Do Silvi-Medicinal Plantations Affect Tree Litter Decomposition and Nutrient Mineralization?

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Litter and Soil Sampling
2.2. Litter Decomposition
2.3. Litter Chemical Analyses and Enzyme Activities
2.4. Data Processing
3. Results
3.1. Effects on the Decomposition of Conifer Litters
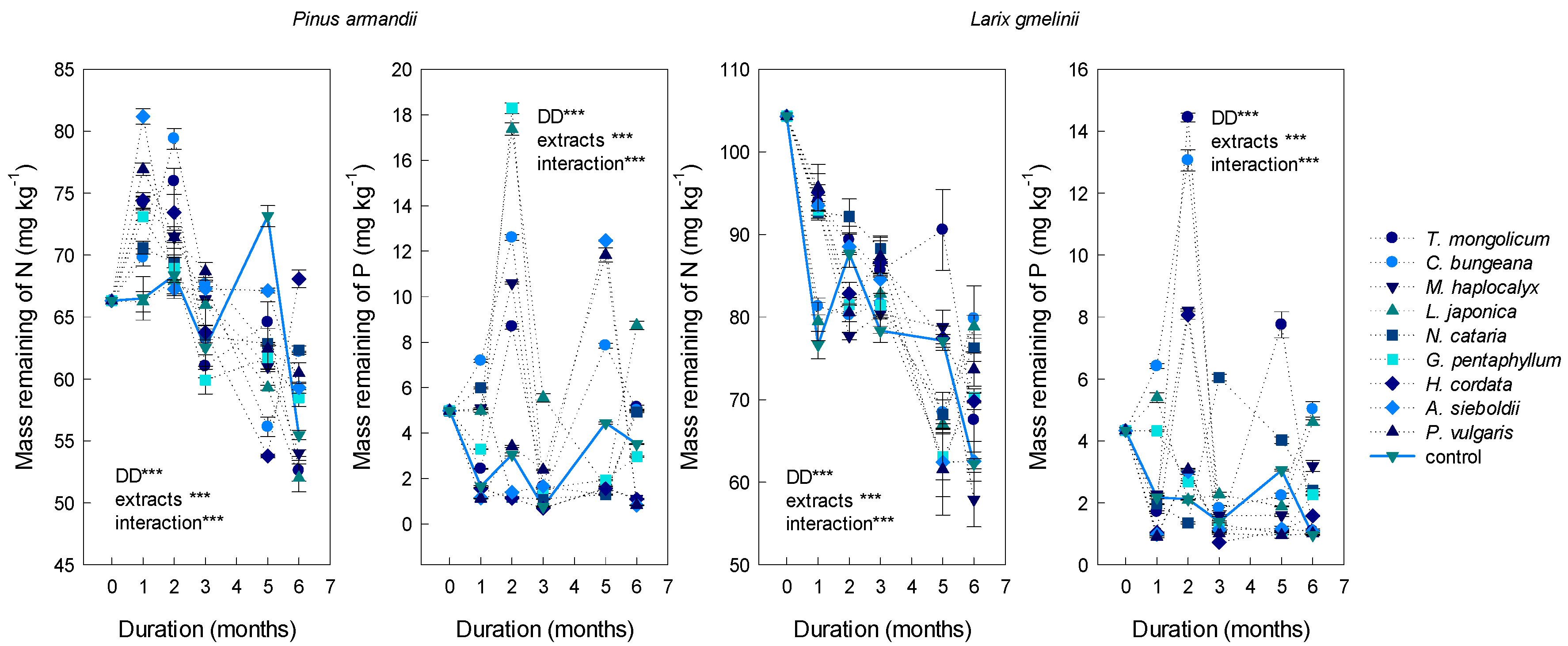
3.2. Effects on the N and P Release of Conifer Litters
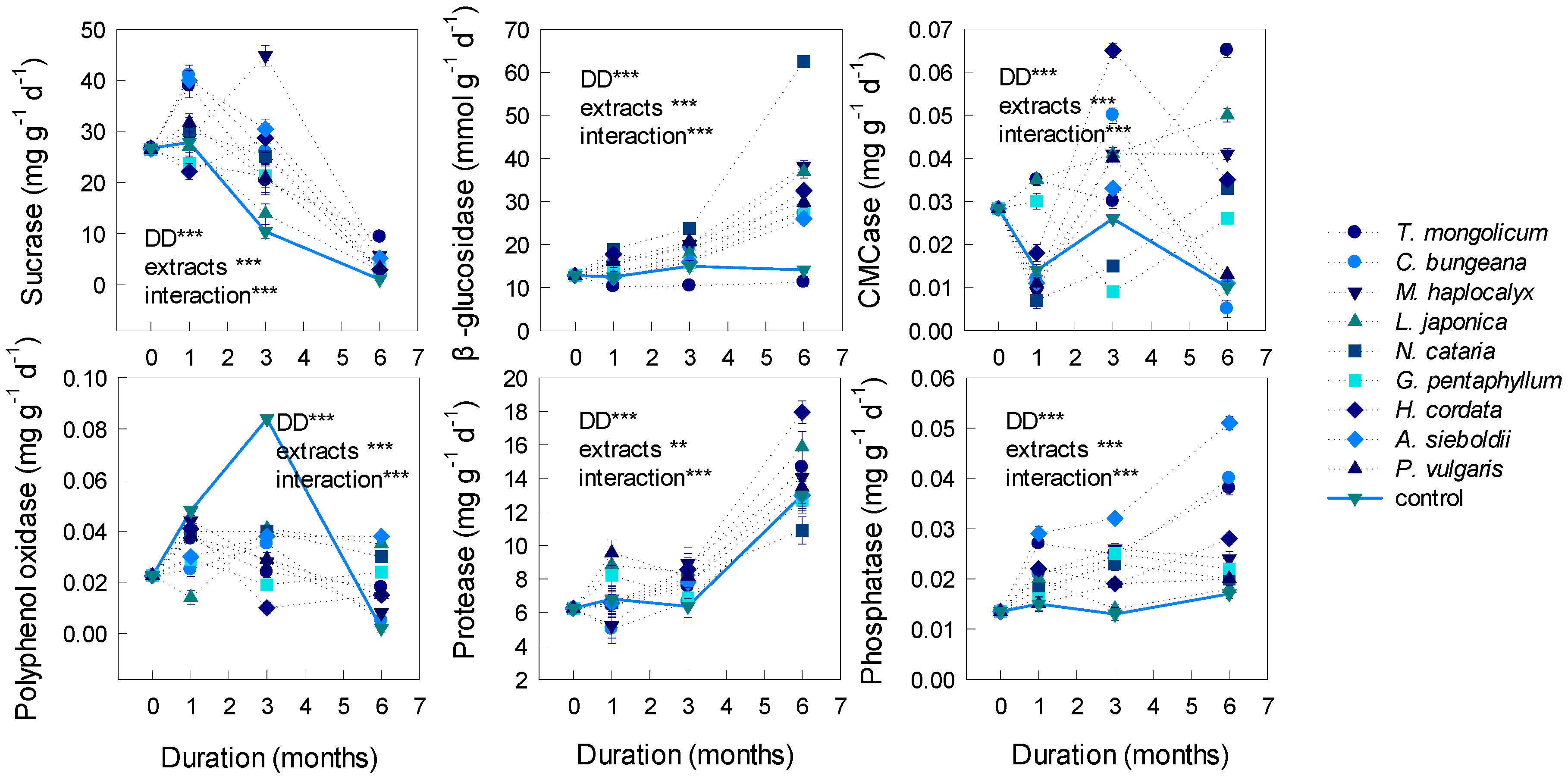
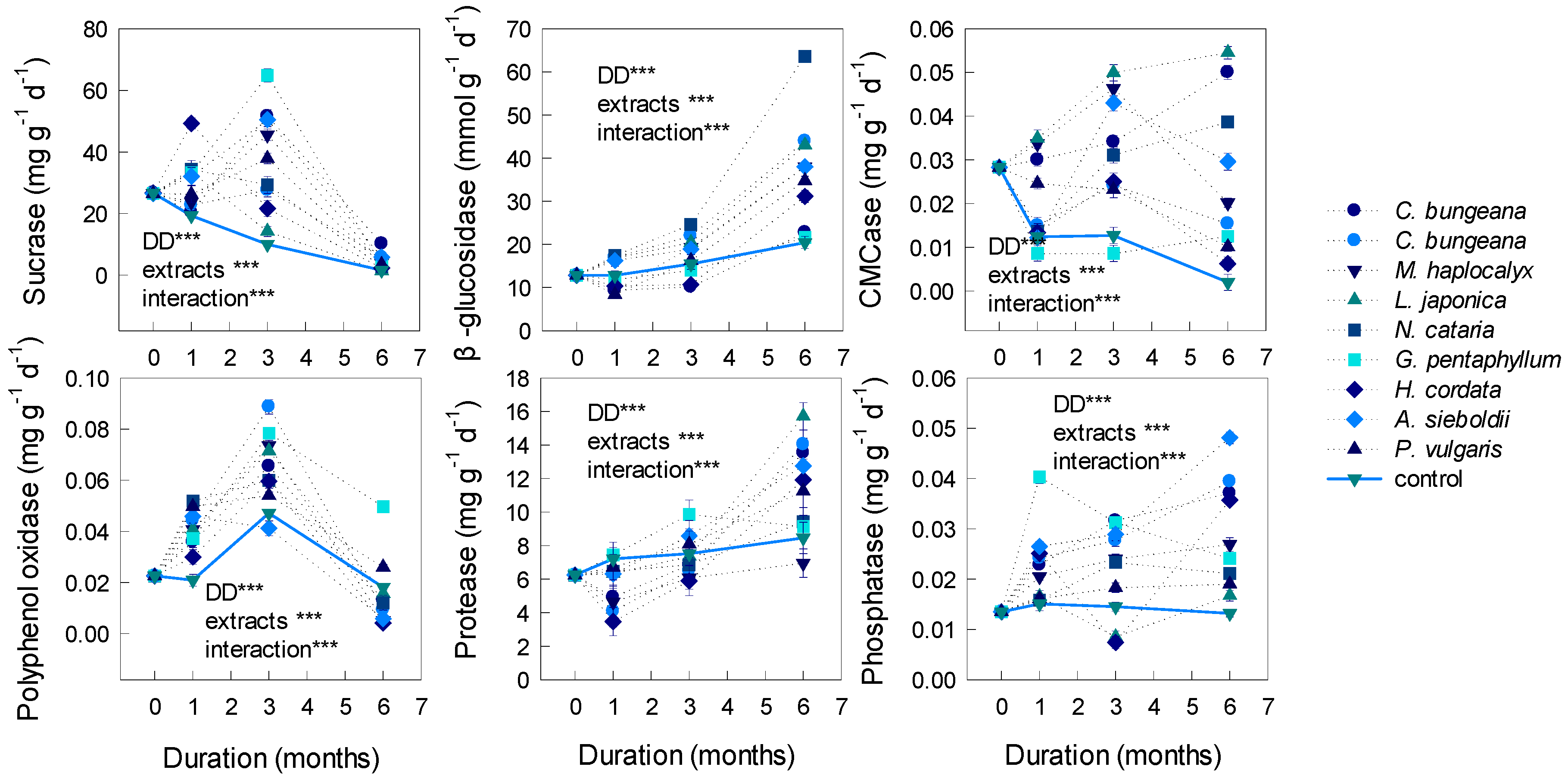
3.3. Effects on Soil Enzymatic Activities
4. Discussion
4.1. Litter Decomposition
4.2. Nutrient Release
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chao, J.; Dai, Y.; Verpoorte, R.; Lam, W.; Cheng, Y.-C.; Pao, L.-H.; Zhang, W.; Chen, S. Major achievements of evidence-based traditional Chinese medicine in treating major diseases. Biochem. Pharmacol. 2017, 139, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Normile, D. The new face of traditional Chinese medicine. Science 2003, 299, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sasse, A.; Sheridan, H. Traditional Chinese Medicine: From Aqueous Extracts to Therapeutic Formulae. In Plant Extracts; IntechOpen: Rijeka, Croatia, 2019. [Google Scholar] [Green Version]
- Duke, J.A. Duke’s Handbook of Medicinal Plants of Latin America; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Li, T.S. Chinese & Related North American Herbs: Phytopharmacology & Therapeutic Values, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Wiśniewski, J.; Szczepanik, M.; Kołodziej, B.; Król, B. Plantation methods effects on common valerian (Valeriana officinalis) yield and quality. J. Anim. Plant Sci. 2016, 26, 177–184. [Google Scholar]
- Hu, L.; Zhang, Z.; Wang, W.; Wang, L.; Yang, H. Ginseng plantations threaten China’s forests. Biodivers Conserv. 2018, 27, 2093–2095. [Google Scholar] [CrossRef]
