
Glyphosate vs. Glyphosate-Based Herbicides Exposure: A Review on Their Toxicity


Abstract
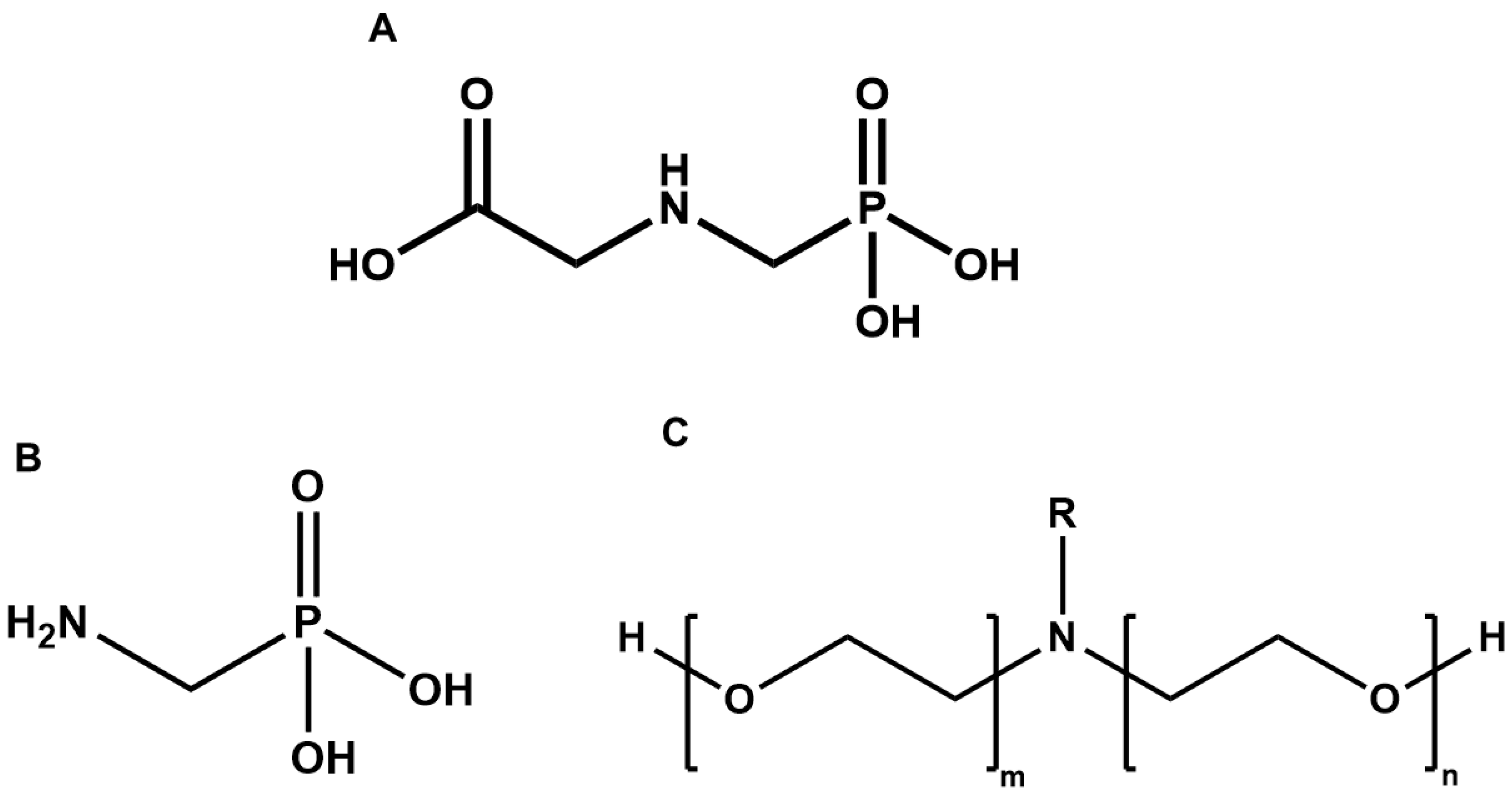
:1. Introduction
2. Regulatory Measures on Glyphosate Use
3. Environmental Persistence of Glyphosate and AMPA
4. In Vitro and In Vivo Studies to Assess Animal Toxicity (2010–2021)

{kind=link}
| Model | Exposure Time | Tested Concentrations | Effects | Ref. | |
|---|---|---|---|---|---|
| Glyphosate (99.8%) | Cherax quadricarinatus | 60 days | 10 and 40 mg/L | Decrease in lipid levels in muscle, as well as protein level in hepatopancreas and muscle | [83] |
| Glyphosate (99%) | Danio rerio | 96 h | 1.7–100 mg/L | Genotoxicity, morphological abnormalities | [84] |
| Glyphosate (˃98%) | Hormone-dependent breast cancer (T47D cell line) | 24 h | 10−9–10−3 mM | Increase in cell proliferation | [85] |
| Glyphosate (95%) | Human keratinocytes (HaCaT cell line) | 24 h | 10–70 mM | Loss of cell integrity, overproduction of H2O2, membrane damage, apoptosis induction, genotoxicity | [86] |
| Glyphosate (95%) | Buccal epithelial cells (TR146 cell line) | 20 min | >10 mg/L | Increased lactate dehydrogenase release, DNA damage | [87] |
| Glyphosate (99%) | Human hepatocarcinoma (HepG2 cell line) | 4 and 24 h | 0.5–3.5 µg/mL | Micronuclei formation, lower antioxidant capacity | [80] |
| Glyphosate (90%) | Sprague Dawley rat | 5 weeks | 5–500 mg/kg | Decreased average daily feed intake and decreased total sperm count | [88] |
| Glyphosate (not specified) | Induced pluripotent stem cells (iPSCs) | 24 h | 1–1000 µM | Increase in blood–brain barrier permeability to fluorescein, changes in neuronal cells metabolic activity and increase of glucose uptake in brain’s microvascular endothelial cells | [89] |
| Glyphosate (40%) | Daphnia magna | 60 days | 0.5–4.05 mg/L of glyphosate | Reduction of juvenile’s size, decreased fecundity and longevity | [90] |
| Glyphosate (not specified) | Danio rerio | 21 days | 10–100 mg/L of glyphosate | Reduced egg production, increase in early-stage embryo mortalities and premature hatching, disruption of the steroidogenic biosynthesis pathway, oxidative stress | [91] |
| Glyphosate (not specified) | Danio rerio | 48 h | 50 µg/mL of glyphosate | Structural abnormalities in the atrium and ventricle, irregular heart looping, situs inversus and decreased heartbeats | [92] |
| Glyphosate (not specified) | Danio rerio | 15 days | 65 µg/mL of glyphosate | Increase in oocytes’ diameter, presence of concentric membranes appearing as myelin-like structures, increase in expression of SF-1 in oocytes | [93] |
| Glyphosate (not specified) | Danio rerio | 96 h | 0.01–0.5 mg/L of glyphosate | Decrease locomotion in adult zebrafish, decreased ocular distance in zebrafish larvae | [94] |
| Glyphosate (not specified) | Sprague Dawley rat | 2 weeks | 50–150 mg/kg of glyphosate | Hypoactivity, decrease in specific binding to D1 dopamine receptors in the nucleus accumbens, decrease in basal extracellular dopamine levels and high-potassium-induced dopamine release in striatum | [95] |
| Roundup (480 g/L) | Piaractus mesopotamicus | 48 h | 3.0–4.5 mg /L of glyphosate | Cytoplasmic vacuolization, lipid accumulation, nuclear and cellular membrane alterations and glycogen depletion in the liver | [96] |
| Touchdown® (523 g/L) | Caenorhabditis elegans | 30 min | 3–10% of glyphosate | Inhibition of mitochondria’s complex II, decrease in ATP levels, increase in H2O2 levels | [97] |
| Roundup (410 g/L) | Human alveolar carcinoma (A549 cell line) | 2 h | 100 µg/L | Inhibition of cell proliferation, collapse of mitochondrial membrane, oxidative DNA damage, DNA single-strand breaks and double-strand breaks | [98] |
| Roundup (180 g/L) | Daphnia magna | 60 days | 0.5–4.05 mg/L of glyphosate | Reduction of juvenile size, growth, fecundity and increased abortion | [90] |
| Roundup (180 g/L) | Drosophila melanogaster | 24 h | 15 µg/mL | Decreased lifespan, fecundity, cell viability of ovarian sheath cells, negative geotaxis response, increase in protein carboxyl levels and enhanced caspase activity indicative of pro-apoptotic process | [99] |
| Herbolex (486 g/L) | Daphnia magna | 48 h | 20–137 µg/L | Increased lipid peroxidation, feed inhibition, increase in antioxidant enzyme activity | [100] |
| Roundup (480 mg/L) | Poecilia reticulata | 96 h | 0.34–5.2 mg/L of glyphosate | Modulation of energy and nucleic acids metabolism, cytoskeleton and proteins; progressive histopathological damage in the gills | [101] |
| Roundup (450 g/L) | Buccal epithelial cells (TR146 cell line) | 20 min | >10 mg/L of glyphosate | Increase in nucleoplasmatic bridges, nuclear aberrations and micronuclei | [87] |
| Roundup (360 g/L) | Albino rats | 12 weeks | 3.6–248.4 mg/kg of glyphosate | Accumulation of glyphosate residue in kidney tissue, histopathological lesions in kidneys, distorted renal cortical histoarchitecture, expanded urinary space due to glomerulosclerosis, and tubular necrosis | [102] |
