
Can Habitat Quality Index Measured Using the InVEST Model Explain Variations in Bird Diversity in an Urban Area?

Abstract
:1. Introduction
2. Materials and Methods
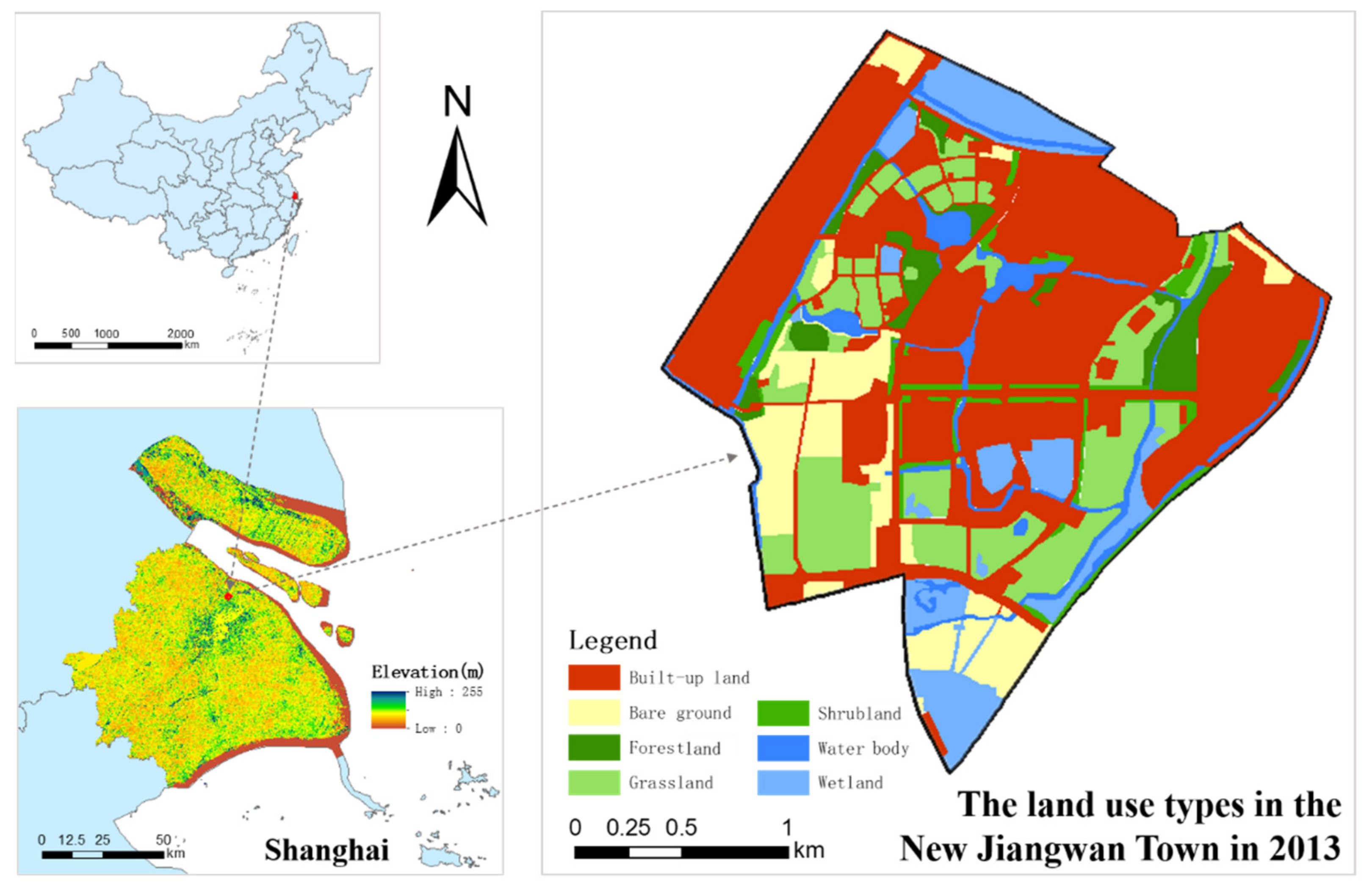
2.1. Study Area
2.2. Data Collection
2.3. Methods
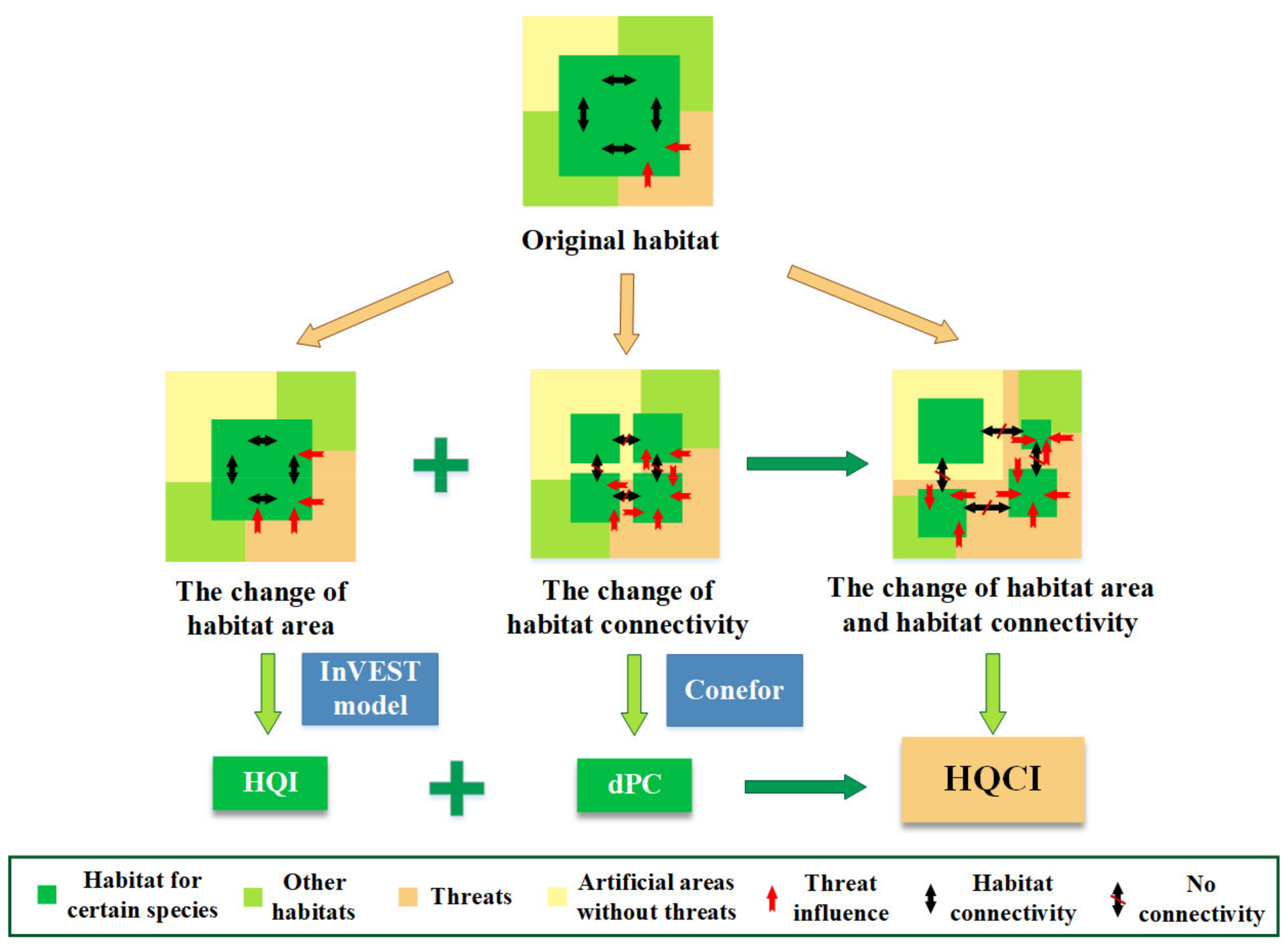
2.3.1. The HQI Calculated Using the InVEST Model
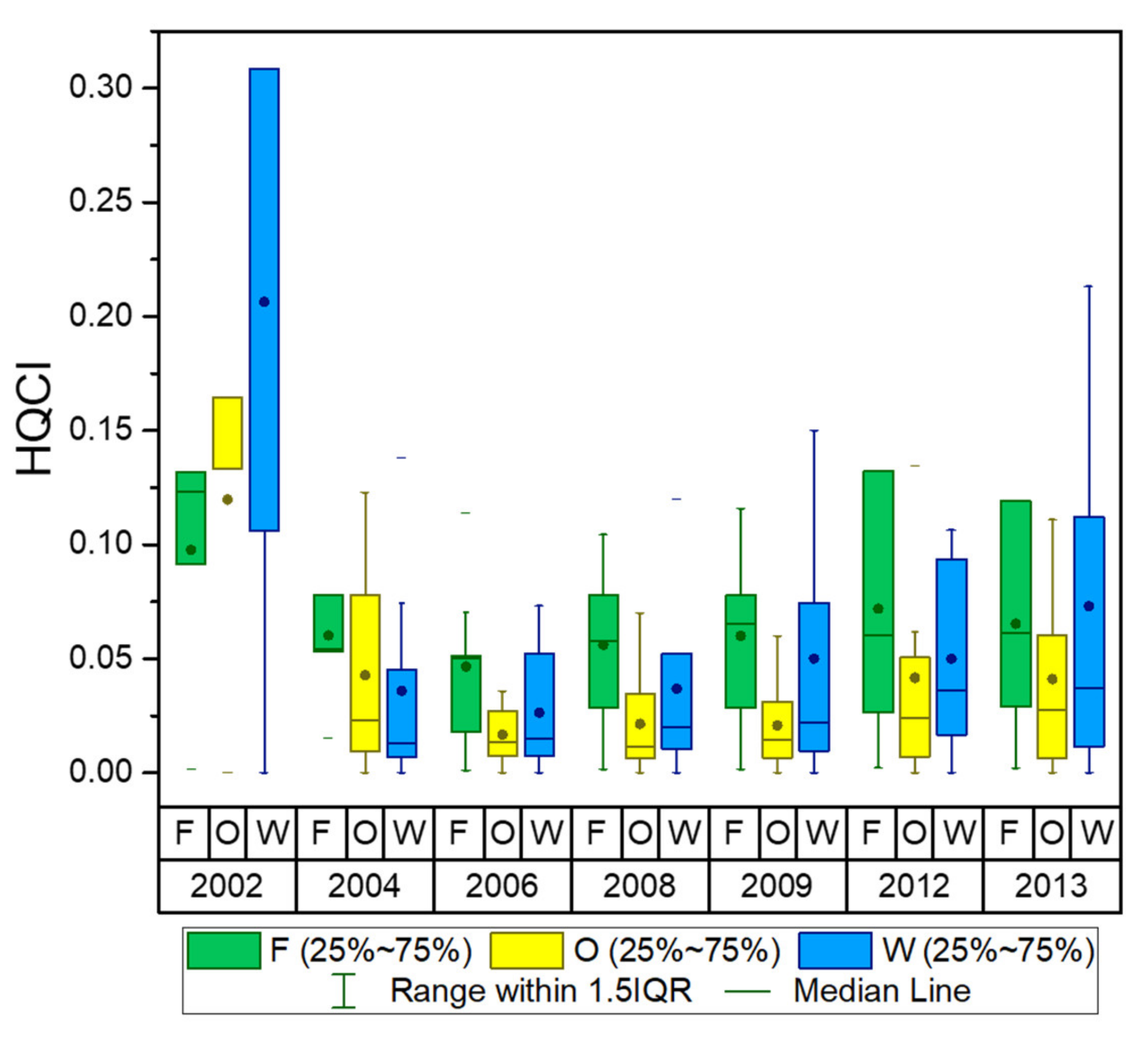
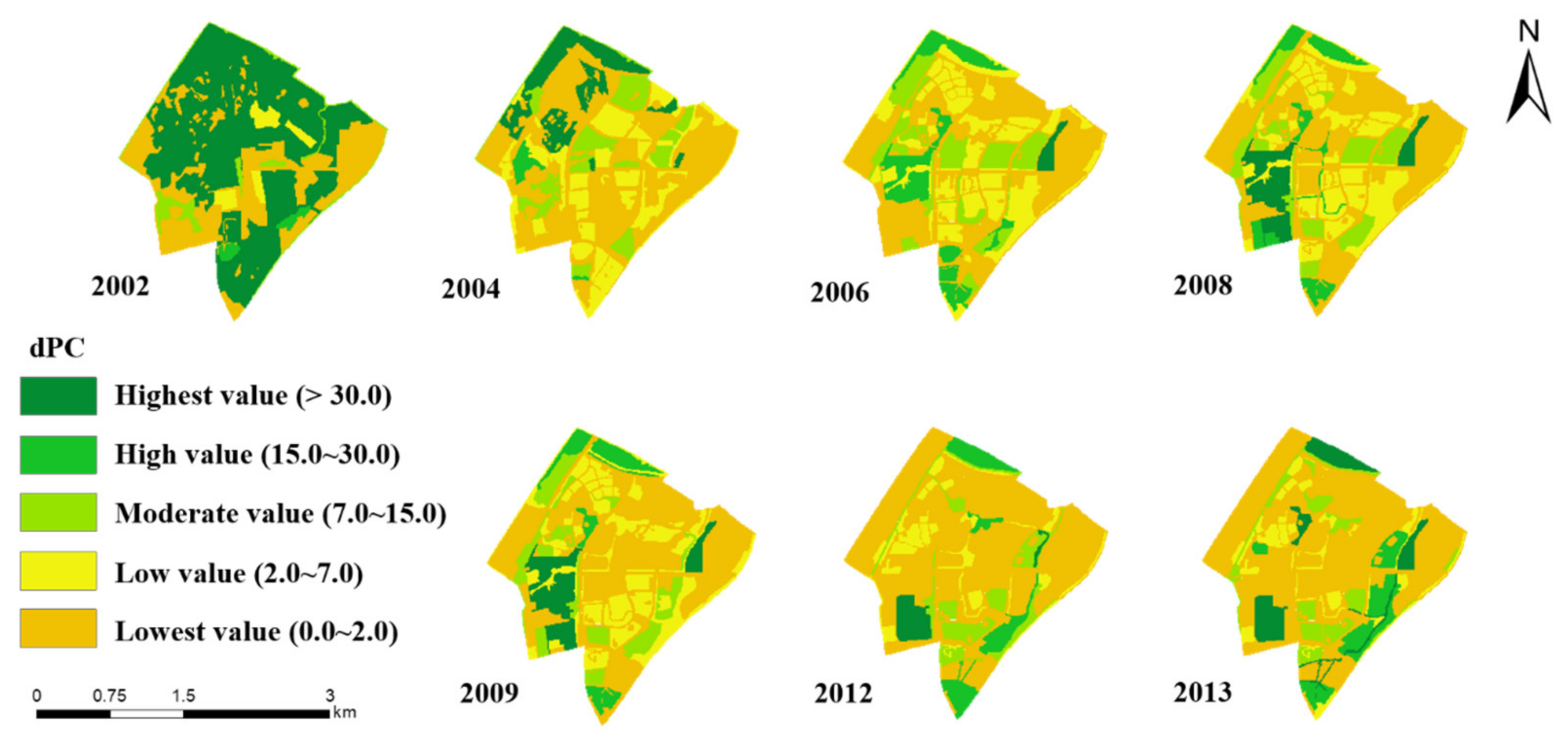
2.3.2. Habitat Connectivity Measured by the Graph-Based Connectivity Index, dPC
2.3.3. The New Compound Indicator Considering Both Habitat Quality and Connectivity
2.3.4. The Calculation of Bird Diversity
2.3.5. The Statistical Methods
3. Results
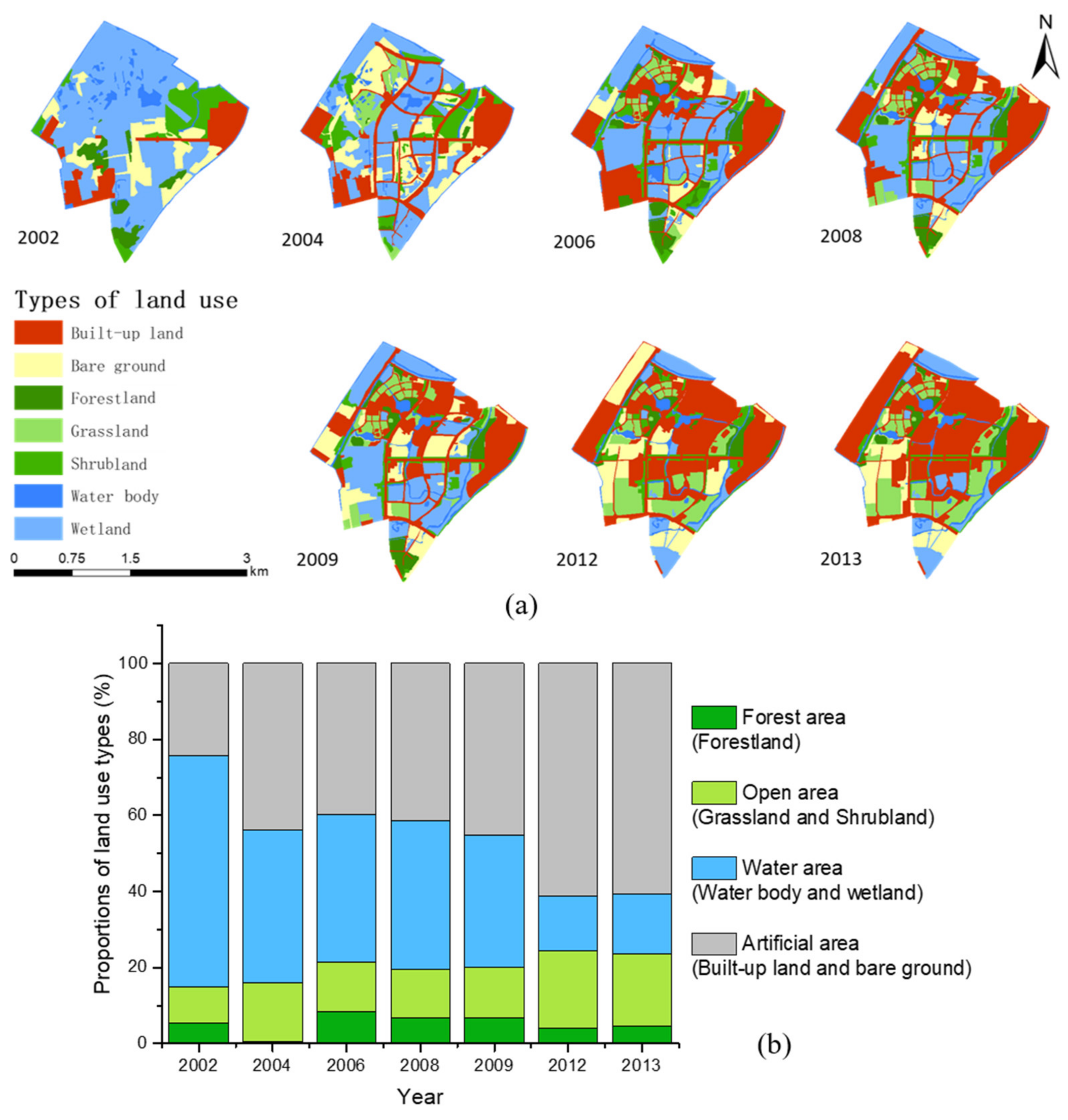
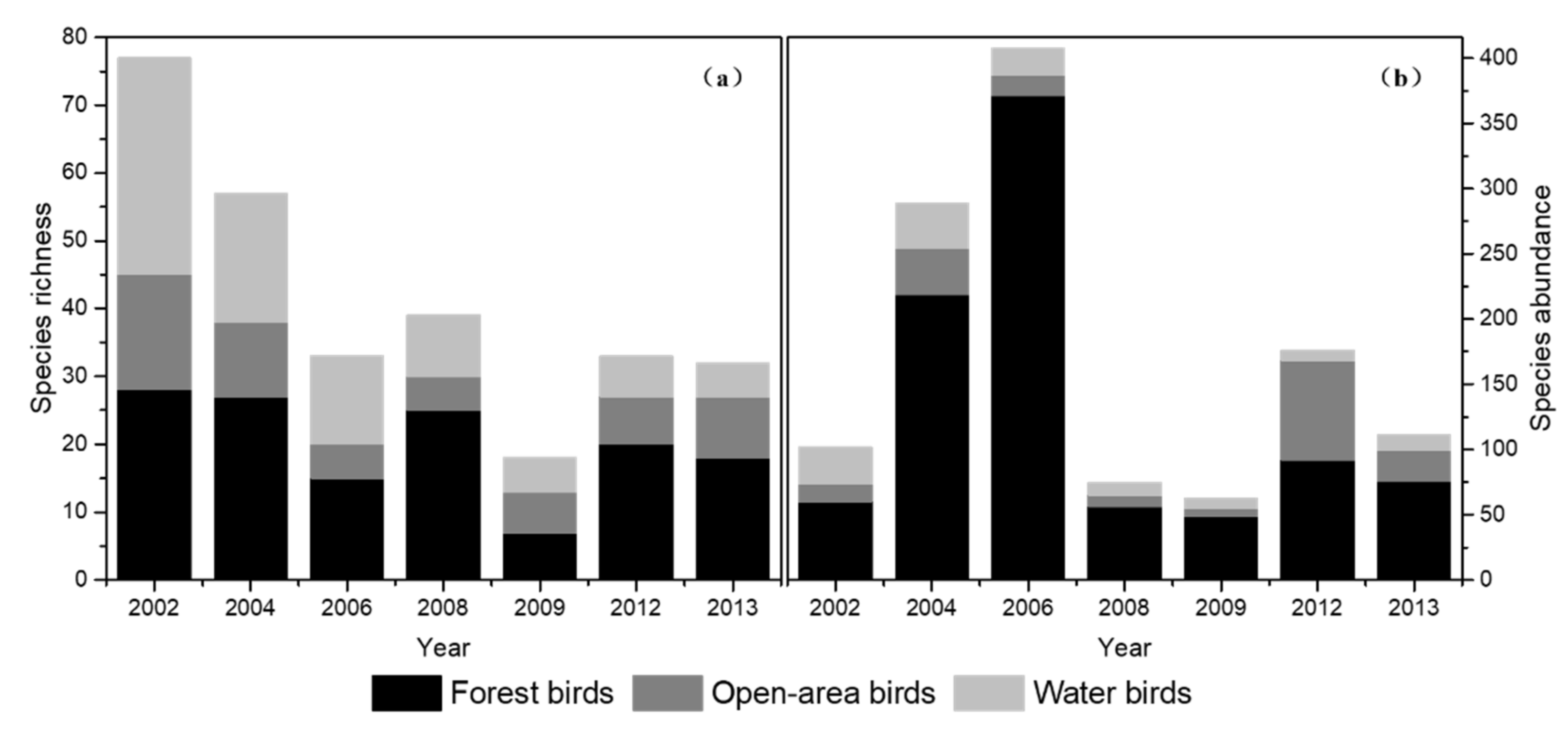
3.1. Variations of Land Use Types and Bird Diversity
- (1)
- The variations of the land use types
- (2)
- The variations of the bird diversity
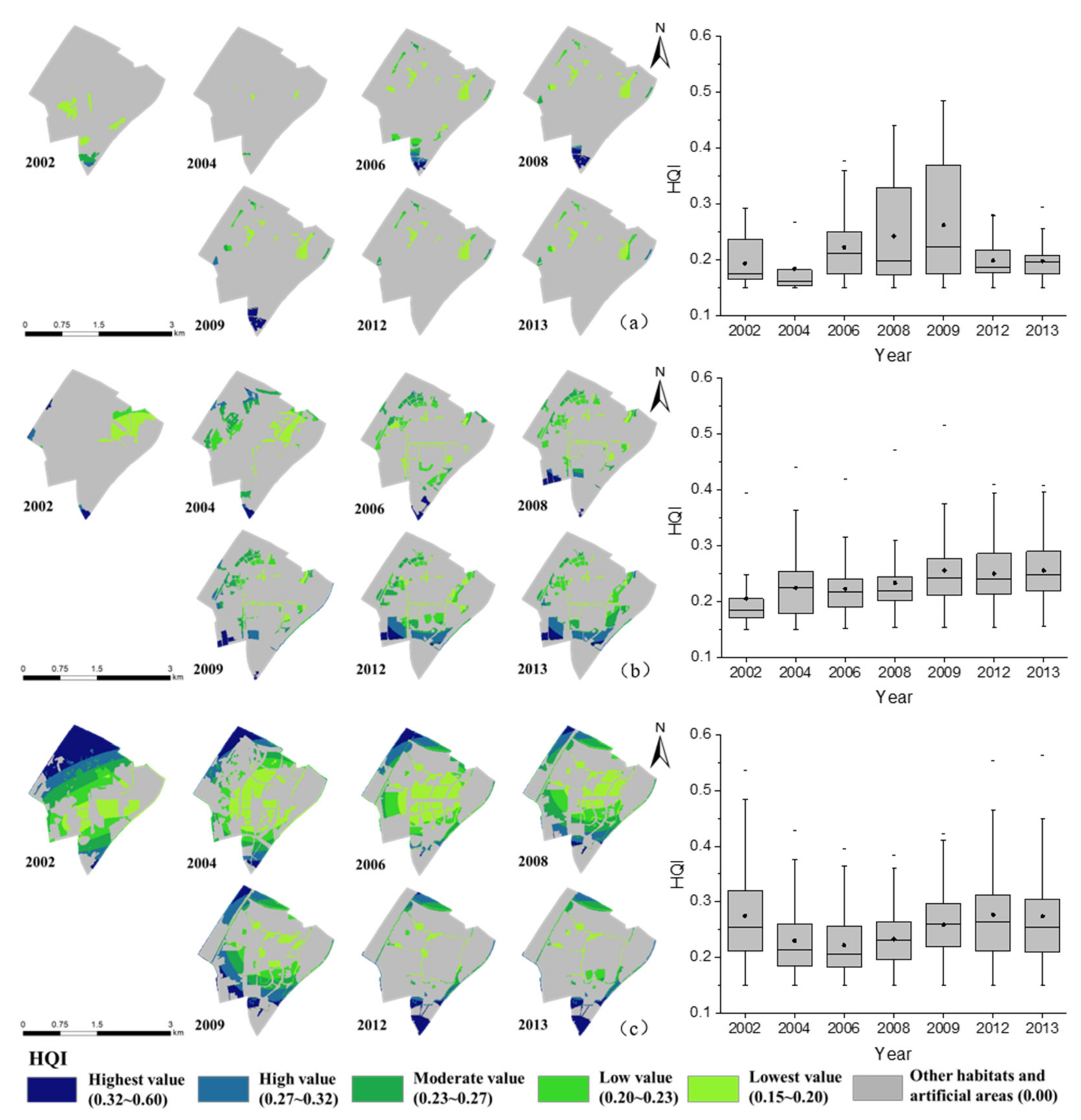
3.2. The Assessments of Habitat Quality Calculated Using the InVEST Model
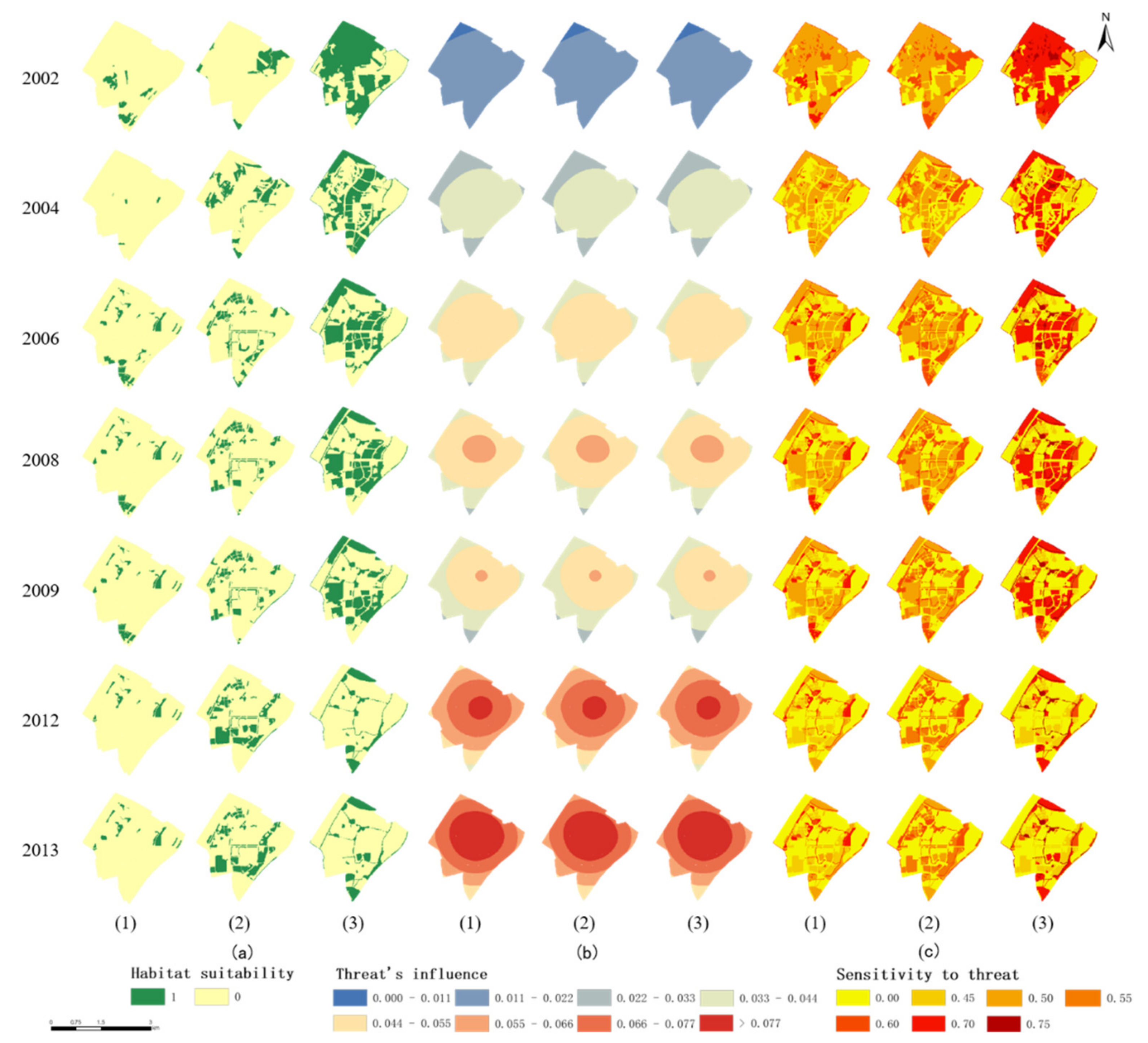
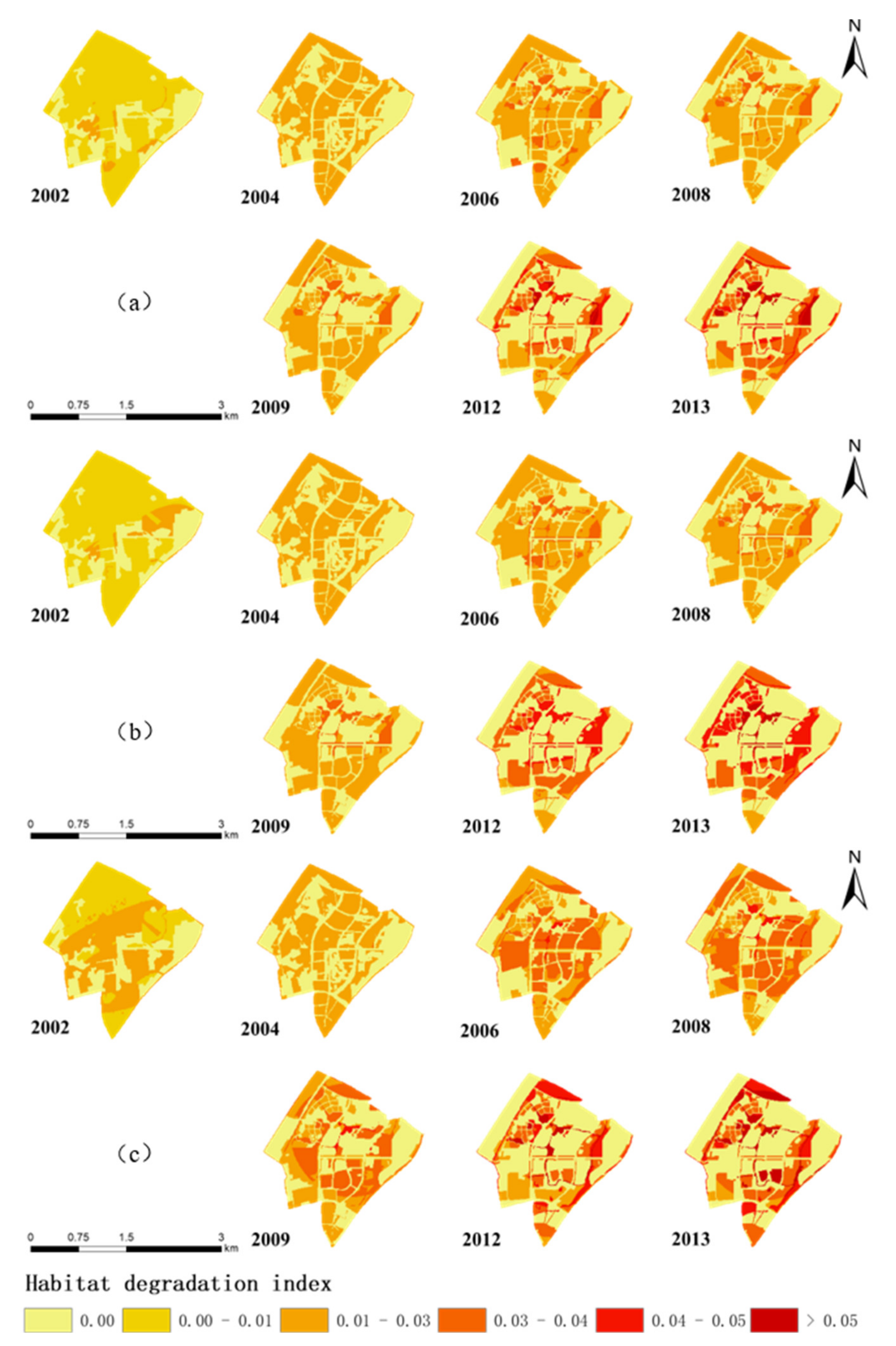
3.2.1. Habitat Quality for the Forest Areas
3.2.2. Habitat Quality for Open Areas
3.2.3. Habitat Quality for Water Area
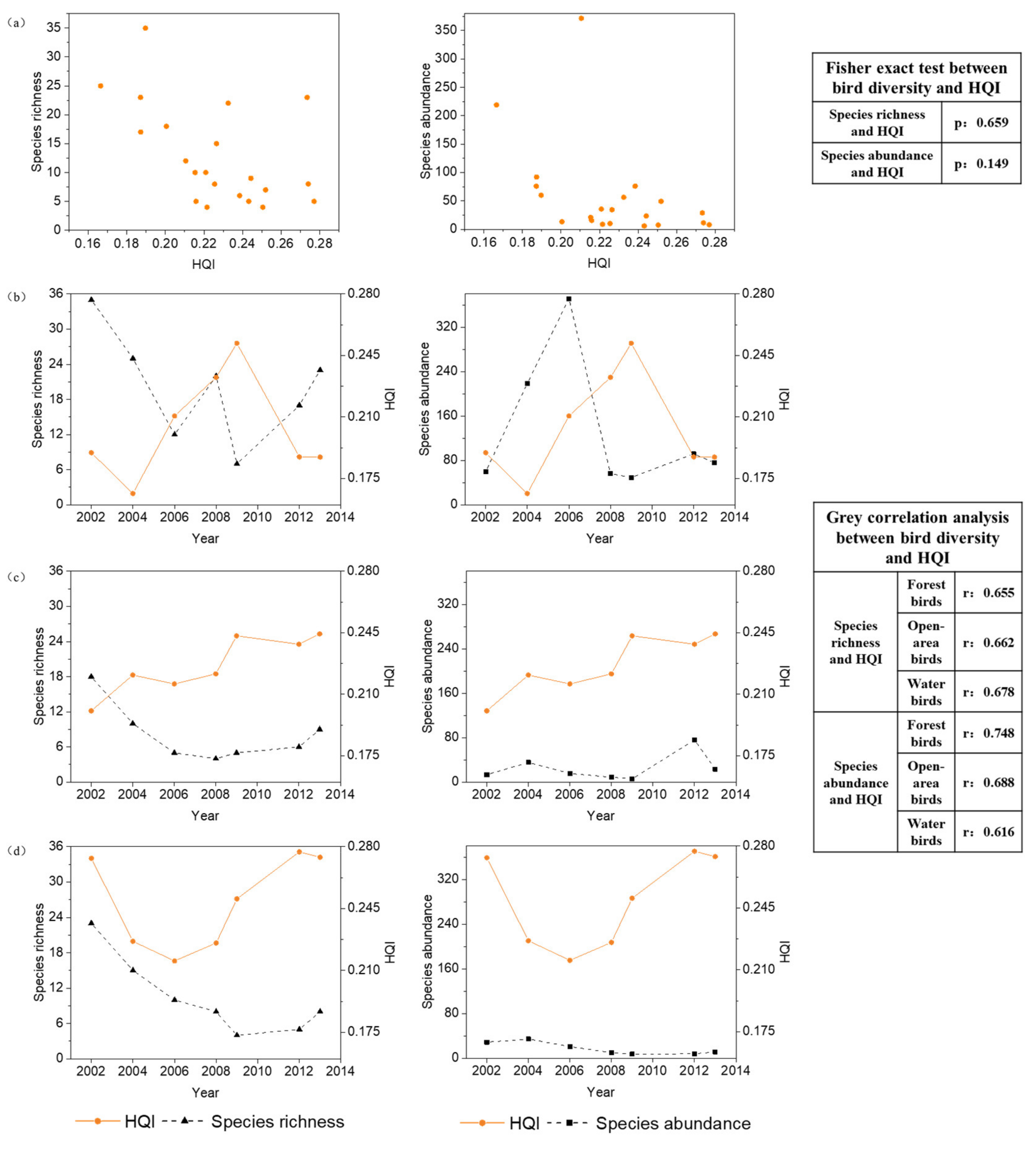
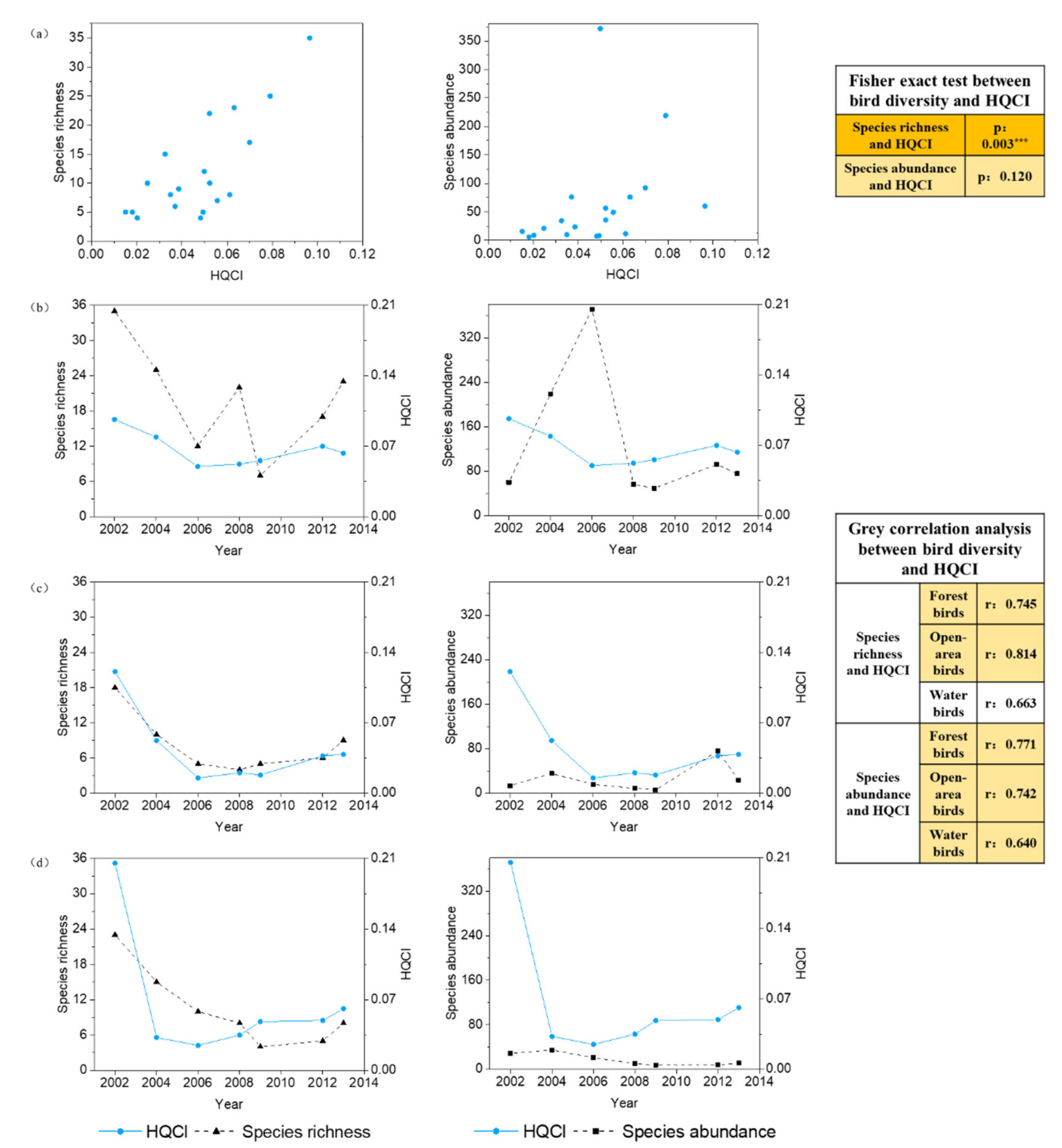
3.3. The Relationships between Bird Diversity and HQI
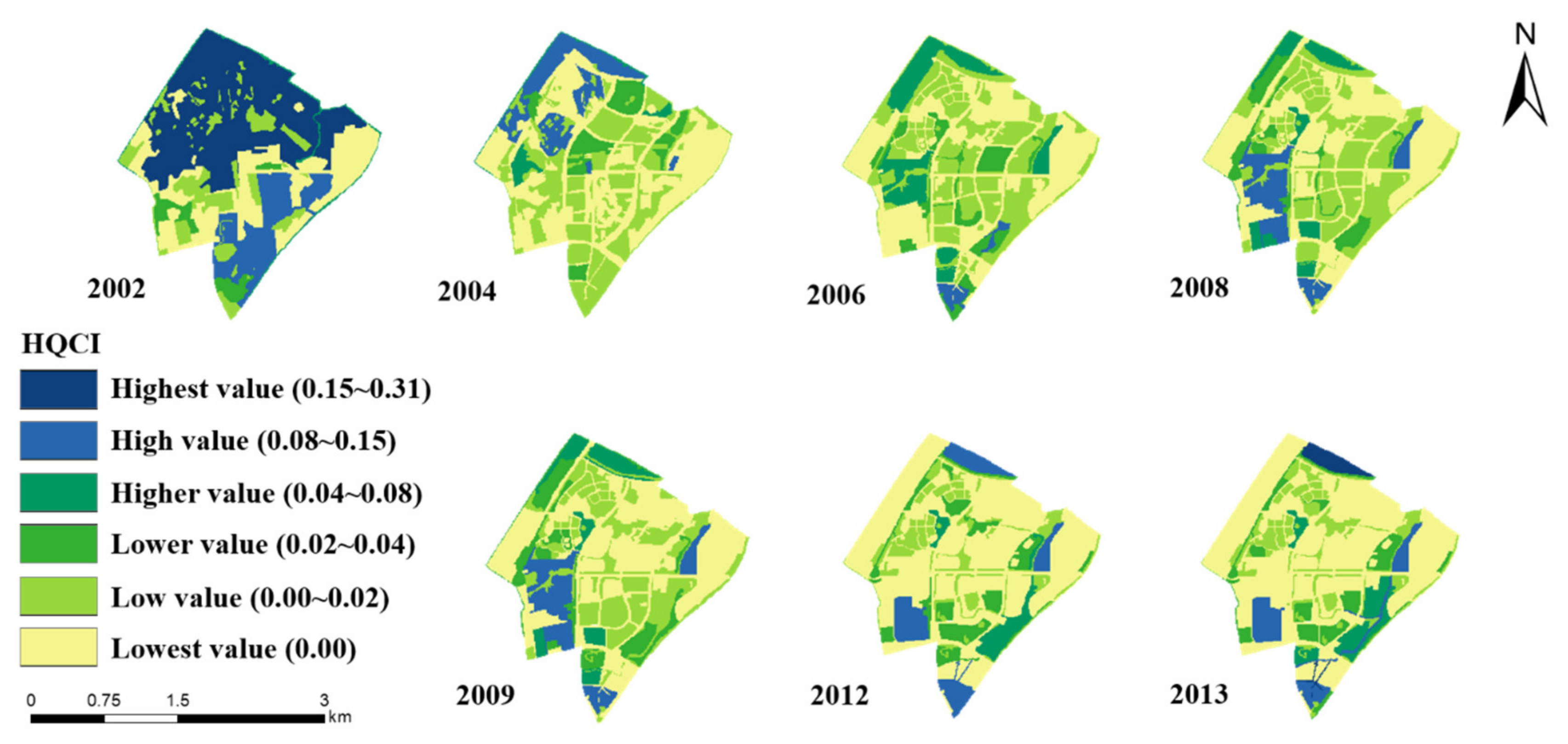
3.4. The New Compound Indicator HQCI and Its Relationship with the Bird Diversity
3.4.1. The Spatial and Temporal Variations of the New Compound Indicator HQCI
3.4.2. Relationship between Bird Diversity and the HQCI
4. Discussion
4.1. The Problems of the Habitat Quality Indicator in Explaining Bird Diversity at the Local Scale
4.2. Applicability of the New Combined Indicator of Habitat Quality and Connectivity in Explaining Bird Diversity
4.3. Limitations of the Present Study and Potential Guidance for Further Studies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Classification of Species and the Explanations of the Habitat Types for Birds
| Name | Family | Species | Habitat Type |
| Japanese Quail | Phasianidae | Coturnix japonica | O1 |
| Common Pheasant | Phasianidae | Phasianus colchicus | F3 |
| Mandarin Duck | Anatidae | Aix galericulata | W1–F2 |
| Mallard | Anatidae | Anas platyrhynchos | W1 |
| Spot-billed Duck | Anatidae | Anas poecilorhyncha | W1 |
| Philippine Duck | Anatidae | Anas luzonica | W1 |
| Tufted duck | Anatidae | Aythya fuligula | W1 |
| Eurasian hoopoe | Upupidae | Upupa epops | O2 |
| Dollarbird | Coraciidae | Eurystomus orientalis | F2 |
| Common Kingfisher | Alcedinidae | Alcedo atthis | W3 |
| Black-capped Kingfisher | Alcedinidae | Halcyon pileata | W3 |
| Pied Kingfisher | Cerylidae | Ceryle rudis | W3 |
| Eurasian Cuckoo | Cuculidae | Cuculus canorus | F2–W3 |
| Lesser Cuckoo | Cuculidae | Cuculus poliocephalus | F2 |
| Lesser Coucal | Centropodidae | Centropus bengalensis | F3–W3 |
| Fork-tailed Swift | Apodidae | Apus pacificus | O1 |
| Indian Jungle Nightjar | Caprimulgidae | Caprimulgus indicus | F1 |
| Oriental Turtle Dove | Columbidae | Streptopelia orientalis | O2 |
| Spotted Dove | Columbidae | Streptopelia chinensis | O2 |
| White-breasted Waterhen | Rallidae | Amaurornis phoenicurus | W3 |
| Common Moorhen | Rallidae | Gallinula chloropus | W1–O1 |
| Common Coot | Rallidae | Fulica atra | W1 |
| Common Snipe | Scolopacidae | Gallinago gallinago | W3 |
| Common Greenshank | Scolopacidae | Tringa nebularia | W2 |
| Green Sandpiper | Scolopacidae | Tringa ochropus | W2 |
| Wood Sandpiper | Scolopacidae | Tringa glareola | W2 |
| Common Sandpiper | Scolopacidae | Actitis hypoleucos | W2 |
| Rufous-necked Stint | Scolopacidae | Calidris ruficollis | W2 |
| Temminck’s Stint | Scolopacidae | Calidris temminckii | W2 |
| Long-toed Stint | Scolopacidae | Calidris subminuta | W2 |
| Pheasant-tailed Jacana | Jacanidae | Hydrophasianus chirurgus | W3 |
| Pacific Golden Plover | Charadriidae | Pluvialis fulva | W2 |
| Little Ringed Plover | Charadriidae | Charadrius dubius | W2 |
| Kentish Plover | Charadriidae | Charadrius alexandrinus | W2 |
| Mew Gul | Laridae | Larus canus | W3 |
| Hen harrier | Accipitridae | Circus cyaneus | W1 |
| Chinese Goshaw | Accipitridae | Accipiter soloensis | F1 |
| Common Buzzard | Accipitridae | Buteo buteo | O1 |
| Common Kestrel | Falconidae | Falco tinnunculus | O1 |
| Little Grebe | Podicipedidae | Tachybaptus ruficollis | W1 |
