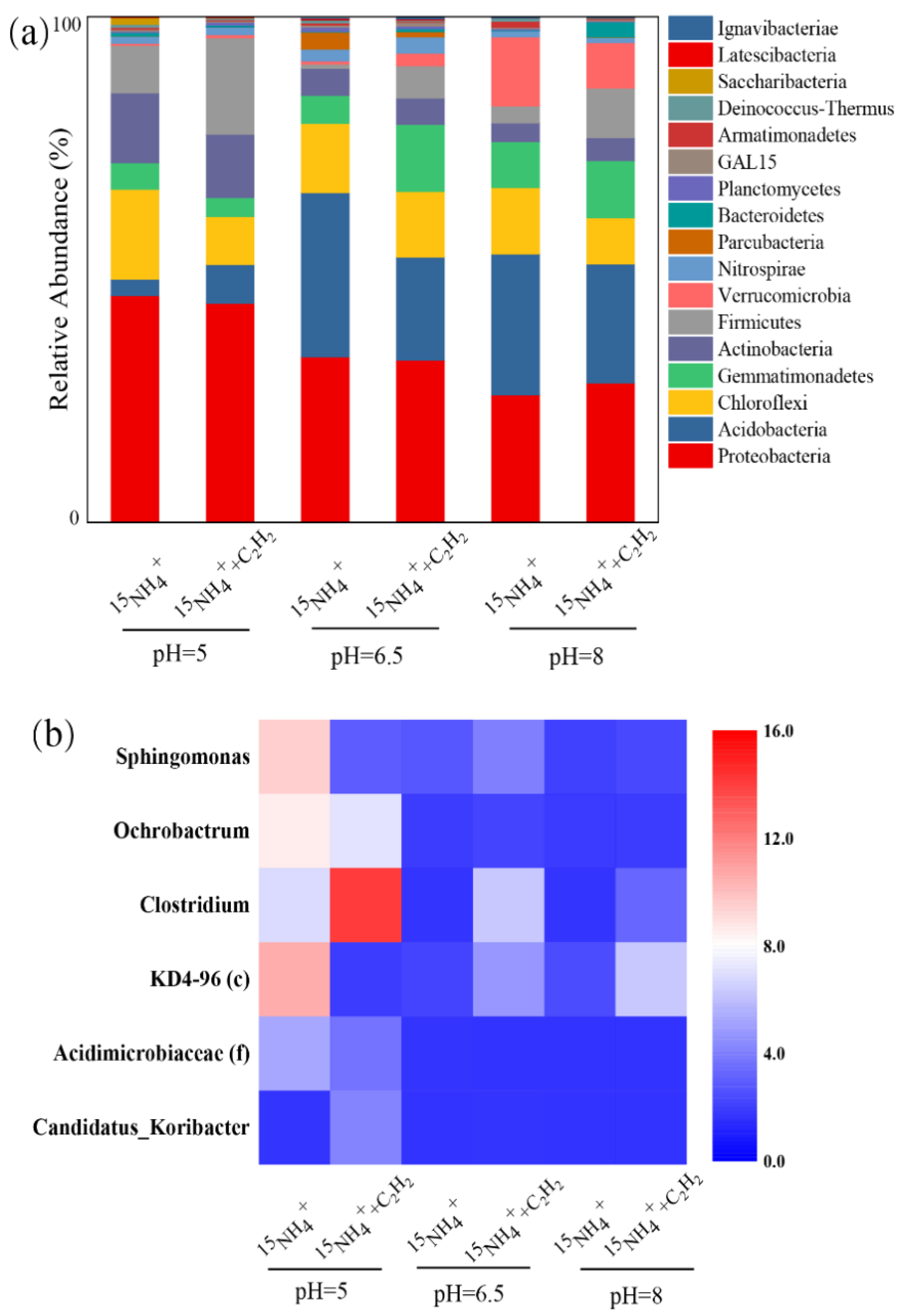
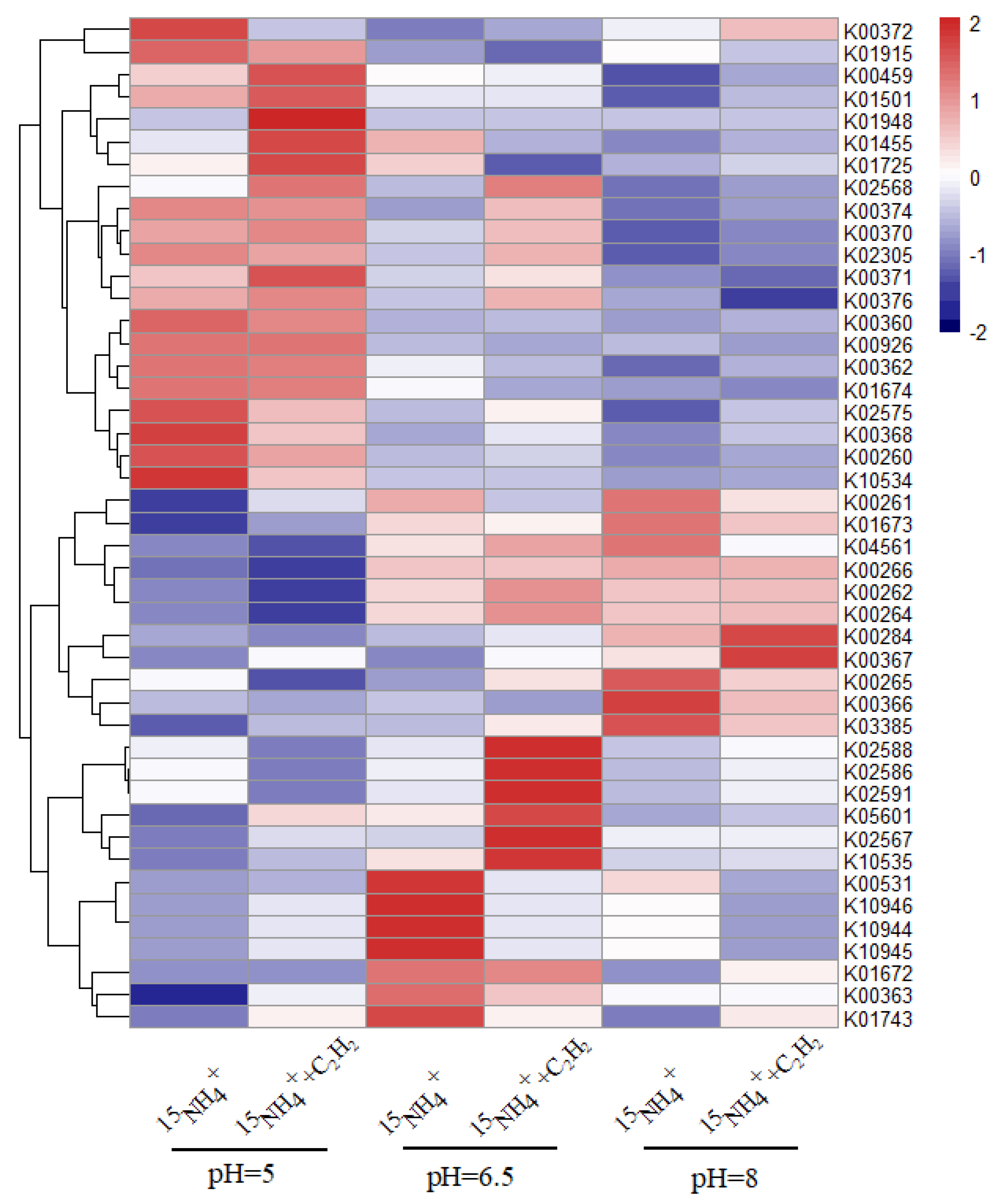
3.3. Evolution of the Microbial Community in Feammox
3.4. Environmental Implications
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, J.; Xu, C.C.; Ridoutt, B.G.; Wang, X.C.; Ren, P.A. Nitrogen and phosphorus losses and eutrophication potential associated with fertilizer application to cropland in China. J. Clean. Prod. 2017, 159, 171–179. [Google Scholar] [CrossRef]
- Yu, C.; Huang, X.; Chen, H.; Godfray, H.C.J.; Wright, J.S.; Hall, J.W.; Gong, P.; Ni, S.; Qiao, S.; Huang, G.; et al. Managing nitrogen to restore water quality in China. Nature 2019, 567, 516–520. [Google Scholar] [CrossRef]
- Guo, J.H.; Liu, X.J.; Zhang, Y.; Shen, J.L.; Han, W.X.; Zhang, W.F.; Christie, P.; Goulding, K.W.T.; Vitousek, P.M.; Zhang, F.S. Significant Acidification in Major Chinese Croplands. Science 2010, 327, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greaver, T.L.; Clark, C.M.; Compton, J.E.; Vallano, D.; Talhelm, A.F.; Weaver, C.P.; Band, L.E.; Baron, J.S.; Davidson, E.A.; Tague, C.L.; et al. Key ecological responses to nitrogen are altered by climate change. Nat. Clim. Chang. 2016, 6, 836–843. [Google Scholar] [CrossRef]
- Zhu, Z.L. Research on soil nitrogen in China. Acta Pedol. Sinica. 2008, 45, 778–783. [Google Scholar]
- Yang, W.H.; Weber, K.A.; Silver, W.L. Nitrogen loss from soil through anaerobic ammonium oxidation coupled to iron reduction. Nat. Geosci. 2012, 5, 538–541. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.J.; An, X.L.; Li, S.; Zhang, G.L.; Zhu, Y.G. Nitrogen loss through anaerobic ammonium oxidation coupled to iron reduction from paddy soils in a chronosequence. Environ. Sci. Technol. 2014, 48, 10641–10647. [Google Scholar] [CrossRef]
- Li, X.; Hou, L.; Liu, M.; Zheng, Y.; Yin, G.; Lin, X.; Cheng, L.; Li, Y.; Hu, X. Evidence of Nitrogen Loss from Anaerobic Ammonium Oxidation Coupled with Ferric Iron Reduction in an Intertidal Wetland. Environ. Sci. Technol. 2015, 49, 11560–11568. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Li, Z.; Qin, Y. Nitrogen loss from anaerobic ammonium oxidation coupled to Iron(III) reduction in a riparian zone. Environ. Pollut. 2017, 231, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Clément, J.; Shrestha, J.; Ehrenfeld, J.; Jaffe, P. Ammonium oxidation coupled to dissimilatory reduction of iron under anaerobic conditions in wetland soils. Soil Biol. Biochem. 2005, 37, 2323–2328. [Google Scholar] [CrossRef]
