
Copper and Nickel Induce Changes in the Lipid and Fatty Acid Composition of Anodonta cygnea

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Design
2.2. Lipid Analysis
2.3. Statistical Analyses
3. Results and Discussion
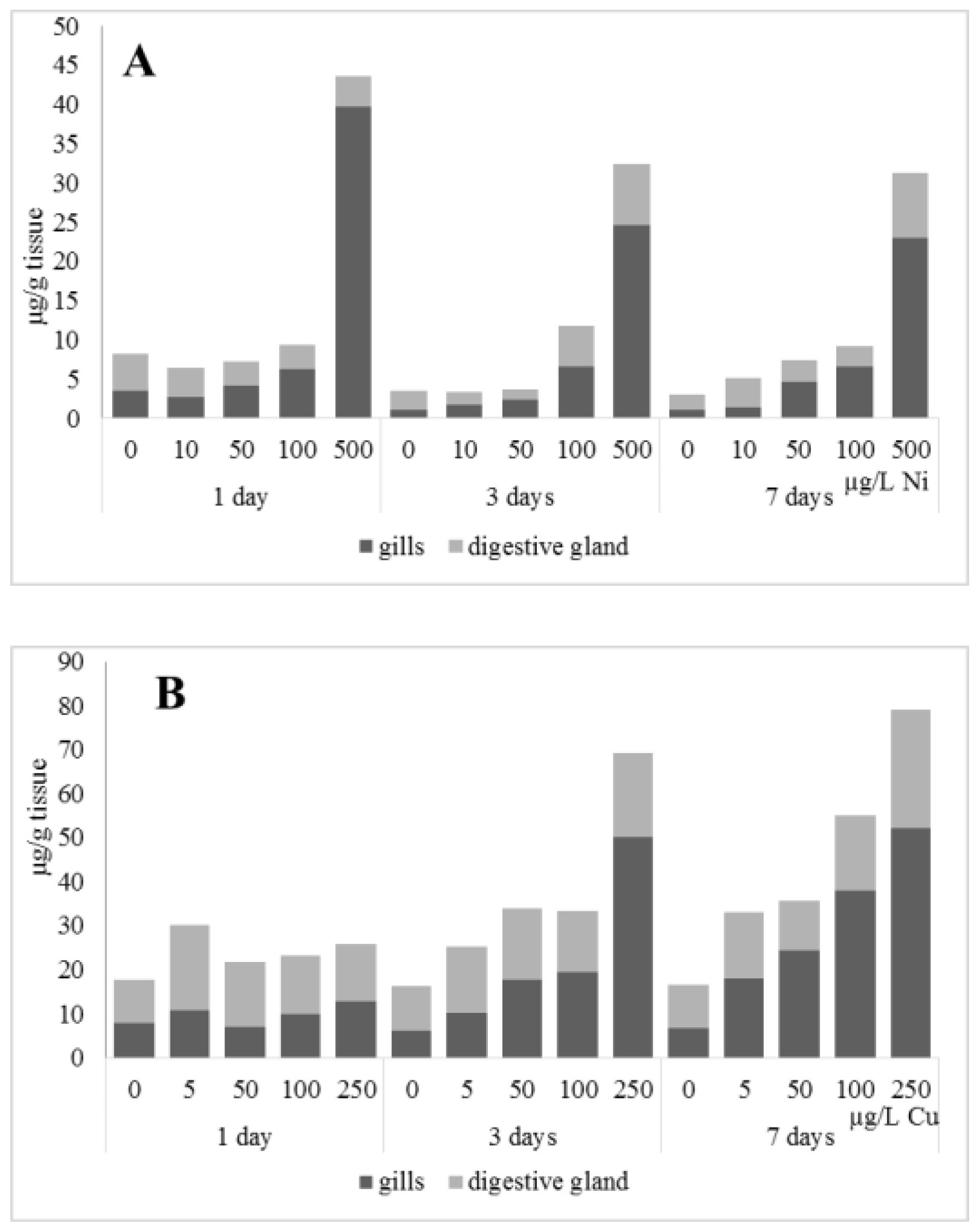
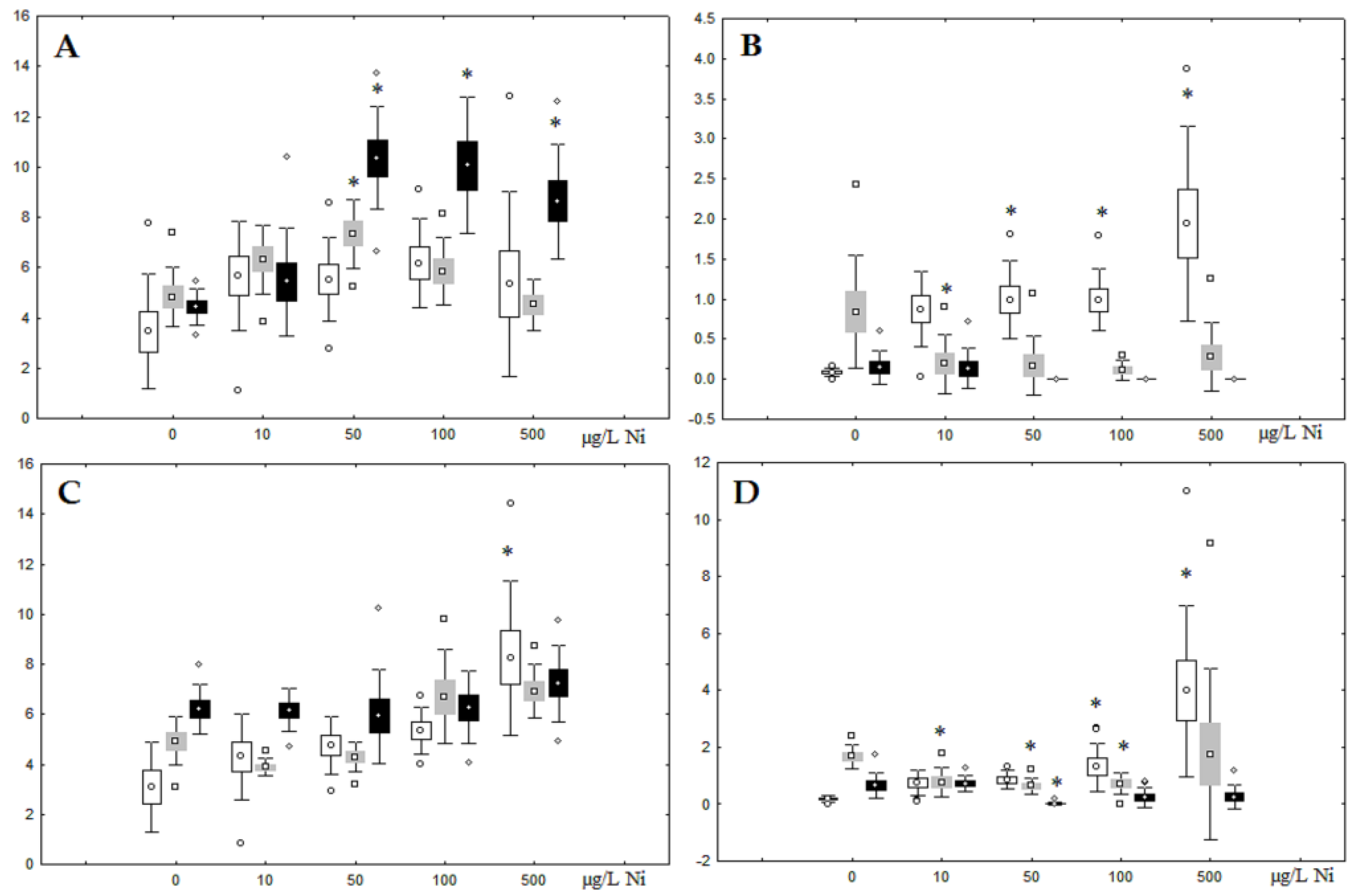
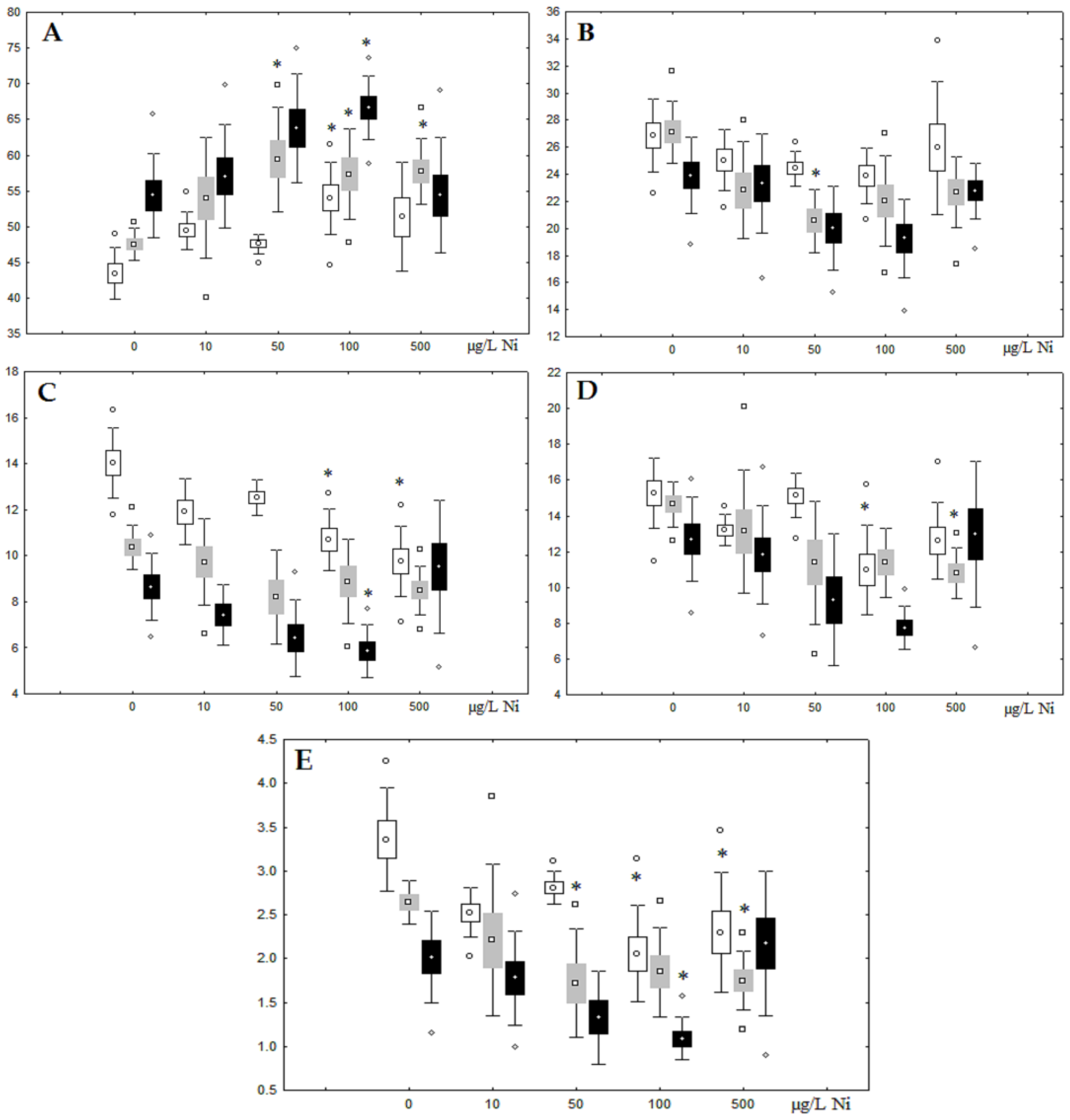
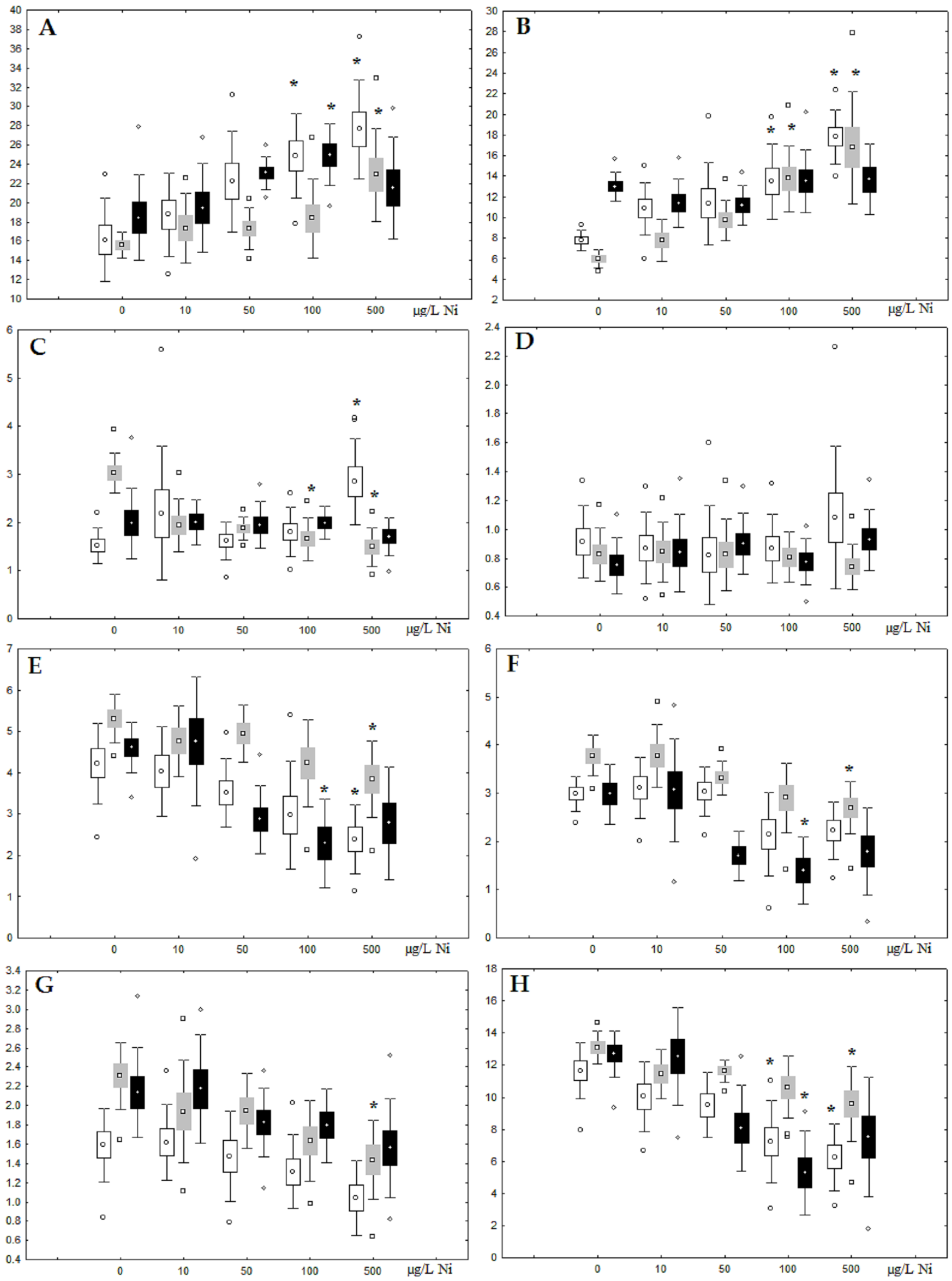
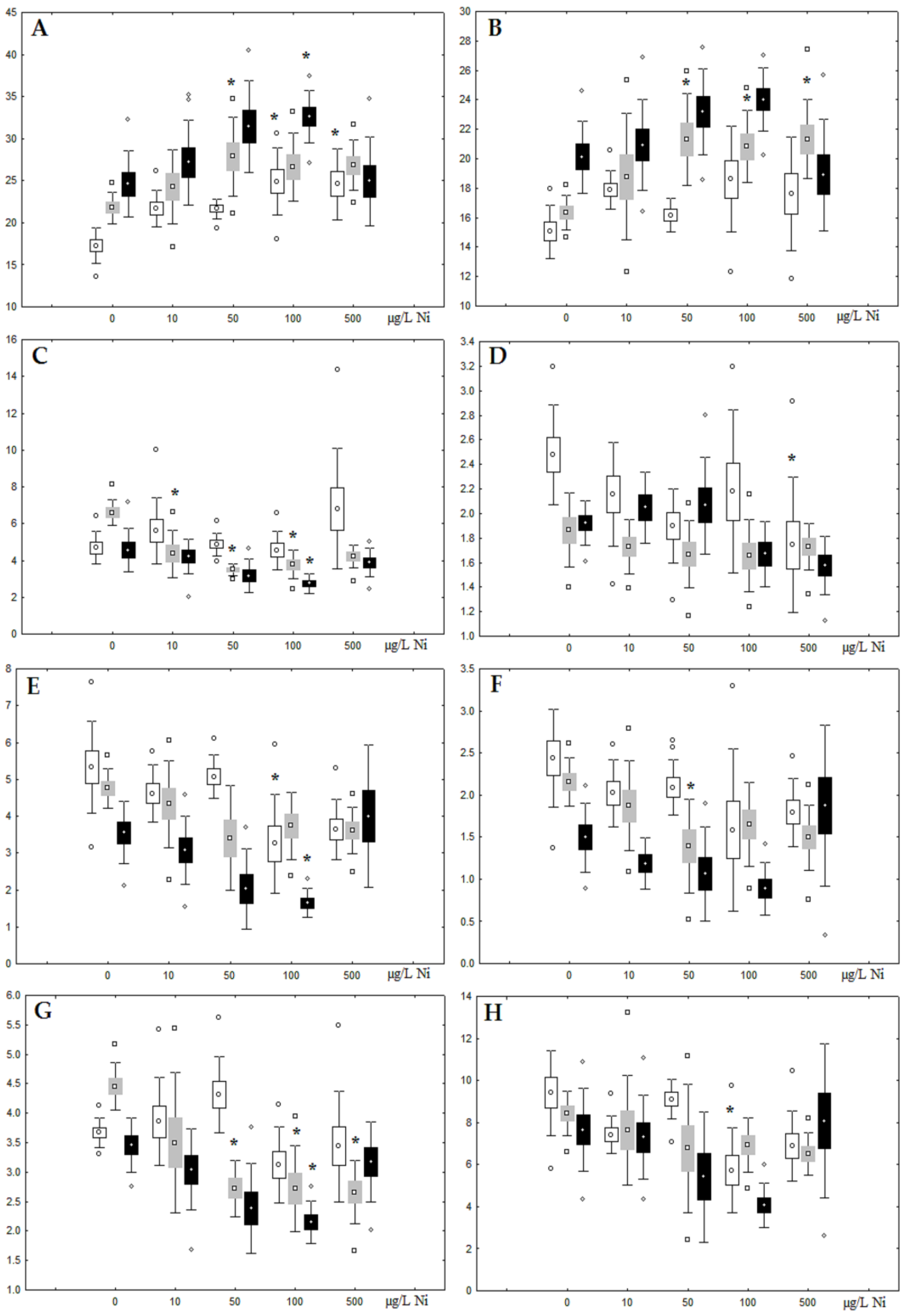
3.1. Effects of Nickel
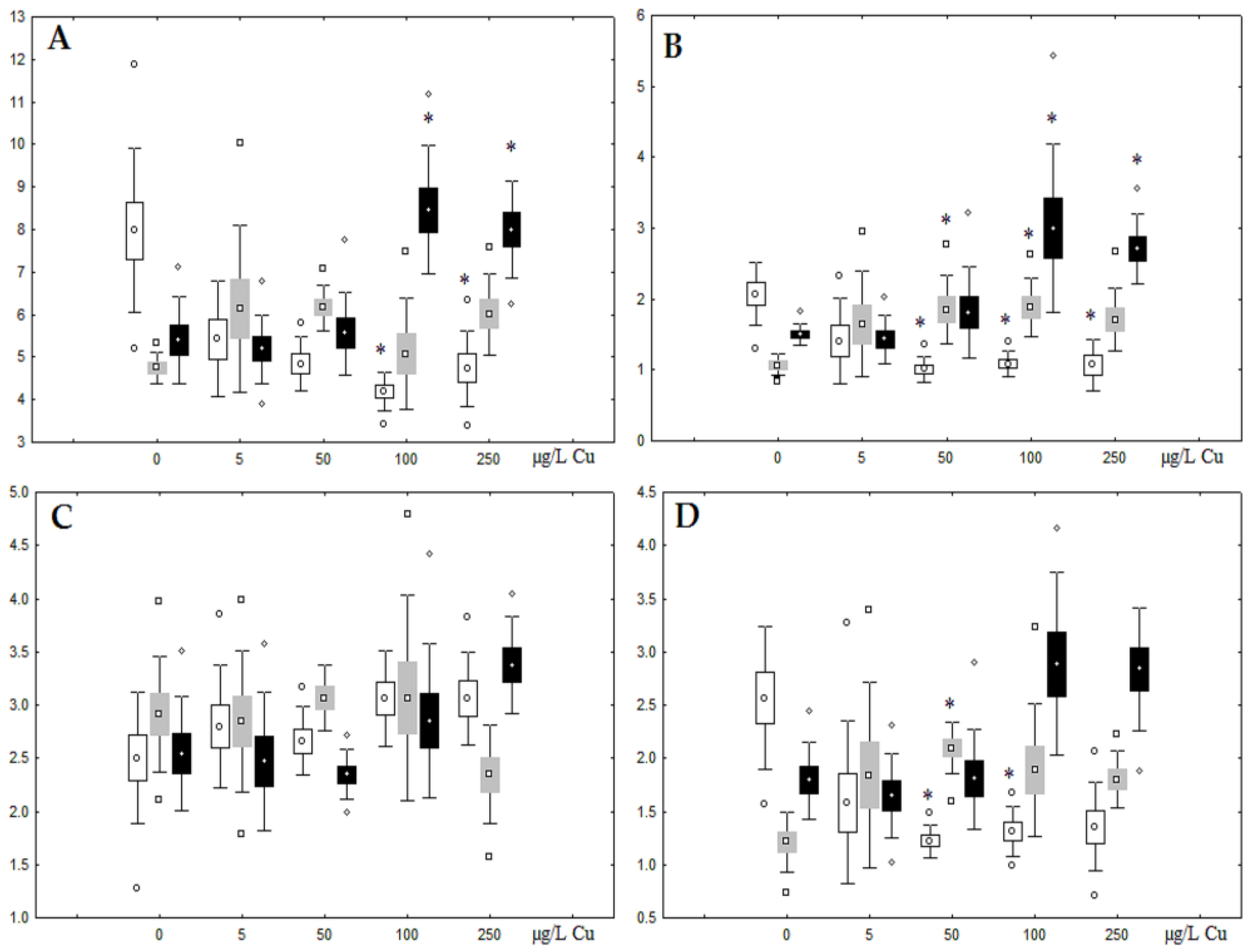
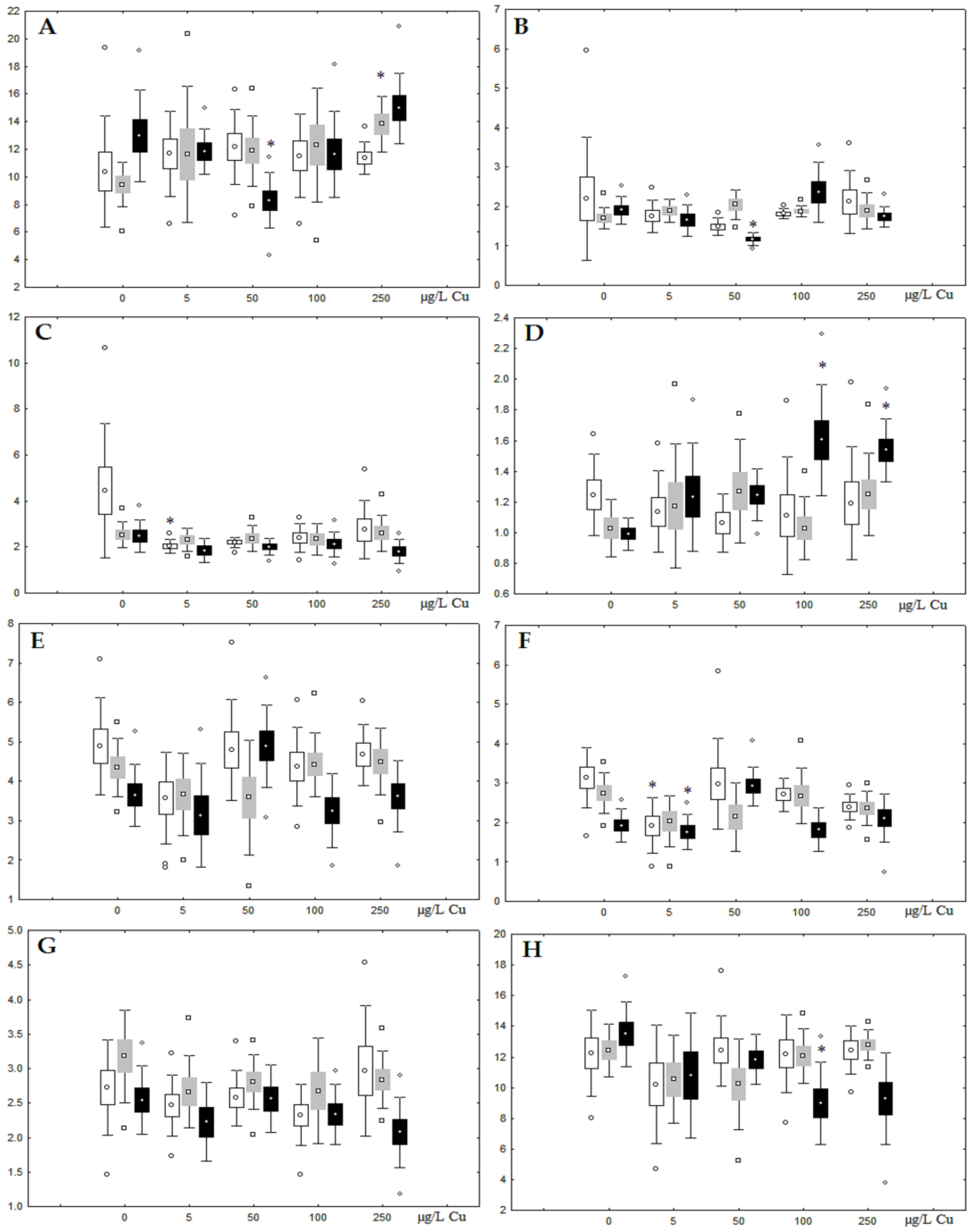
3.2. Copper Effects
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Štefanac, T.; Grgas, D.; Dragičević, T.L. Xenobiotics—Division and Methods of Detection: A Review. J. Xenobiotics 2021, 11, 130–141. [Google Scholar] [CrossRef]
- Duffus, J.H. “Heavy metals” a meaningless term? (IUPAC Technical Report). Pure Appl. Chem. 2002, 74, 793–807. [Google Scholar] [CrossRef] [Green Version]
- Company, R.; Serafim, A.; Cosson, R.P.; Fiala-Médioni, A.; Camus, L.; Colaço, A.; Bebianno, M.J. Antioxidant biochemical responses to long-term copper exposure in Bathymodiolus azoricus from Menez-Gwen hydrothermal vent. Sci. Total Environ. 2008, 389, 407–417. [Google Scholar] [CrossRef]