| Great Crested Grebe | Podicipedidae | Podiceps cristatus | W1 |
| Little Egret | Ardeidae | Egretta garzetta | W2–F2 |
| Gray Heron | Ardeidae | Ardea cinerea | W2–F2 |
| Large Egret | Ardeidae | Ardea alba | W2 |
| Intermediate Egret | Ardeidae | Ardea intermedia | W2 |
| Cattle Egret | Ardeidae | Bubulcus ibis | W2 |
| Chinese Pond Heron | Ardeidae | Ardeola bacchus | W2 |
| Striated Heron | Ardeidae | Butorides striatus | W2–W3 |
| Black-crowned Night Heron | Ardeidae | Nycticorax nycticorax | W2–F2 |
| Yellow Bittern | Ardeidae | Ixobrychus sinensis | W3 |
| Brown Shrike | Laniidae | Lanius cristatus | F3 |
| Long-tailed Shrike | Laniidae | Lanius schach | F3 |
| Azure-winged Magpie | Corvidae | Cyanopica cyanus | F2 |
| Black-billed Magpie | Corvidae | Pica hudsonia | F2 |
| Black Drongo | Dicruridae | Dicrurus macrocercus | F2 |
| Hair-crested Drongo | Dicruridae | Dicrurus hottentottus | F1 |
| White-throated Rock Thrush | Muscicapidae | Monticola gularis | F1 |
| Scaly Thrush | Muscicapidae | Zoothera dauma | F1 |
| Gray-backed Thrush | Muscicapidae | Turdus hortulorum | F1 |
| Japanese Thrush | Turdidae | Turdus cardis | F3 |
| Eurasian Blackbird | Muscicapidae | Turdus merula | F2 |
| Eyebrowed Thrush | Muscicapidae | Turdus obscurus | F1 |
| Pale Thrush | Muscicapidae | Turdus pallidus | F1 |
| Dusky Thrush | Muscicapidae | Turdus eunomus | O1 |
| Gray-streaked Flycatcher | Muscicapidae | Muscicapa griseisticta | F1 |
| Asian Brown Flycatcher | Muscicapidae | Muscicapa dauurica | F1 |
| Narcissus Flycatcher | Muscicapidae | Ficedula narcissina | F1 |
| Mugimaki Flycatcher | Muscicapidae | Ficedula mugimaki | F2 |
| Blue-and-white Flycatcher | Muscicapidae | Cyanoptila cyanomelana | F1 |
| Bluethroat | Muscicapidae | Luscinia svecica | F3 |
| Orange-flanked Bush-Robin | Muscicapidae | Tarsiger cyanurus | F1 |
| Daurian Redstart | Muscicapidae | Phoenicurus auroreus | F3 |
| White-cheeked Starling | Sturnidae | Sturnus cineraceus | O2 |
| Black-collared Starling | Sturnidae | Gracupica nigricollis | O2 |
| Crested Myna | Sturnidae | Acridotheres cristatellus | O1 |
| Chinese Penduline Tit | Remizidae | Remiz consobrinus | W3 |
| Yellow-bellied Tit | Paridae | Pardaliparus venustulus | F3 |
| Great Tit | Paridae | Parus major | F2 |
| Black-throated Tit | Aegithalidae | Aegithalos concinnus | F3 |
| Barn Swallow | Hirundinidae | Hirundo rustica | O1 |
| Red-rumped Swallow | Hirundinidae | Hirundo daurica | O1 |
| Light-vented Bulbul | Pycnonotidae | Pycnonotus sinensis | F2 |
| Himalayan Black Bulbul | Pycnonotidae | Hypsipetes leucocephalus | F1 |
| Zitting Cisticola | Cisticolidae | Cisticola juncidis | O1 |
| Plain Prinia | Cisticolidae | Prinia inornata | W3 |
| Manchurian Bush Warbler | Sylviidae | Horornis canturians | F3 |
| Japanese Bush-Warbler | Sylviidae | Horornis diphone | F3 |
| Brownish-flanked Bush-Warbler | Sylviidae | Horornis fortipes | F3 |
| Oriental Reed Warbler | Sylviidae | Acrocephalus orientalis | W3 |
| Yellow-rumped Warbler | Sylviidae | Setophaga coronata | F3 |
| Yellow-browed Warbler | Sylviidae | Phylloscopus inornatus | F1 |
| Pale-legged Warbler | Sylviidae | Phylloscopus tenellipes | F1–F3 |
| Eastern Crowned Warbler | Sylviidae | Phylloscopus coronatus | F1 |
| Masked Laughingthrush | Sylviidae | Garrulax perspicillatus | F3 |
| Greater Necklaced Laughingthrush | Sylviidae | Pterorhinus pectoralis | F1 |
| Hwamei | Sylviidae | Garrulax canorus | F3 |
| Vinous-throated Parrotbill | Paradoxornis | Sinosuthora webbianus | F3 |
| Eurasian Skylark | Alaudidae | Alauda arvensis | O1 |
| Oriental Skylark | Alaudidae | Alauda gulgula | O1 |
| Eurasian Tree Sparrow | Passeridae | Passer montanus | F2 |
| Forest Wagtail | Motacillidae | Dendronanthus indicus | F1 |
| White Wagtail | Passeridae | Motacilla alba | O1–W2 |
| Yellow Wagtail | Passeridae | Motacilla flava | O1–W2 |
| Gray Wagtail | Passeridae | Motacilla cinerea | O1–W2 |
| Oriental Tree Pipit | Passeridae | Anthus hodgsoni | F2 |
| White-rumped Munia | Passeridae | Lonchura striata | F3 |
| Brambling | Fringillidae | Fringilla montifringilla | F1 |
| Gray-capped Greenfinch | Fringillidae | Carduelis sinica | F2 |
| Yellow-billed Grosbeak | Fringillidae | Eophona migratoria | F2 |
| Meadow Bunting | Fringillidae | Emberiza cioides | F2 |
| Tristram’s Bunting | Fringillidae | Emberiza tristrami | F3 |
| Yellow-browed Bunting | Fringillidae | Emberiza chrysophrys | F3 |
| Rustic Bunting | Fringillidae | Emberiza rustica | F3 |
