
Influence of Nutrient Manipulation on Growth and Biochemical Constituent in Anabaena variabilis and Nostoc muscorum to Enhance Biodiesel Production




Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Culture Conditions
2.2. Determination of Growth
2.3. Effect of Nutrients Manipulation
2.4. Determination of Total Lipid for the Isolated Species
2.5. Estimation of Fatty Acid Methyl Esters
2.6. Evaluation of Biodiesel Properties
2.7. Statistical Analysis
3. Results and Discussion
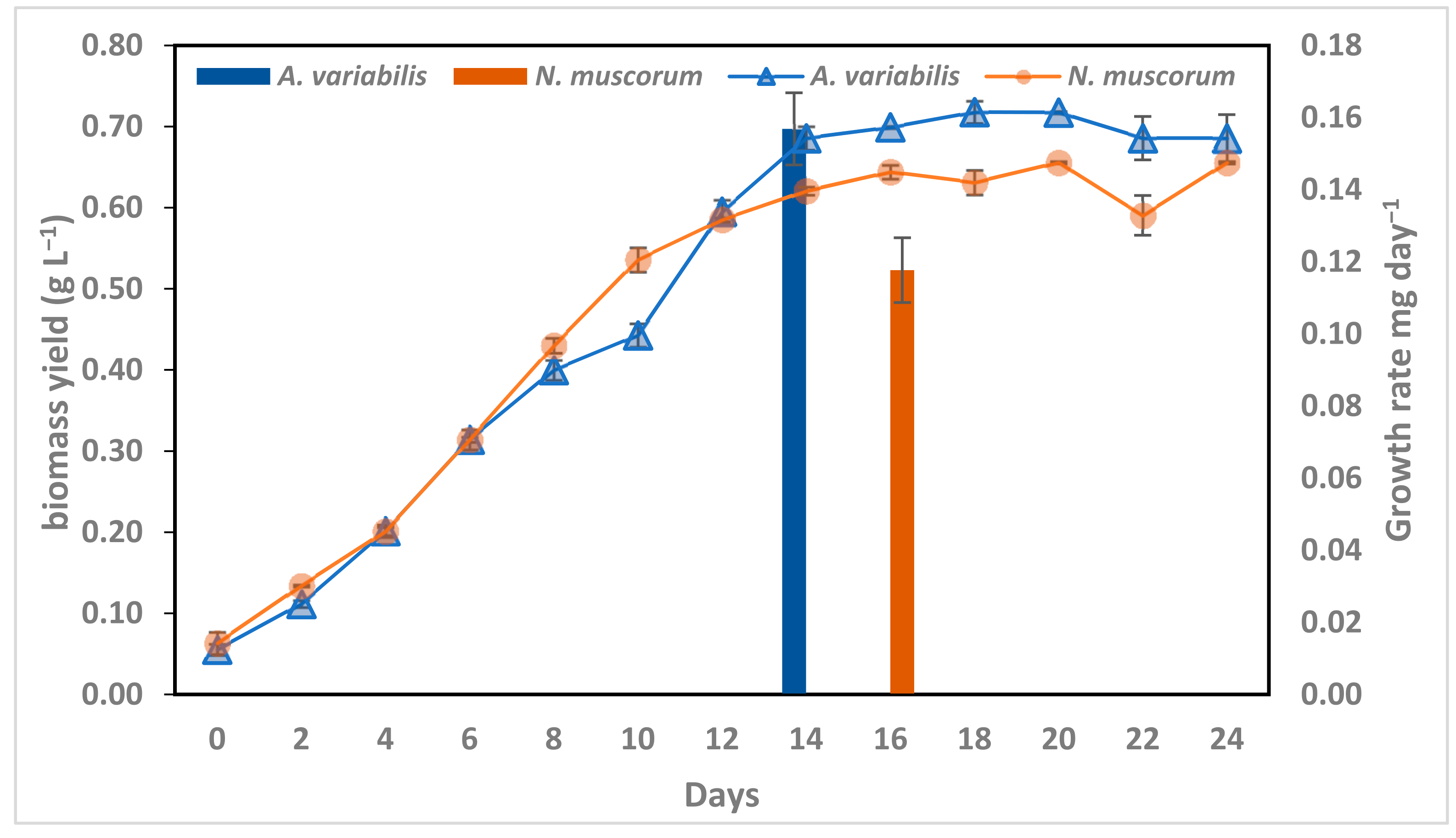
3.1. Growth Curve
3.2. Biomass and Biomass Productivities under Different Nutrient Concentrations
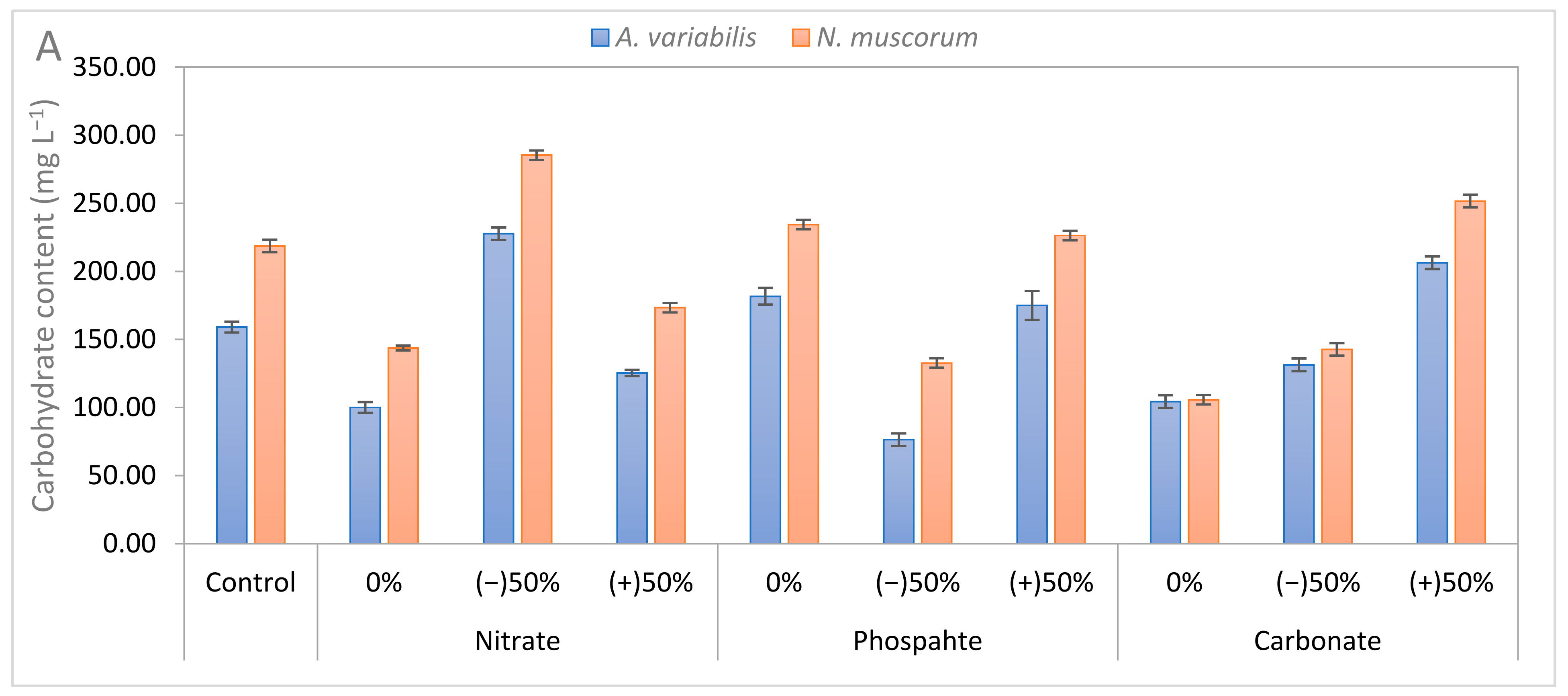
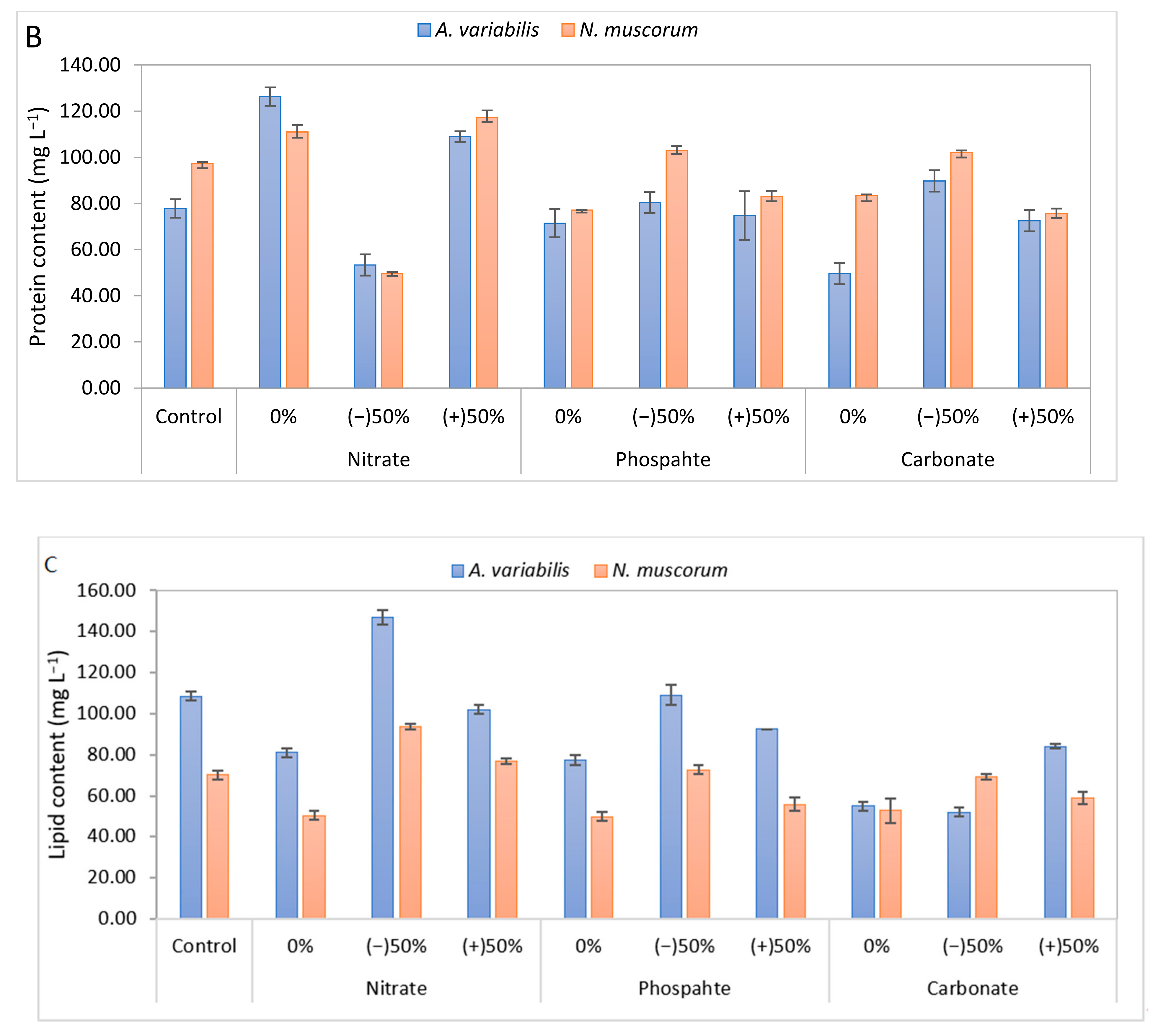
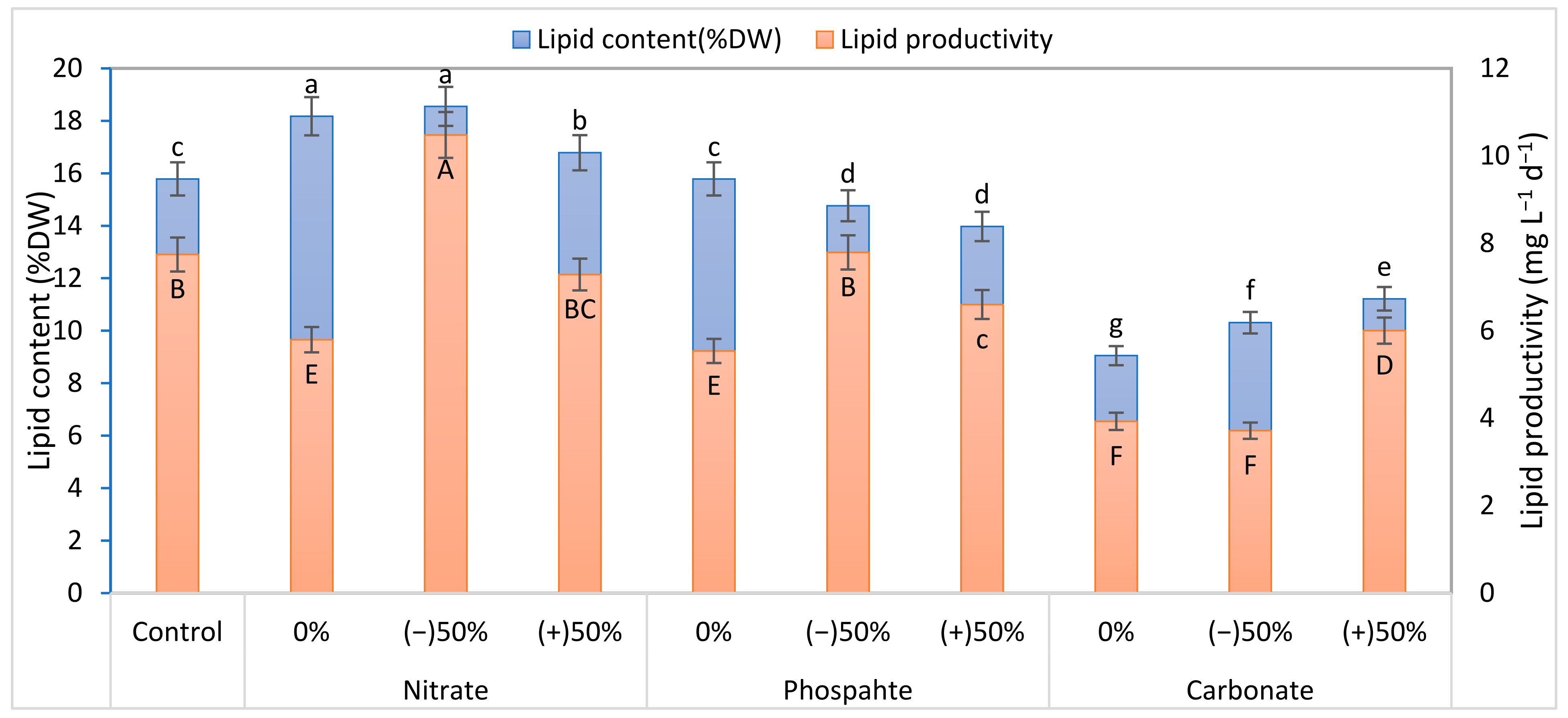
3.3. Biochemical Composition
3.4. Comparative Summary of the Individual Factors Analyses versus Control
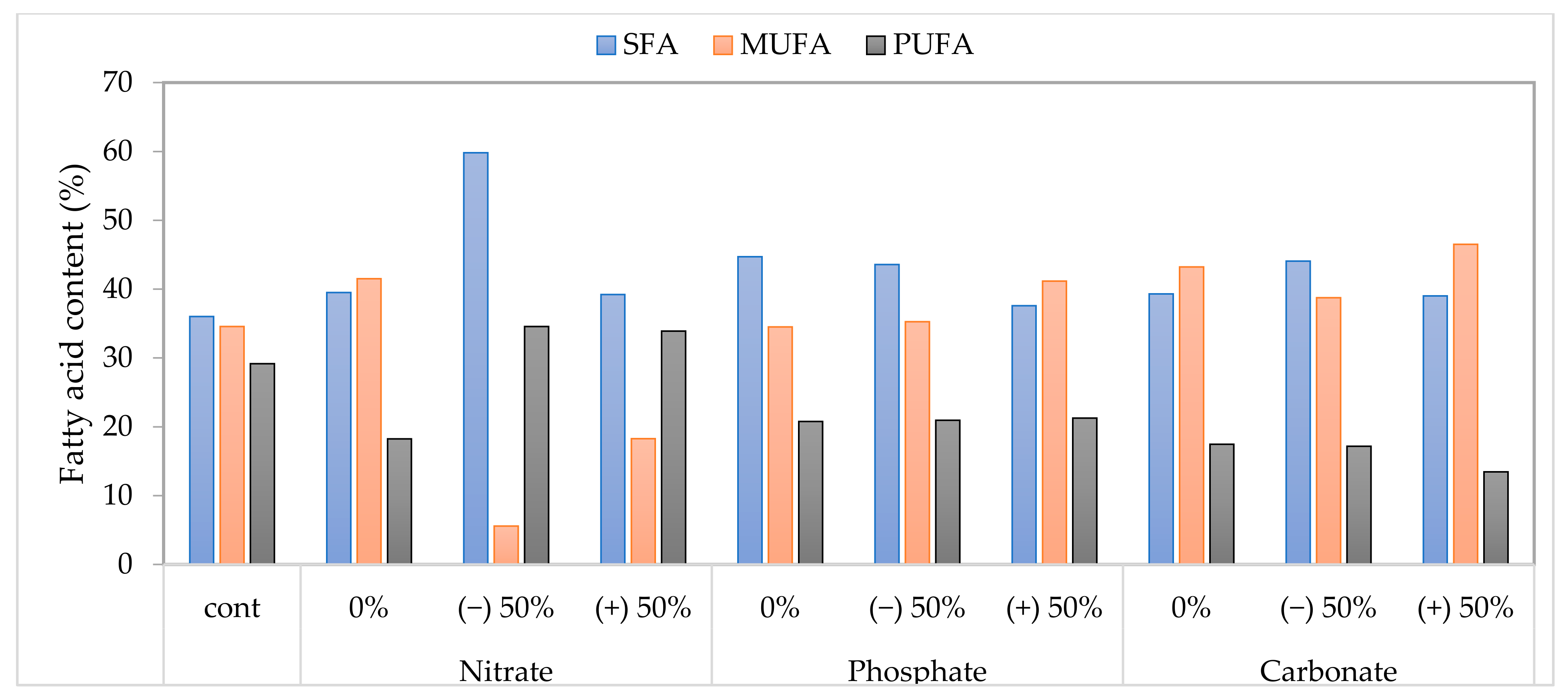
3.5. Fatty Acid Profiles Properties
3.6. Biodiesel Properties
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abdelsalam, I.M.; Elshobary, M.; Eladawy, M.M.; Nagah, M. Utilization of Multi-Tasking Non-Edible Plants for Phytoremediation and Bioenergy Source—A Review. Phyton 2019, 88, 69–90. [Google Scholar] [CrossRef]
- Elshobary, M.E.; Zabed, H.M.; Yun, J.; Zhang, G.; Qi, X. Recent insights into microalgae-assisted microbial fuel cells for generating sustainable bioelectricity. Int. J. Hydrogen Energy 2020, 46, 3135–3159. [Google Scholar] [CrossRef]
- Amaro, H.M.; Guedes, A.C.; Malcata, F.X. Advances and perspectives in using microalgae to produce biodiesel. Appl. Energy 2011, 88, 3402–3410. [Google Scholar] [CrossRef]
- Abomohra, A.-F.; Elshobary, M.E. Biodiesel, Bioethanol and Biobutanol Production from Microalgae Biomass. In Microalgae Biotechnology for Development of Biofuel and Waste Water Treatment; Springer: Singapore, 2019; pp. 293–321. [Google Scholar]
- Deb, D.; Mallick, N.; Bhadoria, P. Engineering culture medium for enhanced carbohydrate accumulation in Anabaena variabilis to stimulate production of bioethanol and other high-value co-products under cyanobacterial refinery approach. Renew. Energy 2020, 163, 1786–1801. [Google Scholar] [CrossRef]
- Yang, J.; Xu, M.; Zhang, X.; Hu, Q.; Sommerfeld, M.; Chen, Y. Life-cycle analysis on biodiesel production from microalgae: Water footprint and nutrients balance. Bioresour. Technol. 2011, 102, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Rittmann, B.E. Opportunities for renewable bioenergy using microorganisms. Biotechnol. Bioeng. 2008, 100, 203–212. [Google Scholar] [CrossRef]
- Rodriguez, H.; Guerrero, M.G. Products and Uses of Cyanobacteria (Bluegreen Algae). In Profiles on Biotechnology; Villa, T.G., Abalde, J., Eds.; Universidad de Santiago de Chile: Santiago, Chile, 1992; pp. 247–260. [Google Scholar]
- Modiri, S.; Hajfarajollah, H.; Zahiri, H.S.; Sharafi, H.; Noghabi, K.A.; Zamanzadeh, Z.; Haghighi, O.; Vali, H.; Alidoust, L.; Azarivand, A. Lipid production and mixotrophic growth features of cyanobacterial strains isolated from various aquatic sites. Microbiology 2015, 161, 662–673. [Google Scholar] [CrossRef] [Green Version]
