Effect of Growing Miscanthus x giganteus on Soil Microbial Communities in Post-Military Soil

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment
2.2. Sampling
2.3. Analyses
2.4. Statistics
3. Results and Discussion
3.1. Soil Nutrients
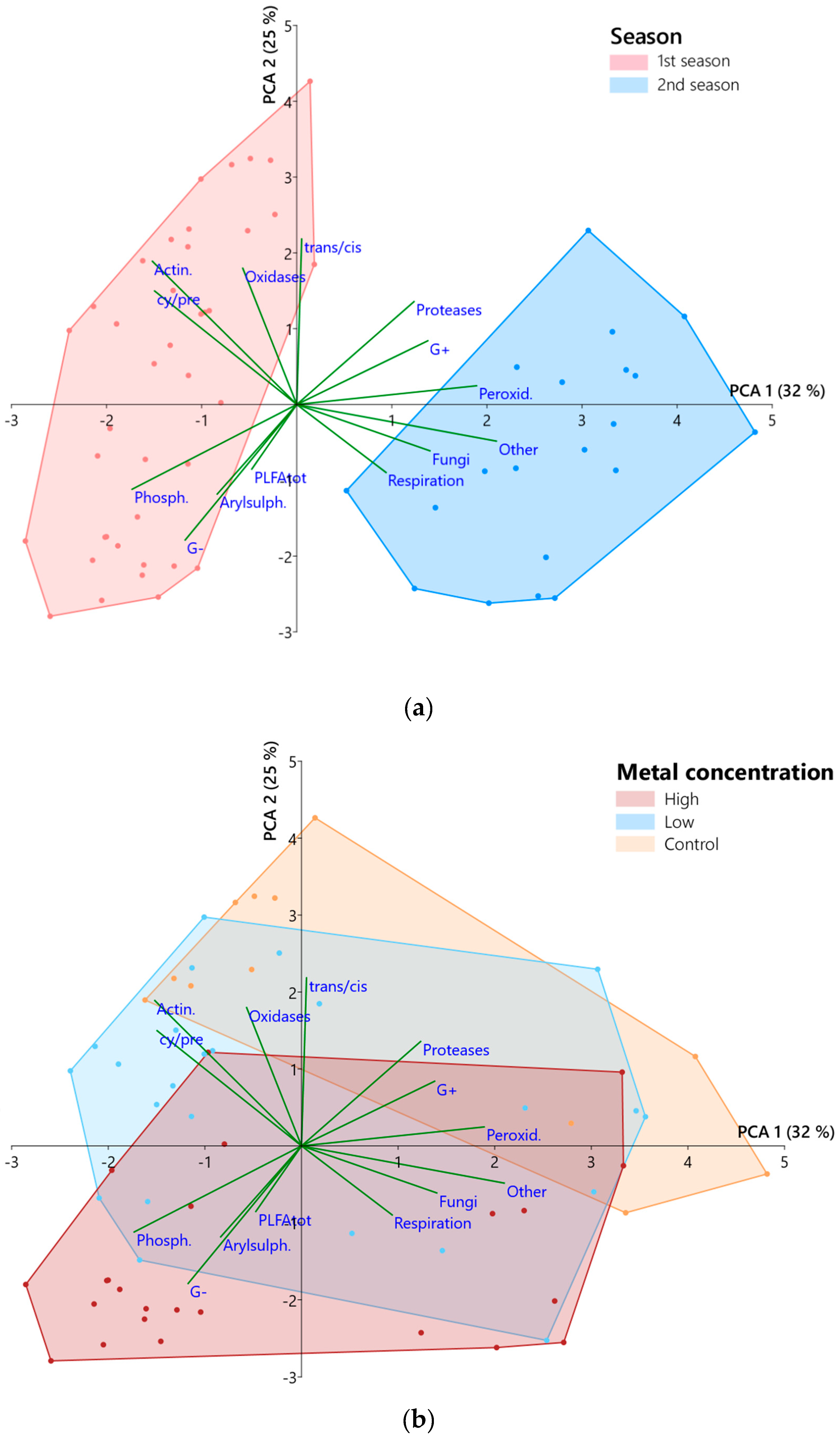
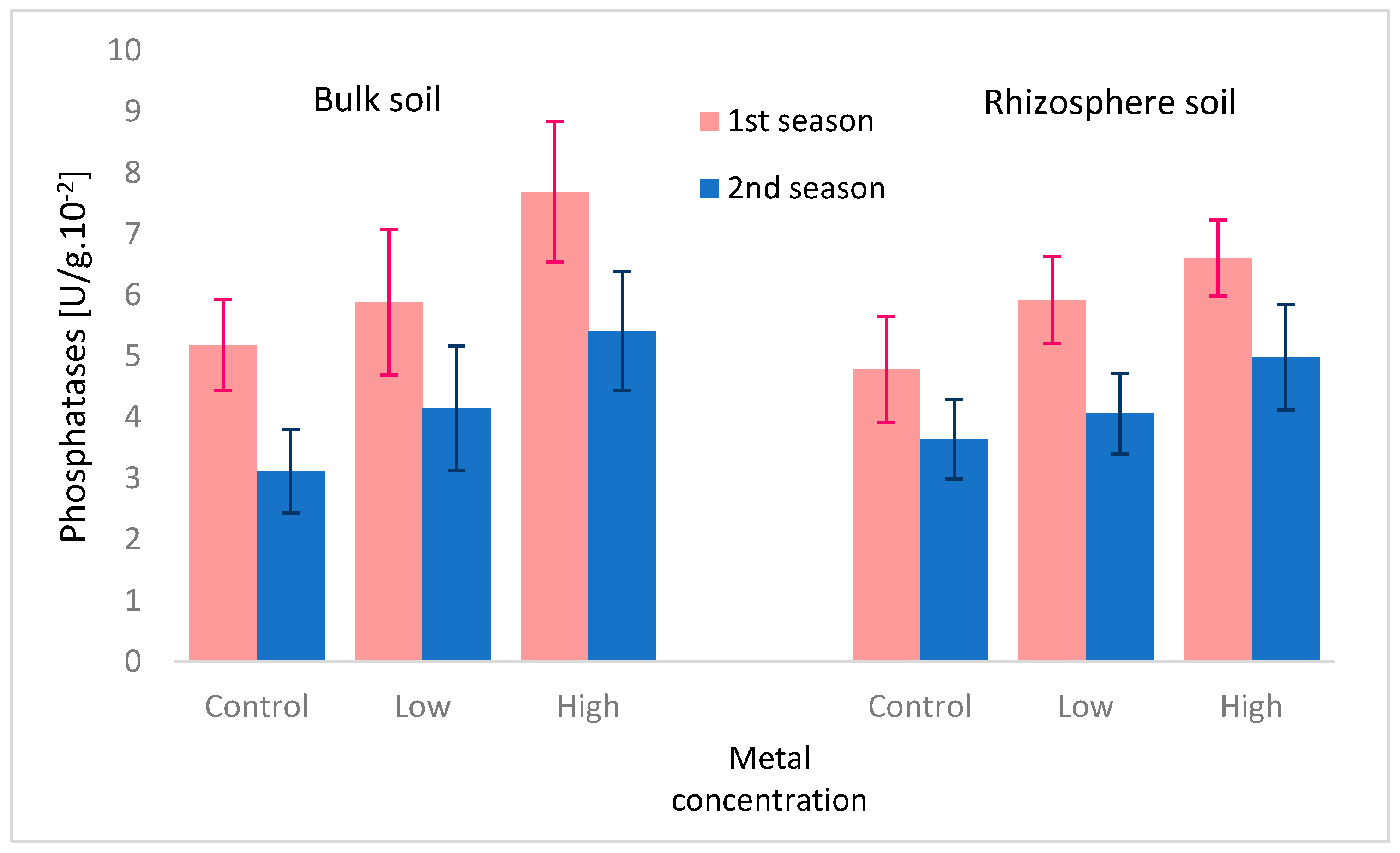
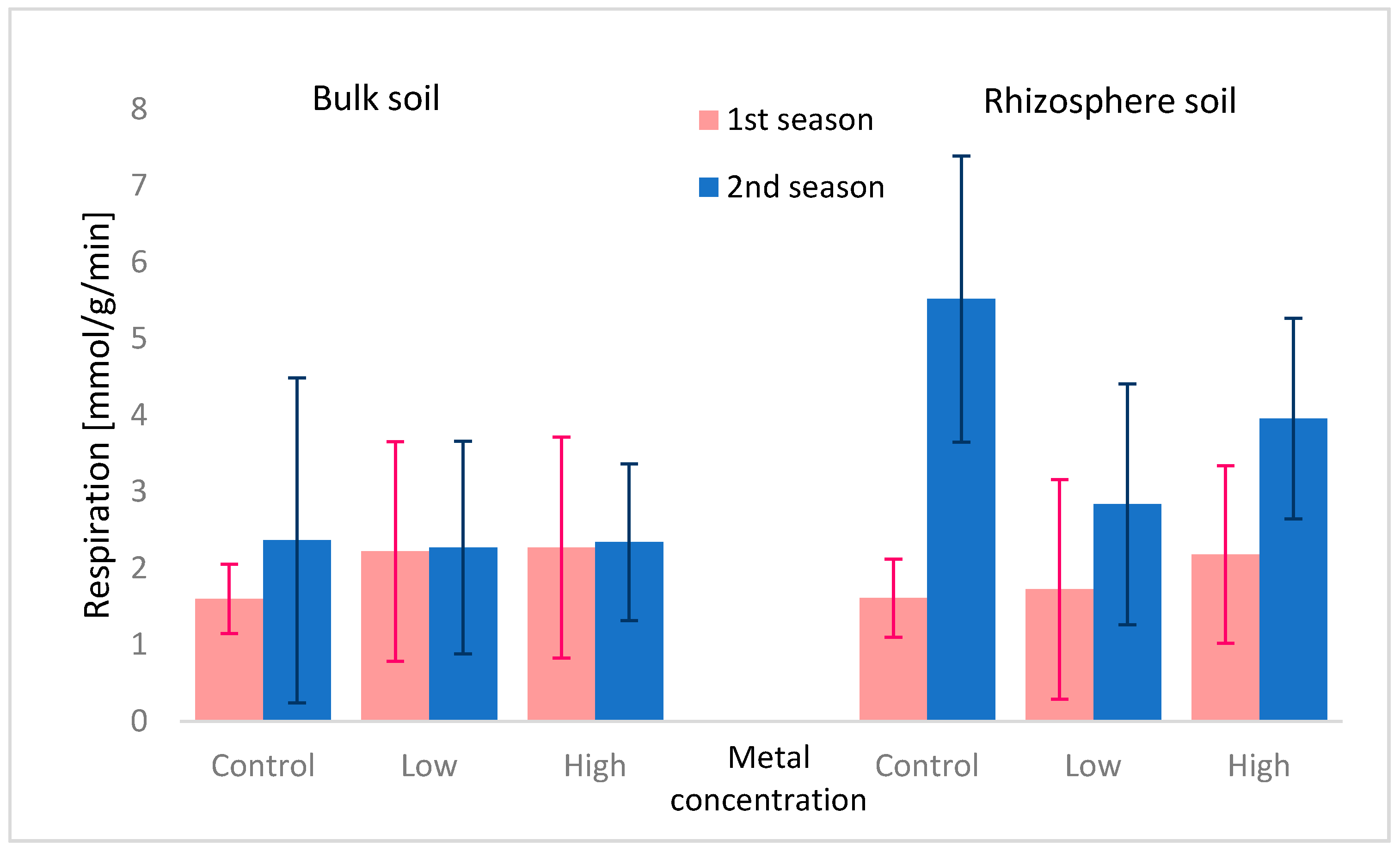
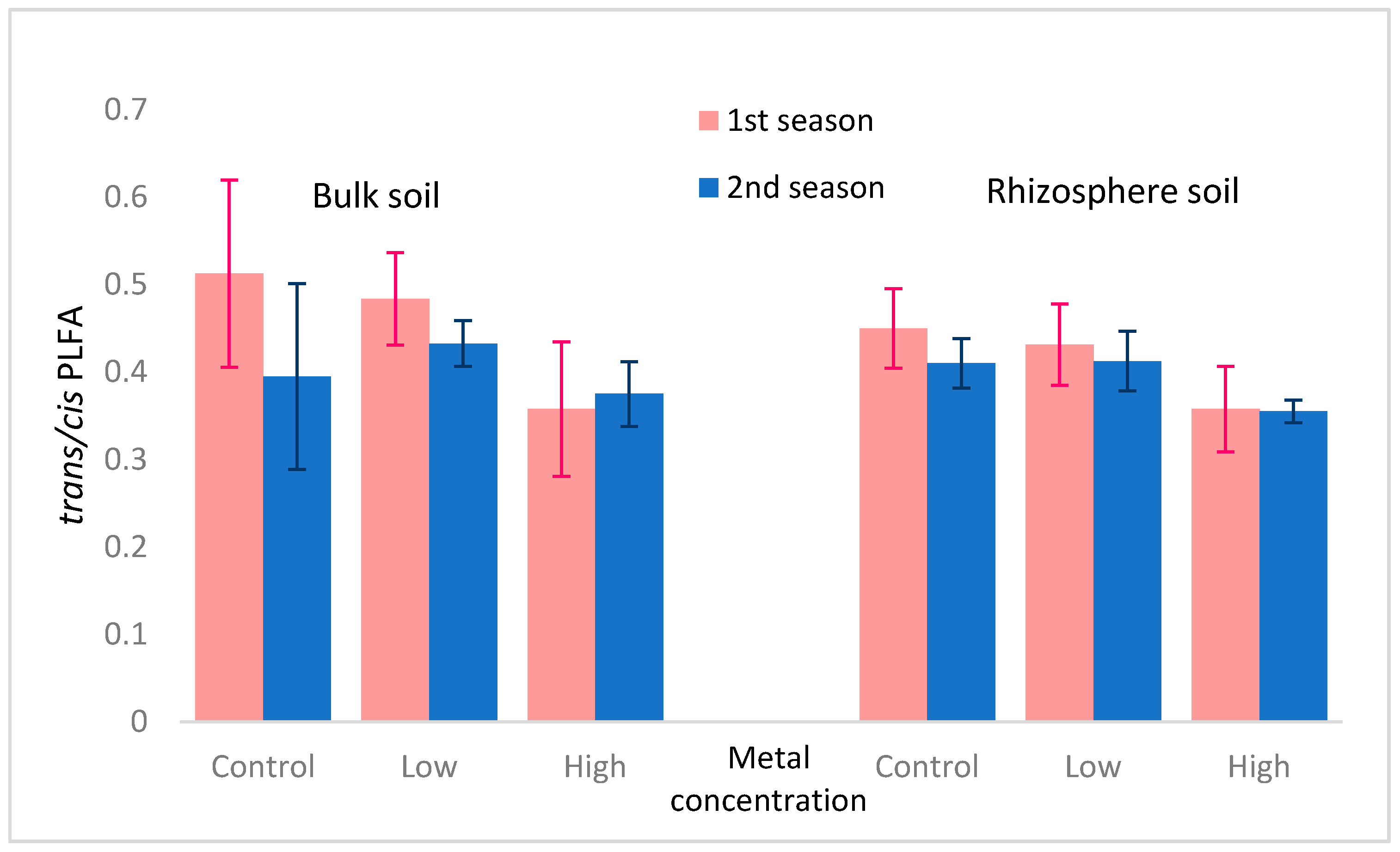
3.2. Microbial Community
3.3. Implications for Sustainable Development
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Appendix A. Explanation of the Physiological Indicators
References
- Fonnum, F.; Paukstys, B.; Zeeb, B.A.; Reimer, K.J. Environmental Contamination and Remediation Practices at Former and Present Military Bases; Paukštys, B., Fonnum, F., Zeeb, B.A., Reimer, K.J., Eds.; Springer: Dordrecht, The Netherlands, 1998; ISBN 978-0-7923-5248-8. [Google Scholar]
- Gazenbeek, A. LIFE, Natura 2000 and the Military; Office for Official Publications of the European Communities: Luxembourg, 2005; ISBN 92-894-9213-9. [Google Scholar]
- Ellwanger, G.; Müller, C.; Ssymank, A.; Vischer-Leopold, M.; Paulsch, C. Management of Natura 2000 Sites on Military Training Areas; Landwirtschaftsverlag: Münster, Germany, 2016; ISBN 978-3-7843-9172-4. [Google Scholar]
- DANCEE; Andersern, J.N. Management of Contaminated Sites and Land in Central and Eastern Europe; Danish Environmental Protection Agency: Copenhagen, Denmark, 2000; ISBN 87-7909-889-4. Available online: https://www2.mst.dk/udgiv/publications/2000/87-7909-888-6/html/default_eng.htm (accessed on 2 November 2018).
