
Natural Products as a Tool to Modulate the Activity and Expression of Multidrug Resistance Proteins of Intestinal Barrier


Abstract
:1. Introduction
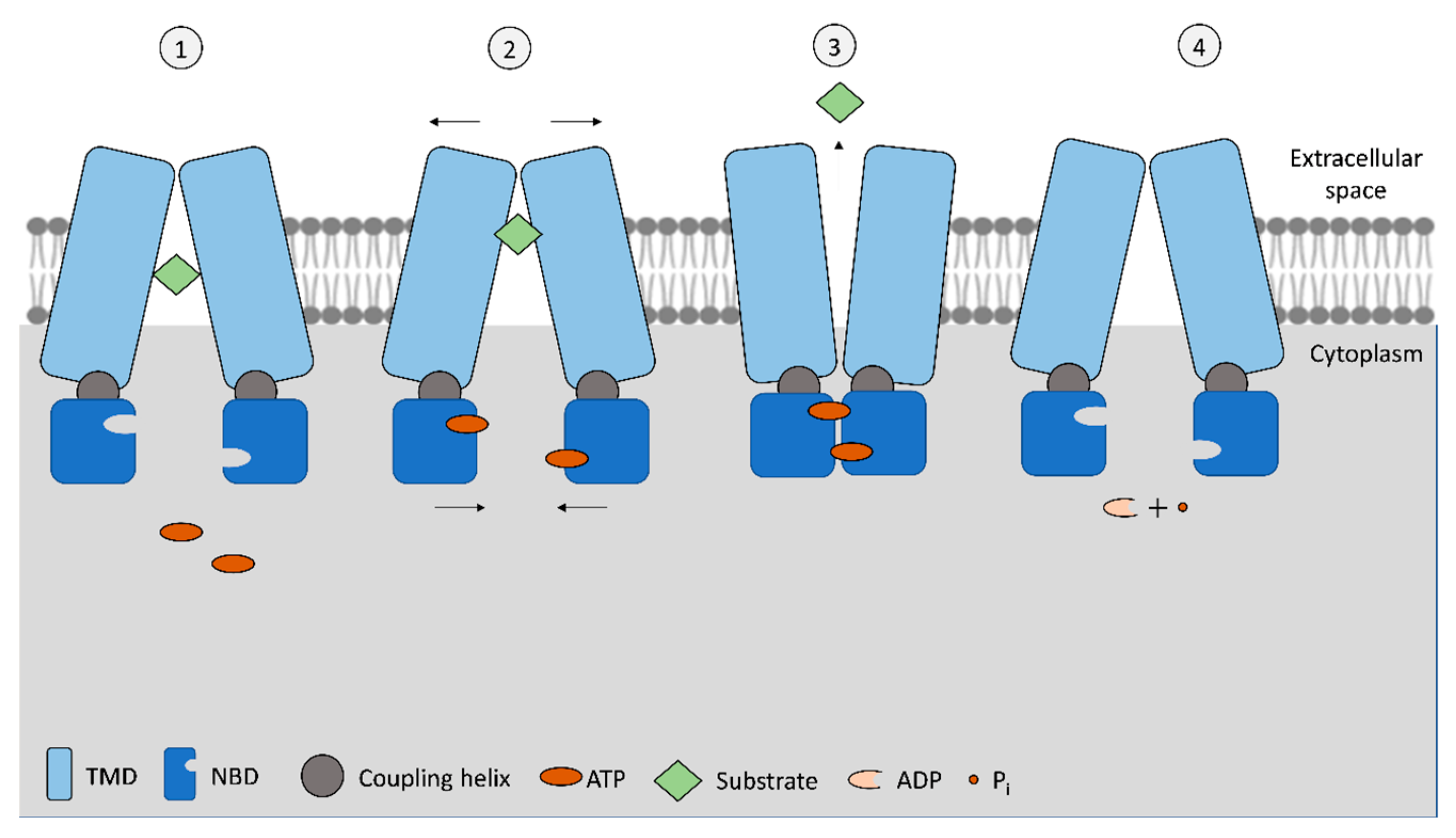
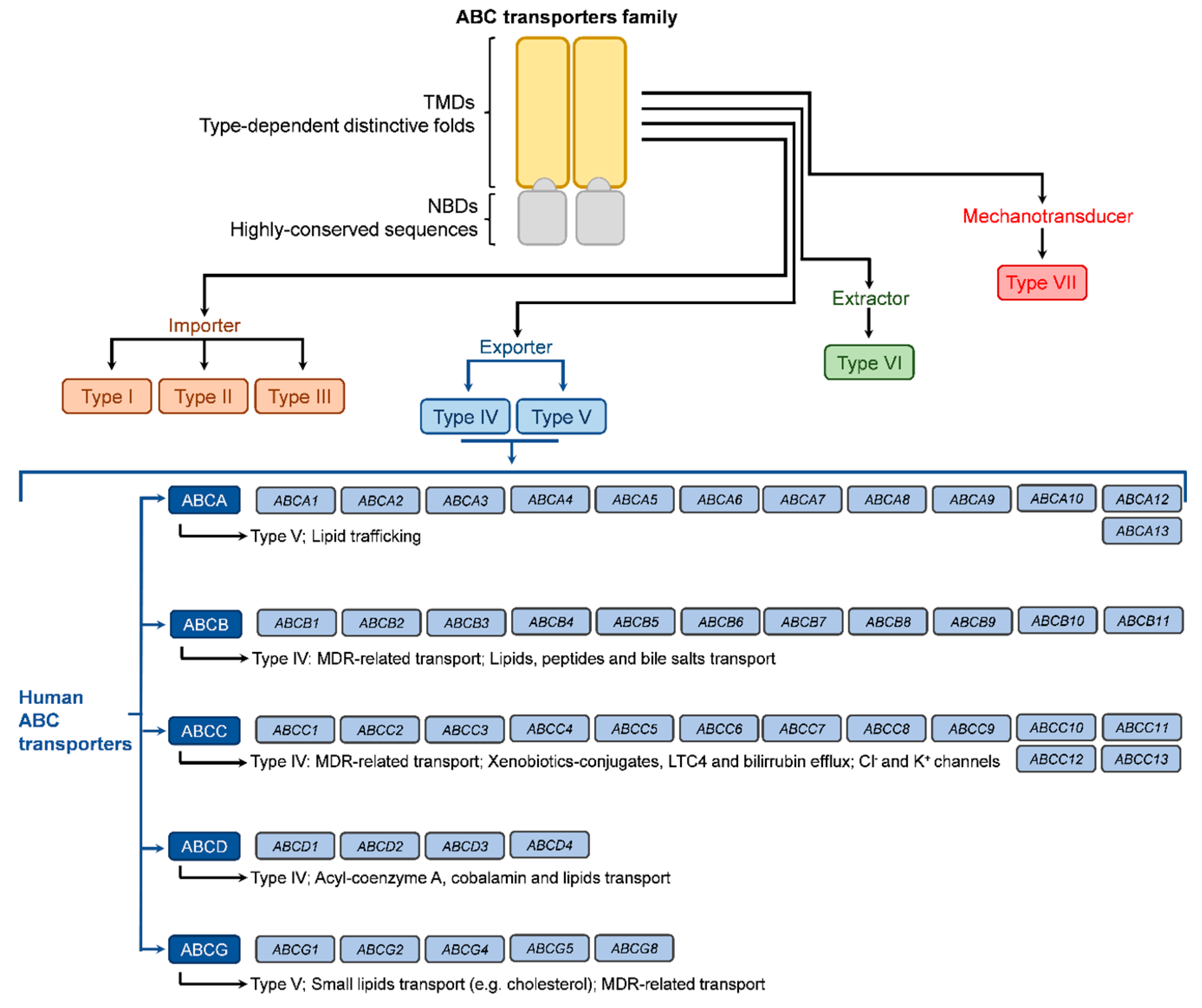
2. ATP Binding Cassette (ABC) Transporters Family
2.1. Main ABC-Transporters Implicated in Multi-Drug Resistance in Colorectal Cancer
2.1.1. P-glycoprotein
2.1.2. Multidrug Resistance-Associated Proteins Subfamily
2.1.3. Subfamily G
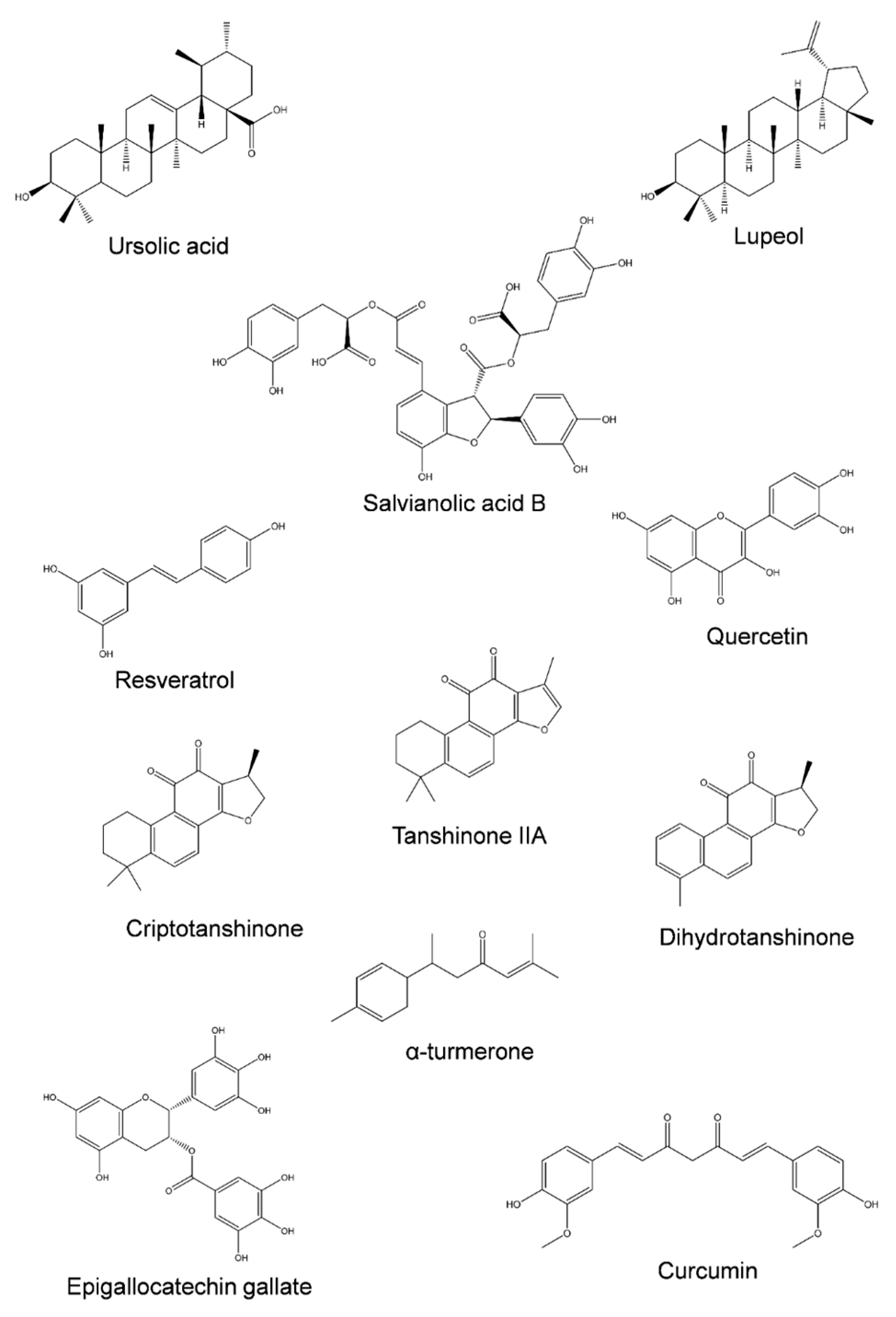
3. Natural Products as Potential Candidates to Overcome Colorectal Cancer MDR

{kind=link}
{kind=link}
{kind=link}
| Compound | Experimental Model | Concentration | Effect | Ref. |
|---|---|---|---|---|
| Ursolic acid | HCT-8 and SW620 cells | 20 µM | Increased effect of oxaliplatin; Reduced P-gp expression | [69] |
| RKO, LoVo, and SW480 cells | 20 µM | Reduced P-gp expression; Increased the sensitivity to 5-fluroacil and oxaliplatin | [95] | |
| Caco-2 cells | >200 µM | Chemosensitizing effect for doxorubicin; P-gp inhibition at high concentrations | [96] | |
| Salvianolic acid B | HCT-8 cells | 20 µg/mL | Increased sensitivity to 5-fluorouracil, cisplatin, vincristine and paclitaxel; Reduced P-gp expression | [107] |
| LoVo and HCT-116 cancer stem cells xenografts in mice | 0.36 g | Reduced ABCG2 expression | [106] | |
| α-turmerone | Caco-2 cells | 50 µg/mL | Reduced P-gp expression | [114] |
| Curcumin | HCT-8 and HCT-8/5-FU cells | 5.5 µg/mL | Increased sensitivity to 5-fluorouracil; Reduced P-gp expression | [115] |
| HCT-8 and HCT-8/5-FU cells | 12.96 µg/mL | Increased 5-fluorouracil effect; Reduced P-gp expression | [116] | |
| SW620 and SW620/Ad300 cells | 5.5 µM | Increased doxorubicin effect; Reduced P-gp activity | [117] | |
| In situ Cancerous Colon Perfusion Rat Model | 50 mg/kg | Reduced P-gp expression | [118] | |
| Quercetin | SW620/Ad300 cells | 33 µM | Increased doxorubicin effect; Inhibited P-gp activity | [119] |
| Tanshinone IIA | SCID mice with Colo205 cell xenograft | 20 mg/kg | Increased 5-fluorouracil effect; Downregulation of P-gp | [104] |
| Cryptotanshinone | Caco-2 and SW620/Ad300 cells | 25 µM | Increased doxorubicin and irinotecan toxicity; P-gp transport inhibition | [103] |
| Dihydrotanshinone | Caco-2 and SW620/Ad300 cells | 25 µM | Increased doxorubicin and irinotecan toxicity; P-gp transport inhibition | [103] |
| Cinobufagin | LoVo/ADR, HCT-116/L-OHP, and Caco-2/ADR cells | 20 nM | Inhibited P-gp activity | [120] |
| Resveratrol | HCT-116/L-OHP cells | 50 µM | Downregulation of P-gp | [108] |
| Epigallocatechin gallate | HCT-116 and DLD-1 cells | 50 µM | Increased 5-fluorouracil effect; Suppressed MDR1 expression | [59] |
| Lupeol | LoVo cells | 50 µM | Increased oxaliplatin effect; ABCG2 suppression | [102] |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ho, G.T.; Moodie, F.M.; Satsangi, J. Multidrug resistance 1 gene (P-glycoprotein 170): An important determinant in gastrointestinal disease? Gut 2003, 52, 759–766. [Google Scholar] [CrossRef] [Green Version]
- Cong, Y.; Baimanov, D.; Zhou, Y.; Chen, C.; Wang, L. Penetration and translocation of functional inorganic nanomaterials into biological barriers. Adv. Drug Deliv. Rev. 2022, 191, 114615. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.C. Molecular mechanisms of cellular cholesterol efflux. J. Biol. Chem. 2014, 289, 24020–24029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, W.-T.; Xu, D.; Wang, L.; Chen, Y.; Chen, Z.-P.; Zhou, C.-Z.; Chen, Y. Plastic structures for diverse substrates: A revisit of human ABC transporters. Proteins Struct. Funct. Bioinform. 2022, 90, 1749–1765. [Google Scholar] [CrossRef] [PubMed]
- Godbey, W.T. Chapter 3—Cellular Transport. In An Introduction to Biotechnology; Godbey, W.T., Ed.; Woodhead Publishing: Cambridge UK, 2014; pp. 35–64. [Google Scholar]
- Choi, C.C.; Ford, R.C. ATP binding cassette importers in eukaryotic organisms. Biol. Rev. 2021, 96, 1318–1330. [Google Scholar] [CrossRef] [PubMed]
- Bukowski, K.; Kciuk, M.; Kontek, R. Mechanisms of Multidrug Resistance in Cancer Chemotherapy. Int. J. Mol. Sci. 2020, 21, 3233. [Google Scholar] [CrossRef]
- Aires, V.; Colin, D.J.; Doreau, A.; Di Pietro, A.; Heydel, J.-M.; Artur, Y.; Latruffe, N.; Delmas, D. P-Glycoprotein 1 Affects Chemoactivities of Resveratrol against Human Colorectal Cancer Cells. Nutrients 2019, 11, 2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunjachan, S.; Rychlik, B.; Storm, G.; Kiessling, F.; Lammers, T. Multidrug resistance: Physiological principles and nanomedical solutions. Adv. Drug Deliv. Rev. 2013, 65, 1852–1865. [Google Scholar] [CrossRef] [Green Version]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef]
- Sun, Y.-L.; Patel, A.; Kumar, P.; Chen, Z.-S. Role of ABC transporters in cancer chemotherapy. Chin. J. Cancer 2012, 31, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Ling, V.; Thompson, L.H. Reduced permeability in CHO cells as a mechanism of resistance to colchicine. J. Cell. Physiol. 1974, 83, 103–116. [Google Scholar] [CrossRef]