| Roundup (360 g/L) | Mice | 6 and 12 weeks | 250 or 500 mg/kg/day | Decrease in body weight gain and locomotor activity, increase of anxiety and depression-like behavior levels | [103] |
| Roundup (360 g/L) | Albino rats | 12 weeks | 3.6–248.4 mg/kg/day of glyphosate | Decrease in the mean level of testosterone, FHS and LH in the blood, and increase of prolactin, excessive production of ROS, reduction in sperm count, percentage mobility and increase in abnormal sperm cells, degenerative testicular lesions | [104] |
| Roundup (360 g/L) | Anguilla anguilla | 1 and 3 days | 18 and 36 µg/L | Increment of catalase activity in gills, decrease of superoxide dismutase activity in liver, increase in DNA damage | [105] |
| Roundup (360 g/L) | Murine Sertoli cells (TM4 cell line) | 24 h | 10–10,000 mg/L | Decrease of succinate dehydrogenase activity, inhibition of glutathione-S-transferase, disruption of cell detoxification systems, increase of cytoplasmatic lipid droplets | [106] |
| Roundup (360 g/L) | Colossoma macropomum | 96 h | 10 and 15 mg/L of glyphosate | Alterations in respiratory epithelium structure, changes in hematological parameters, increase ROS production, increase in DNA damage in red blood cells and inhibition of cholinesterase activity in fish brain | [107] |
| Roundup (120 g/L) | Danio rerio | 21 days | 0.01–10 mg/L of glyphosate | Increase in early-stage embryo mortalities and premature hatching, disruption of the steroidogenic biosynthesis pathway, oxidative stress | [91] |
| Roundup (120 g/L) | Daphnia magna | 48 h | 100–300 mg/L | Loss of whole body enzyme activity and loss of cells integrity | [108] |
| Roundup | Danio rerio | 96 h | 0.01–0.5 mg/L | Decrease in locomotion in adult zebrafish, ocular distance in zebrafish larvae and decrease in aggressive behavior in adult zebrafish, impairment in memory in adult zebrafish | [94] |
| AMPA | Daphnia magna | 21 days | 7.4–120 mg/L | Decreased neonate production | [109] |
| AMPA | Bufo spinosus | 16 days | 0.07–3.6 µg/L | Decrease in embryonic survival, development delay, modification of body morphology | [110] |
| AMPA | Danio rerio | 24–96 h | 1.7–100 mg/L | Genotoxicity, morphological abnormalities | [84] |
| AMPA | Induced pluripotent stem cells (iPSCs) | 24 h | 0.1–1000 µM | Increase in blood–brain barrier permeability to fluorescein, changes in neuronal cells metabolic activity and glucose uptake in brain microvascular endothelial cells | [89] |
| AMPA | Human erythrocytes | 4 and 24 h | 0.01–5 mM | Increased ROS production, hemolysis and hemoglobin oxidation | [111] |
| AMPA | Paracentrotus lividus | 24 and 48 h | 1–100 µg/L | Development delay, increase of respiration rate, reduction in larvae size | [112] |
5. Toxic Effect of Surfactants Used in Glyphosate-Based Herbicides
| Model | Exposure Time | Tested Concentrations | Effects | Ref. |
|---|---|---|---|---|
| Wistar rat | 15 min | 1.28–800 mg/L | Disturbances of the spontaneous motoric activity of isolated jejunum segments | [114] |
| Crassostrea gigas | 35 days | 0.1–100 µg/L | Delay in gametogenesis, connective tissue destructuration, atrophies of the wall of digestive tubules | [115] |
| Danio rerio | 24–96 h | 0.4–16 mg/L | Genotoxicity, morphological abnormalities | [84] |
| Murine Sertoli cells (TM4 cell line) | 24 h | >0.01% | High cytotoxicity (0% cell viability) | [106] |
| Drosophila melanogaster | 24 h | 45 µg/mL | Decrease in lifespan, negative geotaxis response, increase in protein carboxyl levels, decrease in fecundity, decrease of ovarian sheath cells viability, enhanced caspase activity indicative of pro-apoptotic process | [99] |
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Gianessi, L.P. The increasing importance of herbicides in worldwide crop production. Pest Manag. Sci. 2013, 69, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Kraehmer, H.; Laber, B.; Rosinger, C.; Schulz, A. Herbicides as Weed Control Agents: State of the Art: I. Weed Control Research and Safener Technology: The Path to Modern Agriculture. Plant Physiol. 2014, 166, 1119–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, A.E.S.; Grillo, R.; Mello, N.F.S.; Rosa, A.H.; Fraceto, L.F. Application of poly(epsilon-caprolactone) nanoparticles containing atrazine herbicide as an alternative technique to control weeds and reduce damage to the environment. J. Hazard. Mater. 2014, 268, 207–215. [Google Scholar] [CrossRef]
- Trebst, A. Chapter 8—The Mode of Action of Triazine Herbicides in Plants, in the Triazine Herbicides; LeBaron, H.M., McFarland, J.E., Burnside, O.C., Eds.; Elsevier: Amesterdam, The Netherlands, 2008; pp. 101–110. [Google Scholar] [CrossRef]
- Chen, Z.; Zou, Y.; Wang, J.; Li, M.; Wen, Y. Phytotoxicity of chiral herbicide bromacil: Enantioselectivity of photosynthesis in Arabidopsis thaliana. Sci. Total Environ. 2016, 548–549, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, T.R. Mechanisms of resistance to paraquat in plants. Pest Manag. Sci. 2014, 70, 1316–1323. [Google Scholar] [CrossRef]