- Merian, E.; Anke, M.; Ihnat, M.; Stoeppler, M. (Eds.) Elements and Their Compounds in the Environment; Wiley-VHC Verlag: Weinheim, Germany, 2004; pp. 731–750. [Google Scholar]
- Nath, R. Copper deficiency and heart disease: Molecular basis, recent advances and current concepts. Int. J. Biochem. Cell Biol. 1997, 29, 1245–1254. [Google Scholar] [CrossRef]
- Lettieri, G.; Mollo, V.; Ambrosino, A.; Caccavale, F.; Troisi, J.; Febbraio, F.; Piscopo, M. Molecular effects of copper on the reproductive system of mytilus galloprovincialis. Mol. Reprod. Dev. 2019, 86, 1357–1368. [Google Scholar] [CrossRef]
- Piscopo, M.; Trifuoggi, M.; Scarano, C.; Gori, C.; Giarra, A.; Febbraio, F. Relevance of arginine residues in Cu (II)-induced DNA breakage and Proteinase K resistance of H1 histones. Sci. Rep. 2018, 8, 7414. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Li, M.; Zhong, Z.; Chen, H.; Wang, X.; Wang, M.; Xu, Z.; Cao, L.; Lian, C.; Zhang, H.; et al. Biochemical and metabolic responses of the deep-sea mussel Bathymodiolus platifrons to cadmium and copper exposure. Aquat. Toxicol. 2021, 236, 105845. [Google Scholar] [CrossRef]
- Tagliaferro, M.; Rocha, C.; Marques, J.C.; Gonçalves, A.M. Assessment of metal exposure (uranium and copper) in fatty acids and carbohydrate profiles of Calamoceras marsupus larvae (Trichoptera) and Alnus glutinosa leaf litter. Sci. Total Environ. 2022, 836, 155613. [Google Scholar] [CrossRef]