- Li, Y.; Wang, B.; Chang, Y.; Yang, Y.; Yao, C.; Huang, X.; Zhang, J.; Cai, Z.; Zhao, J. Reductive soil disinfestation effectively alleviates the replant failure of Sanqi ginseng through allelochemical degradation and pathogen suppression. Appl. Microbiol. Biotechnol. 2019, 103, 3581–3595. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Xu, J.; Feng, G.; Li, X.; Chen, S. Soil bacterial and fungal community dynamics in relation to Panax notoginseng death rate in a continuous cropping system. Sci. Rep. 2016, 6, 31802. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, S.; Li, B.; Ren, X. Evaluation of important medicinal plant resources in Qinling Mountains based on analytic hierarchy process method. Chin. Traditional. Herb. Drug 2013, 44, 2172–2182. [Google Scholar]
- Zhang, X.; Liu, H.; Wang, B.; Li, J.; Lei, H.; Liu, Z. Characteristics of the mixed decomposition of fresh litter of Picea asperata and broadleaved species. Ecol. Environ. Sci. 2019, 28, 235–244. [Google Scholar]
- Sanwal, C.S.; Kumar, R.; Anwar, R.; Kakade, V.; Kerketta, S.; Bhardwaj, S. Growth and yield of Solanum khasianum in Pinus roxburghii forest based silvi-medicinal system in mid hills of Indian Himalaya. For. Ecosyst. 2016, 3, 19–27. [Google Scholar] [CrossRef]
- Song, X.; Jiang, H.; Yu, S.; Ma, Y.; Zhou, G.; Dou, R.; Guo, P. Litter decomposition of dominant plant species in successional stages in mid-subtropical zone. Chin. J. Appl. Ecol. 2009, 20, 537–542. [Google Scholar]
- Adamczyk, S.; Adamczyk, B.; Kitunen, V.; Smolander, A. Monoterpenes and higher terpenes may inhibit enzyme activities in boreal forest soil. Soil. Biol. Biochem. 2015, 87, 59–66. [Google Scholar] [CrossRef]
- Pitarokili, D.; Couladis, M.; Petsikos-Panayotarou, N.; Tzakou, O. Composition and antifungal activity on soil-borne pathogens of the essential oil of Salvia sclarea from Greece. J. Agric. Food Chem. 2002, 50, 6688–6691. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Qiao, M.; Li, D.; Zhao, C.; Li, Y.; Yin, H.; Liu, Q. Effects of two root-secreted phenolic compounds from a subalpine conifer species on soil enzyme activity and microbial biomass. Chem. Ecol. 2015, 31, 636–649. [Google Scholar] [CrossRef]
- Zhou, B.; Kong, C.-H.; Li, Y.-H.; Wang, P.; Xu, X.-H. Crabgrass (Digitaria sanguinalis) allelochemicals that interfere with crop growth and the soil microbial community. J. Agric. Food Chem. 2013, 61, 5310–5317. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ying, Y.; Zhao, D.; Ding, W. Influence of allelochemicals on microbial community in ginseng cultivating soil. Chin. Herb. Med. 2014, 6, 313–318. [Google Scholar] [CrossRef]