- Green, J.M. Current state of herbicides in herbicide-resistant crops. Pest Manag. Sci. 2014, 70, 1351–1357. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Mohsin, S.M.; Bhuyan, M.H.M.B.; Bhuiyan, T.F.; Anee, T.I.; Masud, A.A.C.; Nahar, K. Chapter 3—Phytotoxicity, environmental and health hazards of herbicides: Challenges and ways forward. In Agrochemicals Detection, Treatment and Remediation; Prasad, M.N.V., Ed.; Butterworth-Heinemann: Hyderabad, India, 2020; pp. 55–99. [Google Scholar] [CrossRef]
- Jablonowski, N.D.; Schäffer, A.; Burauel, P. Still present after all these years: Persistence plus potential toxicity raise questions about the use of atrazine. Environ. Sci. Pollut. Res. 2011, 18, 328–331. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Kumar, V.; Chauhan, A.; Datta, S.; Wani, A.B.; Singh, N.; Singh, J. Toxicity, degradation and analysis of the herbicide atrazine. Environ. Chem. Lett. 2018, 16, 211–237. [Google Scholar] [CrossRef]
- Kucka, M.; Pogrmic-Majkic, K.; Fa, S.; Stojilkovic, S.S.; Kovacevic, R. Atrazine acts as an endocrine disrupter by inhibiting cAMP-specific phosphodiesterase-4. Toxicol. Appl. Pharmacol. 2012, 265, 19–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos-Pereira, F.D.; Oliveira, C.A.; Pigoso, A.A.; Silva-Zacarin, E.C.; Barbieri, R.; Spatti, E.F.; Marin-Morales, M.A.; Severi-Aguiar, G.D. Severi-Aguiar, Early cytotoxic and genotoxic effects of atrazine on Wistar rat liver: A morphological, immunohistochemical, biochemical, and molecular study. Ecotoxicol. Environ. Saf. 2012, 78, 170–177. [Google Scholar] [CrossRef]
- Jowa, L.; Howd, R. Should Atrazine and Related Chlorotriazines Be Considered Carcinogenic for Human Health Risk Assessment? J. Environ. Sci. Health Part C 2011, 29, 91–144. [Google Scholar] [CrossRef]
- Olejnik, A.M.; Marecik, R.; Białas, W.; Cyplik, P.; Grajek, W. In Vitro Studies on Atrazine Effects on Human Intestinal Cells. Water Air Soil Pollut. 2010, 213, 401–411. [Google Scholar] [CrossRef]
- Powell, E.R.; Faldladdin, N.; Rand, A.D.; Pelzer, D.; Schrunk, E.M.; Dhanwada, K.R. Dhanwada, Atrazine exposure leads to altered growth of HepG2 cells. Toxicol. Vitr. 2011, 25, 644–651. [Google Scholar] [CrossRef]
- Park, S.K.; Kang, D.; Beane-Freeman, L.; Blair, A.; Hoppin, J.A.; Sandler, D.P.; Lynch, C.F.; Knott, C.; Gwak, J.; Alavanja, M. Cancer Incidence among Paraquat Exposed Applicators in the Agricultural Health Study: A Prospective Cohort Study. Int. J. Occup. Environ. Health 2009, 15, 274–281. [Google Scholar] [CrossRef]
- Silva, A.M.; Silva, S.; Soares, J.P.; Martins-Gomes, C.; Teixeira, J.P.; Leal, F.; Gaivão, I. Ginkgo biloba L. Leaf Extract Protects HepG2 Cells Against Paraquat-Induced Oxidative DNA Damage. Plants 2019, 8, 556. [Google Scholar] [CrossRef] [Green Version]
- Awadalla, E.A. Efficacy of vitamin C against liver and kidney damage induced by paraquat toxicity. Exp. Toxicol. Pathol. 2012, 64, 431–434. [Google Scholar] [CrossRef]
- Blanco-Ayala, T.; Andérica-Romero, A.C.; Pedraza-Chaverri, J. New insights into antioxidant strategies against paraquat toxicity. Free Radic. Res. 2014, 48, 623–640. [Google Scholar] [CrossRef] [PubMed]
- Suntres, Z.E. Exploring the potential benefit of natural product extracts in paraquat toxicity. Fitoterapia 2018, 131, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Ohtaki, K.; Matsubara, K.; Aoyama, K.; Uezono, T.; Saito, O.; Suno, M.; Ogawa, K.; Hayase, N.; Kimura, K.; et al. Carrier-mediated processes in blood–brain barrier penetration and neural uptake of paraquat. Brain Res. 2001, 906, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Tatjana, V.; Domitille, S.; Jean-Charles, S. Paraquat-induced cholesterol biosynthesis proteins dysregulation in human brain microvascular endothelial cells. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Anderson, T.; Merrill, A.K.; Eckard, M.L.; Marvin, E.; Conrad, K.; Welle, K.; Oberdörster, G.; Sobolewski, M.; Cory-Slechta, D.A. Paraquat Inhalation, a Translationally Relevant Route of Exposure: Disposition to the Brain and Male-Specific Olfactory Impairment in Mice. Toxicol. Sci. 2020, 180, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Febrero, R.; Salvador, J.-P.; Sanchez-Baeza, F.; Marco, M.-P. Rapid method based on immunoassay for determination of paraquat residues in wheat, barley and potato. Food Control 2014, 41, 193–201. [Google Scholar] [CrossRef]
- Ghisi, N.D.C.; Zuanazzi, N.R.; Fabrin, T.M.C.; Oliveira, E.C. Glyphosate and its toxicology: A scientometric review. Sci. Total Environ. 2020, 733, 139359. [Google Scholar] [CrossRef]
- Benbrook, C.M. Trends in glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Duke, S.O. The history and current status of glyphosate. Pest Manag. Sci. 2018, 74, 1027–1034. [Google Scholar] [CrossRef]
- Székács, A.; Darvas, B. Forty years with glyphosate. In Herbicides-Properties, Synthesis and Control of Weeds; IntechOpen: London, UK, 2012; Volume 14, pp. 247–284. Available online: https://www.intechopen.com/chapters/25624 (accessed on 1 December 2021).