| Yellow-throated Bunting | Fringillidae | Emberiza elegans | F1 |
| Yellow-breasted Bunting | Emberizidae | Emberiza aureola | O1 |
| Black-faced Bunting | Fringillidae | Emberiza spodocephala | F3 |
| Where the habitat types of the birds were classified as F—forest area (F1—forest species that only use forested areas, F2—forest species that also use open areas, and F3—forest species that use boscage areas), O—open area (O1—open area species and O2—species that prefer open areas, but also use forested areas), W—water area (W1—swimming birds that use open water, W2—species that conceal themselves in marshes and aquatic areas with high grass, and W3—waders). | |||
Appendix B. Input Parameters, Including Habitat Suitability and Sensitivity to the Threat to Different Habitat Types for Birds

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land Use Types | Habitat Suitability | Sensitivity to Threat | ||||
|---|---|---|---|---|---|---|
| Forest Birds | Open-Area Birds | Water Birds | Forest Birds | Open-Area Birds | Water Birds | |
| Built-up land | 0 | 0 | 0 | 0 | 0 | 0 |
| Forestland | 1 | 0 | 0 | 0.7 | 0.6 | 0.6 |
| Bare ground | 0 | 0 | 0 | 0 | 0 | 0 |
| Shrubland | 0 | 1 | 0 | 0.5 | 0.6 | 0.45 |
| Water body | 0 | 0 | 1 | 0.6 | 0.6 | 0.75 |
| Wetland | 0 | 0 | 1 | 0.5 | 0.5 | 0.7 |
| Grassland | 0 | 1 | 0 | 0.45 | 0.55 | 0.45 |
Appendix C. Input Layers for the Calculation of HQI and Variations to an Intermediate Index, the Habitat Degradation Index, for the Three Types of the Habitats in New Jiangwan Town, from 2002 to 2013


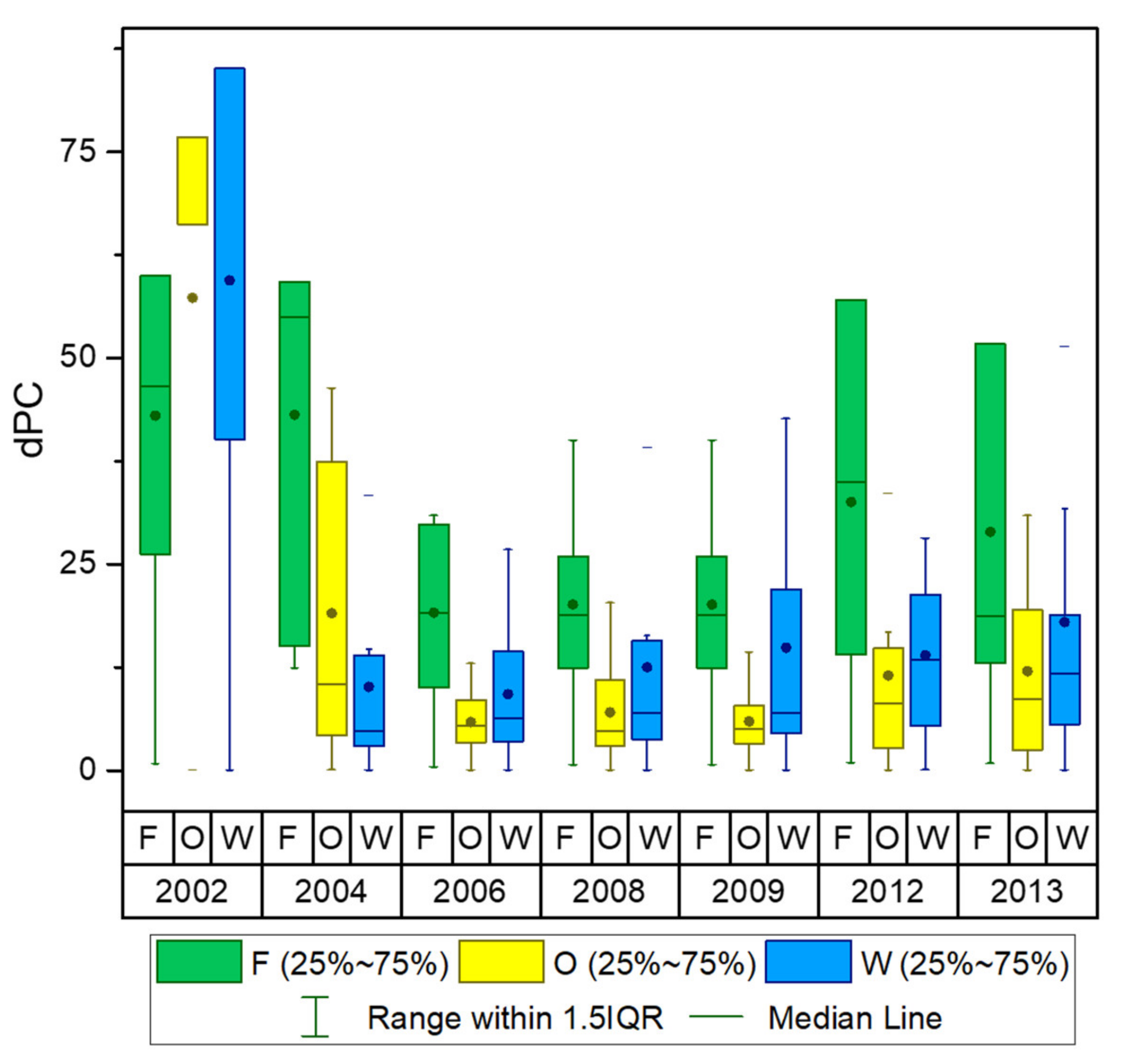
Appendix D. Distribution Maps and the Statistical Box-Plots of the dPC


References
- Mace, G.M.; Norris, K.; Fitter, A.H. Biodiversity and ecosystem services: A multilayered relationship. Trends Ecol. Evol. 2012, 27, 19–26. [Google Scholar] [CrossRef]
- Penvern, S.; Fernique, S.; Cardona, A.; Herz, A.; Ahrenfeldt, E.; Dufils, A.; Jamar, L.; Korsgaard, M.; Kruczyńska, D.; Matray, S.; et al. Farmers’ management of functional biodiversity goes beyond pest management in organic European apple orchards. Agric. Ecosyst. Environ. 2019, 284, 106555. [Google Scholar] [CrossRef] [Green Version]
- Ebeling, A.; Lind, E.W.; Meyer, S.T.; Barnes, A.D.; Borer, E.T.; Eisenhauer, N.; Weisser, W.W. Contrasting effects of plant diversity on beta- and gamma-diversity of grassland invertebrates. Ecology 2020, 101, e03057-10. [Google Scholar] [CrossRef]
- Armenteras, D.; Rodríguez, N.; Retana, J. National and regional relationships of carbon storage and tropical biodiversity. Biol. Conserv. 2015, 192, 378–386. [Google Scholar] [CrossRef]
- Thorup-Kristensen, K.; Dresbøll, D.B.; Kristensen, H.L. Crop yield, root growth, and nutrient dynamics in a conventional and three organic cropping systems with different levels of external inputs and N re-cycling through fertility building crops. Eur. J. Agron. 2012, 37, 66–82. [Google Scholar] [CrossRef] [Green Version]
- Harrison, P.; Berry, P.; Simpson, G.; Haslett, J.; Blicharska, M.; Bucur, M.; Dunford, R.; Egoh, B.; Garcia-Llorente, M.; Geamănă, N.; et al. Linkages between biodiversity attributes and ecosystem services: A systematic review. Ecosyst. Serv. 2014, 9, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Díaz, S.; Demissew, S.; Carabias, J.; Joly, C.; Lonsdale, M.; Ash, N.; Larigauderie, A.; Adhikari, J.R.; Arico, S.; Báldi, A.; et al. The IPBES Conceptual Framework—Connecting nature and people. Curr. Opin. Environ. Sustain. 2015, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Pan, Y.; Liu, Y.; Zhao, H.; Wang, Y. Linking ecological degradation risk to identify ecological security patterns in a rapidly urbanizing landscape. Habitat Int. 2018, 71, 110–124. [Google Scholar] [CrossRef]
- Khoury, C.K.; Amariles, D.; Soto, J.S.; Diaz, M.V.; Sotelo, S.; Sosa, C.C.; Ramírez-Villegas, J.; Achicanoy, H.A.; Velásquez-Tibatá, J.; Guarino, L.; et al. Comprehensiveness of conservation of useful wild plants: An operational indicator for biodiversity and sustainable development targets. Ecol. Indic. 2019, 98, 420–429. [Google Scholar] [CrossRef]
- Heli, S.; Jyri, M.; Turo, H.; Kaisu, A. Participatory multi-criteria decision analysis in valuing peatland ecosystem services-Trade-offs related to peat extraction vs. pristine peatlands in Southern Finland. Ecol. Econ. 2019, 162, 17–28. [Google Scholar]