- Kong, C.; Wang, P.; Zhao, H.; Xu, X.; Zhu, Y. Impact of allelochemical exuded from allelopathic rice on soil microbial community. Soil Biol. Biochem. 2008, 40, 1862–1869. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Jørgensen, H.B. Carbon quality rather than stoichiometry controls litter decomposition in a tropical rain forest. J. Ecol. 2010, 98, 754–763. [Google Scholar] [CrossRef]
- Medvedeff, C.A.; Bridgham, S.D.; Pfeifer-Meister, L.; Keller, J.K. Can Sphagnum leachate chemistry explain differences in anaerobic decomposition in peatlands? Soil Biol. Biochem. 2015, 86, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Ushio, M.; Balser, T.C.; Kitayama, K. Effects of condensed tannins in conifer leaves on the composition and activity of the soil microbial community in a tropical montane forest. Plant Soil 2013, 365, 157–170. [Google Scholar] [CrossRef]
- Triebwasser, D.J.; Tharayil, N.; Preston, C.M.; Gerard, P.D. The susceptibility of soil enzymes to inhibition by leaf litter tannins is dependent on the tannin chemistry, enzyme class and vegetation history. New Phytol. 2012, 196, 1122–1132. [Google Scholar] [CrossRef]
- Adamczyk, S.; Kiikkilä, O.; Kitunen, V.; Smolander, A. Potential response of soil processes to diterpenes, triterpenes and tannins: Nitrification, growth of microorganisms and precipitation of proteins. Appl. Soil Ecol. 2013, 67, 47–52. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, W.; Liu, Z. Soil Response of nutrient release of Periploca sepium litter to soil petroleum contamination. Clean Soil Air Water 2016, 44, 1709–1716. [Google Scholar] [CrossRef]
- Lin, H.; Zhao, Y.; Muyidong, N.; Tian, K.; He, Z.; Kong, X.; Sun, S.; Tian, X. Secondary compounds of Pinus massoniana alter decomposers’ effects on Quercus variabilis litter decomposition. Ecol. Evol. 2018, 8, 9439–9450. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, B.; Karonen, M.; Adamczyk, S.; Engström, M.T.; Laakso, T.; Saranpää, P.; Kitunen, V.; Smolander, A.; Simon, J. Tannins can slow-down but also speed-up soil enzymatic activity in boreal forest. Soil Biol. Biochem. 2017, 107, 60–67. [Google Scholar] [CrossRef]