- Costa, M.; Salnikow, K.; Sutherland, J.E.; Broday, L.; Peng, W.; Zhang, Q.; Kluz, T. The role of oxidative stress in nickel and chromate genotoxicity. In Oxygen/Nitrogen Radicals: Cell Injury and Disease; Springer: Boston, MA, USA, 2002; pp. 265–275. [Google Scholar]
- Denkhaus, E.; Salnikow, K. Nickel essentiality, toxicity, and carcinogenicity. Crit. Rev. Oncol. Hematol. 2002, 42, 35–56. [Google Scholar] [CrossRef]
- Abele, D.; Vazquez-Medina, J.P.; Zenteno-Savin, T. Oxidative Stress in Aquatic Ecosystems; John Wiley & Sons: London, UK, 2011; 548p. [Google Scholar]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [Green Version]
- Fokina, N.N.; Ruokolainen, T.R.; Nemova, N.N.; Bakhmet, I.N. Changes of blue mussels Mytilus edulis L. lipid composition under cadmium and copper toxic effect. Biol. Trace Elem. Res. 2013, 154, 217–225. [Google Scholar] [CrossRef]
- Signa, G.; Di Leonardo, R.; Vaccaro, A.; Tramati, C.D.; Mazzola, A.; Vizzini, S. Lipid and fatty acid biomarkers as proxies for environmental contamination in caged mussels Mytilus galloprovincialis. Ecol. Indic. 2015, 57, 384–394. [Google Scholar] [CrossRef]