- Juliano, R.L.; Ling, V. A surface glycoprotein modulating drug permeability in Chinese hamster ovary cell mutants. Biochim. Biophys. Acta (BBA)—Biomembr. 1976, 455, 152–162. [Google Scholar] [CrossRef]
- Kartner, N.; Shales, M.; Riordan, J.R.; Ling, V. Daunorubicin-resistant Chinese hamster ovary cells expressing multidrug resistance and a cell-surface P-glycoprotein. Cancer Res. 1983, 43, 4413–4419. [Google Scholar]
- Gupta, R.S. Cross-resistance of vinblastine- and taxol-resistant mutants of Chinese hamster ovary cells to other anticancer drugs. Cancer Treat. Rep. 1985, 69, 515–521. [Google Scholar]
- Rice, A.J.; Park, A.; Pinkett, H.W. Diversity in ABC transporters: Type I, II and III importers. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 426–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ter Beek, J.; Guskov, A.; Slotboom, D.J. Structural diversity of ABC transporters. J. Gen. Physiol. 2014, 143, 419–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.; Tampé, R. Structural and Mechanistic Principles of ABC Transporters. Annu. Rev. Biochem. 2020, 89, 605–636. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Seelig, A.; Bernèche, S. Unidirectional Transport Mechanism in an ATP Dependent Exporter. ACS Cent. Sci. 2017, 3, 250–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.; Aller, S.G.; Beis, K.; Carpenter, E.P.; Chang, G.; Chen, L.; Dassa, E.; Dean, M.; Duong Van Hoa, F.; Ekiert, D.; et al. Structural and functional diversity calls for a new classification of ABC transporters. FEBS Lett. 2020, 594, 3767–3775. [Google Scholar] [CrossRef]
- Greene, N.P.; Kaplan, E.; Crow, A.; Koronakis, V. Antibiotic Resistance Mediated by the MacB ABC Transporter Family: A Structural and Functional Perspective. Front. Microbiol. 2018, 9, 590. [Google Scholar] [CrossRef] [Green Version]
- Vasiliou, V.; Vasiliou, K.; Nebert, D.W. Human ATP-binding cassette (ABC) transporter family. Hum. Genom. 2009, 3, 281–290. [Google Scholar] [CrossRef]
- Stockner, T.; Gradisch, R.; Schmitt, L. The role of the degenerate nucleotide binding site in type I ABC exporters. FEBS Lett. 2020, 594, 3815–3838. [Google Scholar] [CrossRef] [PubMed]
- Gerovac, M.; Tampé, R. Control of mRNA Translation by Versatile ATP-Driven Machines. Trends Biochem. Sci. 2019, 44, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Joung, J.-Y.; Cho, J.-H.; Son, C.-G.; Lee, N. Overcoming P-Glycoprotein-Mediated Multidrug Resistance in Colorectal Cancer: Potential Reversal Agents among Herbal Medicines. Evid.-Based Complement. Altern. Med. 2018, 2018, 3412074. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, A.I.; Real, R.; Pérez, M.; Mendoza, G.; Prieto, J.G.; Merino, G. Modulation of the activity of ABC transporters (P-glycoprotein, MRP2, BCRP) by flavonoids and drug response. J. Pharm. Sci. 2010, 99, 598–617. [Google Scholar] [CrossRef]
- Srikant, S.; Gaudet, R. Mechanics and pharmacology of substrate selection and transport by eukaryotic ABC exporters. Nat. Struct. Mol. Biol. 2019, 26, 792–801. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.M.; George, A.M. The Switch and Reciprocating Models for the Function of ABC Multidrug Exporters: Perspectives on Recent Research. Int. J. Mol. Sci. 2023, 24, 2624. [Google Scholar] [CrossRef] [PubMed]
- Elmeliegy, M.; Vourvahis, M.; Guo, C.; Wang, D.D. Effect of P-glycoprotein (P-gp) Inducers on Exposure of P-gp Substrates: Review of Clinical Drug–Drug Interaction Studies. Clin. Pharmacokinet. 2020, 59, 699–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palko-Labuz, A.; Sroda-Pomianek, K.; Uryga, A.; Kostrzewa-Suslow, E.; Michalak, K. Anticancer activity of baicalein and luteolin studied in colorectal adenocarcinoma LoVo cells and in drug-resistant LoVo/Dx cells. Biomed. Pharmacother. 2017, 88, 232–241. [Google Scholar] [CrossRef]
- Cole, S.P.C.; Bhardwaj, G.; Gerlach, J.H.; Mackie, J.E.; Grant, C.E.; Almquist, K.C.; Stewart, A.J.; Kurz, E.U.; Duncan, A.M.V.; Deeley, R.G. Overexpression of a Transporter Gene in a Multidrug-Resistant Human Lung Cancer Cell Line. Science 1992, 258, 1650–1654. [Google Scholar] [CrossRef]
- Borst, P.; Evers, R.; Kool, M.; Wijnholds, J. The multidrug resistance protein family. Biochim. Biophys. Acta (BBA)—Biomembr. 1999, 1461, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Hopper, E.; Belinsky, M.G.; Zeng, H.; Tosolini, A.; Testa, J.R.; Kruh, G.D. Analysis of the structure and expression pattern of MRP7 (ABCC10), a new member of the MRP subfamily. Cancer Lett. 2001, 162, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Bortfeld, M.; Rius, M.; König, J.; Herold-Mende, C.; Nies, A.T.; Keppler, D. Human multidrug resistance protein 8 (MRP8/ABCC11), an apical efflux pump for steroid sulfates, is an axonal protein of the CNS and peripheral nervous system. Neuroscience 2006, 137, 1247–1257. [Google Scholar] [CrossRef] [PubMed]
- Bera Tapan, K.; Iavarone, C.; Kumar, V.; Lee, S.; Lee, B.; Pastan, I. MRP9, an unusual truncated member of the ABC transporter superfamily, is highly expressed in breast cancer. Proc. Natl. Acad. Sci. USA 2002, 99, 6997–7002. [Google Scholar] [CrossRef] [Green Version]