- Nadtochiy, P.P.; Bilyavs’kyy, Y.A.; Myslyva, T.M.; Shmahala, Y.B. The problems of rehabilitation of the ground-landed resources of Zhytomyr oblast, muddy as a result of military activity. ВІСНИК ЖНАЕУ (VISNYK ZHNAEU) 2009, 2, 14–32. (In Ukrainian) [Google Scholar]
- Fazio, S.; Monti, A. Life cycle assessment of different bioenergy production systems including perennial and annual crops. Biomass Bioenergy 2011, 35, 4868–4878. [Google Scholar] [CrossRef]
- van der Weijde, T.; Alvim Kamei, C.L.; Torres, A.F.; Vermerris, W.; Dolstra, O.; Visser, R.G.F.; Trindade, L.M. The potential of C4 grasses for cellulosic biofuel production. Front. Plant Sci. 2013, 4, 107. [Google Scholar] [CrossRef] [PubMed]
- Pidlisnyuk, V.; Trögl, J.; Stefanovska, T.; Shapoval, P.; Erickson, L. Preliminary Results on Growing Second Generation Biofuel Crop Miscanthus x giganteus at The Polluted Military Site in Ukraine. Nova Biotechnol. Chim. 2016, 15, 77–84. [Google Scholar] [CrossRef]
- Pidlisnyuk, V.V.; Erickson, L.E.; Trögl, J.; Shapoval, P.Y.; Popelka, J.; Davis, L.C.; Stefanovska, T.R.; Hettiarachchi, G.M. Metals uptake behaviour in Miscanthus x giganteus plant during growth at the contaminated soil from the military site in Sliač, Slovakia. Pol. J. Chem. Technol. 2018, 20, 1–7. [Google Scholar] [CrossRef]
- Nsanganwimana, F.; Pourrut, B.; Waterlot, C.; Louvel, B.; Bidar, G.; Labidi, S.; Fontaine, J.; Muchembled, J.; Lounès-Hadj Sahraoui, A.; Fourrier, H.; et al. Metal accumulation and shoot yield of Miscanthus×giganteus growing in contaminated agricultural soils: Insights into agronomic practices. Agric. Ecosyst. Environ. 2015, 213, 61–71. [Google Scholar] [CrossRef]
