
Increasing Potato Sustainability to PVY under Water Deficiency by Bacillus Bacteria with Salicylic Acid and Methyl Jasmonate
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
Materials
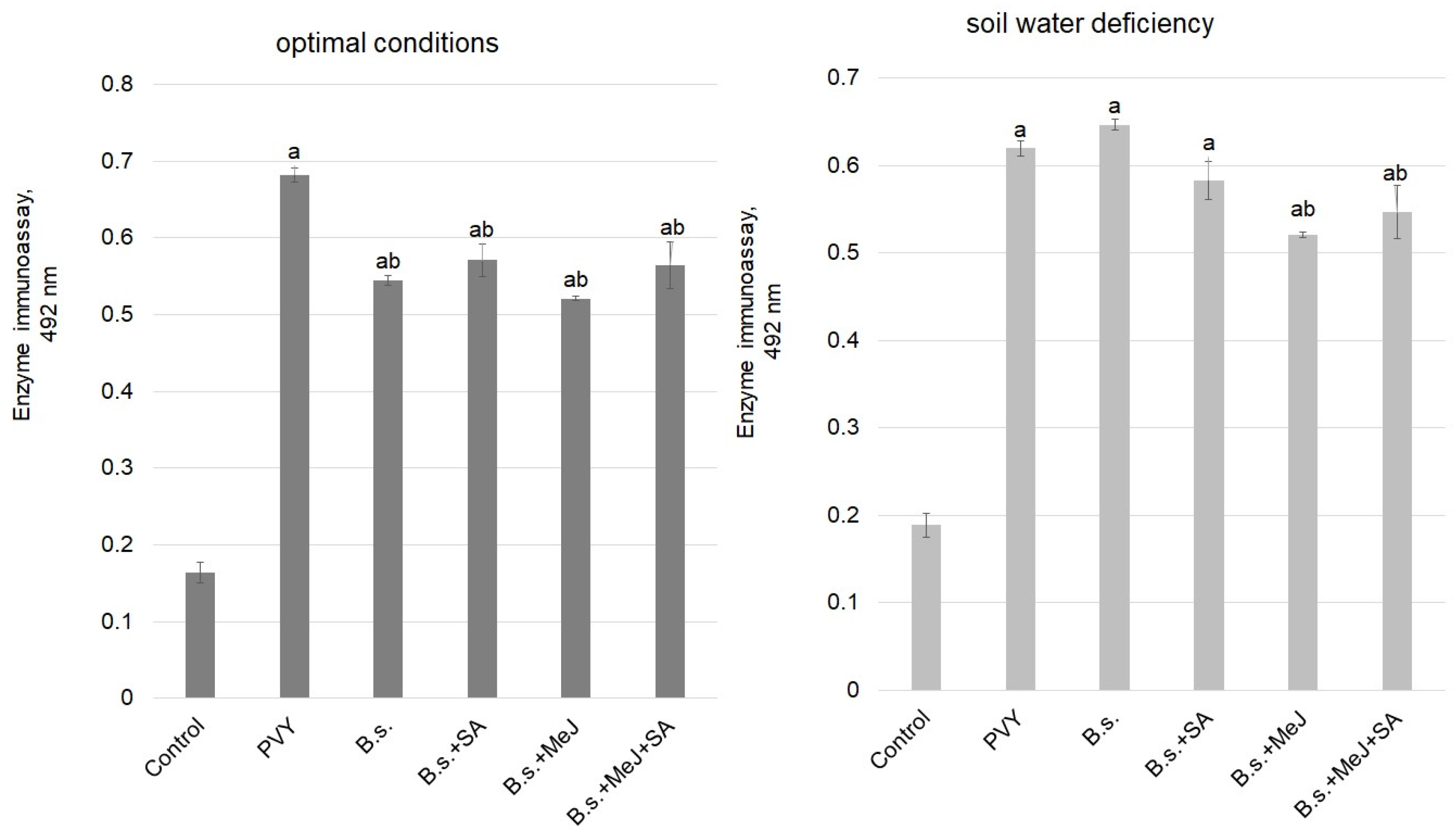