- Sodani, K.; Patel, A.; Kathawala, R.J.; Chen, Z.-S. Multidrug resistance associated proteins in multidrug resistance. Chin. J. Cancer 2012, 31, 58–72. [Google Scholar] [CrossRef] [Green Version]
- Cole, S.P. Targeting multidrug resistance protein 1 (MRP1, ABCC1): Past, present, and future. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 95–117. [Google Scholar] [CrossRef] [PubMed]
- Kruh, G.D.; Belinsky, M.G. The MRP family of drug efflux pumps. Oncogene 2003, 22, 7537–7552. [Google Scholar] [CrossRef]
- He, S.M.; Li, R.; Kanwar, J.R.; Zhou, S.F. Structural and functional properties of human multidrug resistance protein 1 (MRP1/ABCC1). Curr. Med. Chem. 2011, 18, 439–481. [Google Scholar] [CrossRef]
- Cao, D.; Qin, S.; Mu, Y.; Zhong, M. The role of MRP1 in the multidrug resistance of colorectal cancer. Oncol. Lett. 2017, 13, 2471–2476. [Google Scholar] [CrossRef] [Green Version]
- Hasanabady, M.H.; Kalalinia, F. ABCG2 inhibition as a therapeutic approach for overcoming multidrug resistance in cancer. J. Biosci. 2016, 41, 313–324. [Google Scholar] [CrossRef]
- Mo, W.; Zhang, J.-T. Human ABCG2: Structure, function, and its role in multidrug resistance. Int. J. Biochem. Mol. Biol. 2012, 3, 1–27. [Google Scholar] [PubMed]
- Groenen, A.G.; Halmos, B.; Tall, A.R.; Westerterp, M. Cholesterol efflux pathways, inflammation, and atherosclerosis. Crit. Rev. Biochem. Mol. Biol. 2021, 56, 426–439. [Google Scholar] [CrossRef]
- Roundhill, E.A.; Jabri, S.; Burchill, S.A. ABCG1 and Pgp identify drug resistant, self-renewing osteosarcoma cells. Cancer Lett. 2019, 453, 142–157. [Google Scholar] [CrossRef] [PubMed]
- Lou, J.; Zhou, H.; Li, C.; Hu, L.; Lu, X.; Li, J.; Yao, H.; Li, W.; Zhang, X.; Xu, M. ABCA1 and ABCG1 Expression in the Small Intestine of Type 2 Diabetic Rats. Lab. Med. 2014, 45, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özvegy, C.; Litman, T.; Szakács, G.; Nagy, Z.; Bates, S.; Váradi, A.; Sarkadi, B. Functional Characterization of the Human Multidrug Transporter, ABCG2, Expressed in Insect Cells. Biochem. Biophys. Res. Commun. 2001, 285, 111–117. [Google Scholar] [CrossRef]
- Tan, K.W.; Li, Y.; Paxton, J.W.; Birch, N.P.; Scheepens, A. Identification of novel dietary phytochemicals inhibiting the efflux transporter breast cancer resistance protein (BCRP/ABCG2). Food Chem. 2013, 138, 2267–2274. [Google Scholar] [CrossRef]
- Luciani, M.F.; Denizot, F.; Savary, S.; Mattei, M.G.; Chimini, G. Cloning of Two Novel ABC Transporters Mapping on Human Chromosome 9. Genomics 1994, 21, 150–159. [Google Scholar] [CrossRef]
- Qian, H.; Zhao, X.; Cao, P.; Lei, J.; Yan, N.; Gong, X. Structure of the Human Lipid Exporter ABCA1. Cell 2017, 169, 1228–1239.e1210. [Google Scholar] [CrossRef] [Green Version]
- Babashamsi, M.M.; Koukhaloo, S.Z.; Halalkhor, S.; Salimi, A.; Babashamsi, M. ABCA1 and metabolic syndrome; a review of the ABCA1 role in HDL-VLDL production, insulin-glucose homeostasis, inflammation and obesity. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 1529–1534. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, C. Regulation of ABCA1 functions by signaling pathways. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2012, 1821, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Reboul, E. Vitamin E intestinal absorption: Regulation of membrane transport across the enterocyte. IUBMB Life 2019, 71, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Niesor, E.J. Will Lipidation of ApoA1 through Interaction with ABCA1 at the Intestinal Level Affect the Protective Functions of HDL? Biology 2015, 4, 17–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguirre-Portolés, C.; Feliu, J.; Reglero, G.; Ramírez de Molina, A. ABCA1 overexpression worsens colorectal cancer prognosis by facilitating tumour growth and caveolin-1-dependent invasiveness, and these effects can be ameliorated using the BET inhibitor apabetalone. Mol. Oncol. 2018, 12, 1735–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jodoin, J.; Demeule, M.; Béliveau, R. Inhibition of the multidrug resistance P-glycoprotein activity by green tea polyphenols. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2002, 1542, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F.; Huang, W.; Xu, T.; Wu, L.; Chen, Q.; Peng, J.; Liu, X.; Lu, B. Natural P-gp inhibitor EGCG improves the acteoside absorption in Caco-2 cell monolayers and increases the oral bioavailability of acteoside in rats. Food Chem. Toxicol. 2020, 146, 111827. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, S.; Nabekura, T.; Kamiyama, S. Inhibition of P-glycoprotein function by tea catechins in KB-C2 cells. J. Pharm. Pharmacol. 2004, 56, 1001–1005. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.I.G.; Fragoso, L.R.; Esparza, J.R. Effect of EGCG on the expression and activity of P-glycoprotein in breast cancer cells. FASEB J. 2013, 27, 1167.6. [Google Scholar] [CrossRef]