- Thompson, G.A., Jr. Metabolism and control of lipid structure modification. Biochem. Cell Biol. 1986, 64, 66–69. [Google Scholar] [CrossRef]
- Hellou, J.; Law, R.J. Stress on stress response of wild mussels, Mytilus edulis and Mytilus trossulus, as an indicator of ecosystem health. Environ. Pollut. 2003, 126, 407–416. [Google Scholar] [CrossRef]
- Vance, D.E.; Vance, J.E. (Eds.) Biochemistry of Lipids, Lipoproteins and Membranes, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Hochachka, P.M.; Somero, G.N. Biochemical Adaptation; Princeton University Press: Oxford, UK, 2002. [Google Scholar]
- Faggio, C.; Tsarpali, V.; Dailianis, S. Mussel digestive gland as a model tissue for assessing xenobiotics: An overview. Sci. Total Environ. 2018, 636, 220–229. [Google Scholar] [CrossRef]
- Kulikova, T.P. Zooplankton Vodnyh Ob’ektov Bassejna Onezhskogo Ozera; KarRC RAS: Petrozavodsk, Russia, 2007; 223p. (In Russian) [Google Scholar]
- Zorita, I.; Ortiz-Zarragoitia, M.; Soto, M.; Cajaraville, M.P. Biomarkers in mussels from a copper site gradient (Visnes, Norway): An integrated biochemical, histochemical and histological study. Aquat. Toxicol. 2006, 78, S109–S116. [Google Scholar] [CrossRef]