- Dill, G.M.; Sammons, R.D.; Feng, P.C.C.; Kohn, F.; Kretzmer, K.; Mehrsheikh, A.; Bleeke, M.; Honegger, J.L.; Farmer, D.; Wright, D.; et al. Glyphosate: Discovery, Development, Applications, and Properties. In Glyphosate Resistance in Crops and Weeds; Wiley: Hoboken, NJ, USA, 2010; pp. 1–33. [Google Scholar] [CrossRef]
- Sammons, R.D.; Gaines, T.A. Glyphosate resistance: State of knowledge. Pest Manag. Sci. 2014, 70, 1367–1377. [Google Scholar] [CrossRef] [Green Version]
- Cuhra, M.; Bøhn, T.; Cuhra, P. Glyphosate: Too Much of a Good Thing? Front. Environ. Sci. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Heap, I.; Duke, S.O. Overview of glyphosate-resistant weeds worldwide. Pest Manag. Sci. 2018, 74, 1040–1049. [Google Scholar] [CrossRef]
- Green, J.M. The rise and future of glyphosate and glyphosate-resistant crops. Pest Manag. Sci. 2016, 74, 1035–1039. [Google Scholar] [CrossRef]
- Fogliatto, S.; Ferrero, A.; Vidotto, F. Current and future scenarios of glyphosate use in Europe: Are there alternatives? In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2020; Volume 163, pp. 219–278. [Google Scholar] [CrossRef]
- Kudsk, P.; Mathiassen, S.K. Pesticide regulation in the European Union and the glyphosate controversy. Weed Sci. 2020, 68, 214–222. [Google Scholar] [CrossRef]
- Kier, L.D.; Kirkland, D.J. Review of genotoxicity studies of glyphosate and glyphosate-based formulations. Crit. Rev. Toxicol. 2013, 43, 283–315. [Google Scholar] [CrossRef] [PubMed]
- Benbrook, C.M. How did the US EPA and IARC reach diametrically opposed conclusions on the genotoxicity of glyphosate-based herbicides? Environ. Sci. Eur. 2019, 31, 2. [Google Scholar] [CrossRef]
- Tsai, W.-T. Trends in the Use of Glyphosate Herbicide and Its Relevant Regulations in Taiwan: A Water Contaminant of Increasing Concern. Toxics 2019, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Mesnage, R.; Benbrook, C.; Antoniou, M.N. Insight into the confusion over surfactant co-formulants in glyphosate-based herbicides. Food Chem. Toxicol. 2019, 128, 137–145. [Google Scholar] [CrossRef]
- EFSA—European Food Safety Authority. Conclusion on the peer review of the pesticide risk assessment of the active substance glyphosate: Peer review of the pesticide risk assessment of the active substance glyphosate: Peer review of the pesticide risk assessment of the active substance glyphosate. EFSA J. 2015, 13, 4302. [Google Scholar] [CrossRef]
- von Mérey, G.; Manson, P.S.; Mehrsheikh, A.; Sutton, P.; Levine, S.L. Glyphosate and aminomethylphosphonic acid chronic risk assessment for soil biota. Environ. Toxicol. Chem. 2016, 35, 2742–2752. [Google Scholar] [CrossRef]
- Mercurio, P.; Flores, F.; Mueller, J.F.; Carter, S.; Negri, A.P. Glyphosate persistence in seawater. Mar. Pollut. Bull. 2014, 85, 385–390. [Google Scholar] [CrossRef]
- Zhan, H.; Feng, Y.; Fan, X.; Chen, S. Recent advances in glyphosate biodegradation. Appl. Microbiol. Biotechnol. 2018, 102, 5033–5043. [Google Scholar] [CrossRef]
- Viirlaid, E.; Ilisson, M.; Kopanchuk, S.; Mäeorg, U.; Rinken, A.; Rinken, T. Immunoassay for rapid on-site detection of glyphosate herbicide. Environ. Monit. Assess. 2019, 191, 507. [Google Scholar] [CrossRef] [PubMed]
- Torretta, V.; Katsoyiannis, I.; Viotti, P.; Rada, E. Critical Review of the Effects of Glyphosate Exposure to the Environment and Humans through the Food Supply Chain. Sustainability 2018, 10, 950. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.; Boënne, W.; Desmet, N.; Seuntjens, P.; Bronders, J.; van Griensven, A. Quantification and characterization of glyphosate use and loss in a residential area. Sci. Total Environ. 2015, 517, 207–214. [Google Scholar] [CrossRef]
- Struger, J.; Van Stempvoort, D.R.; Brown, S.J. Sources of aminomethylphosphonic acid (AMPA) in urban and rural catchments in Ontario, Canada: Glyphosate or phosphonates in wastewater? Environ. Pollut. 2015, 204, 289–297. [Google Scholar] [CrossRef]
- Gunarathna, S.; Gunawardana, B.; Jayaweera, M.; Manatunge, J.; Zoysa, K. Glyphosate and AMPA of agricultural soil, surface water, groundwater and sediments in areas prevalent with chronic kidney disease of unknown etiology, Sri Lanka. J. Environ. Sci. Health Part B 2018, 53, 729–737. [Google Scholar] [CrossRef]
- Demonte, L.D.; Michlig, N.; Gaggiotti, M.; Adam, C.G.; Beldoménico, H.R.; Repetti, M.R. Determination of glyphosate, AMPA and glufosinate in dairy farm water from Argentina using a simplified UHPLC-MS/MS method. Sci. Total Environ. 2018, 645, 34–43. [Google Scholar] [CrossRef]
- Poiger, T.; Buerge, I.J.; Bächli, A.; Müller, M.D.; Balmer, M.E. Occurrence of the herbicide glyphosate and its metabolite AMPA in surface waters in Switzerland determined with on-line solid phase extraction LC-MS/MS. Environ. Sci. Pollut. Res. 2016, 24, 1588–1596. [Google Scholar] [CrossRef]
- Berman, M.C.; Marino, D.; Quiroga, M.V.; Zagarese, H. Occurrence and levels of glyphosate and AMPA in shallow lakes from the Pampean and Patagonian regions of Argentina. Chemosphere 2018, 200, 513–522. [Google Scholar] [CrossRef]
- Skeff, W.; Neumann, C.; Schulz-Bull, D.E. Glyphosate and AMPA in the estuaries of the Baltic Sea method optimization and field study. Mar. Pollut. Bull. 2015, 100, 577–585. [Google Scholar] [CrossRef]