- Aggarwal, K.; Khanuja, S.; Ahmad, A.; Santha Kumar, T.; Gupta, V.K.; Kumar, S. Antimicrobial activity profiles of the two enantiomers of limonene and carvone isolated from the oils of Mentha spicata and Anethum sowa. Flavour Fragr. J. 2002, 17, 59–63. [Google Scholar] [CrossRef]
- Yang, S.; Jiang, D.; Fan, G.; Zhang, Q.; Li, M.; Cheng, Y.; Peng, L. Antifungal activity against citrus blue mold and its active constituents of solvent extracts from pine needles. Mod. Food Sci. Technol. 2016, 32, 65–69. [Google Scholar]
- Chomel, M.; Guittonny-Larchevêque, M.; Fernandez, C.; Gallet, C.; DesRochers, A.; Paré, D.; Jackson, B.G.; Baldy, V. Plant secondary metabolites: A key driver of litter decomposition and soil nutrient cycling. J. Ecol. 2016, 104, 1527–1541. [Google Scholar] [CrossRef]
- Del Giudice, R.D.; Lindo, Z. Short-term leaching dynamics of three peatland plant species reveals how shifts in plant communities may affect decomposition processes. Geoderma 2017, 285, 110–116. [Google Scholar] [CrossRef]
- Wei, C.; Liu, X.; Lin, C.; Li, X.; Li, Y.; Zheng, Y. Response of soil enzyme activities to litter input changes in two secondary Castanopsis carlessii forests in subtropical China. Chin. J. Plant Ecol. 2018, 42, 692–702. [Google Scholar]
- Li, C. Effect of exogenous allelochemical vanillic acid on soil enzyme activity and soil nutrient for pepper seedling. China Veg. 2009, 2009, 46–49. [Google Scholar]
- Chen, W.; Vermaak, I.; Viljoen, A. Camphor-a fumigant during the black death and a coveted fragrant wood in ancient Egypt and Babylon-a review. Molecules 2013, 18, 5434–5454. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.; Luan, D.; Wang, S.; Zhao, L.; Tao, L.; Yuan, Q.; Wang, X. Borneol-grafted cellulose for antifungal adhesion and fungal growth inhibition. RSC Adv. 2015, 5, 51947–51952. [Google Scholar] [CrossRef]