- Andrejić, G.; Gajić, G.; Prica, M.; Dželetović, Ž.; Rakić, T. Zinc accumulation, photosynthetic gas exchange, and chlorophyll a fluorescence in Zn-stressed Miscanthus × giganteus plants. Photosynthetica 2018, 56, 1249–1258. [Google Scholar] [CrossRef]
- Iqbal, M.; Bermond, A.; Lamy, I. Impact of miscanthus cultivation on trace metal availability in contaminated agricultural soils: Complementary insights from kinetic extraction and physical fractionation. Chemosphere 2013, 91, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Al Souki, K.S.; Louvel, B.; Douay, F.; Pourrut, B. Assessment of Miscanthus x giganteus capacity to restore the functionality of metal-contaminated soilsEx situ experiment. Appl. Soil Ecol. 2017, 115, 44–52. [Google Scholar] [CrossRef]
- Nsanganwimana, F.; Pourrut, B.; Mench, M.; Douay, F. Suitability of Miscanthus species for managing inorganic and organic contaminated land and restoring ecosystem services. A review. J. Environ. Manag. 2014, 143, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Techer, D.; Martinez-Chois, C.; Laval-Gilly, P.; Henry, S.; Bennasroune, A.; D’Innocenzo, M.; Falla, J. Assessment of Miscanthus×giganteus for rhizoremediation of long term PAH contaminated soils. Appl. Soil Ecol. 2012, 62, 42–49. [Google Scholar] [CrossRef]
- Trögl, J.; Pavlorková, J.; Packová, P.; Seják, J.; Kuráň, P.; Popelka, J.; Pacina, J. Indication of Importance of Including Soil Microbial Characteristics into Biotope Valuation Method. Sustainability 2016, 8, 253. [Google Scholar] [CrossRef]