- Karatay, S.E.; Dönmez, G. Microbial oil production from thermophile cyanobacteria for biodiesel production. Appl. Energy 2011, 88, 3632–3635. [Google Scholar] [CrossRef]
- de Oliveira, D.T.; Vasconcelos, C.T.; Feitosa, A.M.T.; Aboim, J.B.; Oliveira, A.D.N.D.; Xavier, L.; Santos, A.S.; Gonçalves, E.C.; Filho, G.N.D.R.; Nascimento, L.A.S.D. Lipid profile analysis of three new Amazonian cyanobacteria as potential sources of biodiesel. Fuel 2018, 234, 785–788. [Google Scholar] [CrossRef]
- Quintana, N.; Van Der Kooy, F.; Van De Rhee, M.D.; Voshol, G.P.; Verpoorte, R. Renewable energy from Cyanobacteria: Energy production optimization by metabolic pathway engineering. Appl. Microbiol. Biotechnol. 2011, 91, 471–490. [Google Scholar] [CrossRef] [Green Version]
- Ruffing, A.M. Improved Free Fatty Acid Production in Cyanobacteria with Synechococcus sp. PCC 7002 as Host. Front. Bioeng. Biotechnol. 2014, 2, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassilev, S.V.; Vassileva, C.G. Composition, properties and challenges of algae biomass for biofuel application: An overview. Fuel 2016, 181, 1–33. [Google Scholar] [CrossRef]
- Kim, H.; Jo, B.Y.; Kim, H.S. Effect of different concentrations and ratios of ammonium, nitrate, and phosphate on growth of the blue-green alga (cyanobacterium) Microcystis aeruginosa isolated from the Nakdong River, Korea. ALGAE 2017, 32, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Erratt, K.J.; Creed, I.F.; Trick, C.G. Comparative effects of ammonium, nitrate and urea on growth and photosynthetic efficiency of three bloom-forming cyanobacteria. Freshw. Biol. 2018, 63, 626–638. [Google Scholar] [CrossRef]
- Ruangsomboon, S. Effect of media and salinity on lipid content of cyanobacterium Hapalosiphon sp. Chiang Mai J. Sci. 2014, 41, 307–315. [Google Scholar]
- Nzayisenga, J.C.; Farge, X.; Groll, S.L.; Sellstedt, A. Effects of light intensity on growth and lipid production in microalgae grown in wastewater. Biotechnol. Biofuels 2020, 13, 1–8. [Google Scholar] [CrossRef]
- Cointet, E.; Wielgosz-Collin, G.; Bougaran, G.; Rabesaotra, V.; Gonçalves, O.; Méléder, V. Effects of light and nitrogen availability on photosynthetic efficiency and fatty acid content of three original benthic diatom strains. PLoS ONE 2019, 14, e0224701. [Google Scholar] [CrossRef]
- Ernst, A.; Deicher, M.; Herman, P.M.J.; Wollenzien, U.I.A. Nitrate and Phosphate Affect Cultivability of Cyanobacteria from Environments with Low Nutrient Levels. Appl. Environ. Microbiol. 2005, 71, 3379–3383. [Google Scholar] [CrossRef] [Green Version]
- Elshobary, M.E.; Abo-Shady, A.M.; Khairy, H.M.; Essa, D.; Zabed, H.M.; Qi, X.; Abomohra, A.E.-F. Influence of nutrient supplementation and starvation conditions on the biomass and lipid productivities of Micractinium reisseri grown in wastewater for biodiesel production. J. Environ. Manag. 2019, 250, 109529. [Google Scholar] [CrossRef]