- Cho, W.-I.; Cheigh, C.-I.; Hwang, H.-J.; Chung, M.-S. Sporicidal activities of various surfactant components against Bacillus subtilis spores. J. Food Prot. 2015, 78, 1221–1225. [Google Scholar] [CrossRef] [PubMed]
- Pattnaik, S.; Subramanyam, V.; Bapaji, M.; Kole, C. Antibacterial and antifungal activity of aromatic constituents of essential oils. Microbios 1997, 89, 39–46. [Google Scholar] [PubMed]
- Mancini, A.; Pucciarelli, S. Antibiotic activity of the antioxidant drink effective Microorganism-X (EM-X) extracts against common nosocomial pathogens: An in vitro study. Nat. Prod. Res. 2018, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Li, Q. Research on Chemical Composition and Decay Resistance Mechanism of Cinnamomum camphora Wood Extracts. Ph.D. Thesis, Fujian Agricultural and Forestry University, Fuzhou, China, 2014. [Google Scholar]
- Skočibušić, M.; Bezić, N.; Dunkić, V. Phytochemical composition and antimicrobial activities of the essential oils from Satureja subspicata Vis. growing in Croatia. Food Chem. 2006, 96, 20–28. [Google Scholar] [CrossRef]
- Liu, P.; Liu, Z.; Wang, C.; Guo, F.; Wang, M.; Zhang, Y.; Dong, L.; Wan, S. Effects of three long-chain fatty acids present in peanut (Arachis hypogaea L.) root exudates on its own growth and the soil enzymes activities. Allelopath. J. 2012, 29, 13–24. [Google Scholar]
- Guan, Z.; Luo, Q.; Chen, X.; Feng, X.; Tang, Z.; Wei, W.; Zheng, Y. Saline soil enzyme activities of four plant communities in Sangong River basin of Xinjiang, China. J. Arid Land 2014, 6, 164–173. [Google Scholar] [CrossRef]
- Yang, K.; Zhu, J.J. Impact of tree litter decomposition on soil biochemical properties obtained from a temperate secondary forest in Northeast China. J. Soils Sediments 2015, 15, 13–23. [Google Scholar] [CrossRef]
- Berger, T.W.; Olivier, D.; Ika, D.; Michael, T.; Gerzabek, M.H.; Franz, Z. Decomposition of beech (Fagus sylvatica) and pine (Pinus nigra) litter along an Alpine elevation gradient: Decay and nutrient release. Geoderma 2015, 251, 92–104. [Google Scholar] [CrossRef]
- Schimel, J.P.; Hättenschwiler, S. Nitrogen transfer between decomposing leaves of different N status. Soil Biol. Biochem. 2007, 39, 1428–1436. [Google Scholar] [CrossRef]