- Vaissière, A.-C.; Levrel, H.; Scemama, P. Biodiversity offsetting: Clearing up misunderstandings between conservation and economics to take further action. Biol. Conserv. 2017, 206, 258–262. [Google Scholar] [CrossRef] [Green Version]
- Morelli, F.; Jiguet, F.; Sabatier, R.; Dross, C.; Princé, K.; Tryjanowski, P.; Tichit, M. Spatial covariance between ecosystem services and biodiversity pattern at a national scale (France). Ecol. Indic. 2017, 82, 574–586. [Google Scholar] [CrossRef]
- Fisher, R.A.; Corbet, A.S.; Williams, C.B. The Relation Between the Number of Species and the Number of Individuals in a Random Sample of an Animal Population. J. Anim. Ecol. 1943, 12, 42. [Google Scholar] [CrossRef]
- Bartkowski, B.; Lienhoop, N.; Hansjürgens, B. Capturing the complexity of biodiversity: A critical review of economic valuation studies of biological diversity. Ecol. Econ. 2015, 113, 1–14. [Google Scholar] [CrossRef]
- Purvis, A.; Hector, A. Getting the measure of biodiversity. Nature 2000, 405, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Hackman, K.O.; Gong, P. Biodiversity estimation of the western region of Ghana using arthropod mean morphospecies abundance. Biodivers. Conserv. 2017, 26, 2083–2097. [Google Scholar] [CrossRef]
- Ren, C.; Zhang, W.; Zhong, Z.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Differential responses of soil microbial biomass, diversity, and compositions to altitudinal gradients depend on plant and soil characteristics. Sci. Total. Environ. 2018, 610–611, 750–758. [Google Scholar] [CrossRef]
- Yue, Y.; Zhuang, Y.; Yeh, A.G.-O.; Xie, J.-Y.; Ma, C.-L.; Li, Q.-Q. Measurements of POI-based mixed use and their relationships with neighbourhood vibrancy. Int. J. Geogr. Inf. Sci. 2017, 31, 658–675. [Google Scholar] [CrossRef] [Green Version]
- Mora, F. The use of ecological integrity indicators within the natural capital index framework: The ecological and economic value of the remnant natural capital of México. J. Nat. Conserv. 2019, 47, 77–92. [Google Scholar] [CrossRef]
- Gao, T.; Nielsen, A.B.; Hedblom, M. Reviewing the strength of evidence of biodiversity indicators for forest ecosystems in Europe. Ecol. Indic. 2015, 57, 420–434. [Google Scholar] [CrossRef]
- Pomerantz, A.; Peñafiel, N.; Arteaga, A.; Bustamante, L.; Pichardo, F.; Coloma, L.A.; Barrio-Amorós, C.L.; Salazar-Valenzuela, D.; Prost, S. Real-time DNA barcoding in a rainforest using nanopore sequencing: Opportunities for rapid biodiversity assessments and local capacity building. GigaScience 2018, 7, giy033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, F.; Lenz, R. Ecological indicators: Theoretical fundamentals of consistent applications in environmental management—Introduction. Ecol. Indic. 2006, 6, 1–5. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Hansen, M.C.; Loveland, T.R. A review of large area monitoring of land cover change using Landsat data. Remote Sens. Environ. 2012, 122, 66–74. [Google Scholar] [CrossRef]
- Ma, L.; Sun, R.; Kazemi, E.; Pang, D.; Zhang, Y.; Sun, Q.; Zhou, J.; Zhang, K. Evaluation of Ecosystem Services in the Dongting Lake Wetland. Water 2019, 11, 2564. [Google Scholar] [CrossRef] [Green Version]
- Pham, H.V.; Sperotto, A.; Torresan, S.; Acuña, V.; Jorda-Capdevila, D.; Rianna, G.; Marcomini, A.; Critto, A. Coupling scenarios of climate and land-use change with assessments of potential ecosystem services at the river basin scale. Ecosyst. Serv. 2019, 40, 101045. [Google Scholar] [CrossRef]
- Redhead, J.W.; May, L.; Oliver, T.H.; Hamel, P.; Sharp, R.; Bullock, J.M. National scale evaluation of the InVEST nutrient retention model in the United Kingdom. Sci. Total. Environ. 2018, 610–611, 666–677. [Google Scholar] [CrossRef]
- Sun, X.; Crittenden, J.C.; Li, F.; Lu, Z.; Dou, X. Urban expansion simulation and the spatio-temporal changes of ecosystem services, a case study in Atlanta Metropolitan area, USA. Sci. Total. Environ. 2018, 622–623, 974–987. [Google Scholar] [CrossRef]
- Terrado, M.; Sabater, S.; Chaplin-Kramer, B.; Mandle, L.; Ziv, G.; Acuña, V. Model development for the assessment of terrestrial and aquatic habitat quality in conservation planning. Sci. Total. Environ. 2016, 540, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Upadhaya, S.; Dwivedi, P. Conversion of forestlands to blueberries: Assessing implications for habitat quality in Alabaha river watershed in Southeastern Georgia, United States. Land Use Policy 2019, 89, 104229. [Google Scholar] [CrossRef]