- Srinuanpan, S.; Cheirsilp, B.; Prasertsan, P.; Kato, Y.; Asano, Y. Strategies to increase the potential use of oleaginous microalgae as biodiesel feedstocks: Nutrient starvations and cost-effective harvesting process. Renew. Energy 2018, 122, 507–516. [Google Scholar] [CrossRef]
- Kamalanathan, M.; Pierangelini, M.; Shearman, L.A.; Gleadow, R.; Beardall, J. Impacts of nitrogen and phosphorus starvation on the physiology of Chlamydomonas reinhardtii. Environ. Boil. Fishes 2015, 28, 1509–1520. [Google Scholar] [CrossRef]
- Khatoon, N.; Pal, R. Microalgae in biotechnological application: A commercial approach. In Plant Biology and Biotechnology; Springer: Berlin/Heidelberg, Germany, 2015; pp. 27–47. [Google Scholar]
- Allen, M.M.E.; Stanier, R.Y. Selective Isolation of Blue-green Algae from Water and Soil. J. Gen. Microbiol. 1968, 51, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Aboshady, A.; Khairy, H.; Abomohra, A.; Elshobary, M.; Essa, D. Influence of algal biotreated wastewater on some growth parameters and metabolites of Vicia faba. Egypt. J. Exp. Boil. (Botany) 2017, 13, 209–217. [Google Scholar] [CrossRef]
- Rogers, S.L.; Burns, R.G. Changes in aggregate stability, nutrient status, indigenous microbial populations, and seedling emergence, following inoculation of soil with Nostoc muscorum. Biol. Fertil. Soils 1994, 18, 209–215. [Google Scholar] [CrossRef]
- Abo-Shady, A.; Khairy, H.M.; Abomohra, A.; Elshobary, M. Potential cultivation of halophilic oleaginous microalgae on industrial wastewater. Egypt. J. Bot. 2018, 58, 205–206. [Google Scholar] [CrossRef] [Green Version]
- Payne, J.K.; Stewart, J.R. The chemical composition of the thallus wall of Characiosiphon rivularis (Characiosiphonaceae, Chlorophyta). Phycologia 1988, 27, 43–49. [Google Scholar] [CrossRef]
- Elshobary, M.E.; Osman, M.E.H.; Abushady, A.M.; Piercey-Normore, M.D. Comparison of Lichen-Forming Cyanobacterial and Green Algal Photobionts with Free-Living Algae. Cryptogam. Algologie 2015, 36, 81–100. [Google Scholar] [CrossRef]
- Elshobary, M.E.; Osman, M.E.; Abo-Shady, A.M.; Komatsu, E.; Perreault, H.; Sorensen, J.; Piercey-Normore, M.D. Algal carbohydrates affect polyketide synthesis of the lichen-forming fungus Cladonia rangiferina. Mycologia 2016, 108, 646–656. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Improved recovery of fatty acid through direct transesterification without prior extraction or purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef]
- Krzemińska, I.; Oleszek, M. Glucose supplementation-induced changes in the Auxenochlorella protothecoides fatty acid composition suitable for biodiesel production. Bioresour. Technol. 2016, 218, 1294–1297. [Google Scholar] [CrossRef]
- Hoekman, S.K.; Broch, A.; Robbins, C.; Ceniceros, E.; Natarajan, M. Review of biodiesel composition, properties, and specifications. Renew. Sustain. Energy Rev. 2012, 16, 143–169. [Google Scholar] [CrossRef]