- Das, N.; Chandran, P. Microbial Degradation of Petroleum Hydrocarbon Contaminants: An Overview. Biotechnol. Res. Int. 2011, 2011, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Giller, K.E.; Witter, E.; Mcgrath, S.P. Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: A review. Soil Biol. Biochem. 1998, 30, 1389–1414. [Google Scholar] [CrossRef]
- Dai, J.; Becquer, T.; Rouiller, J.H.; Reversat, G.; Bernhard-Reversat, F.; Lavelle, P. Influence of heavy metals on C and N mineralisation and microbial biomass in Zn-, Pb-, Cu-, and Cd-contaminated soils. Appl. Soil Ecol. 2004, 25, 99–109. [Google Scholar] [CrossRef]
- Hiroki, M. Effect of arsenic pollution on soil microbial population. Soil Sci. Plant Nutr. 1993, 39, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Goswami, L.; Arul Manikandan, N.; Pakshirajan, K.; Pugazhenthi, G. Simultaneous heavy metal removal and anthracene biodegradation by the oleaginous bacteria Rhodococcus opacus. 3 Biotech 2017, 7, 37. [Google Scholar] [CrossRef] [PubMed]
- Riis, V.; Babel, W.; Pucci, O.H. Influence of heavy metals on the microbial degradation of diesel fuel. Chemosphere 2002, 49, 559–568. [Google Scholar] [CrossRef]
- Zukauskaite, A.; Jakubauskaite, V.; Belous, O.; Ambrazaitiene, D.; Stasiskiene, Z. Impact of heavy metals on the oil products biodegradation process. Waste Manag. Res. 2008, 26, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Chibuike, G.U.; Obiora, S.C. Heavy Metal Polluted Soils: Effect on Plants and Bioremediation Methods. Appl. Environ. Soil Sci. 2014, 2014, 1–12. [Google Scholar] [CrossRef]