- La, X.; Zhang, L.; Li, Z.; Li, H.; Yang, Y. (−)-Epigallocatechin Gallate (EGCG) Enhances the Sensitivity of Colorectal Cancer Cells to 5-FU by Inhibiting GRP78/NF-κB/miR-155-5p/MDR1 Pathway. J. Agric. Food Chem. 2019, 67, 2510–2518. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Sinha, D. EGCG maintained Nrf2-mediated redox homeostasis and minimized etoposide resistance in lung cancer cells. J. Funct. Foods 2019, 62, 103553. [Google Scholar] [CrossRef]
- Moradzadeh, M.; Roustazadeh, A.; Tabarraei, A.; Erfanian, S.; Sahebkar, A. Epigallocatechin-3-gallate enhances differentiation of acute promyelocytic leukemia cells via inhibition of PML-RARα and HDAC1. Phytother. Res. 2018, 32, 471–479. [Google Scholar] [CrossRef]
- Ravikumar Reddy, D.; Khurana, A.; Bale, S.; Ravirala, R.; Samba Siva Reddy, V.; Mohankumar, M.; Godugu, C. Natural flavonoids silymarin and quercetin improve the brain distribution of co-administered P-gp substrate drugs. SpringerPlus 2016, 5, 1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Zhao, Q.; Wang, B.; Yuan, S.; Wang, X.; Li, K. Quercetin reversed MDR in breast cancer cells through down-regulating P-gp expression and eliminating cancer stem cells mediated by YB-1 nuclear translocation. Phytother. Res. 2018, 32, 1530–1536. [Google Scholar] [CrossRef]
- van Zanden, J.J.; van der Woude, H.; Vaessen, J.; Usta, M.; Wortelboer, H.M.; Cnubben, N.H.P.; Rietjens, I.M.C.M. The effect of quercetin phase II metabolism on its MRP1 and MRP2 inhibiting potential. Biochem. Pharmacol. 2007, 74, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Mohos, V.; Fliszár-Nyúl, E.; Ungvári, O.; Kuffa, K.; Needs, P.W.; Kroon, P.A.; Telbisz, Á.; Özvegy-Laczka, C.; Poór, M. Inhibitory Effects of Quercetin and Its Main Methyl, Sulfate, and Glucuronic Acid Conjugates on Cytochrome P450 Enzymes, and on OATP, BCRP and MRP2 Transporters. Nutrients 2020, 12, 2306. [Google Scholar] [CrossRef]
- Wang, W.-J.; Sui, H.; Qi, C.; Li, Q.; Zhang, J.; Wu, S.-F.; Mei, M.-Z.; Lu, Y.-Y.; Wan, Y.-T.; Chang, H.; et al. Ursolic acid inhibits proliferation and reverses drug resistance of ovarian cancer stem cells by downregulating ABCG2 through suppressing the expression of hypoxia-inducible factor-1α in vitro. Oncol. Rep. 2016, 36, 428–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, J.-h.; Wei, X.-h.; Sheng, X.-y.; Zhou, D.-q.; Peng, H.-w.; Lu, Y.-n.; Zhou, J. Effect of Ursolic Acid on Breast Cancer Resistance Protein-mediated Transport of Rosuvastatin In Vivo and Vitro. Chin. Med. Sci. J. 2015, 30, 218–225. [Google Scholar] [CrossRef]
- Nabekura, T.; Yamaki, T.; Hiroi, T.; Ueno, K.; Kitagawa, S. Inhibition of anticancer drug efflux transporter P-glycoprotein by rosemary phytochemicals. Pharmacol. Res. 2010, 61, 259–263. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, L.; Shi, H.; Chen, H.; Tao, J.; Shen, R.; Wang, T. Ursolic acid enhances the therapeutic effects of oxaliplatin in colorectal cancer by inhibition of drug resistance. Cancer Sci. 2018, 109, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Villar, V.H.; Vögler, O.; Barceló, F.; Gómez-Florit, M.; Martínez-Serra, J.; Obrador-Hevia, A.; Martín-Broto, J.; Ruiz-Gutiérrez, V.; Alemany, R. Oleanolic and maslinic acid sensitize soft tissue sarcoma cells to doxorubicin by inhibiting the multidrug resistance protein MRP-1, but not P-glycoprotein. J. Nutr. Biochem. 2014, 25, 429–438. [Google Scholar] [CrossRef] [Green Version]
- Braga, F.; Ayres-Saraiva, D.; Gattass, C.R.; Capella, M.A.M. Oleanolic acid inhibits the activity of the multidrug resistance protein ABCC1 (MRP1) but not of the ABCB1 (P-glycoprotein): Possible use in cancer chemotherapy. Cancer Lett. 2007, 248, 147–152. [Google Scholar] [CrossRef]
- An, G.; Gallegos, J.; Morris, M.E. The Bioflavonoid Kaempferol Is an Abcg2 Substrate and Inhibits Abcg2-Mediated Quercetin Efflux. Drug Metab. Dispos. 2011, 39, 426. [Google Scholar] [CrossRef] [Green Version]
- Nabekura, T.; Kamiyama, S.; Kitagawa, S. Effects of dietary chemopreventive phytochemicals on P-glycoprotein function. Biochem. Biophys. Res. Commun. 2005, 327, 866–870. [Google Scholar] [CrossRef]
- Liu, C.-M.; Kao, C.-L.; Tseng, Y.-T.; Lo, Y.-C.; Chen, C.-Y. Ginger Phytochemicals Inhibit Cell Growth and Modulate Drug Resistance Factors in Docetaxel Resistant Prostate Cancer Cell. Molecules 2017, 22, 1477. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Ye, M.; Lu, Y.; Zhang, H.; Chen, Q.; Huang, S.; Su, S. Curcumin Improves the Tumoricidal Effect of Mitomycin C by Suppressing ABCG2 Expression in Stem Cell-Like Breast Cancer Cells. PLoS ONE 2015, 10, e0136694. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Mukherjee, S. Reversal of resistance towards cisplatin by curcumin in cervical cancer cells. Asian Pac. J. Cancer Prev. APJCP 2014, 15, 1403–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.-L.; Takahashi, K.; Tanaka, K.; Tougou, K.; Qiu, F.; Komatsu, K.; Takahashi, K.; Azuma, J. Curcuma drugs and curcumin regulate the expression and function of P-gp in Caco-2 cells in completely opposite ways. Int. J. Pharm. 2008, 358, 224–229. [Google Scholar] [CrossRef]