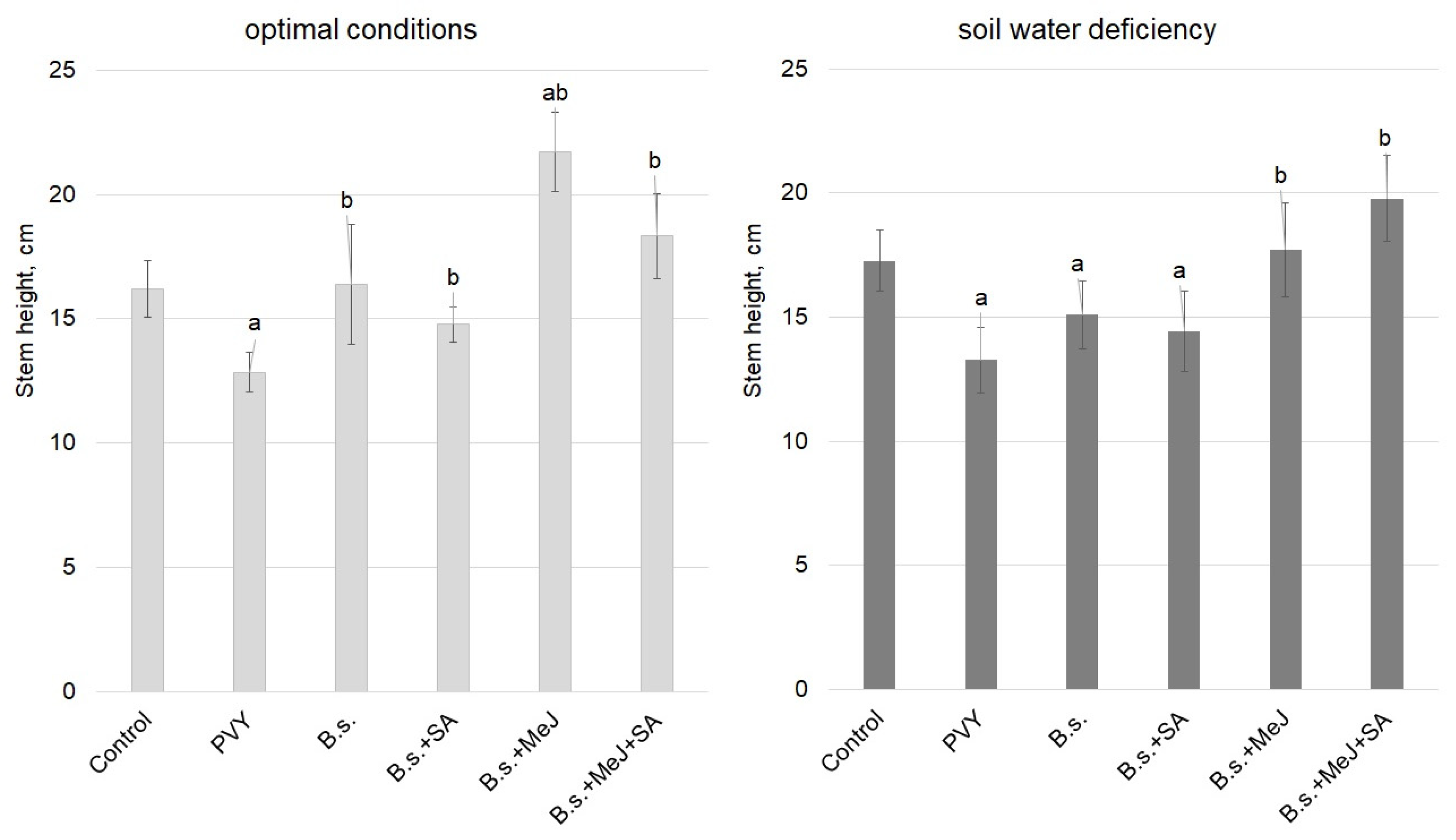
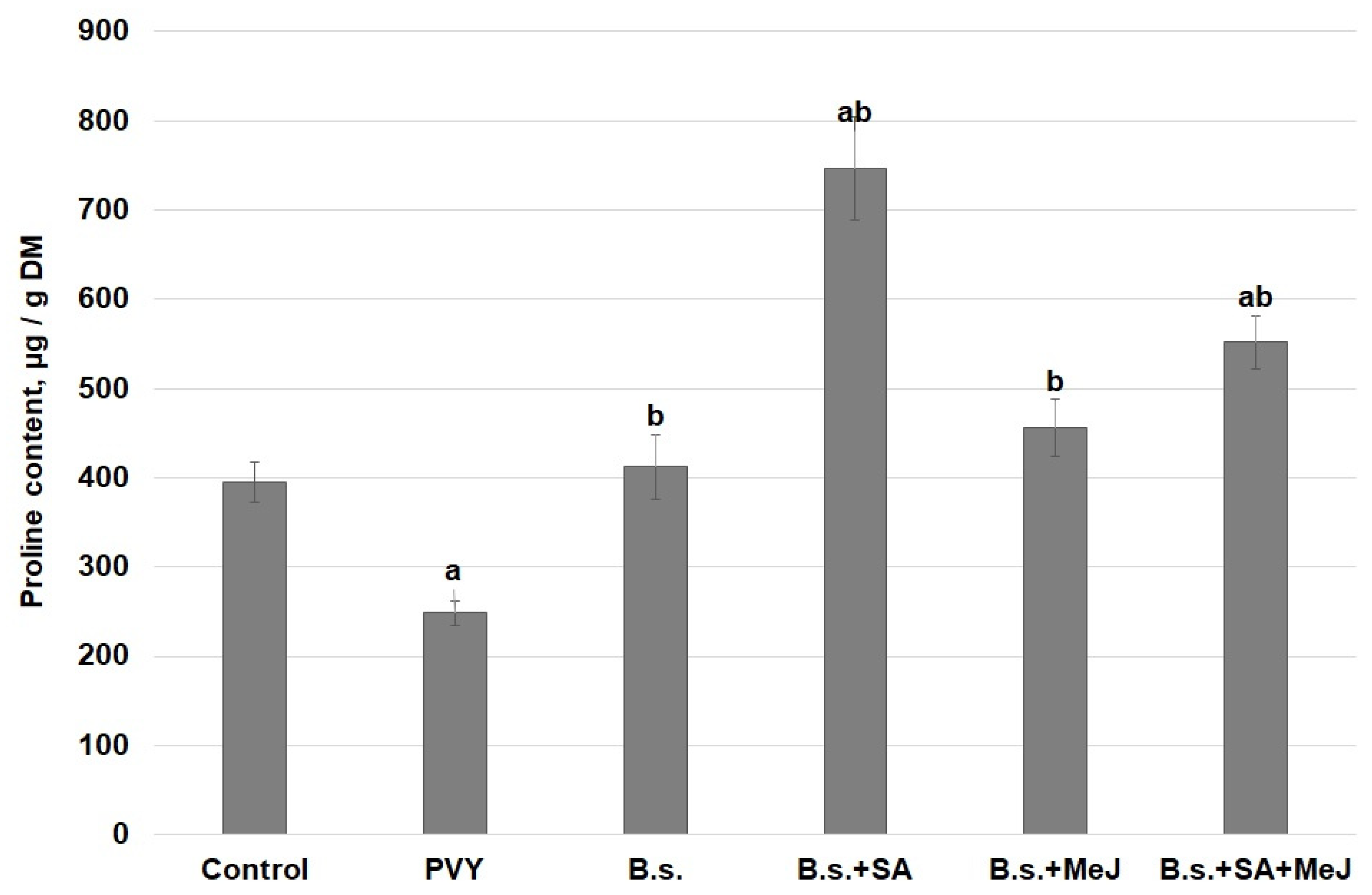
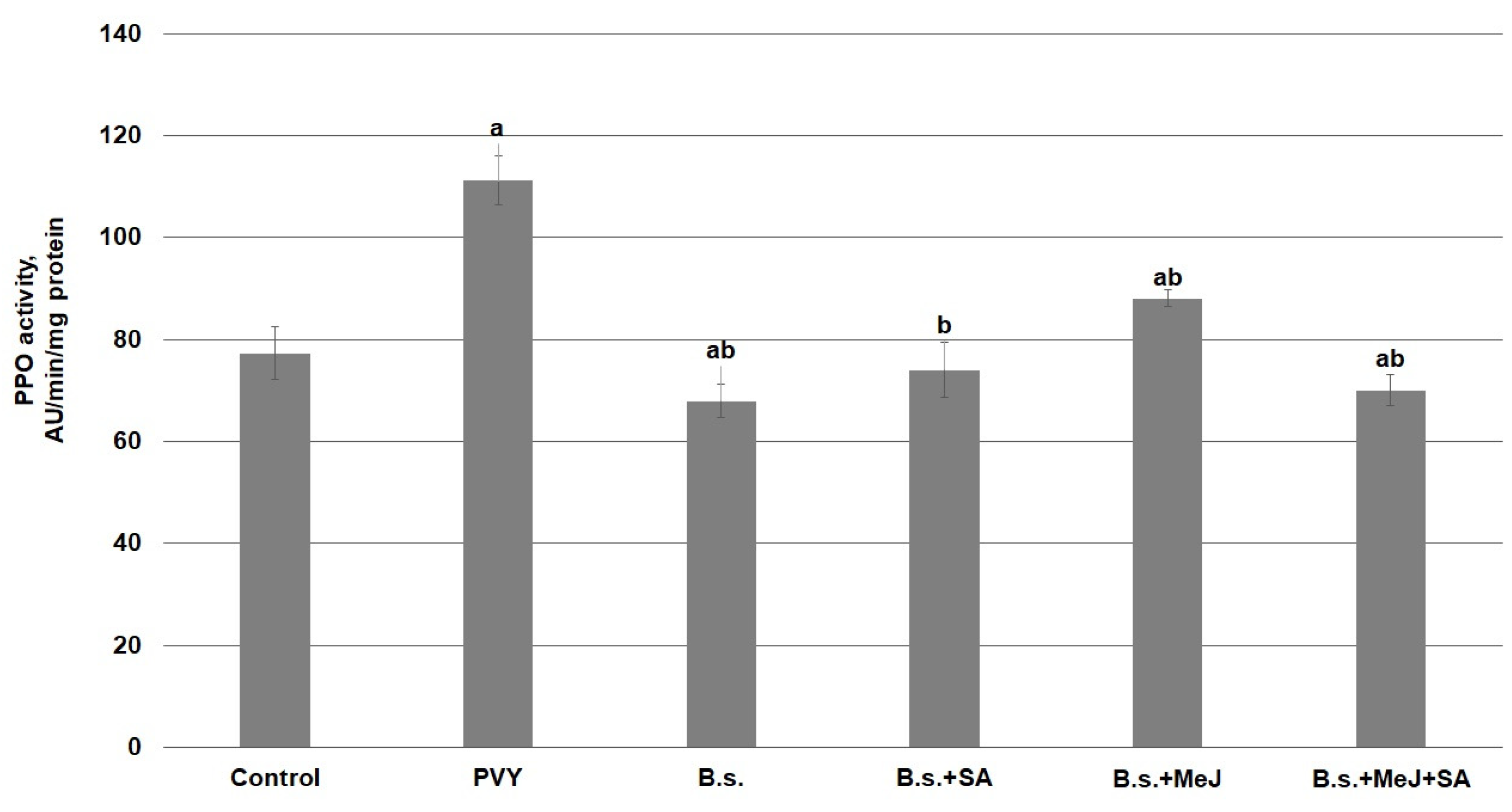
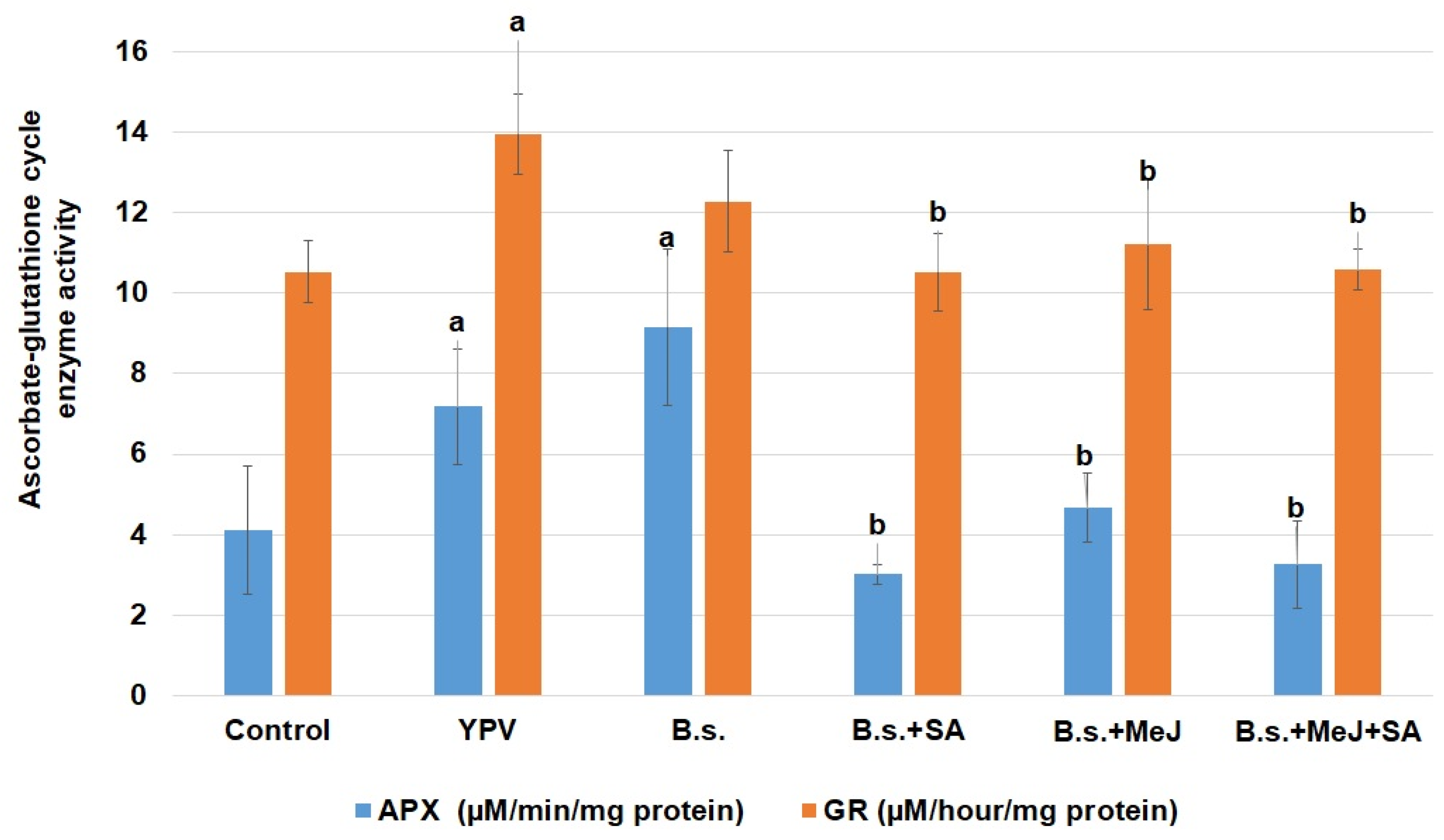
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO (Food and Agriculture Organization of the United Nations). FAOSTAT—Food and Agriculture Organization of the United Nations Statistics Division. FAOSTAT, Statistical Database. 2021. Available online: http://faostat.fao.org/ (accessed on 1 December 2022).