- Smolander, A.; Kanerva, S.; Adamczyk, B.; Kitunen, V. Nitrogen transformations in boreal forest soils—Does composition of plant secondary compounds give any explanations? Plant Soil 2012, 350, 1–26. [Google Scholar] [CrossRef]
- Lu, Y.; Xu, J.; Zhang, X.; Wang, B.; Xie, B.; Liu, Z. Effects of leachate from understory medicinal plants on litter decomposition and soil enzyme activities of Betula albo-sinensis and Eucommia ulmoides. Chin. J. Plant Ecol. 2017, 41, 639–649. [Google Scholar]
- Yang, Q.; Li, R.; Zhang, W.; Zheng, W.; Wang, Q.; Chen, L.; Guan, X.; Xu, M.; Wang, S. Decomposition of harvest residue needles of different needle ages in a Chinese fir (Cunninghamia lanceolata) plantation. Plant Soil 2018, 423, 273–284. [Google Scholar] [CrossRef]
- Shao, X.; Cheng, S.; Wang, H.; Yu, D.; Mungai, C. The possible mechanism of antifungal action of tea tree oil on Botrytis cinerea. J. Appl. Microbiol. 2013, 114, 1642–1649. [Google Scholar] [CrossRef] [PubMed]
- Cox, S.; Mann, C.; Markham, J.; Bell, H.; Gustafson, J.; Warmington, J.; Wyllie, S. The mode of antimicrobial action of the essential oil of Melaleuca alternifolia (tea tree oil). J. Appl. Microbiol. 2000, 88, 170–175. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Needle Litters | Concentration (mg·g−1) | C/N | C/P | Lignin/N | |||
|---|---|---|---|---|---|---|---|
| C | N | P | Lignin | ||||
| P. armandii | 446.46 ± 10.56 | 11.05 ± 0.83 * | 0.83 ± 0.01 * | 264.66 ± 5.38 * | 40.40 * | 538.5 * | 23.95 * |
| L. gmelinii | 455.63 ± 8.95 | 20.86 ± 0.45 | 0.72 ± 0.01 | 198.51 ± 3.67 | 21.84 | 631.07 | 9.52 |
| Extracts of Medicinal Herbs | Detected Metabolites * | N Concentration (mg L−1) | P Concentration (mg L−1) |
|---|---|---|---|
| T. mongolicum | 24,25-Dihydroxycholecalciferol; Caffeic acid; Camphor; Caryophyllene; Chlorogenic acid; Mignonette; Quercetin; | 24.53 ± 1.13 | 2.72 ± 0.12 |
| C. bungeana | 2,3-Dihydrobenzofuran; 3-Oxygen-β-ionone; 4-(3-Hydroxybutyl)-3,5,5-trimethyl-2-cyclohexene-1-ketone; 4-Hydroxy-3,5,6-trimethyl-4-(3-oxo-1-butenyl)-2-cyclohexene-1-ketone; Coniferyl alcohol; Linalool; β-Pinene; β-Sitosterol | 23.58 ± 1.49 | 1.14 ± 0.97 |