- Jia, Y.; Liu, Z.; Wang, C.; Meng, Q.; Huo, X.; Liu, Q.; Sun, H.; Sun, P.; Yang, X.; Ma, X.; et al. P-gp, MRP2 and OAT1/OAT3 mediate the drug-drug interaction between resveratrol and methotrexate. Toxicol. Appl. Pharmacol. 2016, 306, 27–35. [Google Scholar] [CrossRef]
- El-Readi, M.Z.; Eid, S.; Abdelghany, A.A.; Al-Amoudi, H.S.; Efferth, T.; Wink, M. Resveratrol mediated cancer cell apoptosis, and modulation of multidrug resistance proteins and metabolic enzymes. Phytomedicine 2019, 55, 269–281. [Google Scholar] [CrossRef]
- Kweon, S.H.; Song, J.H.; Kim, T.S. Resveratrol-mediated reversal of doxorubicin resistance in acute myeloid leukemia cells via downregulation of MRP1 expression. Biochem. Biophys. Res. Commun. 2010, 395, 104–110. [Google Scholar] [CrossRef]
- Shankar, S.; Nall, D.; Tang, S.-N.; Meeker, D.; Passarini, J.; Sharma, J.; Srivastava, R.K. Resveratrol Inhibits Pancreatic Cancer Stem Cell Characteristics in Human and KrasG12D Transgenic Mice by Inhibiting Pluripotency Maintaining Factors and Epithelial-Mesenchymal Transition. PLoS ONE 2011, 6, e16530. [Google Scholar] [CrossRef] [Green Version]
- Marley, A.R.; Nan, H. Epidemiology of colorectal cancer. Int. J. Mol. Epidemiol. Genet. 2016, 7, 105–114. [Google Scholar] [PubMed]
- Granados-Romero, J.J.; Valderrama-Treviño, A.I.; Contreras-Flores, E.H.; Barrera-Mera, B.; Herrera Enríquez, M.; Uriarte-Ruíz, K.; Ceballos-Villalba, J.; Estrada-Mata, A.G.; Alvarado Rodríguez, C.; Arauz-Peña, G. Colorectal cancer: A review. Int. J. Res. Med. Sci. 2017, 5, 4667–4676. [Google Scholar] [CrossRef] [Green Version]
- Favoriti, P.; Carbone, G.; Greco, M.; Pirozzi, F.; Pirozzi, R.E.M.; Corcione, F. Worldwide burden of colorectal cancer: A review. Updat. Surg. 2016, 68, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.-Y.; Lee, J.-S.; Son, C.-G.; Lee, N.-H. Combating Drug Resistance in Colorectal Cancer Using Herbal Medicines. Chin. J. Integr. Med. 2021, 27, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Aniogo, E.; George, B.P.; Abrahamse, H. Plant-Based Compounds as Alternative Adjuvant Therapy for Multidrug-Resistant Cancer. In Phytomedicine: Research and Development; CRC Press: Boca Raton, FL, USA, 2020; pp. 7–12. [Google Scholar]
- Cort, A.; Ozben, T. Natural Product Modulators to Overcome Multidrug Resistance in Cancer. Nutr. Cancer 2015, 67, 411–423. [Google Scholar] [CrossRef]
- Costea, T.; Vlad, O.C.; Miclea, L.-C.; Ganea, C.; Szöllősi, J.; Mocanu, M.-M. Alleviation of Multidrug Resistance by Flavonoid and Non-Flavonoid Compounds in Breast, Lung, Colorectal and Prostate Cancer. Int. J. Mol. Sci. 2020, 21, 401. [Google Scholar] [CrossRef] [Green Version]
- Deferme, S.; Augustijns, P. The effect of food components on the absorption of P-gp substrates: A review. J. Pharm. Pharmacol. 2003, 55, 153–162. [Google Scholar] [CrossRef]
- Kwatra, D.; Venugopal, A.; Standing, D.; Ponnurangam, S.; Dhar, A.; Mitra, A.; Anant, S. Bitter Melon Extracts Enhance the Activity of Chemotherapeutic Agents Through the Modulation of Multiple Drug Resistance. J. Pharm. Sci. 2013, 102, 4444–4454. [Google Scholar] [CrossRef] [Green Version]
- Ben Toumia, I.; Sobeh, M.; Ponassi, M.; Banelli, B.; Dameriha, A.; Wink, M.; Chekir Ghedira, L.; Rosano, C. A Methanol Extract of Scabiosa atropurpurea Enhances Doxorubicin Cytotoxicity against Resistant Colorectal Cancer Cells In Vitro. Molecules 2020, 25, 5265. [Google Scholar] [CrossRef]
- González-Vallinas, M.; Molina, S.; Vicente, G.; de la Cueva, A.; Vargas, T.; Santoyo, S.; García-Risco, M.R.; Fornari, T.; Reglero, G.; Ramírez de Molina, A. Antitumor effect of 5-fluorouracil is enhanced by rosemary extract in both drug sensitive and resistant colon cancer cells. Pharmacol. Res. 2013, 72, 61–68. [Google Scholar] [CrossRef]
- Perloff, M.D.; von Moltke, L.L.; Störmer, E.; Shader, R.I.; Greenblatt, D.J. Saint John’s wort: An in vitro analysis of P-glycoprotein induction due to extended exposure. Br. J. Pharmacol. 2001, 134, 1601–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedl, A.; Schlederer, M.; Pudelko, K.; Stadler, M.; Walter, S.; Unterleuthner, D.; Unger, C.; Kramer, N.; Hengstschläger, M.; Kenner, L.; et al. Comparison of cancer cells in 2D vs. 3D culture reveals differences in AKT–mTOR–S6K signaling and drug responses. J. Cell Sci. 2017, 130, 203–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, J.-z.; Xuan, Y.-y.; Zhang, Q.; Huang, J.-j. Ursolic acid sensitized colon cancer cells to chemotherapy under hypoxia by inhibiting MDR1 through HIF-1α. J. Zhejiang Univ.—Sci. B 2016, 17, 672–682. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.-X.; Wink, M. Reversal of Multidrug Resistance in Human Colon Cancer and Human Leukemia Cells by Three Plant Extracts and Their Major Secondary Metabolites. Medicines 2018, 5, 123. [Google Scholar] [CrossRef] [Green Version]
- Martins-Gomes, C.; Taghouti, M.; Schäfer, J.; Bunzel, M.; Silva, A.M.; Nunes, F.M. Chemical characterization and bioactive properties of decoctions and hydroethanolic extracts of Thymus carnosus Boiss. J. Funct. Foods 2018, 43, 154–164. [Google Scholar] [CrossRef]