- Kaur, A.; Chaudhary, A.; Kaur, A.; Choudhary, R.; Kaushik, R. Phospholipid fatty acid—A bioindicator of environment monitoring and assessment in soil ecosystem. Curr. Sci. 2005, 89, 1103–1112. [Google Scholar] [CrossRef]
- Pažická, A.; Brčeková, J.; Klimko, T.; Jurkovič, Ľ.; Peťková, K.; Kordík, J.; Slaninka, I. Characteristics of geogenic and anthropogenic sources of potentially toxic trace elements in selected stream sediments of Slovakia based on geochemical and mineralogical evaluation. Miner. Slovaca 2014, 45, 117–126. [Google Scholar]
- DSTU ISO 11464-2001: Soil Quality. Preliminary Preparation of Samples for Physicalchemical Analysis; DSTU: Kyiv, Ukraine, 2001.
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Kukla, J.; Holec, M.; Trögl, J.; Holcová, D.; Hofmanová, D.; Kuráň, P.; Popelka, J.; Pacina, J.; Kříženecká, S.; Usťak, S.; et al. Tourist Traffic Significantly Affects Microbial Communities of Sandstone Cave Sediments in the Protected Landscape Area “Labské Pískovce” (Czech Republic): Implications for Regulatory Measures. Sustainability 2018, 10, 396. [Google Scholar] [CrossRef]
- Frouz, J.; Cajthaml, T.; Kříbek, B.; Schaeffer, P.; Bartuška, M.; Galertová, R.; Rojík, P.; Krištůfek, V. Deep, subsurface microflora after excavation respiration and biomass and its potential role in degradation of fossil organic matter. Folia Microbiol. 2011, 56, 389–396. [Google Scholar] [CrossRef] [PubMed]
- 275/1998 Sb. Vyhláška Ministerstva Zemědělství o Agrochemickém Zkoušení Zemědělských půd a Zjišťování Půdních Vlastností Lesních Pozemků. Available online: http://eagri.cz/public/web/ws_content?contentKind=regulation§ion=1&id=46970&name=275/1998 (accessed on 31 October 2018).