- Ashour, M.; Elshobary, M.E.; El-Shenody, R.; Kamil, A.e.W.; Abomohra, A.E.-F. Evaluation of a native oleaginous marine microalga Nannochloropsis oceanica for dual use in biodiesel production and aquaculture feed. Biomass Bioenergy 2018, 120, 439–447. [Google Scholar] [CrossRef]
- Singh, P.; Kumar, D. Biomass and Lipid Productivities of Cyanobacteria-Leptolyngbya foveolarum HNBGU001. BioEnergy Res. 2020, 14, 278–291. [Google Scholar] [CrossRef]
- Beale, S.I. Enzymes of chlorophyll biosynthesis. Photosynth. Res. 1999, 60, 43–73. [Google Scholar] [CrossRef]
- Kim, H.; Oh, C.; Bae, H. Comparison of red microalgae (Porphyridium cruentum) culture conditions for bioethanol production. Bioresour. Technol. 2017, 233, 44–50. [Google Scholar] [CrossRef]
- Sarkar, A.; Rajarathinam, R.; Venkateshan, R.B. A comparative assessment of growth, pigment and enhanced lipid production by two toxic freshwater cyanobacteria Anabaena circinalis FSS 124 and Cylindrospermopsis raciborskii FSS 127 under various combinations of nitrogen and phosphorous inputs. Environ. Sci. Pollut. Res. 2020, 28, 15923–15933. [Google Scholar] [CrossRef]
- Herrero, A.; Muro-Pastor, A.M.; Flores, E. Nitrogen Control in Cyanobacteria. J. Bacteriol. 2001, 183, 411–425. [Google Scholar] [CrossRef] [Green Version]
- Xie, T.; Xia, Y.; Zeng, Y.; Li, X.; Zhang, Y. Nitrate concentration-shift cultivation to enhance protein content of heterotrophic microalga Chlorella vulgaris: Over-compensation strategy. Bioresour. Technol. 2017, 233, 247–255. [Google Scholar] [CrossRef]
- Markou, G.; Vandamme, D.; Muylaert, K. Microalgal and cyanobacterial cultivation: The supply of nutrients. Water Res. 2014, 65, 186–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markou, G.; Angelidaki, I.; Georgakakis, D. Microalgal carbohydrates: An overview of the factors influencing carbohydrates production, and of main bioconversion technologies for production of biofuels. Appl. Microbiol. Biotechnol. 2012, 96, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, H.; Chu, H.; Yu, S. A batch study on the bio-fixation of carbon dioxide in the absorbed solution from a chemical wet scrubber by hot spring and marine algae. Chemosphere 2007, 66, 878–886. [Google Scholar] [CrossRef]
- de Morais, M.G.; Costa, J.A.V. Carbon dioxide fixation by Chlorella kessleri, C. vulgaris, Scenedesmus obliquus and Spirulina sp. cultivated in flasks and vertical tubular photobioreactors. Biotechnol. Lett. 2007, 29, 1349–1352. [Google Scholar] [CrossRef] [PubMed]
- Giordano, M.; Beardall, J.; Raven, J.A. CO2 concentrating mechanisms in algae: Mechanisms, Environmental Modulation, and Evolution. Annu. Rev. Plant Biol. 2005, 56, 99–131. [Google Scholar] [CrossRef] [Green Version]
- Deb, D.; Mallick, N.; Bhadoria, P. Analytical studies on carbohydrates of two cyanobacterial species for enhanced bioethanol production along with poly-β-hydroxybutyrate, C-phycocyanin, sodium copper chlorophyllin, and exopolysaccharides as co-products. J. Clean. Prod. 2019, 221, 695–709. [Google Scholar] [CrossRef]