- Primost, J.E.; Marino, D.J.G.; Aparicio, V.C.; Costa, J.L.; Carriquiriborde, P. Glyphosate and AMPA, “pseudo-persistent” pollutants under real-world agricultural management practices in the Mesopotamic Pampas agroecosystem, Argentina. Environ. Pollut. 2017, 229, 771–779. [Google Scholar] [CrossRef]
- Mac Loughlin, T.M.; Peluso, M.L.; Aparicio, V.C.; Marino, D.J. Contribution of soluble and particulate-matter fractions to the total glyphosate and AMPA load in water bodies associated with horticulture. Sci. Total Environ. 2020, 703, 134717. [Google Scholar] [CrossRef]
- Aparicio, V.C.; De Gerónimo, E.; Marino, D.; Primost, J.; Carriquiriborde, P.; Costa, J.L. Environmental fate of glyphosate and aminomethylphosphonic acid in surface waters and soil of agricultural basins. Chemosphere 2013, 93, 1866–1873. [Google Scholar] [CrossRef]
- Chang, F.-C.; Simcik, M.F.; Capel, P.D. Occurrence and fate of the herbicide glyphosate and its degradate aminomethylphosphonic acid in the atmosphere. Environ. Toxicol. Chem. 2010, 30, 548–555. [Google Scholar] [CrossRef]
- Morshed, M.; Omar, R.D.; Mohamad, S.; Abd, S.W. Determination of glyphosate through passive and active sampling methods in a treated field atmosphere. Afr. J. Agric. Res. 2011, 6, 4010–4018. [Google Scholar] [CrossRef]
- Cruz, J.M.; Murray, J.A. Determination of glyphosate and AMPA in oat products for the selection of candidate reference materials. Food Chem. 2020, 342, 128213. [Google Scholar] [CrossRef] [PubMed]
- Botero-Coy, A.M.; Ibañez, M.; Sancho, J.V.; Hernandez, F. Direct liquid chromatography–tandem mass spectrometry determination of underivatized glyphosate in rice, maize and soybean. J. Chromatogr. A 2013, 1313, 157–165. [Google Scholar] [CrossRef]
- Xu, J.; Smith, S.; Smith, G.; Wang, W.; Li, Y. Glyphosate contamination in grains and foods: An overview. Food Control 2019, 106, 106710. [Google Scholar] [CrossRef]
- Zoller, O.; Rhyn, P.; Rupp, H.; Zarn, J.; Geiser, C. Glyphosate residues in Swiss market foods: Monitoring and risk evaluation. Food Addit. Contam. Part B 2018, 11, 83–91. [Google Scholar] [CrossRef]
- Ehling, S.; Reddy, T.M. Analysis of Glyphosate and Aminomethylphosphonic Acid in Nutritional Ingredients and Milk by Derivatization with Fluorenylmethyloxycarbonyl Chloride and Liquid Chromatography–Mass Spectrometry. J. Agric. Food Chem. 2015, 63, 10562–10568. [Google Scholar] [CrossRef]
- Bøhn, T.; Cuhra, M.; Traavik, T.; Sanden, M.; Fagan, J.; Primicerio, R. Compositional differences in soybeans on the market: Glyphosate accumulates in Roundup Ready GM soybeans. Food Chem. 2014, 153, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; John, J. Glyphosate monitoring in water, foods, and urine reveals an association between urinary glyphosate and tea drinking: A pilot study. Int. J. Environ. Health Eng. 2018, 7, 2. [Google Scholar] [CrossRef]
- de Souza, A.P.F.; Rodrigues, N.R.; Reyes, F.G.R. Glyphosate and aminomethylphosphonic acid (AMPA) residues in Brazilian honey. Food Addit. Contam. Part B 2020, 14, 40–47. [Google Scholar] [CrossRef]
- Thompson, T.S.; Heever, J.P.V.D.; Limanowka, R.E. Determination of glyphosate, AMPA, and glufosinate in honey by online solid-phase extraction-liquid chromatography-tandem mass spectrometry. Food Addit. Contam. Part A 2019, 36, 434–446. [Google Scholar] [CrossRef]
- Zhao, J.; Pacenka, S.; Wu, J.; Richards, B.K.; Steenhuis, T.; Simpson, K.; Hay, A.G. Detection of glyphosate residues in companion animal feeds. Environ. Pollut. 2018, 243, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Connolly, A.; Basinas, I.; Jones, K.; Galea, K.S.; Kenny, L.; McGowan, P.; Coggins, A.M. Characterising glyphosate exposures among amenity horticulturists using multiple spot urine samples. Int. J. Hyg. Environ. Health 2018, 221, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Soukup, S.T.; Merz, B.; Bub, A.; Hoffmann, I.; Watzl, B.; Steinberg, P.; Kulling, S.E. Glyphosate and AMPA levels in human urine samples and their correlation with food consumption: Results of the cross-sectional KarMeN study in Germany. Arch. Toxicol. 2020, 94, 1575–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvez, S.; Gerona, R.R.; Proctor, C.; Friesen, M.; Ashby, J.L.; Reiter, J.L.; Lui, Z.; Winchester, P.D. Glyphosate exposure in pregnancy and shortened gestational length: A prospective Indiana birth cohort study. Environ. Health 2018, 17, 1–12. [Google Scholar] [CrossRef]
- Zoller, O.; Rhyn, P.; Zarn, J.A.; Dudler, V. Urine glyphosate level as a quantitative biomarker of oral exposure. Int. J. Hyg. Environ. Health 2020, 228, 113526. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, L.E.; Hansen, P.W.; Mizrak, S.; Hansen, H.K.; Mørck, T.A.; Nielsen, F.; Siersma, V.; Mathiesen, L. Biomonitoring of Danish school children and mothers including biomarkers of PBDE and glyphosate. Rev. Environ. Health 2017, 32, 279–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshioka, N.; Asano, M.; Kuse, A.; Mitsuhashi, T.; Nagasaki, Y.; Ueno, Y. Rapid determination of glyphosate, glufosinate, bialaphos, and their major metabolites in serum by liquid chromatography–tandem mass spectrometry using hydrophilic interaction chromatography. J. Chromatogr. A 2011, 1218, 3675–3680. [Google Scholar] [CrossRef]