- Devaux, A.; Kromann, P.; Ortiz, O. Potatoes for sustainable global food security. Potato Res. 2014, 57, 185–199. [Google Scholar] [CrossRef]
- Valkonen, J.P.T.; Gebhardt, C.H.; Zimnoch-Guzowska, E.; Watanabe, K.N. Resistance to Potato Virus Y in potato. In Potato Virus Y: Biodiversity, Pathogenicity, Epidemiology and Management; Lacomme, C., Glais, L., Bellstedt, D., Dupuis, B., Karasev, A., Jacquot, E., Eds.; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Termorshuizen, A.J. Chapter 29—Fungal and Fungus-Like Pathogens of Potato. In Potato Biology and Biotechnology; Vreugdenhil, D., Bradshaw, J., Gebhardt, C., Govers, F., Mackerron, D.K.L., Taylor, M.A., Ross, H.A., Eds.; Elsevier Science B.V.: Amsterdam, The Netherlands, 2007; pp. 643–665. [Google Scholar]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of Combined Abiotic and Biotic Stresses on Plant Growth and Avenues for Crop Improvement by Exploiting Physio-morphological Traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef] [Green Version]
- Hamooh, B.T.; Sattar, F.A.; Wellman, G.; Mousa, M.A.A. Metabolomic and Biochemical Analysis of Two Potato (Solanum tuberosum L.) Cultivars Exposed to In Vitro Osmotic and Salt Stresses. Plants 2021, 10, 98. [Google Scholar] [CrossRef]
- Burton, W.G. Challenges for stress physiology in potato. Am. Potato J. 1981, 58, 3–14. [Google Scholar] [CrossRef]
- Jones, R.A.C. Global Plant Virus Disease Pandemics and Epidemics. Plants 2021, 10, 233. [Google Scholar] [CrossRef] [PubMed]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef]
- Saeed, Q.; Xiukang, W.; Haider, F.U.; Kučerik, J.; Mumtaz, M.Z.; Holatko, J.; Naseem, M.; Kintl, A.; Ejaz, M.; Naveed, M.; et al. Rhizosphere Bacteria in Plant Growth Promotion, Biocontrol, and Bioremediation of Contaminated Sites: A Comprehensive Review of Effects and Mechanisms. Int. J. Mol. Sci. 2021, 22, 10529. [Google Scholar] [CrossRef] [PubMed]
- Burkhanova, G.F.; Veselova, S.V.; Sorokan, A.V.; Blagova, D.K.; Nuzhnaya, T.V.; Maksimov, I.V. Strains of Bacillus ssp. regulate wheat resistance to Septoria nodorum Berk. Appl. Biochem. Microbiol. 2017, 53, 346–352. [Google Scholar] [CrossRef]
- Abdel-Rahman, S.S.A.; Abdel-Kader, A.A.S.; Khalil, S.E. Response of Three Sweet Basil Cultivars to Inoculation with Bacillus subtilis and Arbuscular Mycorrhizal Fungi under Salt Stress Conditions. Nat. Sci. 2011, 9, 93–111. [Google Scholar]
- Veselova, S.V.; Burkhanova, G.F.; Rumyantsev, S.D.; Blagova, D.K.; Maksimov, I.V. Strains of Bacillus ssp. Regulate Wheat Resistance to Greenbug Schizaphis graminum Rond. Appl. Biochem. Microbiol. 2019, 55, 41–47. [Google Scholar] [CrossRef]
- Riera, N.; Wang, H.; Li, Y.; Li, J.; Pelz-Stelinski, K.; Wang, N. Induced Systemic Resistance Against Citrus Canker Disease by Rhizobacteria. Phytopathology 2018, 108, 1038–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, S.; Jiang, H.; Ding, T.; Xu, Q.; Chai, W.; Cheng, B. Bacillus amyloliquefaciens FZB42 represses plant miR846 to induce systemic resistance via a jasmonic acid-dependent signal-ling pathway. Mol. Plant Pathol. 2018, 19, 1612–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalhais, L.C.; Dennis, P.G.; Badri, D.V.; Tyson, G.W.; Vivanco, J.M.; Schenk, P.M. Activation of the jasmonic acid plant defence pathway alters the composition of rhizosphere bacterial communities. PLoS ONE 2013, 8, e56457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichmann, R.; Richards, L.; Schäfer, P. Hormones as go-betweens in plant microbiome assembly. Plant J. 2021, 105, 518–541. [Google Scholar] [CrossRef] [PubMed]
- Sorokan, A.; Burkhanova, G.; Gordeev, A.; Maksimov, I. Exploring the Role of Salicylic Acid in Regulating the Colonization Ability of Bacillus subtilis 26D in Potato Plants and Defense against Phytophthora infestans. Int. J. Plant Biol. 2023, 14, 242–253. [Google Scholar] [CrossRef]
- Barkai-Golan, R. Postharvest diseases of fruit and vegetables. In Development and Control, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2001; p. 442. [Google Scholar]
- Maksimov, I.V.; Abizgildina, R.R.; Pusenkova, L.I. Plant growth promoting rhizobacteria as alternative to chemical crop protectors from pathogens (Review). Appl. Biochem. Microbiol. 2011, 47, 333–345. [Google Scholar] [CrossRef]