- Wu, C.F.; Lin, Y.P.; Chiang, L.C.; Huang, T. Assessing highway’s impacts on landscape patterns and ecosystem services: A case study in Puli Township, Taiwan. Landsc. Urban Plan. 2014, 128, 60–71. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, X.; Qu, L.; Li, S.; Lin, Y.; Yao, R.; Zhou, X.; Li, J. Land use/cover predictions incorporating ecological security for the Yangtze River Delta region, China. Ecol. Indic. 2020, 119, 106841. [Google Scholar] [CrossRef]
- Gong, J.; Xie, Y.C.; Cao, E.J.; Huang, Q.Y.; Li, H.Y. Integration of InVEST-habitat quality model with landscape pattern indexes to assess mountain plant biodiversity change: A case study of Bailongjiang watershed in Gansu Province. J. Geogr. Sci. 2019, 29, 1193–1210. [Google Scholar] [CrossRef] [Green Version]
- Tallis, H.; Polasky, S. Mapping and Valuing Ecosystem Services as an Approach for Conservation and Natural-Resource Management. Ann. N. Y. Acad. Sci. 2009, 1162, 265–283. [Google Scholar] [CrossRef] [PubMed]
- Verhagen, W.; Van Teeffelen, A.J.A.; Compagnucci, A.B.; Poggio, L.; Gimona, A.; Verburg, P. Effects of landscape configuration on mapping ecosystem service capacity: A review of evidence and a case study in Scotland. Landsc. Ecol. 2016, 31, 1457–1479. [Google Scholar] [CrossRef]
- Duarte, G.T.; Santos, P.M.; Cornelissen, T.G.; Ribeiro, M.C.; Paglia, A.P. The effects of landscape patterns on ecosystem services: Meta-analyses of landscape services. Landsc. Ecol. 2018, 33, 1247–1257. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhao, J. Investigating the Spatiotemporally Varying Correlation between Urban Spatial Patterns and Ecosystem Services: A Case Study of Nansihu Lake Basin, China. ISPRS Int. J. Geo-Inf. 2019, 8, 346. [Google Scholar] [CrossRef] [Green Version]
- Lenore, F.; Jacques, B.; Lluís, B.; Burel, F.G.; Crist, T.O.; Fuller, R.J.; Clelia, S.; Siriwardena, G.M.; Jean-Louis, M. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Lett. 2011, 14, 101–112. [Google Scholar]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of habitat fragmentation on biodiversity [Review]. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef] [Green Version]
- Debinski, D.M.; Holt, R.D. A Survey and Overview of Habitat Fragmentation Experiments. Conserv. Biol. 2000, 14, 342–355. [Google Scholar] [CrossRef]
- Theodorou, P.; Radzevičiūtė, R.; Lentendu, G.; Kahnt, B.; Husemann, M.; Bleidorn, C.; Settele, J.; Schweiger, O.; Grosse, I.; Wubet, T.; et al. Urban areas as hotspots for bees and pollination but not a panacea for all insects. Nat. Commun. 2020, 11, 576. [Google Scholar] [CrossRef] [PubMed]
- Reis, E.; López-Iborra, G.M.; Pinheiro, R.T. Changes in bird species richness through different levels of urbanization: Implications for biodiversity conservation and garden design in Central Brazil. Landsc. Urban Plan. 2012, 107, 31–42. [Google Scholar] [CrossRef]
- Bregman, T.P.; Sekercioglu, C.H.; Tobias, J.A. Global patterns and predictors of bird species responses to forest fragmentation: Implications for ecosystem function and conservation. Biol. Conserv. 2014, 169, 372–383. [Google Scholar] [CrossRef]
- Lopes, E.V.; Mendonça, L.B.; Junior, M.A.D.S.; López-Iborra, G.M.; Dos Anjos, L. Effects of Connectivity on the Forest Bird Communities of Adjacent Fragmented Landscapes. Ardeola 2016, 63, 279–293. [Google Scholar] [CrossRef] [Green Version]
- Pereira, J.; Saura, S.; Jordán, F. Single-node vs. multi-node centrality in landscape graph analysis: Key habitat patches and their protection for 20 bird species in NE Spain. Methods Ecol. Evol. 2017, 8, 1458–1467. [Google Scholar] [CrossRef]
- Saura, S.; Pascual-Hortal, L. A new habitat availability index to integrate connectivity in landscape conservation planning: Comparison with existing indices and application to a case study. Landsc. Urban Plan. 2007, 83, 91–103. [Google Scholar] [CrossRef]
- Yang, Y.C.; Da, L.J.; Ji, F. Study on diversity of plant community at Jiangwan Airport, Shanghai. Shanghai Environ. Sci. 2003, 9, 615–618. (In Chinese) [Google Scholar]
- Jin, X.B.; Zhou, B.C.; Qin, X.K.; Cui, Z.X.; Xia, J.H.; Si, Q.; Liu, M.P. Biodiversity of the Jiangwan Airport in Shanghai. In Proceedings of the 6th National Workshop on Biodiversity Conservation and Sustainable Use, Li Jiang, China, 12–14 May 2004; Biodiversity Committee, Chinese Academy of Sciences: Li Jiang, China, 2004; pp. 1–36. (In Chinese). [Google Scholar]