- Taghouti, M.; Martins-Gomes, C.; Schäfer, J.; Félix, L.M.; Santos, J.A.; Bunzel, M.; Nunes, F.M.; Silva, A.M. Thymus pulegioides L. as a rich source of antioxidant, anti-proliferative and neuroprotective phenolic compounds. Food Funct. 2018, 9, 3617–3629. [Google Scholar] [CrossRef]
- Raudone, L.; Zymone, K.; Raudonis, R.; Vainoriene, R.; Motiekaityte, V.; Janulis, V. Phenological changes in triterpenic and phenolic composition of Thymus L. species. Ind. Crops Prod. 2017, 109, 445–451. [Google Scholar] [CrossRef]
- Kontogianni, V.G.; Tomic, G.; Nikolic, I.; Nerantzaki, A.A.; Sayyad, N.; Stosic-Grujicic, S.; Stojanovic, I.; Gerothanassis, I.P.; Tzakos, A.G. Phytochemical profile of Rosmarinus officinalis and Salvia officinalis extracts and correlation to their antioxidant and anti-proliferative activity. Food Chem. 2013, 136, 120–129. [Google Scholar] [CrossRef]
- Kalaycıoğlu, Z.; Uzaşçı, S.; Dirmenci, T.; Erim, F.B. α-Glucosidase enzyme inhibitory effects and ursolic and oleanolic acid contents of fourteen Anatolian Salvia species. J. Pharm. Biomed. Anal. 2018, 155, 284–287. [Google Scholar] [CrossRef]
- Chen, M.-C.; Hsu, H.-H.; Chu, Y.-Y.; Cheng, S.-F.; Shen, C.-Y.; Lin, Y.-J.; Chen, R.-J.; Viswanadha, V.P.; Lin, Y.-M.; Huang, C.-Y. Lupeol alters ER stress-signaling pathway by downregulating ABCG2 expression to induce Oxaliplatin-resistant LoVo colorectal cancer cell apoptosis. Environ. Toxicol. 2018, 33, 587–593. [Google Scholar] [CrossRef]
- Hu, T.; To, K.K.W.; Wang, L.; Zhang, L.; Lu, L.; Shen, J.; Chan, R.L.Y.; Li, M.; Yeung, J.H.K.; Cho, C.H. Reversal of P-glycoprotein (P-gp) mediated multidrug resistance in colon cancer cells by cryptotanshinone and dihydrotanshinone of Salvia miltiorrhiza. Phytomedicine 2014, 21, 1264–1272. [Google Scholar] [CrossRef]
- Su, C.-C. Tanshinone IIA potentiates the efficacy of 5-FU in Colo205 colon cancer cells in vivo through downregulation of P-gp and LC3-II. Exp. Ther. Med. 2012, 3, 555–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, P.; Vishwakarma, R.A.; Bharate, S.B. Natural alkaloids as P-gp inhibitors for multidrug resistance reversal in cancer. Eur. J. Med. Chem. 2017, 138, 273–292. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Wang, J.; Gao, W.; Liu, X.; Wu, S.; Wan, B.; Xu, L.; Li, Y. Salvianolic acid B reverses multidrug resistance in nude mice bearing human colon cancer stem cells. Mol. Med. Rep. 2018, 18, 1323–1334. [Google Scholar] [CrossRef] [Green Version]
- Guo, P.; Wang, S.; Liang, W.; Wang, W.; Wang, H.; Zhao, M.; Liu, X. Salvianolic acid B reverses multidrug resistance in HCT-8/VCR human colorectal cancer cells by increasing ROS levels. Mol. Med. Rep. 2017, 15, 724–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Zhang, L.; Ni, Z.; Sun, J.; Gao, H.; Cheng, Z.; Xu, J.; Yin, P. Resveratrol induces AMPK-dependent MDR1 inhibition in colorectal cancer HCT116/L-OHP cells by preventing activation of NF-κB signaling and suppressing cAMP-responsive element transcriptional activity. Tumor Biol. 2015, 36, 9499–9510. [Google Scholar] [CrossRef]
- Li, M.; Yin, D.; Li, J.; Shao, F.; Zhang, Q.; Jiang, Q.; Zhang, M.; Yang, Y. Rosmarinic acid, the active component of Salvia miltiorrhizae, improves gliquidone transport by regulating the expression and function of P-gp and BCRP in Caco-2 cells. Die Pharm.—Int. J. Pharm. Sci. 2020, 75, 18–22. [Google Scholar] [CrossRef]
- Wu, J.; Zhu, Y.; Li, F.; Zhang, G.; Shi, J.; Ou, R.; Tong, Y.; Liu, Y.; Liu, L.; Lu, L.; et al. Spica prunellae and its marker compound rosmarinic acid induced the expression of efflux transporters through activation of Nrf2-mediated signaling pathway in HepG2 cells. J. Ethnopharmacol. 2016, 193, 1–11. [Google Scholar] [CrossRef]
- Li, Y.; Revalde, J.L.; Reid, G.; Paxton, J.W. Interactions of dietary phytochemicals with ABC transporters: Possible implications for drug disposition and multidrug resistance in cancer. Drug Metab. Rev. 2010, 42, 590–611. [Google Scholar] [CrossRef]
- Critchfield, J.W.; Welsh, C.J.; Phang, J.M.; Chao Yeh, G. Modulation of adriamycin® accumulation and efflux by flavonoids in HCT-15 colon cells: Activation of P-glycoprotein as a putative mechanism. Biochem. Pharmacol. 1994, 48, 1437–1445. [Google Scholar] [CrossRef]
- Lohner, K.; Schnäbele, K.; Daniel, H.; Oesterle, D.; Rechkemmer, G.; Göttlicher, M.; Wenzel, U. Flavonoids alter P-gp expression in intestinal epithelial cells in vitro and in vivo. Mol. Nutr. Food Res. 2007, 51, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Yue, G.G.L.; Cheng, S.-W.; Yu, H.; Xu, Z.-S.; Lee, J.K.M.; Hon, P.-M.; Lee, M.Y.H.; Kennelly, E.J.; Deng, G.; Yeung, S.K.; et al. The Role of Turmerones on Curcumin Transportation and P-Glycoprotein Activities in Intestinal Caco-2 Cells. J. Med. Food 2011, 15, 242–252. [Google Scholar] [CrossRef]