- Lastochkina, O.; Baymiev, A.; Shayahmetova, A.; Garshina, D.; Koryakov, I.; Shpirnaya, I.; Pusenkova, L.; Mardanshin, I.; Kasnak, C.; Palamutoglu, R. Effects of Endophytic Bacillus subtilis and Salicylic Acid on Postharvest Diseases (Phytophthora infestans, Fusarium oxysporum) Development in Stored Potato Tubers. Plants 2020, 9, 76. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Guo, Z.; Li, H.; Wang, M. Phytochrome A and B function antagonistically to regulate cold tolerance via abscisic acid-dependent jasmonate signaling. Plant Physiol. 2016, 170, 459–471. [Google Scholar] [CrossRef] [Green Version]
- Fageria, M.; Nie, X.; Gallagher, A.; Singh, M. Mechanical Transmission of Potato Virus Y (PVY) through Seed Cutting and Plant Wounding. Am. J. Potato Res. 2015, 92, 143–147. [Google Scholar] [CrossRef]
- Wassu, M. Specific Gravity, Dry Matter Content, and Starch Content of Potato (Solanum tuberosum L.) Varieties Cultivated in Eastern Ethiopia. East Afr. J. Sci. 2016, 10, 87–102. [Google Scholar]
- Lichtenthaler, K.; Welburn, R. Determination of Total Carotenoids and Chlorophylls A and B of Leaf Extracts in Different Solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldern, R.P.; Teare, D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Kumar, V.B.A.; Mohan, T.C.K.; Murugan, K. Purification and kinetic characterization of polyphenol oxidase from Barbados cherry (Malpighia glabra L.). Food Chem. 2008, 110, 328–333. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Aono, M.; Kubo, A.; Saji, H. Resistance to active oxygen toxicity of transgenic Nicotiana tabacum that express the gene for glutathione reductase from Escherichia coli. Plant Cell Physiol. 1991, 32, 691–697. [Google Scholar] [CrossRef]
- Rahman Khan, M.M.; Rahman, M.M.; Islam, M.S.; Begum, S.A. A Simple UV-spectrophotometric Method for the Determination of Vitamin C Content in Various Fruits and Vegetables at Sylhet Area in Bangladesh. J. Biol. Sci. 2006, 6, 388–392. [Google Scholar] [CrossRef] [Green Version]
- Fedina, E.O.; Karimova, F.G.; Tarchevsky, I.A.; Toropygin, I.Y.; Khripach, V.A. Effect of epibrassinolide on tyrosine phosphorylation of the Calvin cycle enzymes. Rus. J. Plant Phys. 2008, 55, 193–200. [Google Scholar] [CrossRef]
- Yanchevskaya, T.G.; Grits, A.N.; Kolomiets, E.I.; Romanovskaya, T.V.; Yarullina, L.G.; Ibragimov, R.I.; Tsvetkov, V.O. Stimulation of Cellular Mechanisms of Potato Antivirus Resistance by the Action of a Preparation Based on Bacillus subtilis Bacteria. Appl. Biochem. Microbiol. 2018, 54, 324–330. [Google Scholar] [CrossRef]
- Li, T.; Huang, Y.; Xu, Z.S.; Wang, F.; Xiong, A.S. Salicylic acid-induced differential resistance to the Tomato yellow leaf curl virus among resistant and susceptible tomato cultivars. BMC Plant Biol. 2019, 19, 173. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, N.; Basak, J. Exogenous application of methyl jasmonate induces defense response and develops tolerance against mungbean yellow mosaic India virus in Vigna mungo. Funct. Plant Biol. 2018, 46, 69–81. [Google Scholar] [CrossRef]
- Yarullina, L.G.; Tsvetkov, V.O.; Khabibullina, V.O.; Cherepanova, E.A.; Burkhanova, G.F.; Zaikina, E.A.; Kalatskaya, J.N. Impact of Bacillus subtilis Bacteria in Combination with Salicylic and Jasmonic Acids on Changing the Proteome of Potato Leaves when Infected by Phytophthora infestans (Mont.) De Bary and with a Moisture Deficit. Russ. J. Plant Physiol. 2022, 69, 81. [Google Scholar] [CrossRef]
- Abdelgawad, Z.A.; Khalafaallah, A.A.; Abdallah, M.M. Impact of methyl jasmonate on antioxidant activity and some biochemical aspects of maize plant grown under water stress condition. Agric. Sci. 2014, 5, 1077–1088. [Google Scholar] [CrossRef] [Green Version]
- Veselova, S.V.; Sorokan, A.V.; Burkhanova, G.F.; Rumyantsev, S.D.; Cherepanova, E.A.; Alekseev, V.Y.; Sarvarova, E.R.; Kasimova, A.R.; Maksimov, I.V. By Modulating the Hormonal Balance and Ribonuclease Activity of Tomato Plants Bacillus subtilis Induces Defense Response against Potato Virus X and Potato Virus, Y. Biomolecules 2022, 12, 288. [Google Scholar] [CrossRef] [PubMed]
- Blake, C.; Nordgaard, C.M.; Kovacs, A.T. Molecular Aspects of Plant Growth Promotion and Protection by Bacillus subtilis. Mol. Plant-Microbe Interact. 2021, 34, 15–25. [Google Scholar] [CrossRef]