| M. haplocalyx | (-)-Carvone; (+)-Limonene; Caryophyllene oxide; Caryophyllene; DL-menthol acetate; L-menthol; Menthol; Myrcene; Neodecanoic acid; Pulegone; Pyridazine; α-Cadinol; α-Pinene; β-Pinene | 13.80 ± 0.95 | 0.31 ± 0.02 |
| H. cordata | (2S,3S)-2-Amino-3-methylpentanoic acid; Aporphines; Artemisinin; Caryophyllene; Chlorogenic acid; Cineole; DL-Limonene; Hyperoside; Isorhamnetin; Kaempferol-3-O-β-D-glucoside; L-glutamic acid; Linalool; Linoleic acid; Methyl-nonylketone; Myrcene; Ocimene; Palmitic acid; Pyridine; Quercetin; Rutin; Stearic acid; α-Pinene; β-Sitosterol | 22.19 ± 1.35 | 2.28 ± 0.74 |
| A. sieboldii | 3,4,5-Trimethoxytoluene; 3,5-Dimethoxytoluene; Elemicin; Methyl eugenol; Myristicin; Safrole | 15.81 ± 1.17 | 2.40 ± 0.59 |
| L. japonica | 1,6-Anhydro-β-D-glucopyranose; 2-Vinylpyridine; 3-tert-Butyl-4-hydroxyanisole; Allose; Benzyl alcohol; Benzyl benzoate; Borneol; Limonene; Linalool; Palmitic acid; Phenethyl alcohol; Phenylacetaldehyde; | 25.40 ± 0.93 | 2.64 ± 0.34 |
| N. cataria | (+)-Dipentene; 1-Octen-3-ol; 1-Octen-3-yl acetate; Caryophyllene oxide; Isomenthone; Menthenone; Menthone; Menthone; Palmitic acid; Pyridazine; β-Sitosterol | 10.12 ± 0.67 | 0.54 ± 0.02 |
| G. pentaphyllum | Arachidic Acid; Benzaldehyde; Diisobutyl phthalate; Dodecenylsuccinic acid; Fitone; Geranylacetone; Linalool; Naphthalene; Palmitic acid; Pentadecanoic acid; β-Sitosterol | 12.56 ± 1.69 | 0.71 ± 0.04 |
| P. vulgaris | 1,8-Cineole; 7-Hydroxycoumarin; Caffeic acid; Lauric acid; Linalool; Linalyl acetate; Luteolin; Methyl oleanolate; Myrcene; Palmitic acid; α- Phellandrene; β-Pinene; β-Sitosterol | 38.69 ± 2.15 | 8.35 ± 1.13 |
| Medicinal Herb Extract | Litter Decomposition Model | T0.95 (yr) | T0.50 (yr) |
|---|---|---|---|
| T. mongolicum | R = 0.29e−8.86t + 0.71e−0.22t | 19.90 ± 2.58 | 2.67 ± 0.60 |
| C. bungeana | R = 0.29e−9.36t + 0.71e−0.22t | 13.16 ± 2.27 | 1.71 ± 0.42 * |
| M. haplocalyx | R = 0.17e−11.26t + 0.83e−0.46t | 13.34 ± 2.19 | 1.74 ± 0.41 * |
| H. cordata | R = 0.22e−17.88t + 0.78e−0.49t | 7.78 ± 1.79 * | 1.24 ± 0.22 ** |
| A. sieboldii | R = 0.22e−9.84t + 0.78e−0.28t | 9.16 ± 1.82 * | 1.44 ± 0.37 ** |