- He, W.-t.; Zhu, Y.-h.; Zhang, T.; Abulimiti, P.; Zeng, F.-y.; Zhang, L.-p.; Luo, L.-j.; Xie, X.-m.; Zhang, H.-l. Curcumin Reverses 5-Fluorouracil Resistance by Promoting Human Colon Cancer HCT-8/5-FU Cell Apoptosis and Down-regulating Heat Shock Protein 27 and P-Glycoprotein. Chin. J. Integr. Med. 2019, 25, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.X.; Abulimiti, P.; Zhang, H.L.; Zhou, Y.K.; Zhu, L. Mechanism of reversal of multidrug resistance by curcumin in human colorectal cancer cell line HCT-8/5-FU. Genet. Mol. Res. GMR 2017, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Gao, M.; Wang, Z.; Zhang, J.; Cui, W.; Li, J.; Zhu, X.; Zhang, H.; Yang, D.-H.; Xu, X. Curcumin reverses doxorubicin resistance in colon cancer cells at the metabolic level. J. Pharm. Biomed. Anal. 2021, 201, 114129. [Google Scholar] [CrossRef]
- Neerati, P.; Sudhakar, Y.A.; Kanwar, J.R. Curcumin Regulates Colon Cancer by Inhibiting P-Glycoprotein in In-situ Cancerous Colon Perfusion Rat Model. J. Cancer Sci. 2013, 5, 313–319. [Google Scholar]
- Zhou, Y.; Zhang, J.; Wang, K.; Han, W.; Wang, X.; Gao, M.; Wang, Z.; Sun, Y.; Yan, H.; Zhang, H.; et al. Quercetin overcomes colon cancer cells resistance to chemotherapy by inhibiting solute carrier family 1, member 5 transporter. Eur. J. Pharmacol. 2020, 881, 173185. [Google Scholar] [CrossRef]
- Yuan, Z.; Shi, X.; Qiu, Y.; Jia, T.; Yuan, X.; Zou, Y.; Liu, C.; Yu, H.; Yuan, Y.; He, X.; et al. Reversal of P-gp-mediated multidrug resistance in colon cancer by cinobufagin. Oncol. Rep. 2017, 37, 1815–1825. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.-T.; Shi, X.-J.; Yuan, Y.-X.; Qiu, Y.-Y.; Zou, Y.; Liu, C.; Yu, H.; He, X.; Xu, K.; Yin, P.-H. Bufalin reverses ABCB1-mediated drug resistance in colorectal cancer. Oncotarget 2017, 8, 48012–48026. [Google Scholar] [CrossRef] [Green Version]
- Al-Abbasi, F.A.; Alghamdi, E.A.; Baghdadi, M.A.; Alamoudi, A.J.; El-Halawany, A.M.; El-Bassossy, H.M.; Aseeri, A.H.; Al-Abd, A.M. Gingerol Synergizes the Cytotoxic Effects of Doxorubicin against Liver Cancer Cells and Protects from Its Vascular Toxicity. Molecules 2016, 21, 886. [Google Scholar] [CrossRef]
- Mukkavilli, R.; Gundala, S.R.; Yang, C.; Donthamsetty, S.; Cantuaria, G.; Jadhav, G.R.; Vangala, S.; Reid, M.D.; Aneja, R. Modulation of Cytochrome P450 Metabolism and Transport across Intestinal Epithelial Barrier by Ginger Biophenolics. PLoS ONE 2014, 9, e108386. [Google Scholar] [CrossRef]
- Martins-Gomes, C.; Souto, E.B.; Silva, A.M. Chapter 15—Nanophytosomes: A novel approach for the delivery of herbal drugs. In Systems of Nanovesicular Drug Delivery; Nayak, A.K., Hasnain, M.S., Aminabhavi, T.M., Torchilin, V.P., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 239–257. [Google Scholar]
- Holst, B.; Williamson, G. Nutrients and phytochemicals: From bioavailability to bioefficacy beyond antioxidants. Curr. Opin. Biotechnol. 2008, 19, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Izzo, A.A. Interactions between Herbs and Conventional Drugs: Overview of the Clinical Data. Med. Princ. Pract. 2012, 21, 404–428. [Google Scholar] [CrossRef]
- Jermini, M.; Dubois, J.; Rodondi, P.-Y.; Zaman, K.; Buclin, T.; Csajka, C.; Orcurto, A.; Rothuizen, L.E. Complementary medicine use during cancer treatment and potential herb-drug interactions from a cross-sectional study in an academic centre. Sci. Rep. 2019, 9, 5078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.-W.; Fan, W.; Ko, S.-G.; Song, L.; Bian, Z.-X. Evidence-Based Management of Herb-Drug Interaction in Cancer Chemotherapy. Explore 2010, 6, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-Y.; Hsieh, C.-H.; Tsai, T.-H. Concurrent administration of anticancer chemotherapy drug and herbal medicine on the perspective of pharmacokinetics. J. Food Drug Anal. 2018, 26, S88–S95. [Google Scholar] [CrossRef]
- Herraez, E.; Gonzalez-Sanchez, E.; Vaquero, J.; Romero, M.R.; Serrano, M.A.; Marin, J.J.G.; Briz, O. Cisplatin-Induced Chemoresistance in Colon Cancer Cells Involves FXR-Dependent and FXR-Independent Up-Regulation of ABC Proteins. Mol. Pharm. 2012, 9, 2565–2576. [Google Scholar] [CrossRef]
- Husain, I.; Dale, O.R.; Martin, K.; Gurley, B.J.; Adams, S.J.; Avula, B.; Chittiboyina, A.G.; Khan, I.A.; Khan, S.I. Screening of medicinal plants for possible herb-drug interactions through modulating nuclear receptors, drug-metabolizing enzymes and transporters. J. Ethnopharmacol. 2023, 301, 115822. [Google Scholar] [CrossRef]
- Theile, D.; Wizgall, P. Acquired ABC-transporter overexpression in cancer cells: Transcriptional induction or Darwinian selection? Naunyn-Schmiedeberg’s Arch. Pharmacol. 2021, 394, 1621–1632. [Google Scholar] [CrossRef] [PubMed]
- Murthy, S.; Born, E.; Mathur, S.N.; Field, F.J. LXR/RXR activation enhances basolateral efflux of cholesterol in CaCo-2 cells. J. Lipid Res. 2002, 43, 1054–1064. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Flores, M.; Portilla-Martínez, A.; Cabrera-Pérez, F.; Nájera, N.; Meaney, E.; Villarreal, F.; Pérez-Durán, J.; Ceballos, G. PXR is a target of (-)-epicatechin in skeletal muscle. Heliyon 2020, 6, e05357. [Google Scholar] [CrossRef] [PubMed]
- Al-Dosari, M.S.; Parvez, M.K. Novel plant inducers of PXR-dependent cytochrome P450 3A4 expression in HepG2 cells. Saudi Pharm. J. 2018, 26, 1069–1072. [Google Scholar] [CrossRef]