- Yang, Y.C.; Wang, J.; Da, L.J. Diversity, spatial pattern and dynamics of vegetation under urbanization in Shanghai (II): Study on the flora of Jiangwan Airport, an abondoned land, Shanghai (In Chinese). J. East China Norm. Univ. (Nat. Sci.) 2008, 4, 40–48. [Google Scholar]
- Xu, X.; Xie, Y.; Qi, K.; Luo, Z.; Wang, X. Detecting the response of bird communities and biodiversity to habitat loss and fragmentation due to urbanization. Sci. Total. Environ. 2018, 624, 1561–1576. [Google Scholar] [CrossRef]
- Zhou, D.; Fung, T.; Chu, L. Avian community structure of urban parks in developed and new growth areas: A landscape-scale study in Southeast Asia. Landsc. Urban Plan. 2012, 108, 91–102. [Google Scholar] [CrossRef]
- Luan, X.F. Studies on Avian Community of Shanghai and Planning of Conservation. Ph.D. Thesis, East China Normal University Shanghai, Shanghai, China, 2003. [Google Scholar]
- Fahrig, L. Ecological Responses to Habitat Fragmentation Per Se. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 1–23. [Google Scholar] [CrossRef]
- Baral, H.; Keenan, R.J.; Sharma, S.K.; Stork, N.E.; Kasel, S. Spatial assessment and mapping of biodiversity and conservation priorities in a heavily modified and fragmented production landscape in north-central Victoria, Australia. Ecol. Indic. 2014, 36, 552–562. [Google Scholar] [CrossRef]
- He, J.; Huang, J.; Li, C. The evaluation for the impact of land use change on habitat quality: A joint contribution of cellular automata scenario simulation and habitat quality assessment model. Ecol. Model. 2017, 366, 58–67. [Google Scholar] [CrossRef]
- Yang, S.; Zhao, W.; Liu, Y.; Wang, S.; Wang, J.; Zhai, R. Influence of land use change on the ecosystem service trade-offs in the ecological restoration area: Dynamics and scenarios in the Yanhe watershed, China. Sci. Total. Environ. 2018, 644, 556–566. [Google Scholar] [CrossRef]
- Rimal, B.; Sharma, R.; Kunwar, R.; Keshtkar, H.; Stork, N.E.; Rijal, S.; Rahman, S.A.; Baral, H. Effects of land use and land cover change on ecosystem services in the Koshi River Basin, Eastern Nepal. Ecosyst. Serv. 2019, 38, 12. [Google Scholar] [CrossRef]
- Wu, J.; Feng, Z.; Gao, Y.; Peng, J. Hotspot and relationship identification in multiple landscape services: A case study on an area with intensive human activities. Ecol. Indic. 2013, 29, 529–537. [Google Scholar] [CrossRef]
- Xu, W.Q. Effects of Landscape Pattern Dynamics on the Habitat of Migratory Waterbirds—A Case Study in National Nature Reserve of Hunan Xinxiang Yellow River Wetland. Master’s Thesis, Henan University, Kaifeng, China, 2016. (In Chinese). [Google Scholar]
- Liu, Y.; Zhou, Y.; Du, Y.T. Study on the spatio-temporal patterns of habitat quality and its terrain gradient effects of the middle of the Yangtze river economic belt based on InVEST model. Resour. Environ. Yangtze Basin 2019, 28, 2429–2440. (In Chinese) [Google Scholar]
- Li, F.; Wang, L.; Chen, Z.; Clarke, K.C.; Li, M.; Jiang, P. Extending the SLEUTH model to integrate habitat quality into urban growth simulation. J. Environ. Manag. 2018, 217, 486–498. [Google Scholar] [CrossRef] [Green Version]
- Sharp, R.; Tallis, H.T.; Ricketts, T.; Guerry, A.D.; Wood, S.A.; Chaplin-Kramer, R.; Vogl, A.L. InVEST Users Guide; Stanford: The Natural Capital Project: Stanford University, University of Minnesota, The Nature Conservancy, and World Wildlife Fund, Stanford, CA, USA, 2014. [Google Scholar]
- Mitchell, M.G.E.; Suarez-Castro, A.F.; Martinez-Harms, M.; Maron, M.; McAlpine, C.; Gaston, K.J.; Johansen, K.; Rhodes, J.R. Reframing landscape fragmentation’s effects on ecosystem services. Trends Ecol. Evol. 2015, 30, 190–198. [Google Scholar] [CrossRef] [Green Version]
- Volk, X.K.; Gattringer, J.P.; Otte, A.; Harvolk-Schöning, S. Connectivity analysis as a tool for assessing restoration success. Landsc. Ecol. 2018, 33, 371–387. [Google Scholar] [CrossRef]
- Watson, J.E.M.; Whittaker, R.J.; Freudenberger, D. Bird community responses to habitat fragmentation: How consistent are they across landscapes? J. Biogeogr. 2005, 32, 1353–1370. [Google Scholar] [CrossRef]
- Cadavid-Florez, L.; Laborde, J.; Mclean, D.J. Isolated trees and small woody patches greatly contribute to connectivity in highly fragmented tropical landscapes. Landsc. Urban Plan. 2020, 196, 103745. [Google Scholar] [CrossRef]
- Radford, J.Q.; Bennett, A.F. Thresholds in landscape parameters: Occurrence of the white-browed treecreeper Climacteris affinis in Victoria, Australia. Biol. Conserv. 2004, 117, 375–391. [Google Scholar] [CrossRef]