- Yang, S.; Xu, J.; Wang, Z.-M.; Bao, L.-J.; Zeng, E.Y. Cultivation of oleaginous microalgae for removal of nutrients and heavy metals from biogas digestates. J. Clean. Prod. 2017, 164, 793–803. [Google Scholar] [CrossRef]
- Subramanian, S.; Barry, A.N.; Pieris, S.; Sayre, R.T. Comparative energetics and kinetics of autotrophic lipid and starch metabolism in chlorophytic microalgae: Implications for biomass and biofuel production. Biotechnol. Biofuels 2013, 6, 150. [Google Scholar] [CrossRef] [Green Version]
- Price, G.D.; Badger, M.; Woodger, F.J.; Long, B. Advances in understanding the cyanobacterial CO2-concentrating-mechanism (CCM): Functional components, Ci transporters, diversity, genetic regulation and prospects for engineering into plants. J. Exp. Bot. 2007, 59, 1441–1461. [Google Scholar] [CrossRef] [PubMed]
- Pancha, I.; Chokshi, K.; George, B.; Ghosh, T.; Paliwal, C.; Maurya, R.; Mishra, S. Nitrogen stress triggered biochemical and morphological changes in the microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2014, 156, 146–154. [Google Scholar] [CrossRef]
- Zarrinmehr, M.J.; Farhadian, O.; Heyrati, F.P.; Keramat, J.; Koutra, E.; Kornaros, M.; Daneshvar, E. Effect of nitrogen concentration on the growth rate and biochemical composition of the microalga, Isochrysis galbana. Egypt. J. Aquat. Res. 2019, 46, 153–158. [Google Scholar] [CrossRef]
- An, M.; Gao, L.; Zhao, W.; Chen, W.; Li, M. Effects of Nitrogen Forms and Supply Mode on Lipid Production of Microalga Scenedesmus obliquus. Energies 2020, 13, 697. [Google Scholar] [CrossRef] [Green Version]
- Yeesang, C.; Cheirsilp, B. Effect of nitrogen, salt, and iron content in the growth medium and light intensity on lipid production by microalgae isolated from freshwater sources in Thailand. Bioresour. Technol. 2011, 102, 3034–3040. [Google Scholar] [CrossRef]
- Ahlgren, G.; Hyenstrand, P. Nitrogen limitation effects of different nitrogen sources on nutritional quality of two freshwater organisms, Scenedesmus quadricauda (Chlorophyceae) and Synechococcus sp. (Cyanophyceae). J. Phycol. 2003, 39, 906–917. [Google Scholar] [CrossRef]
- Banerjee, A.; Sharma, R.; Chisti, Y.; Banerjee, U.C. Botryococcus braunii: A Renewable Source of Hydrocarbons and Other Chemicals. Crit. Rev. Biotechnol. 2002, 22, 245–279. [Google Scholar] [CrossRef]
- Dayananda, C.; Sarada, R.; Bhattacharya, S.; Ravishankar, G.A. Effect of media and culture conditions on growth and hydrocarbon production by Botryococcus braunii. Process Biochem. 2005, 40, 3125–3131. [Google Scholar] [CrossRef]
- Zaki, M.; Ashour, M.; Heneash, A.; Mabrouk, M.; Alprol, A.; Khairy, H.; Nour, A.; Mansour, A.; Hassanien, H.; Gaber, A.; et al. Potential Applications of Native Cyanobacterium Isolate (Arthrospira platensis NIOF17/003) for Biodiesel Production and Utilization of Its Byproduct in Marine Rotifer (Brachionus plicatilis) Production. Sustainability 2021, 13, 1769. [Google Scholar] [CrossRef]