- Manly, R.; George, W.O. The occurrence of some heavy metals in populations of the freshwater mussel Anodonta anatina (L.) from the River Thames. Environ. Pollut. 1977, 14, 139–154. [Google Scholar] [CrossRef]
- Slukovskii, Z.I.; Polyakova, T.N. Analysis of accumulation of heavy metals from river bottom sediments of the urban environment in the bodies of oligochaetes. Inland Water Biol. 2017, 10, 315–322. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloan-Stanley, G.H. A simple method for the isolation and purification of total lipids animal tissue (for brain, liver and muscle). J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Sidorov, V.S.; Lizenko, E.I.; Bolgova, O.M.; Nefedova, Z.A. Fish Lipids. 1. Analysis Technique; Karelian Branch of the USSR Academy of Science: Petrozavodsk, Russia, 1972; pp. 150–162. (In Russian) [Google Scholar]
- Engelbrecht, F.M.; Mari, F.; Anderson, J.T. Cholesterol. Determination in serum. A rapid direction method. S. Afr. Med. J. 1974, 48, 250–256. [Google Scholar]
- Cosson, R.P.; Thiébaut, É.; Company, R.; Castrec-Rouelle, M.; Colaço, A.; Martins, I.; Bebianno, M.J. Spatial variation of metal bioaccumulation in the hydrothermal vent mussel Bathymodiolus azoricus. Mar. Environ. Res. 2008, 65, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Rajalakshmi, S.; Mohandas, A. Copper-induced changes in tissue enzyme activity in a freshwater mussel. Ecotoxicol. Environ. Saf. 2005, 62, 140–143. [Google Scholar] [CrossRef]