- Valle, A.L.; Mello, F.C.C.; Alves-Balvedi, R.P.; Rodrigues, L.P.; Goulart, L.R. Glyphosate detection: Methods, needs and challenges. Environ. Chem. Lett. 2018, 17, 291–317. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.C.; He, M.M.; Shin, K.; Mai, V.; Jeong, K.C.; Finckh, M.R.; Morris, J.G., Jr. Environmental and health effects of the herbicide glyphosate. Sci. Total Environ. 2018, 616, 255–268. [Google Scholar] [CrossRef]
- Mesnage, R.; Defarge, N.; de Vendômois, J.S.; Séralini, G. Potential toxic effects of glyphosate and its commercial formulations below regulatory limits. Food Chem. Toxicol. 2015, 84, 133–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, J.P.; Antoniou, M.N.; Blumberg, B.; Carroll, L.; Colborn, T.; Everett, L.G.; Hansen, M.; Landrigan, P.J.; Lanphear, B.P.; Mesnage, R.; et al. Concerns over use of glyphosate-based herbicides and risks associated with exposures: A consensus statement. Environ. Health 2016, 15, 19. [Google Scholar] [CrossRef] [Green Version]
- Agostini, L.P.; Dettogni, R.S.; dos Reis, R.S.; Stur, E.; dos Santos, E.V.; Ventorim, D.P.; Garcia, F.M.; Cardoso, R.C.; Graceli, J.B.; Louro, I.D. Effects of glyphosate exposure on human health: Insights from epidemiological and in vitro studies. Sci. Total Environ. 2019, 705, 135808. [Google Scholar] [CrossRef]
- Gill, J.P.K.; Sethi, N.; Mohan, A.; Datta, S.; Girdhar, M. Glyphosate toxicity for animals. Environ. Chem. Lett. 2017, 16, 401–426. [Google Scholar] [CrossRef]
- Kašuba, V.; Milić, M.; Rozgaj, R.; Kopjar, N.; Mladinić, M.; Žunec, S.; Vrdoljak, A.L.; Pavicic, I.; Čermak, A.M.M.; Pizent, A.; et al. Effects of low doses of glyphosate on DNA damage, cell proliferation and oxidative stress in the HepG2 cell line. Environ. Sci. Pollut. Res. 2017, 24, 19267–19281. [Google Scholar] [CrossRef] [PubMed]
- Paz-Y.-Miño, C.; Sanchez, M.-E.; Arévalo, M.; Muñoz, M.J.; Witte, T.; De-La-Carrera, G.O.; Leone, P.E. Evaluation of DNA damage in an Ecuadorian population exposed to glyphosate. Genet. Mol. Biol. 2007, 30, 456–460. [Google Scholar] [CrossRef]
- Qiu, S.; Fu, H.; Zhou, R.; Yang, Z.; Bai, G.; Shi, B. Toxic effects of glyphosate on intestinal morphology, antioxidant capacity and barrier function in weaned piglets. Ecotoxicol. Environ. Saf. 2020, 187, 109846. [Google Scholar] [CrossRef]
- Avigliano, L.; Fassiano, A.V.; Medesani, D.A.; De Molina, M.C.R.; Rodríguez, E.M. Effects of Glyphosate on Growth Rate, Metabolic Rate and Energy Reserves of Early Juvenile Crayfish, Cherax quadricarinatus M. Bull. Environ. Contam. Toxicol. 2014, 92, 631–635. [Google Scholar] [CrossRef]
- Rodrigues, L.D.B.; Costa, G.G.; Thá, E.L.; da Silva, L.R.; de Oliveira, R.; Leme, D.M.; Cestari, M.M.; Grisolia, C.K.; Valadares, M.C.; de Oliveira, G.A.R. Impact of the glyphosate-based commercial herbicide, its components and its metabolite AMPA on non-target aquatic organisms. Mutat. Res. Toxicol. Environ. Mutagen. 2019, 842, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Thongprakaisang, S.; Thiantanawat, A.; Rangkadilok, N.; Suriyo, T.; Satayavivad, J. Glyphosate induces human breast cancer cells growth via estrogen receptors. Food Chem. Toxicol. 2013, 59, 129–136. [Google Scholar] [CrossRef]
- Elie-Caille, C.; Heu, C.; Guyon, C.; Nicod, L.; Nicod, L.P. Morphological damages of a glyphosate-treated human keratinocyte cell line revealed by a micro- to nanoscale microscopic investigation. Cell Biol. Toxicol. 2009, 26, 331–339. [Google Scholar] [CrossRef]
- Koller, V.J.; Fürhacker, M.; Nersesyan, A.; Mišík, M.; Eisenbauer, M.; Knasmueller, S. Cytotoxic and DNA-damaging properties of glyphosate and Roundup in human-derived buccal epithelial cells. Arch. Toxicol. 2012, 86, 805–813. [Google Scholar] [CrossRef]
- Dai, P.; Hu, P.; Tang, J.; Li, Y.; Li, C. Effect of glyphosate on reproductive organs in male rat. Acta Histochem. 2016, 118, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Martinez, A.; Al-Ahmad, A.J. Effects of glyphosate and aminomethylphosphonic acid on an isogeneic model of the human blood-brain barrier. Toxicol. Lett. 2018, 304, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Cuhra, M.; Traavik, T.; Bøhn, T. Clone- and age-dependent toxicity of a glyphosate commercial formulation and its active ingredient in Daphnia magna. Ecotoxicology 2012, 22, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Webster, T.U.; Laing, L.V.; Florance, H.; Santos, E. Effects of Glyphosate and its Formulation, Roundup, on Reproduction in Zebrafish (Danio rerio). Environ. Sci. Technol. 2013, 48, 1271–1279. [Google Scholar] [CrossRef]
- Roy, N.M.; Ochs, J.; Zambrzycka, E.; Anderson, A. Glyphosate induces cardiovascular toxicity in Danio rerio. Environ. Toxicol. Pharmacol. 2016, 46, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Armiliato, N.; Ammar, D.; Nezzi, L.; Straliotto, M.; Müller, Y.M.R.; Nazari, E.M. Changes in Ultrastructure and Expression of Steroidogenic Factor-1 in Ovaries of Zebrafish Danio rerio Exposed to Glyphosate. J. Toxicol. Environ. Health Part A 2014, 77, 405–414. [Google Scholar] [CrossRef]
- Bridi, D.; Altenhofen, S.; Gonzalez, J.B.; Reolon, G.K.; Bonan, C.D. Glyphosate and Roundup® alter morphology and behavior in zebrafish. Toxicology 2017, 392, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Plata, I.; Giordano, M.; Díaz-Muñoz, M.; Rodríguez, V.M. The herbicide glyphosate causes behavioral changes and alterations in dopaminergic markers in male Sprague-Dawley rat. NeuroToxicology 2015, 46, 79–91. [Google Scholar] [CrossRef]