| L. japonica | R = 0.30e−10.16t + 0.71e−0.11t | 10.79 ± 1.82 | 1.73 ± 0.38 * |
| N. cataria | R = 0.28e−13.07t + 0.72e−0.13t | 23.80 ± 0.76 ** | 3.07 ± 0.12 |
| G. pentaphyllum | R = 0.24e−10.78t + 0.75e−0.33t | 21.18 ± 1.42 * | 2.90 ± 0.32 |
| P. vulgaris | R = 0.19e−49.73t + 0.81e−0.25t | 9.82 ± 2.59 | 1.44±0.52 ** |
| Control | R = 0.28e−10.13t + 0.72e−0.16t | 15.17 ± 2.06 | 2.51 ± 0.50 |
| Medicinal Herb Extract | Litter Decomposition Model | T0.95 (yr) | T0.50 (yr) |
|---|---|---|---|
| T. mongolicum | R = 0.28e−22.18t + 0.72e−0.55t | 6.52 ± 1.12 | 0.72 ± 0.08 |
| C. bungeana | R = 0.36e−20.32t + 0.63e−0.30t | 5.35 ± 0.30 | 0.73 ± 0.05 |
| M. haplocalyx | R = 0.36e−18.21t + 0.64e−0.21t | 9.28 ± 1.88 | 0.85 ± 0.15 |
| H. cordata | R = 0.26e−34.67t + 0.74e−0.56t | 12.17 ± 0.90 * | 1.16 ± 0.12 ** |
| A. sieboldii | R = 0.27e−19.51t + 0.73e−0.78t | 4.91 ± 0.47 | 0.72 ± 0.05 |
| L. japonica | R = 0.40e−13.13t + 0.60e−0.22t | 3.52 ± 0.31 | 0.48 ± 0.02 |
| N. cataria | R = 0.22e−4284.04t + 0.79e−0.95t | 12.56 ± 2.31 ** | 0.91 ± 0.15 |
| G. pentaphyllum | R = 0.31e−19.94t + 0.69e−0.50t | 2.95 ± 0.26 * | 0.48 ± 0.03 |
| P. vulgaris | R = 0.32e−17.63t + 0.68e−0.42t | 6.03 ± 1.16 | 0.68 ± 0.11 |
| Control | R = 0.34e−14.85t + 0.66e−0.38t | 6.37 ± 0.28 | 0.74 ± 0.03 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Lei, H.; Chong, Y.; Hu, J.; Che, W.; Hu, M.; Xu, S.; Zhang, P.; Zhang, L.; Xu, J.; et al. Do Silvi-Medicinal Plantations Affect Tree Litter Decomposition and Nutrient Mineralization? Sustainability 2019, 11, 5138. https://doi.org/10.3390/su11185138
Zhang X, Lei H, Chong Y, Hu J, Che W, Hu M, Xu S, Zhang P, Zhang L, Xu J, et al. Do Silvi-Medicinal Plantations Affect Tree Litter Decomposition and Nutrient Mineralization? Sustainability. 2019; 11(18):5138. https://doi.org/10.3390/su11185138
Chicago/Turabian StyleZhang, Xiaoxi, Hangyu Lei, Yujie Chong, Jiawei Hu, Wenrong Che, Man Hu, Shuoyu Xu, Pan Zhang, Lingling Zhang, Jiyuan Xu, and et al. 2019. "Do Silvi-Medicinal Plantations Affect Tree Litter Decomposition and Nutrient Mineralization?" Sustainability 11, no. 18: 5138. https://doi.org/10.3390/su11185138