- Bourgeault, A.; Gourlay-Francé, C.; Ayrault, S.; Tusseau-Vuillemin, M.H. Bioaccumulation of waterborne Ni in Dreissena polymorpha: A stable isotope experiment to assess the effect of zinc, calcium, and dissolved organic matter. Environ. Toxicol. Chem. 2012, 31, 819–827. [Google Scholar] [CrossRef]
- Perić, L.; Nerlović, V.; Žurga, P.; Žilić, L.; Ramšak, A. Variations of biomarkers response in mussels Mytilus galloprovincialis to low, moderate and high concentrations of organic chemicals and metals. Chemosphere 2017, 174, 554–562. [Google Scholar] [CrossRef]
- Moore, M.N.; Viarengo, A.; Donkin, P.; Hawkins, A.J. Autophagic and lysosomal reactions to stress in the hepatopancreas of blue mussels. Aquat. Toxicol. 2007, 84, 80–91. [Google Scholar] [CrossRef]
- Chan, C.Y.; Wang, W.X. A lipidomic approach to understand copper resilience in oyster Crassostrea hongkongensis. Aquat. Toxicol. 2018, 204, 160–170. [Google Scholar] [CrossRef]
- Wang, W.X.; Yang, Y.; Guo, X.; He, M.; Guo, F.; Ke, C. Copper and zinc contamination in oysters: Subcellular distribution and detoxification. Environ. Toxicol. Chem. 2011, 30, 1767–1774. [Google Scholar] [CrossRef]