- Huo, S.; Basheer, S.; Liu, F.; Elshobary, M.; Zhang, C.; Qian, J.; Xu, L.; Arslan, M.; Cui, F.; Zan, X.; et al. Bacterial intervention on the growth, nutrient removal and lipid production of filamentous oleaginous microalgae Tribonema sp. Algal Res. 2020, 52, 102088. [Google Scholar] [CrossRef]
- Anahas, A.M.P.; Muralitharan, G. Characterization of heterocystous cyanobacterial strains for biodiesel production based on fatty acid content analysis and hydrocarbon production. Energy Convers. Manag. 2018, 157, 423–437. [Google Scholar] [CrossRef]
- Paliwal, C.; Mitra, M.; Bhayani, K.; Bharadwaj, S.V.; Ghosh, T.; Dubey, S.; Mishra, S. Abiotic stresses as tools for metabolites in microalgae. Bioresour. Technol. 2017, 244, 1216–1226. [Google Scholar] [CrossRef] [PubMed]
- Yodsuwan, N.; Sawayama, S.; Sirisansaneeyakul, S. Effect of nitrogen concentration on growth, lipid production and fatty acid profiles of the marine diatom Phaeodactylum tricornutum. Agric. Nat. Resour. 2017, 51, 190–197. [Google Scholar] [CrossRef]
- Knothe, G.; Dunn, R. MO Bagby Biodiesel: The Use of vegetable Oils and Their Derivatives as Alternative Diesel Fuels. In Fuels and Chemicals from Biomass; Saha, B.C., Woodward, J., Eds.; ACS Publications: Washington, DC, USA, 1997. [Google Scholar]
- Haberle, I.; Hrustić, E.; Petrić, I.; Pritišanac, E.; Šilović, T.; Magić, L.; Geček, S.; Budiša, A.; Blažina, M. Adriatic cyanobacteria potential for cogeneration biofuel production with oil refinery wastewater remediation. Algal Res. 2020, 50, 101978. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.; Caetano, N. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Graboski, M.S.; McCormick, R. Combustion of fat and vegetable oil derived fuels in diesel engines. Prog. Energy Combust. Sci. 1998, 24, 125–164. [Google Scholar] [CrossRef]
- Ganesan, R.; Manigandan, S.; Samuel, M.S.; Shanmuganathan, R.; Brindhadevi, K.; Chi, N.T.L.; Duc, P.A.; Pugazhendhi, A. A review on prospective production of biofuel from microalgae. Biotechnol. Rep. 2020, 27, e00509. [Google Scholar] [CrossRef]
- Sanchez, A.; Maceiras, R.; Cancela-Carral, A.; Pérez, A. Culture aspects of Isochrysis galbana for biodiesel production. Appl. Energy 2013, 101, 192–197. [Google Scholar] [CrossRef]
- Anahas, A.M.P.; Muralitharan, G. Isolation and screening of heterocystous cyanobacterial strains for biodiesel production by evaluating the fuel properties from fatty acid methyl ester (FAME) profiles. Bioresour. Technol. 2015, 184, 9–17. [Google Scholar] [CrossRef]
- Ismail, M.M.; Ismail, G.A.; El-Sheekh, M.M. Potential assessment of some micro and macroalgal species for bioethanol and biodiesel production. Energy Sources Part A Recover. Util. Environ. Eff. 2020, 1–17. [Google Scholar] [CrossRef]
- Karpagam, R.; Preeti, R.; Ashokkumar, B.; Varalakshmi, P. Enhancement of lipid production and fatty acid profiling in Chlamydomonas reinhardtii, CC1010 for biodiesel production. Ecotoxicol. Environ. Saf. 2015, 121, 253–257. [Google Scholar] [CrossRef]