- Shiogiri, N.S.; Paulino, M.G.; Carraschi, S.P.; Baraldi, F.G.; da Cruz, C.; Fernandes, M.N. Acute exposure of a glyphosate-based herbicide affects the gills and liver of the Neotropical fish, Piaractus mesopotamicus. Environ. Toxicol. Pharmacol. 2012, 34, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Burchfield, S.L.; Bailey, D.C.; Todt, C.E.; Denney, R.D.; Negga, R.; Fitsanakis, V.A. Acute exposure to a glyphosate-containing herbicide formulation inhibits Complex II and increases hydrogen peroxide in the model organism Caenorhabditis elegans. Environ. Toxicol. Pharmacol. 2019, 66, 36–42. [Google Scholar] [CrossRef]
- Hao, Y.; Zhang, Y.; Cheng, J.; Xu, W.; Xu, Z.; Gao, J.; Tao, L. Adjuvant contributes Roundup’s unexpected effects on A549 cells. Environ. Res. 2020, 184, 109306. [Google Scholar] [CrossRef]
- Bednářová, A.; Kropf, M.; Krishnan, N. The surfactant polyethoxylated tallowamine (POEA) reduces lifespan and inhibits fecundity in Drosophila melanogaster—In vivo and in vitro study. Ecotoxicol. Environ. Saf. 2019, 188, 109883. [Google Scholar] [CrossRef] [PubMed]
- Puértolas, L.; Damásio, J.; Barata, C.; Soares, A.M.; Prat, N. Evaluation of side-effects of glyphosate mediated control of giant reed (Arundo donax) on the structure and function of a nearby Mediterranean river ecosystem. Environ. Res. 2010, 110, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Rocha, T.L.; dos Santos, A.P.R.; Yamada, P.T.; Soares, C.M.D.A.; Borges, C.L.; Bailão, A.M.; Sabóia-Morais, S.M.T. Proteomic and histopathological response in the gills of Poecilia reticulata exposed to glyphosate-based herbicide. Environ. Toxicol. Pharmacol. 2015, 40, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Dedeke, G.A.; Owagboriaye, F.O.; Ademolu, K.O.; Olujimi, O.O.; Aladesida, A. Comparative Assessment on Mechanism Underlying Renal Toxicity of Commercial Formulation of Roundup Herbicide and Glyphosate Alone in Male Albino Rat. Int. J. Toxicol. 2018, 37, 285–295. [Google Scholar] [CrossRef]
- Bali, Y.A.; Ba-Mhamed, S.; Bennis, M. Behavioral and Immunohistochemical Study of the Effects of Subchronic and Chronic Exposure to Glyphosate in Mice. Front. Behav. Neurosci. 2017, 11, 146. [Google Scholar] [CrossRef]
- Owagboriaye, F.O.; Dedeke, G.A.; Ademolu, K.O.; Olujimi, O.O.; Ashidi, J.S.; Adeyinka, A.A. Reproductive toxicity of Roundup herbicide exposure in male albino rat. Exp. Toxicol. Pathol. 2017, 69, 461–468. [Google Scholar] [CrossRef]
- Guilherme, S.; Gaivão, I.; Santos, M.; Pacheco, M. DNA damage in fish (Anguilla anguilla) exposed to a glyphosate-based herbicide—Elucidation of organ-specificity and the role of oxidative stress. Mutat. Res. Toxicol. Environ. Mutagen. 2012, 743, 1–9. [Google Scholar] [CrossRef]
- Vanlaeys, A.; Dubuisson, F.; Seralini, G.-E.; Travert, C. Formulants of glyphosate-based herbicides have more deleterious impact than glyphosate on TM4 Sertoli cells. Toxicol. In Vitro 2018, 52, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Braz-Mota, S.; Sadauskas-Henrique, H.; Duarte, R.; Val, A.; Almeida-Val, V.M. Roundup® exposure promotes gills and liver impairments, DNA damage and inhibition of brain cholinergic activity in the Amazon teleost fish Colossoma macropomum. Chemosphere 2015, 135, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Ørsted, M.; Roslev, P. A fluorescence-based hydrolytic enzyme activity assay for quantifying toxic effects of Roundup® toDaphnia magna. Environ. Toxicol. Chem. 2015, 34, 1841–1850. [Google Scholar] [CrossRef] [PubMed]
- Levine, S.L.; von Mérey, G.; Minderhout, T.; Manson, P.; Sutton, P. Aminomethylphosphonic acid has low chronic toxicity to Daphnia magna and Pimephales promelas. Environ. Toxicol. Chem. 2015, 34, 1382–1389. [Google Scholar] [CrossRef]
- Cheron, M.; Brischoux, F. Aminomethylphosphonic acid alters amphibian embryonic development at environmental concentrations. Environ. Res. 2020, 190, 109944. [Google Scholar] [CrossRef]
- Kwiatkowska, M.; Huras, B.; Bukowska, B. The effect of metabolites and impurities of glyphosate on human erythrocytes (in vitro). Pestic. Biochem. Physiol. 2014, 109, 34–43. [Google Scholar] [CrossRef]
- Asnicar, D.; Cappelli, C.; Sallehuddin, A.S.; Maznan, N.A.; Marin, M.G. Effects of Glyphosate-Based and Derived Products on Sea Urchin Larval Development. J. Mar. Sci. Eng. 2020, 8, 661. [Google Scholar] [CrossRef]
- Tush, D.; Loftin, K.A.; Meyer, M. Characterization of polyoxyethylene tallow amine surfactants in technical mixtures and glyphosate formulations using ultra-high performance liquid chromatography and triple quadrupole mass spectrometry. J. Chromatogr. A 2013, 1319, 80–87. [Google Scholar] [CrossRef]
- Chłopecka, M.; Mendel, M.; Dziekan, N.; Karlik, W. The effect of glyphosate-based herbicide Roundup and its co-formulant, POEA, on the motoric activity of rat intestine—In vitro study. Environ. Toxicol. Pharmacol. 2017, 49, 156–162. [Google Scholar] [CrossRef]
- Séguin, A.; Mottier, A.; Perron, C.; Lebel, J.M.; Serpentini, A.; Costil, K. Sub-lethal effects of a glyphosate-based commercial formulation and adjuvants on juvenile oysters (Crassostrea gigas) exposed for 35 days. Mar. Pollut. Bull. 2017, 117, 348–358. [Google Scholar] [CrossRef]