- Huang, S.; Jaffe, P.R. Isolation and characterization of an ammonium-oxidizing iron reducer: Acidimicrobiaceae sp. A6. PLoS ONE 2018, 13, e0194007. [Google Scholar]
- Tang, Y.; Zhou, C.; Ziv-El, M.; Rittmann, B.E. A pH-control model for heterotrophic and hydrogen-based autotrophic denitrification. Water Res. 2011, 45, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhu-Barker, X.; Horwath, W.R.; Faeflen, S.J.; Luo, H.; Xin, X.; Jiang, X. Effect of iron oxide on nitrification in two agricultural soils with different pH. Biogeosciences 2016, 13, 5609–5617. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Chen, C.; Peng, X.; Jaffé, P.R. Environmental factors affecting the presence of Acidimicrobiaceae and ammonium removal under iron-reducing conditions in soil environments. Soil Biol. Biochem. 2016, 98, 148–158. [Google Scholar] [CrossRef] [Green Version]
- Jensen, M.M.; Thamdrup, B.; Dalsgaard, T. Effects of specific inhibitors on anammox and denitrification in marine sediments. Appl. Environ. Microbiol. 2007, 73, 3151–3158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, S.P.; Hu, C.S.; Oenema, O. Quantifying the under estimation of soil denitrification potential as determined by the acetylene inhibition. Soil Biol. Biochem. 2012, 47, 14–17. [Google Scholar] [CrossRef]
- Kalembasa, S.J.; Jenkinson, D.S. A comparative study of titrimetric and gravimetric methods for the determination of organic carbon in soil. J. Sci. Food Agric. 1973, 24, 1085–1090. [Google Scholar] [CrossRef]
- Li, H.; Su, J.Q.; Yang, X.R.; Zhou, G.W.; Lassen, S.B.; Zhu, Y.G. Rna stable isotope probing of potential feammox population in paddy soil. Environ. Sci. Technol. 2019, 53, 4841–4849. [Google Scholar] [CrossRef]
- Guan, Q.S.; Cao, W.Z.; Wang, M.; Wu, G.J.; Wang, F.F.; Jiang, C.; Tao, Y.R.; Gao, Y. Nitrogen loss through anaerobic ammonium oxidation coupled with iron reduction in a mangrove wetland. Eur. J. Soil Sci. 2018, 69, 732–741. [Google Scholar] [CrossRef]