- Tayyab, N.; Naz, R.; Yasmin, H.; Nosheen, A.; Keyani, R.; Sajjad, M.; Hassan, M.H.; Roberts, T.H. Combined seed and foliar pre-treatments with exogenous methyl jasmonate and salicylic acid mitigate drought-induced stress in maize. PLoS ONE 2020, 15, e0232269. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Al-Askar, A.A.; Arishi, A.A.; Behiry, S.I. Trichoderma hamatum Strain Th23 Promotes Tomato Growth and Induces Systemic Resistance against Tobacco Mosaic Virus. J. Fungi 2022, 8, 228. [Google Scholar] [CrossRef]
- Sofy, A.R.; Dawoud, R.A.; Sofy, M.R.; Mohamed, H.I.; Hmed, A.A.; El-Dougdoug, N.K. Improving Regulation of Enzymatic and Non-Enzymatic Antioxidants and Stress-Related Gene Stimulation in Cucumber mosaic cucumovirus-Infected Cucumber Plants Treated with Glycine Betaine, Chitosan and Combination. Molecules 2020, 25, 2341. [Google Scholar] [CrossRef]
- Kumar, S.; Chauhan, P.S.; Agrawal, L.; Raj, R.; Srivastava, A.; Gupta, S.; Mishra, S.K.; Yadav, S.; Singh, P.C.; Raj, S.K. Paenibacillus lentimorbus Inoculation Enhances Tobacco Growth and Extenuates the Virulence of Cucumber mosaic virus. PLoS ONE 2016, 11, e0149980. [Google Scholar] [CrossRef]
- Javaheri, M.; Mashayekhi, K.; Dadkhah, A.; Tavallaee, F. Effects of salicylic acid on yield and quality characters of tomato fruit (Lycopersicum esculentum Mill.). Int. J. Agric. Crop Sci. 2012, 4, 1184–1187. [Google Scholar]
- Sirhindi, G.; Mushtaq, R.; Gill, S.S.; Sharma, P.; Abd Allah, E.F.; Ahmad, P. Jasmonic acid and methyl jasmonate modulate growth, photosynthetic activity and expression of photosystem II subunit genes in Brassica oleracea L. Sci. Rep. 2020, 10, 9322. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- Signorelli, S.; Coitino, E.L.; Borsani, O.; Monza, J. Molecular Mechanisms for the Reaction Between •OH Radicals and Proline: Insights on the Role as Reactive Oxygen Species Scavenger in Plant Stress. J. Phys. Chem. 2014, 118, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Zandi, P.; Schnug, E. Reactive Oxygen Species, Antioxidant Responses and Implications from a Microbial Modulation Perspective. Biology 2022, 11, 155. [Google Scholar] [CrossRef] [PubMed]
- Sofy, M.R.; Sharaf, A.M.A.; Noufl, M.; Sofy, A.R. Physiological and biochemical responses in Cucurbita pepo leaves associated with some elicitors-induced systemic resistance against Zucchhini yellow mosaic virus. Int. J. Mod. Bot. 2014, 4, 61–74. [Google Scholar] [CrossRef]
- Manacorda, C.A.; Gudesblat, G.; Sutka, M.; Alemano, S.; Peluso, F.; Oricchio, P.; Baroli, I.; Asurmendi, S. TuMV triggers stomatal closure but reduces drought tolerance in Arabidopsis. Plant Cell Environ. 2021, 44, 1399–1416. [Google Scholar] [CrossRef]
- Torun, H.; Novák, O.; Mikulík, J.; Strnad, M.; Ayaz, F.A. The Effects of Exogenous Salicylic Acid on Endogenous Phytohormone Status in Hordeum vulgare L. under Salt Stress. Plants 2022, 11, 618. [Google Scholar] [CrossRef]
- Misra, N.; Misra, R.; Singh, O.P. Effect of exogenous application of salicylic acid on proline metabolism in salt stressed Chamomile recutita. Niger. J. Technol. Res. 2010, 5, 30–45. [Google Scholar] [CrossRef] [Green Version]
- Thipyapong, P.; Stout, M.J.; Attajarusit, J. Functional analysis of polyphenol oxidases by antisense/sense technology. Molecules 2007, 12, 1569–1595. [Google Scholar] [CrossRef]
- Yoruk, R.; Marshall, M.R. Physicochemical properties and function of plant polyphenol oxidase: A review. J. Food Biochem. 2003, 27, 361–422. [Google Scholar] [CrossRef]
- Polovnikova, M.G.; Voskresenskaya, O.L. Activities of antioxidant system components and polyphenol oxidase in ontogeny of lawn grasses under megapolis conditions. Russ. J. Plant Physiol. 2008, 55, 699–705. [Google Scholar] [CrossRef]
- Kang, S.-M.; Khan, A.L.; Waqas, M.; You, Y.-H.; Kim, J.-H.; Kim, J.-G.; Hamayun, M.; Lee, I.-J. Plant growth-promoting rhizobacteria reduce adverse effects of salinity and osmotic stress by regulating phytohormones and antioxidants in Cucumis sativus. J. Plant Interact. 2014, 9, 673–682. [Google Scholar] [CrossRef] [Green Version]
- Dolatabadian, A.; Modarres Sanavy, S.A.M.; Sharifi, M. Effect of salicylic acid and salt on wheat seed germination. Acta Agric. Scand. 2009, 59, 456–464. [Google Scholar] [CrossRef]
- Guan, Y.; Hu, W.; Jiang, A.; Xu, Y.; Sa, R.; Feng, K.; Zhao, M.; Yu, J.; Ji, Y.; Hou, M.; et al. Effect of Methyl Jasmonate on Phenolic Accumulation in Wounded Broccoli. Molecules 2019, 24, 3537. [Google Scholar] [CrossRef] [Green Version]