- Sharma, A.K.; Sahoo, P.K.; Singhal, S.; Joshi, G. Exploration of upstream and downstream process for microwave assisted sustainable biodiesel production from microalgae Chlorella vulgaris. Bioresour. Technol. 2016, 216, 793–800. [Google Scholar] [CrossRef]
- Ashokkumar, V.; Salam, Z.; Tiwari, O.; Chinnasamy, S.; Mohammed, S.; Ani, F.N. An integrated approach for biodiesel and bioethanol production from Scenedesmus bijugatus cultivated in a vertical tubular photobioreactor. Energy Convers. Manag. 2015, 101, 778–786. [Google Scholar] [CrossRef]
- Mittelbach, M.; Remschmidt, C. Biodiesel: The Comprehensive Handbook; Martin Mittelbach: Gratz, Austria, 2004. [Google Scholar]
- Zhang, L.; Cheng, J.; Pei, H.; Pan, J.; Jiang, L.; Hou, Q.; Han, F. Cultivation of microalgae using anaerobically digested effluent from kitchen waste as a nutrient source for biodiesel production. Renew. Energy 2017, 115, 276–287. [Google Scholar] [CrossRef]
- Osman, M.E.H.; Abo-Shady, A.M.; Elshobary, M.E.; El-Ghafar, M.O.A.; Abomohra, A.E.-F. Screening of seaweeds for sustainable biofuel recovery through sequential biodiesel and bioethanol production. Environ. Sci. Pollut. Res. 2020, 27, 32481–32493. [Google Scholar] [CrossRef] [PubMed]
- Elshobary, M.E.; El-Shenody, R.A.; Abomohra, A.E. Sequential biofuel production from seaweeds enhances the energy recovery: A case study for biodiesel and bioethanol production. Int. J. Energy Res. 2020, 45, 6457–6467. [Google Scholar] [CrossRef]
- ASTM D6751. ASTM International Standard Specification for Biodiesel Fuel Blend Stock (B100) for Middle Distillate Fuels, ASTM D6751-08; ASTM International: West Conshohocken, PA, USA, 2008. [Google Scholar]
- EN 14214 Fatty Acid Methyl Esters [FAME] for Diesel Engines. In Requirements and Test Methods; European Committee for Standardization: Brussels, Belgium, 2008.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrients | Zero% | −50% | Control | +50% |
|---|---|---|---|---|
| NaNO3 | 0 | 750 | 1500 | 2250 |
| K2HPO4·3H2O | 0 | 19.5 | 39 | 58.5 |
| Na2CO3 | 0 | 10 | 20 | 30 |
| Nutrients | Treatments | A. variabilis | N. muscorum | ||
|---|---|---|---|---|---|
| CDW (g L−1) | BP (mg g−1 d−1) | CDW (gL−1) | BP (mg g−1 d−1) | ||
| Control | 0.69 ± 0.026 c | 0.031 ± 0.003 c | 0.64 ± 0.006 c | 0.029 ± 0.001 c | |
| Nitrate | 0% | 0.31 ± 0.016 f | 0.010 ± 0.001 f | 0.30 ± 0.002 g | 0.008 ± 0.002 h |
| (−)50% | 0.56 ± 0.007 d | 0.025 ± 0.003 de | 0.56 ± 0.003 de | 0.021 ± 0.003 e | |
| (+)50% | 0.79 ± 0.026 a | 0.042 ± 0.004 a | 0.74 ± 0.002 a | 0.034 ± 0.002 a | |
| Phosphate | 0% | 0.55 ± 0.003 de | 0.025 ± 0.003 de | 0.53 ± 0.003 ef | 0.019 ± 0.002 f |
| (−)50% | 0.56 ± 0.011 d | 0.026 ± 0.003 d | 0.55 ± 0.002 | 0.021 ± 0.003 e | |
| (+)50% | 0.74 ± 0.023 b | 0.035 ± 0.003 b | 0.68 ± 0.005 b | 0.032 ± 0.001 b | |
| Carbonate | 0% | 0.51 ± 0.012 e | 0.022 ± 0.002 e | 0.51 ± 0.001 f | 0.017 ± 0.002 g |
| (−)50% | 0.57 ± 0.042 d | 0.027 ± 0.003 d | 0.58 ± 0.002 d | 0.023 ± 0.003 de | |
| (+)50% | 0.75 ± 0.013 b | 0.039 ± 0.004 a | 0.66 ± 0.004 b | 0.032 ± 0.000 b | |