- Schwertmann, U.; Cornell, R.M. Iron Oxides in the Laboratory. Soil Sci. 1991, 156, 281–282. [Google Scholar] [CrossRef]
- Yang, Y.; Xiao, C.; Lu, J.; Zhang, Y. Fe(III)/Fe(II) forwarding a new anammox-like process to remove high-concentration ammonium using nitrate as terminal electron acceptor. Water Res. 2020, 172, 115528. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Jaffé, P.R. Characterization of incubation experiments and development of an enrichment culture capable of ammonium oxidation under iron-reducing conditions. Biogeosciences 2015, 12, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Tai, Y.L.; Dempsey, B.A. Nitrite reduction with hydrous ferric oxide and Fe(II): Stoichiometry, rate, and mechanism. Water Res. 2009, 43, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Lim, N.Y.N.; Frostegård, A.; Bakken, L.R. Nitrite kinetics during anoxia: The role of abiotic reactions versus microbial reduction. Soil Biol. Biochem. 2018, 119, 203–209. [Google Scholar] [CrossRef]
- Wimmer, F.L. Book Review: Inorganic Chemistry, 2nd ed.; Shriver, D.F., Atkins, P.W., Langford, C.H., Eds.; SAGE Journals: New York, NY, USA, 1995; pp. 599–600. [Google Scholar]
- Lu, Y.; Lu, X.; Shu, W.; Zhou, J.; Qian, G. Microbial mediated iron redox cycling in fe (hydr)oxides for nitrite removal. Bioresour. Technol. 2016, 224, 34–40. [Google Scholar] [CrossRef]
- Sultan, S.; Hasnain, S. Reduction of toxic hexavalent chromium by Ochrobactrum intermedium strain SDCr-5 stimulated by heavy metals. Bioresour. Technol. 2007, 98, 340–344. [Google Scholar] [CrossRef]
- Lei, X.; Jia, Y.; Chen, Y.; Hu, Y. Simultaneous nitrification and denitrification without nitrite accumulation by a novel isolated Ochrobactrum anthropic LJ81. Bioresour. Technol. 2019, 272, 442–450. [Google Scholar] [CrossRef]
- Zhou, J.; Sun, Q.; Chen, D.; Wang, H.Y.; Yang, K. Ochrobactrum anthropi used to control ammonium for nitrate removal by starch-stabilized nanoscale zero valent iron. Water Sci. Technol. 2017, 76, 1827–1832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doi, Y.; Takaya, N.; Takizawa, N. Novel Denitrifying Bacterium Ochrobactrum anthropi YD50.2 Tolerates High Levels of Reactive Nitrogen Oxides. Appl. Environ. Microb. 2009, 75, 5186–5194. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Park, S.; Yoon, Y.; Lee, J.H.; Wu, W.M.; Nguyen Phuoc, D.; Sadowsky, M.J.; Hur, H.G. Methanogenesis Facilitated by Geobiochemical Iron Cycle in a Novel Syntrophic Methanogenic Microbial Community. Environ. Sci. Technol. 2013, 47, 10078–10084. [Google Scholar] [CrossRef]