- Fokina, N.; Vasil’eva, O.; Sukhovskaya, I.; Kurpe, S. Cd and Ni modulate fatty acid composition and oxidative status in the freshwater mussel Anodonta cygnea. Toxicol. Environ. Health Sci. 2020, 12, 169–176. [Google Scholar] [CrossRef]
- Fokina, N.N.; Sukhovskaja, I.V.; Vasil’eva, O.B.; Nemova, N.N. Changes in the Gill Lipid Composition of Freshwater Mussel Anodonta cygnea under the Influence of Copper in Various Concentrations. Inland Water Biol. 2020, 13, 655–663. [Google Scholar] [CrossRef]
- Kowalczyk-Pecka, D.; Kowalczuk-Vasilev, E.; Pecka, S. The effect of heterogeneous copper micro-supplementation on fatty acid profiles in the tissues of snails Helix pomatia (Gastropoda Pulmonata). Ecol. Indic. 2017, 76, 335–343. [Google Scholar] [CrossRef]
- Kowalczyk-Pecka, D.; Kowalczuk-Vasilev, E.; Puchalski, A.; Klebaniuk, R. Peroxidation and unsaturation indices as potential biomarkers of multifarious zinc and copper micro-supplementation in Helix pomatia L. Ecotoxicol. Environ. Saf. 2018, 148, 713–720. [Google Scholar] [CrossRef]
- Maazouzi, C.; Masson, G.; Izquierdo, M.S.; Pihan, J.C. Chronic copper exposure and fatty acid composition of the amphipod Dikerogammarus villosus: Results from a field study. Environ. Pollut. 2008, 156, 221–226. [Google Scholar] [CrossRef]