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldrian, P.; Trögl, J.; Frouz, J.; Šnajdr, J.; Valášková, V.; Merhautová, V.; Cajthaml, T.; Herinková, J. Enzyme activities and microbial biomass in topsoil layer during spontaneous succession in spoil heaps after brown coal mining. Soil Biol. Biochem. 2008, 40, 2107–2115. [Google Scholar] [CrossRef]
- Robertson, A.D.; Davies, C.A.; Smith, P.; Stott, A.W.; Clark, E.L.; McNamara, N.P. Carbon Inputs from Miscanthus Displace Older Soil Organic Carbon without Inducing Priming. Bioenergy Res. 2017, 10, 86–101. [Google Scholar] [CrossRef]
- Técher, D.; Laval-Gilly, P.; Henry, S.; Bennasroune, A.; Formanek, P.; Martinez-Chois, C.; D’Innocenzo, M.; Muanda, F.; Dicko, A.; Rejšek, K.; et al. Contribution of Miscanthus x giganteus root exudates to the biostimulation of PAH degradation: An in vitro study. Sci. Total Environ. 2011, 409, 4489–4495. [Google Scholar] [CrossRef] [PubMed]
- Kuráň, P.; Trögl, J.; Nováková, J.; Pilařová, V.; Dáňová, P.; Pavlorková, J.; Kozler, J.; Novák, F.; Popelka, J. Biodegradation of Spilled Diesel Fuel in Agricultural Soil: Effect of Humates, Zeolite, and Bioaugmentation. Sci. World J. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed]
- United Nations. A/RES/70/1 Transforming Our World: The 2030 Agenda for Sustainable Development; United Nations: New York, NY, USA, 2015. [Google Scholar]
- Pidlisnyuk, V.; Stefanovska, T.; Lewis, E.E.; Erickson, L.E.; Davis, L.C. Miscanthus as a Productive Biofuel Crop for Phytoremediation. CRC Crit. Rev. Plant Sci. 2014, 33, 1–19. [Google Scholar] [CrossRef]
- Bastia, G.; Al Souki, K.; Liné, C.; Pourrut, B. Phytoremediation studies: Comparison between in situ and ex situ experiments results. In Proceedings of the 15th International Phytotechnology Conference (Phytotechnologies and Forestry: Sustainable Approaches to Mitigating the Environmental Consequences of Climate Change), Novi Sad, Serbia, 1–5 October 2018. [Google Scholar]
- Lee, W.-C.; Kuan, W.-C. Miscanthus as cellulosic biomass for bioethanol production. Biotechnol. J. 2015, 10, 840–854. [Google Scholar] [CrossRef] [PubMed]
- Cappelletto, P.; Mongardini, F.; Barberi, B.; Sannibale, M.; Brizzi, M.; Pignatelli, V. Papermaking pulps from the fibrous fraction of Miscanthus x Giganteus. Ind. Crops Prod. 2000, 11, 205–210. [Google Scholar] [CrossRef]