- Seok, S.-J.; Park, J.-S.; Hong, J.-R.; Gil, H.-W.; Yang, J.-O.; Lee, E.-Y.; Song, H.-Y.; Hong, S.-Y. Surfactant volume is an essential element in human toxicity in acute glyphosate herbicide intoxication. Clin. Toxicol. 2011, 49, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Defarge, N.; de Vendômois, J.S.; Séralini, G. Toxicity of formulants and heavy metals in glyphosate-based herbicides and other pesticides. Toxicol. Rep. 2017, 5, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Seralini, G.-E.; Jungers, G. Toxic compounds in herbicides without glyphosate. Food Chem. Toxicol. 2020, 146, 111770. [Google Scholar] [CrossRef] [PubMed]

| Compound | Sample | Concentration | Detection Method | Ref. |
|---|---|---|---|---|
| Glyphosate | Rainwater | 6.1 µg/L | LC–MSn | [46] |
| Stream water | 41 ng/L | IC/MSn | [47] | |
| Groundwater | 4 µg/L | LC–MS | [48] | |
| Groundwater | 21.2 µg/L | UHPLC–MSn | [49] | |
| Groundwater | 0.025 µg/L | LC–MSn | [50] | |
| Lake water | 4.52 µg/L | HPLC–MSn | [51] | |
| Lake water | 45 µg/L | LC–MS | [48] | |
| Marine water | 1.7 µg/L | LC–MSn | [52] | |
| Suspended particulate matter | 584 µg/kg | UHPLC–MSn | [53] | |
| Water | 17 µg/L | UHPLC–MSn | [54] | |
| Suspended particulate matter | 0.13 µg/L | HPLC–MSn | [51] | |
| Sediment | 20.34 µg/kg | HPLC–MSn | [51] | |
| Sediment | 3294 µg/kg | UHPLC–MSn | [53] | |
| Sediment | 1000 µg/kg | LC–MS | [48] | |
| Soil | 8105 µg/kg | UHPLC–MSn | [53] | |
| Soil | 1502 µg/kg | UHPLC–MSn | [55] | |
| Soil | 690 µg/kg | LC–MS | [48] | |
| Air | 0.48 ng/m3 | HPLC-MS | [56] | |
| Air | 0.24 ng/m3 | HPLC-MS | [56] | |
| Air (application) | 42.96 µg/m3 | HPLC-FD | [57] | |
| Air (0–4 h after application) | 0.1 µg/m3 | HPLC-FD | [57] | |
| Air (4–8 h after application) | 0.05 µg/m3 | HPLC-FD | [57] | |
| Organic oat flour | 11 µg/kg | LC–MSn | [58] | |
| Oatmeal | 1100 µg/kg | LC–MSn | [58] | |
| Oat-based cereals | 901 µg/kg | LC–MSn | [58] | |
| Oat flour | 554 µg/kg | LC–MSn | [58] | |
| Wheat | 670 µg/kg | LC–MSn | [59] | |
| Durum wheat | 421 µg/kg | LC–MSn | [60] | |
| Breakfast cereal | 291 µg/kg | LC–MSn | [61] | |
| Soy protein isolate | 105 µg/kg | UHPLC–MSn | [62] | |
| Soy protein concentrate | 850 µg/kg | UHPLC–MSn | [62] | |
| Soybean | 8800 µg/kg | HPLC-FD | [63] | |
| Corn | 1.6 µg/kg | ELISA kit | [64] | |
| Coffee | 26.32 µg/kg | ELISA kit | [64] | |
| Pea | 60 µg/kg | LC–MSn | [59] | |
| Wine | 18.9 µg/kg | LC–MSn | [61] | |
| Beer | 2.8 µg/kg | ELISA kit | [64] | |
| Tea leaves | 40.43 µg/kg | ELISA kit | [64] | |
| Tea bag | 728.2 µg/kg | ELISA kit | [64] | |
| Bread | 45.8 µg/kg | LC–MSn | [61] | |
| Honey | 220 µg/kg | HPLC-FD | [65] | |
| Honey | 49.8 µg/kg | LC–MSn | [66] | |
| Cat and dog food | 0.03 µg/kg | ELISA kit | [67] | |
| Human urine | 7.4 µg/L | LC–MSn | [68] | |
| Human urine | 1.36 µg/L | LC–MSn | [69] | |
| Human urine | 7.2 µg/L | LC–MSn | [70] | |
| Human urine | 5.6 µg/L | LC–MSn | [71] | |
| Human urine | 3.3 ng/L | ELISA kit | [72] | |
| Human serum | 1477 µg/mL | LC–MSn | [73] | |
| Human serum | 89 µg/mL | LC–MSn | [73] | |
| AMPA | Rainwater | 5.8 µg/L | LC–MS | [46] |
| Groundwater | 6.5 µg/L | UHPLC–MSn | [49] | |
| Groundwater | 0.65 µg/L | LC–MSn | [50] | |
| Groundwater | 11 µg/L | LC–MS | [48] | |
| Lake water | 0.90 µg/L | HPLC–MSn | [51] | |
| Marine water | 4.2 µg/L | LC–MSn | [52] | |
| Water | 4.5 µg/L | UHPLC–MSn | [54] | |
| Suspended particulate matter | 475 µg/kg | UHPLC–MSn | [53] | |
| Suspended particulate matter | 0.07 µg/L | HPLC–MSn | [51] | |
| Sediment | 7219 µg/kg | UHPLC–MSn | [53] | |
| Sediment | 15 µg/kg | LC–MS | [48] | |
| Sediment | 32.89 µg/kg | HPLC–MSn | [51] | |
| Soil | 38,939 µg/kg | UHPLC–MSn | [53] | |
| Soil | 2256 µg/kg | UHPLC–MSn | [55] | |
| Soil | 8 µg/kg | LC–MS | [48] | |
| Air | 0.06 ng/m3 | HPLC-MS | [56] | |
| Air | 0.02 ng/m3 | HPLC-MS | [56] | |
| Oatmeal | 40 µg/kg | LC–MSn | [58] | |
| Oat-based cereals | 25 µg/kg | LC–MSn | [58] | |
| Oat flour | 25 µg/kg | LC–MSn | [58] | |
| Breakfast cereal | 10 µg/kg | LC–MSn | [61] | |
| Durum wheat | 247 µg/kg | LC–MSn | [60] | |
| Soy protein isolate | 210 µg/kg | UHPLC–MSn | [62] | |
| Soy protein concentrate | 2710 µg/kg | UHPLC–MSn | [62] | |
| Soybean | 10,000 µg/kg | HPLC-FD | [63] | |
| Wine | 3.4 µg/kg | LC–MSn | [61] | |
| Honey | 100 µg/kg | HPLC-FD | [65] | |
| Honey | 50.1 µg/kg | LC–MSn | [66] | |
| Human urine | 1.53 µg/L | LC–MSn | [69] | |
| Human serum | 1.5 µg/mL | LC–MSn | [73] | |
| Human serum | 0.07 µg/mL | LC–MSn | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins-Gomes, C.; Silva, T.L.; Andreani, T.; Silva, A.M. Glyphosate vs. Glyphosate-Based Herbicides Exposure: A Review on Their Toxicity. J. Xenobiot. 2022, 12, 21-40. https://doi.org/10.3390/jox12010003
Martins-Gomes C, Silva TL, Andreani T, Silva AM. Glyphosate vs. Glyphosate-Based Herbicides Exposure: A Review on Their Toxicity. Journal of Xenobiotics. 2022; 12(1):21-40. https://doi.org/10.3390/jox12010003
Chicago/Turabian StyleMartins-Gomes, Carlos, Tânia L. Silva, Tatiana Andreani, and Amélia M. Silva. 2022. "Glyphosate vs. Glyphosate-Based Herbicides Exposure: A Review on Their Toxicity" Journal of Xenobiotics 12, no. 1: 21-40. https://doi.org/10.3390/jox12010003