- Nugroho, A.P.; Frank, H. Uptake, distribution, and bioaccumulation of copper in the freshwater mussel Anodonta anatina. Toxicol. Environ. Chem. 2011, 93, 1838–1850. [Google Scholar] [CrossRef]
- Millward, G.E.; Kadam, S.; Jha, A.N. Tissue-specific assimilation, depuration and toxicity of nickel in Mytilus edulis. Environ. Pollut. 2012, 162, 406–412. [Google Scholar] [CrossRef]
- Blewett, T.A.; Wood, C.M. Salinity-dependent nickel accumulation and oxidative stress responses in the euryhaline killifish (Fundulus heteroclitus). Arch. Environ. Contam. Toxicol. 2015, 68, 382–394. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fokina, N. Copper and Nickel Induce Changes in the Lipid and Fatty Acid Composition of Anodonta cygnea. J. Xenobiot. 2023, 13, 132-147. https://doi.org/10.3390/jox13010011
Fokina N. Copper and Nickel Induce Changes in the Lipid and Fatty Acid Composition of Anodonta cygnea. Journal of Xenobiotics. 2023; 13(1):132-147. https://doi.org/10.3390/jox13010011
Chicago/Turabian StyleFokina, Natalia. 2023. "Copper and Nickel Induce Changes in the Lipid and Fatty Acid Composition of Anodonta cygnea" Journal of Xenobiotics 13, no. 1: 132-147. https://doi.org/10.3390/jox13010011