- Chiappero, J.; del Rosario Cappellari, L.; Sosa Alderete, L.G.; Palermo, T.B.; Banchio, E. Plant growth promoting rhizobacteria improve the antioxidant status in Mentha piperita grown under drought stress leading to an enhancement of plant growth and total phenolic content. Ind. Crops Prod. 2019, 139, 111553. [Google Scholar] [CrossRef]
- Song, J.; Kong, Z.Q.; Zhang, D.D.; Chen, J.Y.; Dai, X.F.; Li, R. Rhizosphere Microbiomes of Potato Cultivated under Bacillus subtilis Treatment Influence the Quality of Potato Tubers. Int. J. Mol. Sci. 2021, 22, 12065. [Google Scholar] [CrossRef]
- Tkachenko, O.V.; Evseeva, N.V.; Terentyeva, E.V.; Burygin, G.L.; Shirokov, A.A.; Burov, A.M.; Matora, L.Y.; Shchyogolev, S.Y. Improved production of high-quality potato seeds in aeroponics with plant-growth-promoting rhizobacteria. Potato Res. 2021, 64, 55–66. [Google Scholar] [CrossRef]
- Yuan, J.; Cheng, L.; Li, H.; An, C.; Wang, Y.; Zhang, F. Physiological and protein profiling analysis provides insight into the underlying molecular mechanism of potato tuber development regulated by jasmonic acid in vitro. BMC Plant Biol. 2022, 22, 481. [Google Scholar] [CrossRef]
- Sánchez-Rojo, S.; López-Delgado, H.A.; Mora-Herrera, M.E.; Almeyda-León, H.I.; Zavaleta-Mancera, H.A.; Espinosa-Victoria, D. Salicylic acid protects potato plants-from phytoplasma-associated stress and improves tuber photosynthate assimilation. Am. J. Potato Res. 2011, 88, 175–183. [Google Scholar] [CrossRef]
- Aksenova, N.P.; Konstantinova, T.N.; Golyanovskaya, S.A.; Sergeeva, L.I.; Romanov, G.A. Hormonal regulation of tuber formation in potato plants. Russ. J. Plant Physiol. 2012, 59, 451–466. [Google Scholar] [CrossRef]
- Upadhyaya, C.P.; Akula, N.; Young, K.E.; Chun, S.C.; Kim, D.H.; Park, S.W. Enhanced ascorbic acid accumulation in transgenic potato confers tolerance to various abiotic stresses. Biotechnol. Lett. 2010, 32, 321–330. [Google Scholar] [CrossRef]
- Naeem, M.; Basit, A.; Ahmad, I.; Mohamed, H.I.; Wasila, H. Effect of Salicylic Acid and Salinity Stress on the Performance of Tomato Plants. Gesunde Pflanz. 2020, 72, 393–402. [Google Scholar] [CrossRef]
- Youssif, S.B.D. Response of potatoes to foliar spray with cobalamin, folic acid and ascorbic acid under North Sinai conditions. Middle East J. Agric. Res. 2017, 6, 662–672. [Google Scholar]
- Zeeman, S.C.; Thorneycroft, D.; Schupp, N.; Chapple, A.; Weck, M.; Dunstan, H.; Haldimann, P.; Bechtold, N.; Smith, A.M.; Smith, S.M. Plastidial alpha-glucan phosphorylase is not required for starch degradation in Arabidopsis leaves but has a role in the tolerance of abiotic stress. Plant Physiol. 2004, 135, 849–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, H.; Shibahara, K.; Tokunaga, T.; Nishi, A.; Tasaki, M.; Hwang, S.K.; Okita, T.W.; Kaneko, N.; Fujita, N.; Yoshida, M.; et al. Mutation of the plastidial alpha-glucan phosphorylase gene in rice affects the synthesis and structure of starch in the endosperm. Plant Cell 2008, 20, 1833–1849. [Google Scholar] [CrossRef] [Green Version]
- Berka, M.; Kopecká, R.; Berková, V.; Brzobohatý, B.; Černý, M. Regulation of heat shock proteins 70 and their role in plant immunity. J. Exp. Bot. 2022, 73, 1894–1909. [Google Scholar] [CrossRef]
- Shi, H.; He, S.; He, X. An eukaryotic elongation factor 2 from Medicago falcata (MfEF2) confers cold tolerance. BMC Plant Biol. 2019, 19, 218. [Google Scholar] [CrossRef]
- Kim, D.S.; Jeun, Y.; Hwang, B.K. The pepper patatin-like phospholipase CaPLP1 functions in plant cell death and defense signaling. Plant Mol. Biol. 2014, 84, 329–344. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Ismail, I.; Dessoky, E.; El-Hallous, E.; Hafez, E.A. Tomato kinesin-like protein is associated with Tobacco mosaic virus infection. Biotechnol. Biotechnol. Equip. 2019, 33, 1424–1433. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.M.; Liu, W.C.; Lu, Y.T. CATALASE2 Coordinates SA-Mediated Repression of Both Auxin Accumulation and JA Biosynthesis in Plant Defenses. Cell Host Microbe 2017, 21, 143–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Vemanna, R.S.; Oh, S.; Rojas, C.M.; Oh, Y.; Kaundal, A.; Kwon, T.; Lee, H.K.; Senthil-Kumar, M.; Mysore, K.S. Functional role of formate dehydrogenase 1 (FDH1) for host and nonhost disease resistance against bacterial pathogens. PLoS ONE 2022, 17, e0264917. [Google Scholar] [CrossRef] [PubMed]