- Harmsen, S.; Meijerman, I.; Febus, C.L.; Maas-Bakker, R.F.; Beijnen, J.H.; Schellens, J.H.M. PXR-mediated induction of P-glycoprotein by anticancer drugs in a human colon adenocarcinoma-derived cell line. Cancer Chemother. Pharmacol. 2010, 66, 765–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippert, A.; Renner, B. Herb–Drug Interaction in Inflammatory Diseases: Review of Phytomedicine and Herbal Supplements. J. Clin. Med. 2022, 11, 1567. [Google Scholar]
- NCT00920803—A Phase 1, Double-Blind, Randomized Clinical Study to Assess the Safety, Pharmacokinetics, and Pharmacodynamics of SRT501 in Subjects with Colorectal Cancer and Hepatic Metastases. Available online: https://clinicaltrials.gov/ct2/show/NCT00920803?term=resveratrol&cond=Colorectal+Cancer&draw=2&rank=2 (accessed on 4 March 2023).
- NCT00433576—Phase I Repeat-Dose Study of Resveratrol in Colorectal Cancer Patients: Tolerability, Target Tissue Levels and Pharmacodynamics. Available online: https://clinicaltrials.gov/ct2/show/NCT00433576?term=resveratrol&cond=Colorectal+Cancer&draw=2&rank=1 (accessed on 4 March 2023).
- NCT02891538—A Pilot Study to Evaluate the Chemopreventive Effects of Epigallocatechin Gallate (EGCG) in Colorectal Cancer (CRC) Patients With Curative Resections. Available online: https://clinicaltrials.gov/ct2/show/NCT02891538 (accessed on 3 March 2023).
- NCT01239095—Perioperative Administration of Oral Green Tea Extract/Milk Thistle Extract to Colorectal Cancer Patients Undergoing Colorectal Cancer Resection, a Phase 1 Study. Available online: https://clinicaltrials.gov/ct2/show/NCT01239095?term=egcg&cond=Colorectal+Cancer&draw=2&rank=3 (accessed on 3 March 2023).
- NCT02321969—Green Tea Extracts for the Prevention of Colorectal Adenomas and Colorectal Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT02321969?term=egcg&cond=Colorectal+Cancer&draw=2&rank=2 (accessed on 3 March 2023).
- NCT00027495—Phase I Pharmacokinetic Trial of Curcuminoids Administered in a Capsule Formulation. Available online: https://clinicaltrials.gov/ct2/show/NCT00027495?term=curcumin&cond=Colorectal+Cancer&draw=2&rank=6 (accessed on 3 March 2023).
- NCT00973869—A Pilot Study of Administration of Curcumin to Determine Colonic Curcumin Tissue Levels in Patients Awaiting Colorectal Endoscopy or Patients With Colorectal Cancer Awaiting Resection. Available online: https://clinicaltrials.gov/ct2/show/NCT00973869?term=curcumin&cond=Colorectal+Cancer&draw=2&rank=2 (accessed on 4 March 2023).
- NCT01344538—Phase II Study of the Effects of Ginger Root Extract on Eicosanoids in Colon Mucosa in People at Normal Risk for Colorectal Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT01344538?term=extract&cond=Colorectal+Cancer&draw=2&rank=2 (accessed on 5 March 2023).
- NCT01916239—Phase I–II Study of Pomegranate Extract Formulations in Colorectal Cancer Patients: Metabolic and Gene Expression Profiling in Tumoral and Normal Colon Tissues. Available online: https://clinicaltrials.gov/ct2/show/NCT01916239?term=extract&cond=Colorectal+Cancer&draw=2&rank=1 (accessed on 4 March 2023).
- Dhupal, M.; Chowdhury, D. Phytochemical-Based Nanomedicine for Advanced Cancer Theranostics: Perspectives on Clinical Trials to Clinical Use. Int. J. Nanomed. 2020, 15, 9125–9157. [Google Scholar] [CrossRef]
- Xie, J.; Yang, Z.; Zhou, C.; Zhu, J.; Lee, R.J.; Teng, L. Nanotechnology for the delivery of phytochemicals in cancer therapy. Biotechnol. Adv. 2016, 34, 343–353. [Google Scholar] [CrossRef]



| Phytochemical | MDR-Related Transporter Inhibition | Ref. | ||
|---|---|---|---|---|
| P-gp | MRP1 | ABCG2 | ||
| Epigallocatechin gallate | Caco-2 CHRC5 KB-C2 MCF-7 HepG2 | A549 DLD-1 HL-60 NB4 | MCF-7 | [55,56,57,58,59,60,61] |
| Quercetin | Caco-2 MCF-7 | Sf9 inside-out vesicles | HEK293 Sf9 inside-out vesicles | [62,63,64,65,66] |
| Ursolic acid | HCT-8 SW480 KB-C2 | HEK293 Ovarian CSCs MDCK | [47,66,67,68,69] | |
| Oleanolic acid | Ma104 SW982 SK-UT-1 | HEK293 | [47,70,71] | |
| Kaempferol | HEK293 MDCK | [47,72] | ||
| Gingerol | KB-C2 | PC-3 | [73,74] | |
| Curcumin | Caco-2 SiHa | SiHa MCF-7 MDA-MB-23 | MCF-7 MDA-MB-23 | [75,76,77] |
| Resveratrol | KB-C2 Caco-2 rat everted gut sac model | Caco-2 AML-2 | Pancreatic CSC Caco-2 | [78,79,80,81] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins-Gomes, C.; Silva, A.M. Natural Products as a Tool to Modulate the Activity and Expression of Multidrug Resistance Proteins of Intestinal Barrier. J. Xenobiot. 2023, 13, 172-192. https://doi.org/10.3390/jox13020014
Martins-Gomes C, Silva AM. Natural Products as a Tool to Modulate the Activity and Expression of Multidrug Resistance Proteins of Intestinal Barrier. Journal of Xenobiotics. 2023; 13(2):172-192. https://doi.org/10.3390/jox13020014
Chicago/Turabian StyleMartins-Gomes, Carlos, and Amélia M. Silva. 2023. "Natural Products as a Tool to Modulate the Activity and Expression of Multidrug Resistance Proteins of Intestinal Barrier" Journal of Xenobiotics 13, no. 2: 172-192. https://doi.org/10.3390/jox13020014