- Xu, Y.; He, Y.; Feng, X.; Liang, L.; Xu, J.; Brookes, P.C.; Wu, J. Enhanced abiotic and biotic contributions to dechlorination of pentachlorophenol during Fe(III) reduction by an iron-reducing bacterium Clostridium beijerinckii Z. Sci. Total Environ. 2014, 473, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Zhang, Y.; Quan, X.; Chen, S. Anaerobic biodecolorization of AO7 by a newly isolated Fe(III)-reducing bacterium Sphingomonas strain DJ. J. Chem. Technol. Biot. 2015, 90, 158–165. [Google Scholar] [CrossRef]
- Ding, B.; Luo, W.; Qin, Y.; Li, Z. Effects of the addition of nitrogen and phosphorus on anaerobic ammonium oxidation coupled with iron reduction (Feammox) in the farmland soils. Sci. Total Environ. 2020, 737, 139849. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Wang, B.; Zhang, L.; Wen, S.; Xu, M.; Misselbrook, T.H.; Carswell, A.M.; Gao, S. Striking a balance between N sources: Mitigating soil acidification and accumulation of phosphorous and heavy metals from manure. Sci. Total Environ. 2021, 754, 142189. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Carswell, A.; Zhu, Q.; Zhang, F.; de Vries, W. Modelling long-term impacts of fertilization and liming on soil acidification at Rothamsted experimental station. Sci. Total Environ. 2020, 713, 136249. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, D.; Wang, J.; Xue, J.; Yue, Z.; Xia, S.; Song, L.; Gao, H. Effects of Soil pH on Gaseous Nitrogen Loss Pathway via Feammox Process. Sustainability 2021, 13, 10393. https://doi.org/10.3390/su131810393
Ma D, Wang J, Xue J, Yue Z, Xia S, Song L, Gao H. Effects of Soil pH on Gaseous Nitrogen Loss Pathway via Feammox Process. Sustainability. 2021; 13(18):10393. https://doi.org/10.3390/su131810393
Chicago/Turabian StyleMa, Ding, Jin Wang, Jun Xue, Zhengbo Yue, Shaofeng Xia, Lei Song, and Hongwen Gao. 2021. "Effects of Soil pH on Gaseous Nitrogen Loss Pathway via Feammox Process" Sustainability 13, no. 18: 10393. https://doi.org/10.3390/su131810393