| F-value | 468.564 ** | 181.82 ** | 671.088 * | 458.538 * | |
| Fatty Acids | Types | Ctrl | Nitrate | Phosphate | Carbonate | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0% | (−)50% | (+)50% | 0% | (−)50% | (+)50% | 0% | (−)50% | (+)50% | |||
| Lauric acid | C12:0 | - | - | - | - | - | 6.2 | - | - | - | - |
| Tridecanoic acid | C13:0 | - | - | - | 4.08 | 5.64 | - | - | - | - | - |
| Myristic acid | C14:0 | 0.92 | 8.55 | 14.48 | - | - | 3.5 | - | - | - | - |
| Pentadecanoic acid | C15:0 | 4.42 | 5.8 | - | - | - | 7.5 | - | 4.23 | - | |
| Palmitic acid | C16:0 | 30.4 | 15.77 | 24.64 | 28.53 | 30.8 | 22.5 | 20.14 | 26.13 | 12.66 | 21.73 |
| Stearic acid | C18:0 | 4.3 | 3.93 | 4.55 | 4.11 | 5.53 | 2.88 | 8.15 | 12.32 | 14.57 | 11.6 |
| Archidic acid | C20:0 | 0.4 | 6.85 | 10.34 | 2.5 | 2.73 | 2.5 | 7.8 | 0.85 | 12.6 | 5.7 |
| Palmitoleic acid | C16:1 | 1.3 | 2 | - | 1.78 | 6.4 | 4.75 | 8.86 | 1.78 | 9.01 | 1.45 |
| Heptadecanoic acid | C17:1 | 5.21 | 8.06 | - | - | - | 2.08 | - | - | - | - |
| Oleic acid | C18:1 | 27.8 | 28.22 | 5.58 | 13.37 | 27.7 | 32.3 | 22.55 | 40.84 | 14.5 | 45.047 |
| Erucic acid | C22:1 | 0.25 | 3.23 | - | 3.13 | 0.4 | 2.03 | 3.85 | 0.61 | 15.25 | - |
| Linoleic acid | C18:2 | 27.7 | 13.1 | 33.5 | 28.54 | 16.3 | 18.1 | 15.5 | 16.02 | 17.18 | 12.53 |
| Lenolenic acid | C18:3 | 1.48 | 5.15 | 1.07 | 5.36 | 4.46 | 3.16 | 5.44 | 1.44 | - | 0.91 |
| Biodiesel Parameters | ADU | KV (mm2 s−1) | CN | IV (g I2100 g−1 Oil) | Cp (℃) | ρ (g m−3) | HHV (MJ kg−1) | Reference |
|---|---|---|---|---|---|---|---|---|
| Control | 0.94 | 4.61 | 56.58 | 82.92 | 7.39 | 0.88 | 40.20 | This study |
| +50% NO3 | 0.76 | 4.73 | 57.82 | 69.08 | 9.87 | 0.88 | 39.87 | This study |
| ASTM D6751 | - | 1.9–6.0 | ≥47 | - | - | 0.85–0.90 | - | [80] |
| EN 14214 | - | 3.5–5 | ≥51 | ≤120 | >4 | 0.86–0.90 | - | [81] |
| A. variabilis MBDU 013 | 1.56 | 69.09 | 30.93 | 9.15 | 0.87 | [62] | ||
| Arthrospira platensis NIOF17/003 | 19.3 | 52.9 | 85.5 | - | 40.71 | [61] | ||
| Micractinium reisseri | 0.33 | 5 | 60.65 | 37.55 | 15.53 | 0.87 | 39.19 | [21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Shafay, S.M.; Gaber, A.; Alsanie, W.F.; Elshobary, M.E. Influence of Nutrient Manipulation on Growth and Biochemical Constituent in Anabaena variabilis and Nostoc muscorum to Enhance Biodiesel Production. Sustainability 2021, 13, 9081. https://doi.org/10.3390/su13169081
El Shafay SM, Gaber A, Alsanie WF, Elshobary ME. Influence of Nutrient Manipulation on Growth and Biochemical Constituent in Anabaena variabilis and Nostoc muscorum to Enhance Biodiesel Production. Sustainability. 2021; 13(16):9081. https://doi.org/10.3390/su13169081
Chicago/Turabian StyleEl Shafay, Shimaa M., Ahmed Gaber, Walaa F. Alsanie, and Mostafa E. Elshobary. 2021. "Influence of Nutrient Manipulation on Growth and Biochemical Constituent in Anabaena variabilis and Nostoc muscorum to Enhance Biodiesel Production" Sustainability 13, no. 16: 9081. https://doi.org/10.3390/su13169081