- Courard, L.; Parmentier, V. Carbonated miscanthus mineralized aggregates for reducing environmental impact of lightweight concrete blocks. Sustain. Build. 2017, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Moore-Kucera, J.; Dick, R.P. PLFA Profiling of Microbial Community Structure and Seasonal Shifts in Soils of a Douglas-fir Chronosequence. Microb. Ecol. 2008, 55, 500–511. [Google Scholar] [CrossRef] [PubMed]
- Frostegård, Å.; Tunlid, A.; Bååth, E. Use and misuse of PLFA measurements in soils. Soil Biol. Biochem. 2011, 43, 1621–1625. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Variant | Control | Low | High | ||
|---|---|---|---|---|---|
| control soil | 100% | 75% | 50% | 25% | 0% |
| post-military soil | 0% | 25% | 50% | 75% | 100% |
| As [mg/kg] | 290 ± 40 | 515 ± 15 | 430 ± 79 | 465 ± 15 | 425 ± 45 |
| Cu [mg/kg] | 310 ± 0 | 380 ± 60 | 395 ± 5 | 440 ± 100 | 565 ± 125 |
| Fe [mg/kg] | 174,555 ± 5395 | 194,485 ± 8645 | 205,640 ± 7540 | 209,480 ± 6830 | 215,210 ± 5720 |
| Mn [mg/kg] | 2995 ± 185 | 3605 ± 485 | 4110 ± 340 | 4495 ± 555 | 4660 ± 500 |
| Sr [mg/kg] | 685 ± 65 | 695 ± 15 | 925 ± 15 | 1185 ± 75 | 1200 ± 40 |
| Ti [mg/kg] | 20,620 ± 0 | 24,410 ± 260 | 25,935 ± 615 | 27,940 ± 2420 | 28,170 ± 530 |
| Zn [mg/kg] | 960 ± 90 | 1025 ± 45 | 1115 ± 115 | 1205 ± 335 | 1015 ± 235 |
| Zr [mg/kg] | 1275 ± 275 | 1455 ± 75 | 1345 ± 205 | 1500 ± 60 | 1625 ± 225 |
| Soil Nutrients | Metal Concentration | 1st Season | 2nd Season | ||
|---|---|---|---|---|---|
| Rhizosphere Soil | Bulk Soil | Rhizosphere Soil | Bulk Soil | ||
| Available P [mg/kg] | control | 49.9 ± 38.2 a | 45.8 ± 38.0 a | 18.2 ± 15.2 a | 43.0 ± 46.6 a |
| low | 90.1 ± 43.5 b | 96.3 ± 50.2 b | 90.1 ± 46.2 b | 55.1 ± 29.0 b | |
| high | 128.0 ± 107.0 b | 81.1 ± 92.7 b | 127.1 ± 110.5 b | 86.8 ± 82.2 b | |
| Available K [mg/kg] | control | 313.1 ± 31.4 a | 315.8 ± 51.9 a | 269.4 ± 18.5 a | 299.9 ± 73.5 a |
| low | 377.0 ± 81.5 b | 386.5 ± 89.4 b | 327.2 ± 27.4 b | 317.6 ± 55.7 b | |
| high | 416.1 ± 169.5 b | 381.1 ± 170.8 b | 346.4 ± 121.3 b | 369.7 ± 182.4 b | |
| Available Ca [mg/kg] | control | 3130.0 ± 30.3 a | 3273.0 ± 233.7 a | 3106.4 ± 49.6 a | 3185.3 ± 578.2 a |
| low | 3322.5 ± 121.3 b | 3362.5 ± 248.4 b | 3484.1 ± 233.9 b | 3223.4 ± 271.4 b | |
| high | 3276.9 ± 81.9 b | 3251.3 ± 421.2 b | 3291.6 ± 384.3 b | 3200.1 ± 139.3 b | |
| Available Mg [mg/kg] | control | 625.1 ± 3.5 a | 664.4 ± 64.9 a | 677.3 ± 24.1 a | 697.6 ± 144.3 a |
| low | 658.1 ± 24.6 b | 673.0 ± 51.5 b | 713.7 ± 104.3 b | 738.3 ± 76.2 b | |
| high | 634.5 ± 54.1 b | 721.4 ± 120.1 b | 653.9 ± 30.0 b | 683.4 ± 68.7 b | |
| Ntot [%] | control | 0.18 ± 0.08 a | 0.19 ± 0.09 a | 0.14 ± 0.01 a | 0.16 ± 0.09 a |