- Holzschuh, A.; Steffan-Dewenter, I.; Tscharntke, T. How do landscape composition and configuration, organic farming and fallow strips affect the diversity of bees, wasps and their parasitoids? J. Anim. Ecol. 2010, 79, 491–500. [Google Scholar] [CrossRef]
- Hoyer, M.V.; Canfield, D.E. Bird abundance and species richness on Florida lakes: Influence of trophic status, lake morphology, and aquatic macrophytes. Hydrobiologia 1994, 279, 107–119. [Google Scholar] [CrossRef]
- Fahrig, L. Rethinking patch size and isolation effects: The habitat amount hypothesis. J. Biogeogr. 2013, 40, 1649–1663. [Google Scholar] [CrossRef]
- Howell, C.A.; Latta, S.C.; Donovan, T.M.; Porneluzi, P.A.; Parks, G.R.; Faaborg, J. Landscape effects mediate breeding bird abundance in midwestern forests. Landsc. Ecol. 2000, 15, 547–562. [Google Scholar] [CrossRef]
- Thiele, J.; Kellner, S.; Buchholz, S.; Schirmel, J. Connectivity or area: What drives plant species richness in habitat corridors? Landsc. Ecol. 2018, 33, 173–181. [Google Scholar] [CrossRef]
- Martensen, A.C.; Ribeiro, M.C.; Banks-Leite, C.; Prado, P.I.D.K.L.D.; Metzger, J.P.W. Associations of Forest Cover, Fragment Area, and Connectivity with Neotropical Understory Bird Species Richness and Abundance. Conserv. Biol. 2012, 26, 1100–1111. [Google Scholar] [CrossRef]
- Kammerlander, B.; Breiner, H.-W.; Filker, S.; Sommaruga, R.; Sonntag, B.; Stoeck, T. High diversity of protistan plankton communities in remote high mountain lakes in the European Alps and the Himalayan mountains. FEMS Microbiol. Ecol. 2015, 91, fiv010. [Google Scholar] [CrossRef] [Green Version]
- Kaleebu, P.; Nankya, I.L.; Yirrell, D.L.; Shafer, L.A.; Kyosiimire-Lugemwa, J.; Lule, D.B.; Morgan, D.; Beddows, S.; Weber, J.; Whitworth, J.A.G. Relation between chemokine receptor use, disease stage, and HIV-1 subtypes A and D—Results from a rural Ugandan cohort. Jaids J. Acquir. Immune Defic. Syndr. 2007, 45, 28–33. [Google Scholar] [CrossRef]
- Angeli, S.I. Phenotype/Genotype Correlations in a DFNB1 Cohort with Ethnical Diversity. Laryngoscope 2008, 118, 2014–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.; Dong, Y.-X.; Yang, R. Urbanization impact on carbon emissions in the Pearl River Delta region: Kuznets curve relationships. J. Clean. Prod. 2018, 180, 514–523. [Google Scholar] [CrossRef]
- Cotter, M.; Häuser, I.; Harich, F.; He, P.; Sauerborn, J.; Treydte, A.; Martin, K.; Cadisch, G. Biodiversity and ecosystem services−A case study for the assessment of multiple species and functional diversity levels in a cultural landscape. Ecol. Indic. 2017, 75, 111–117. [Google Scholar] [CrossRef]
- Posner, S.; Verutes, G.; Koh, I.; Denu, D.; Ricketts, T. Global use of ecosystem service models. Ecosyst. Serv. 2016, 17, 131–141. [Google Scholar] [CrossRef]
- Sultana, M.; Corlatti, L.; Storch, I. The interaction of imperviousness and habitat heterogeneity drives bird richness patterns in south Asian cities. Urban Ecosyst. 2020, 1–10. [Google Scholar] [CrossRef]
- Ruoso, L.-E.; Plant, R.; Maurel, P.; Dupaquier, C.; Roche, P.K.; Bonin, M. Reading Ecosystem Services at the Local Scale through a Territorial Approach: The Case of Peri-Urban Agriculture in the Thau Lagoon, Southern France. Ecol. Soc. 2015, 20, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Morante-Filho, J.C.; Benchimol, M.; Faria, D. Landscape composition is the strongest determinant of bird occupancy patterns in tropical forest patches. Landsc. Ecol. 2021, 36, 105–117. [Google Scholar] [CrossRef]
- Prugh, L.R.; Hodges, K.E.; Sinclair, A.R.E.; Brashares, J.S. Effect of habitat area and isolation on fragmented animal populations. Proc. Natl. Acad. Sci. USA 2008, 105, 20770–20775. [Google Scholar] [CrossRef] [Green Version]
- Opdam, P.; Wascher, D. Climate change meets habitat fragmentation: Linking landscape and biogeographical scale levels in research and conservation. Biol. Conserv. 2004, 117, 285–297. [Google Scholar] [CrossRef]
- Chace, J.F.; Walsh, J.J. Urban effects on native avifauna: A review. Landsc. Urban Plan. 2006, 74, 46–69. [Google Scholar] [CrossRef]
- Ramos, D.L.; Pizo, M.A.; Ribeiro, M.C.; Cruz, R.S.; Morales, J.M.; Ovaskainen, O. Forest and connectivity loss drive changes in movement behavior of bird species. Ecography 2020, 43, 1203–1214. [Google Scholar] [CrossRef]