- Osmani, Z.; Sabet, M.S.; Nakahara, K.S. Aspartic protease inhibitor enhances resistance to potato virus Y and A in transgenic potato plants. BMC Plant Biol. 2022, 22, 241. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, J.; Laohavisit, A.; Macpherson, N.; Webb, A.; Brownlee, C.; Battey, N.; Davies, J. Annexins: Multifunctional components of growth and adaptation. J. Exp. Bot. 2008, 59, 533–544. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
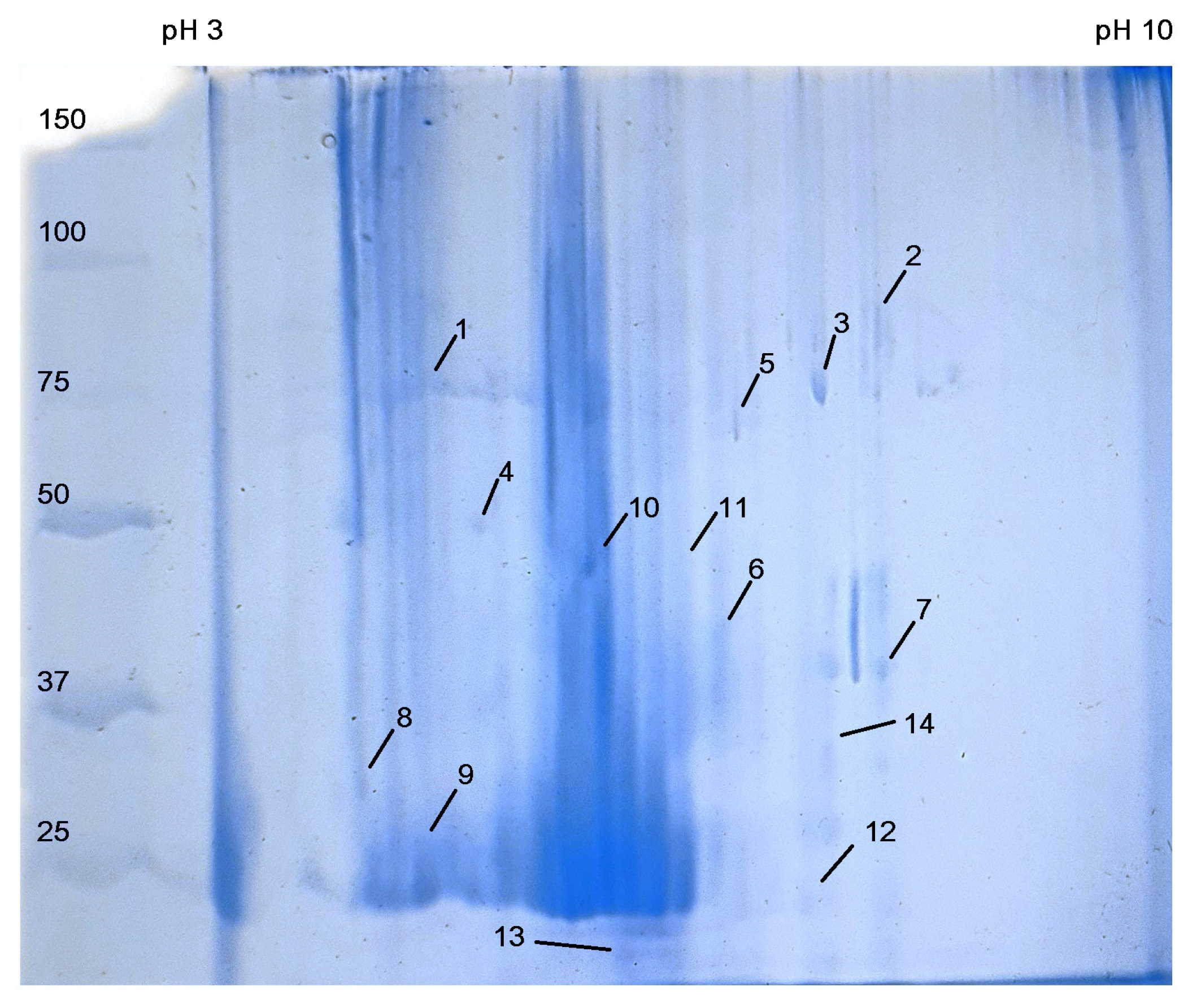{kind=link}
| Optimal Condition | Water Deficiency | |||
|---|---|---|---|---|
| Treatment | Dry Matter % | Chlorophylls Content, mg/g DM | Dry Matter % | Chlorophylls Content, mg/g DM |
| Control | 12.8 ± 0.39 | 12.7 ± 0.61 | 13.7 ± 0.37 | 20.1 ± 0.61 |
| PVY | 13.5 ± 0.11 a | 11.2 ± 0.71 | 13.7 ± 0.22 | 13.1 ± 0.71 a |
| B. subtilis | 13.3 ± 0.12 a | 11.5 ± 0.15 | 13.8 ± 0.24 | 15.5 ± 0.15 ab |
| B. subtilis + SA | 13.4 ± 0.13 a | 10.3 ± 0.56 a | 14.5 ± 0.08 ab | 15.7 ± 0.56 ab |
| B. subtilis + MeJ | 12.2 ± 0.18 b | 11.7 ± 0.34 | 15.3 ± 0.10 ab | 16.8 ± 0.34 ab |
| B. subtilis + MeJ + SA | 12.8 ± 0.25 b | 10.9 ± 0.71 | 14.5 ± 0.25 ab | 18.7 ± 0.71 b |
| Treatment | Number of Minitubers, pcs/Container | Weight of Minitubers, g/Container | Starch Content, % | Ascorbic Acid Content, µg/g WM |
|---|---|---|---|---|
| Control | 28.5 ± 2.2 | 365.0 ± 23.7 | 12.9 ± 0.78 | 26.3 ± 3.3 |
| PVY | 31.0 ± 2.5 | 337.9 ± 26.1 | 12.6 ± 0.10 | 21.4 ± 3.1 |
| B. subtilis | 39.5 ± 3.2 | 351.6 ± 26.4 | 14.0 ± 0.29 | 27.5 ± 4.2 |
| B. subtilis + SA | 36.0 ± 3.4 | 360.0 ± 25.5 | 13.3 ± 0.69 | 32.0 ± 6.1 |
| B. subtilis + MeJ | 38.5 ± 2.8 | 358.8.0 ± 27.3 | 14.4 ± 0.20 | 26.5 ± 5.0 |
| B. subtilis + MeJ + SA | 42.5 ± 3.7 | 361.2 ± 25.3 | 12.0 ± 0.11 | 30.1 ± 5.8 |
| No. | The Name of the Protein According to Uniprot | pI | Protein Content, µg/g Fresh Weight | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Optimal Conditions | Soil Water Deficiency | |||||||||||||
| Control | Infection PVY | Infection + B. subtilis | Infection + B. subtilis + SA | Infection + B. subtilis + MeJ | Infection + B. subtilis + SA + MeJ | Control | Infection | Infection + B. subtilis | Infection + B. subtilis + SA | Infection + B. subtilis + MeJ | Infection + B. subtilis + SA + MeJ | |||
| 1 | Alpha-glucan phosphorylase | 4 | - * | - | - | - | - | 0.1 | - | - | - | - | - | - |
| 2 | 7 | - | - | - | - | - | 0.1 | - | - | - | - | - | - | |
| 3 | 9 | - | - | - | - | - | - | - | - | 0.1 | - | - | 0.1 | |
| 4 | Stromal 70 kDa heat shock-related protein, chloroplastic | 4.5 | - | - | 0.1 | 0.1 | - | - | - | - | - | - | - | - |
| 5 | Elongation factor 2 | 6 | - | - | - | - | - | - | - | - | 0.1 | - | - | - |
| 6 | Kinesin-like protein KIN-5D | 7 | - | - | - | - | - | - | - | - | - | 6 | - | - |
| 7 | Catalase isozyme 2 | 8 | - | - | - | - | - | 0.2 | - | - | - | 0.1 | 0.1 | 0.1 |
| 8 | Patatin | 3.5 | - | - | 0.1 | 0.1 | - | 0.1 | - | - | 4.3 | - | - | - |
| 9 | 4 | - | 2.1 | - | 0.1 | 0.1 | 2.5 | - | 2.3 | 3.5 | 2.5 | - | 0.1 | |
| 10 | 5 | - | 3.2 | - | - | - | - | - | - | - | 2.6 | - | 0.1 | |
| 11 | 6 | - | - | - | - | - | - | - | - | 2.4 | 3.4 | - | - | |
| 12 | Aspartic protease inhibitor 8 | 7 | - | - | 3.6 | 5.7 | 3.1 | - | 0.2 | - | - | 2.8 | 0.1 | - |
| 13 | Annexin D2-like | 5 | 5.5 | 5.6 | 3.5 | - | 6.1 | 6.0 | - | 5.8 | - | 4.1 | - | - |
| 14 | Formate dehydrogenase, mitochondrial | 8 | - | 0.2 | - | - | - | - | - | 3.0 | - | - | 0.1 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalatskaja, J.N.; Baliuk, N.V.; Rybinskaya, K.I.; Herasimovich, K.M.; Yalouskaya, N.A.; Yarullina, L.G.; Tsvetkov, V.O. Increasing Potato Sustainability to PVY under Water Deficiency by Bacillus Bacteria with Salicylic Acid and Methyl Jasmonate. Int. J. Plant Biol. 2023, 14, 312-328. https://doi.org/10.3390/ijpb14010026
Kalatskaja JN, Baliuk NV, Rybinskaya KI, Herasimovich KM, Yalouskaya NA, Yarullina LG, Tsvetkov VO. Increasing Potato Sustainability to PVY under Water Deficiency by Bacillus Bacteria with Salicylic Acid and Methyl Jasmonate. International Journal of Plant Biology. 2023; 14(1):312-328. https://doi.org/10.3390/ijpb14010026
Chicago/Turabian StyleKalatskaja, Joanna N., Natallia V. Baliuk, Katsiaryna I. Rybinskaya, Kanstantsin M. Herasimovich, Ninel A. Yalouskaya, Lubov G. Yarullina, and Vyacheslav O. Tsvetkov. 2023. "Increasing Potato Sustainability to PVY under Water Deficiency by Bacillus Bacteria with Salicylic Acid and Methyl Jasmonate" International Journal of Plant Biology 14, no. 1: 312-328. https://doi.org/10.3390/ijpb14010026