| low | 0.23 ± 0.04 b | 0.24 ± 0.05 b | 0.21 ± 0.01 b | 0.20 ± 0.05 b | |
| high | 0.24 ± 0.04 b | 0.28 ± 0.07 b | 0.22 ± 0.04 b | 0.23 ± 0.03 b | |
| S [%] | control | 0.03 ± 0.01 a | 0.04 ± 0.03 a | 0.02 ± 0.00 a | 0.02 ± 0.02 a |
| low | 0.04 ± 0.01 b | 0.04 ± 0.01 b | 0.03 ± 0.01 b | 0.03 ± 0.01 b | |
| high | 0.03 ± 0.00 b | 0.05 ± 0.02 b | 0.03 ± 0.00 b | 0.03 ± 0.01 b | |
| TOC [%] | control | 2.27 ± 1.01 a | 2.07 ± 1.04 a | 2.28 ± 0.36 a | 1.97 ± 0.88 a |
| low | 2.64 ± 0.68 b | 2.89 ± 0.74 b | 2.87 ± 0.46 b | 3.06 ± 1.26 b | |
| high | 3.05 ± 0.11 b | 3.40 ± 0.50 b | 3.36 ± 0.72 b | 3.14 ± 0.79 b | |
| Effect | P | K | Ca | Mg | S | Ntot | TOC |
|---|---|---|---|---|---|---|---|
| 1st season × 2nd season | 0.27 | 1.90 | 0.03 | 1.72 | 9.75↓ *** | 5.78↓ ** | 0.14 |
| rhizosphere × bulk | 1.02 | 0.00 | 0.32 | 2.82 | 2.15 | 0.67 | 0.08 |
| metals concentration | 2.72 | 1.49 | 1.37 | 0.63 | 1.38 | 7.74↑ *** | 6.76↑ *** |
| Effect | Phosp. | Arylsulp. | Proteas. | Oxidas. | Peroxidas. | Respirat. |
|---|---|---|---|---|---|---|
| 1st season × 2nd season | 55.71↓ *** | 2.54 | 12.42↑ *** | 18.55↓ *** | 117.98↑ *** | 8.03↑ *** |
| rhizosphere × bulk | 2.52 | 0.48 | 0.01 | 0.51 | 0.16 | 0.94 |
| metals concentration | 23.61↑ *** | 7.07↑ *** | 9.70↓ *** | 17.82↓ *** | 11.44↓ *** | 0.43 |
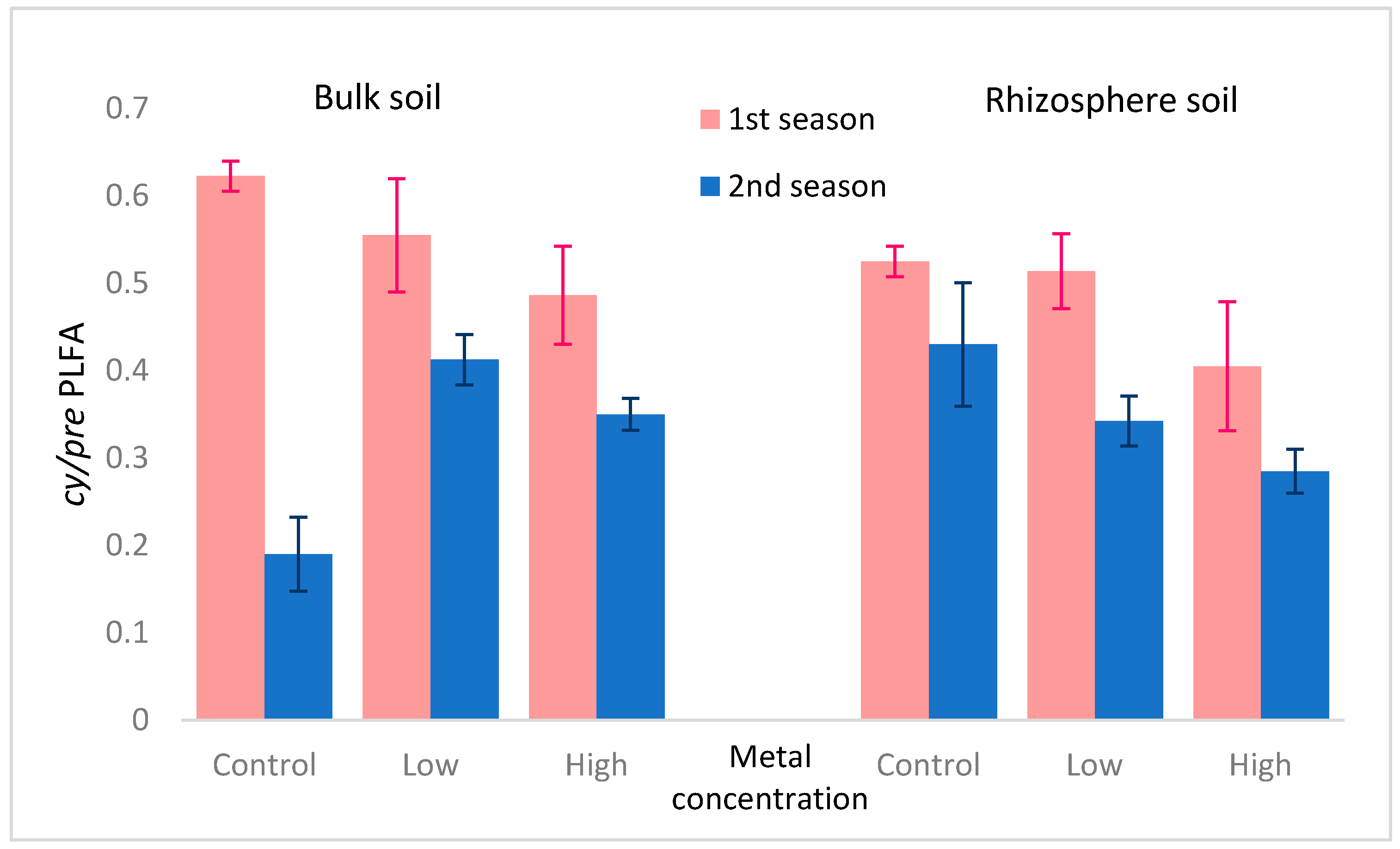
| Effect | PLFAG+ | PLFAG− | PLFAAc | PLFAFungi | trans/cis | cy/pre |
|---|---|---|---|---|---|---|
| 1st season × 2nd season | 13.82↑ *** | 6.27↓ ** | 176.94↓ *** | 27.64↑ *** | 2.92↓ * | 82.77↓ *** |
| rhizosphere × bulk | 0.00 | 0.29 | 0.58 | 0.98 | 3.27↑ * | 7.61↑ *** |
| metals concentration | 1.01 | 8.87↑ *** | 28.77↓ *** | 0.19 | 17.54↓ *** | 10.41↓ *** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nebeská, D.; Trögl, J.; Pidlisnyuk, V.; Popelka, J.; Veronesi Dáňová, P.; Usťak, S.; Honzík, R. Effect of Growing Miscanthus x giganteus on Soil Microbial Communities in Post-Military Soil. Sustainability 2018, 10, 4021. https://doi.org/10.3390/su10114021
Nebeská D, Trögl J, Pidlisnyuk V, Popelka J, Veronesi Dáňová P, Usťak S, Honzík R. Effect of Growing Miscanthus x giganteus on Soil Microbial Communities in Post-Military Soil. Sustainability. 2018; 10(11):4021. https://doi.org/10.3390/su10114021
Chicago/Turabian StyleNebeská, Diana, Josef Trögl, Valentina Pidlisnyuk, Jan Popelka, Petra Veronesi Dáňová, Sergej Usťak, and Roman Honzík. 2018. "Effect of Growing Miscanthus x giganteus on Soil Microbial Communities in Post-Military Soil" Sustainability 10, no. 11: 4021. https://doi.org/10.3390/su10114021