- Newbold, T.; Scharlemann, J.P.W.; Butchart, S.H.M.; Şekercioğlu, Ç.H.; Alkemade, R.; Booth, H.; Purves, D.W. Ecological traits affect the response of tropical forest bird species to land-use intensity. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122131. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, I.; Proença, V.; Serra, P.; Palma, J.; Domingo-Marimon, C.; Pons, X.; Domingos, T. Remotely sensed indicators and open-access biodiversity data to assess bird diversity patterns in Mediterranean rural landscapes. Sci. Rep. 2019, 9, 6826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanderWerf, E.A. Demography of hawai’i ‘elepaio: Variation with habitat disturbance and population density. Ecology 2004, 85, 770–783. [Google Scholar] [CrossRef]
- Belisle, M.; Desrochers, A.; Fortin, M.J. Influence of forest cover on the movements of forest birds: A homing experiment. Ecology 2001, 82, 1893–1904. [Google Scholar] [CrossRef]
- Partridge, D.R.; Clark, J.A. Urban green roofs provide habitat for migrating and breeding birds and their arthropod prey. PLoS ONE 2018, 13, e0202298. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.; Soffianian, A.R.; Koupaei, S.S.; Pourmanafi, S.; Saatchi, S. Wetland restoration prioritizing, a tool to reduce negative effects of drought; An application of multicriteria-spatial decision support system (MC-SDSS). Ecol. Eng. 2018, 112, 132–139. [Google Scholar] [CrossRef]
- Li, D.; Chen, S.; Lloyd, H.; Zhu, S.; Shan, K.; Zhang, Z. The importance of artificial habitats to migratory waterbirds within a natural/artificial wetland mosaic, Yellow River Delta, China. Bird Conserv. Int. 2013, 23, 184–198. [Google Scholar] [CrossRef] [Green Version]
- Shreeve, T.G.; Dennis, R.L.H. Landscape scale conservation: Resources, behaviour, the matrix and opportunities. Lepid. Conserv. Chang. World 2010, 15, 261–270. [Google Scholar] [CrossRef]
- Xie, Y.; Yu, X.; Ng, N.C.; Li, K.; Fang, L. Exploring the dynamic correlation of landscape composition and habitat fragmentation with surface water quality in the Shenzhen river and deep bay cross-border watershed, China. Ecol. Indic. 2018, 90, 231–246. [Google Scholar] [CrossRef]
- Pfeifer, M.; Boyle, M.J.; Dunning, S.; Olivier, P.I. Forest floor temperature and greenness link significantly to canopy attributes in South Africa’s fragmented coastal forests. PeerJ. 2019, 7, e6190. [Google Scholar] [CrossRef]
- Calderon, M.R.; Almeida, C.A.; González, P.; Jofré, M.B. Influence of water quality and habitat conditions on amphibian community metrics in rivers affected by urban activity. Urban Ecosyst. 2019, 22, 743–755. [Google Scholar] [CrossRef]
- Daw, T.M.; Hicks, C.C.; Brown, K.; Chaigneau, T.; Januchowski-Hartley, F.A.; Cheung, W.W.L.; Rosendo, S.; Crona, B.; Coulthard, S.; Sandbrook, C.; et al. Elasticity in ecosystem services: Exploring the variable relationship between ecosystems and human well-being. Ecol. Soc. 2016, 21, 11–13. [Google Scholar] [CrossRef]
- Buschke, F.T.; Esterhuyse, S.; Kemp, M.E.; Seaman, M.T.; Brendonck, L.; Vanschoenwinkel, B. The dynamics of mountain rock pools—Are aquatic and terrestrial habitats alternative stable states? Acta Oecologica 2013, 47, 24–29. [Google Scholar] [CrossRef]
- Boesing, A.L.; Nichols, E.; Metzger, J.P. Biodiversity extinction thresholds are modulated by matrix type. Ecography 2018, 41, 1520–1533. [Google Scholar] [CrossRef] [Green Version]
- Fahrig, L. How much habitat is enough? Biol. Conserv. 2001, 100, 65–74. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The Population Biology of Invasive Species. Annu. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.A.E.; Si, X.; Smith, Y.C.E.; Kalle, R.; Ramesh, T.; Downs, C.T. Patterns of avian diversity across a decreasing patch-size gradient in a critically endangered subtropical forest system. J. Biogeogr. 2018, 45, 2118–2132. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Sun, W.; Xia, F.; Yang, Y.; Xie, Y. Can Habitat Quality Index Measured Using the InVEST Model Explain Variations in Bird Diversity in an Urban Area? Sustainability 2021, 13, 5747. https://doi.org/10.3390/su13105747
Li D, Sun W, Xia F, Yang Y, Xie Y. Can Habitat Quality Index Measured Using the InVEST Model Explain Variations in Bird Diversity in an Urban Area? Sustainability. 2021; 13(10):5747. https://doi.org/10.3390/su13105747
Chicago/Turabian StyleLi, Dehuan, Wei Sun, Fan Xia, Yixuan Yang, and Yujing Xie. 2021. "Can Habitat Quality Index Measured Using the InVEST Model Explain Variations in Bird Diversity in an Urban Area?" Sustainability 13, no. 10: 5747. https://doi.org/10.3390/su13105747