
Comparative Evaluation of Paclobutrazol and Rhizobacterium Variovorax sp. YNA59 Effects on Mitigating Drought Stress in Chinese Cabbage (Brassica rapa ssp. Pekinensis)


Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Materials and Selection of Rhizobacterium and Paclobutrazol
2.2. Experimental Design
2.3. Plant Experiment
2.4. Analysis of Biochemical Properties
2.4.1. Chlorophyll Measurement
2.4.2. Relative Water Content Measurement
2.4.3. Relative Water Loss (ELWL) Calculation
2.4.4. ABA Analysis
2.4.5. Antioxidant Activities
- Glutathione (GSH) Measurement
- Catalase (CAT) Measurement
- Polyphenol oxidase (PPO) measurement
- Peroxidase (POD) measurement
2.5. Statistical Analysis
3. Results
3.1. Morphological Characteristics
3.2. Chlorophyll Measurement
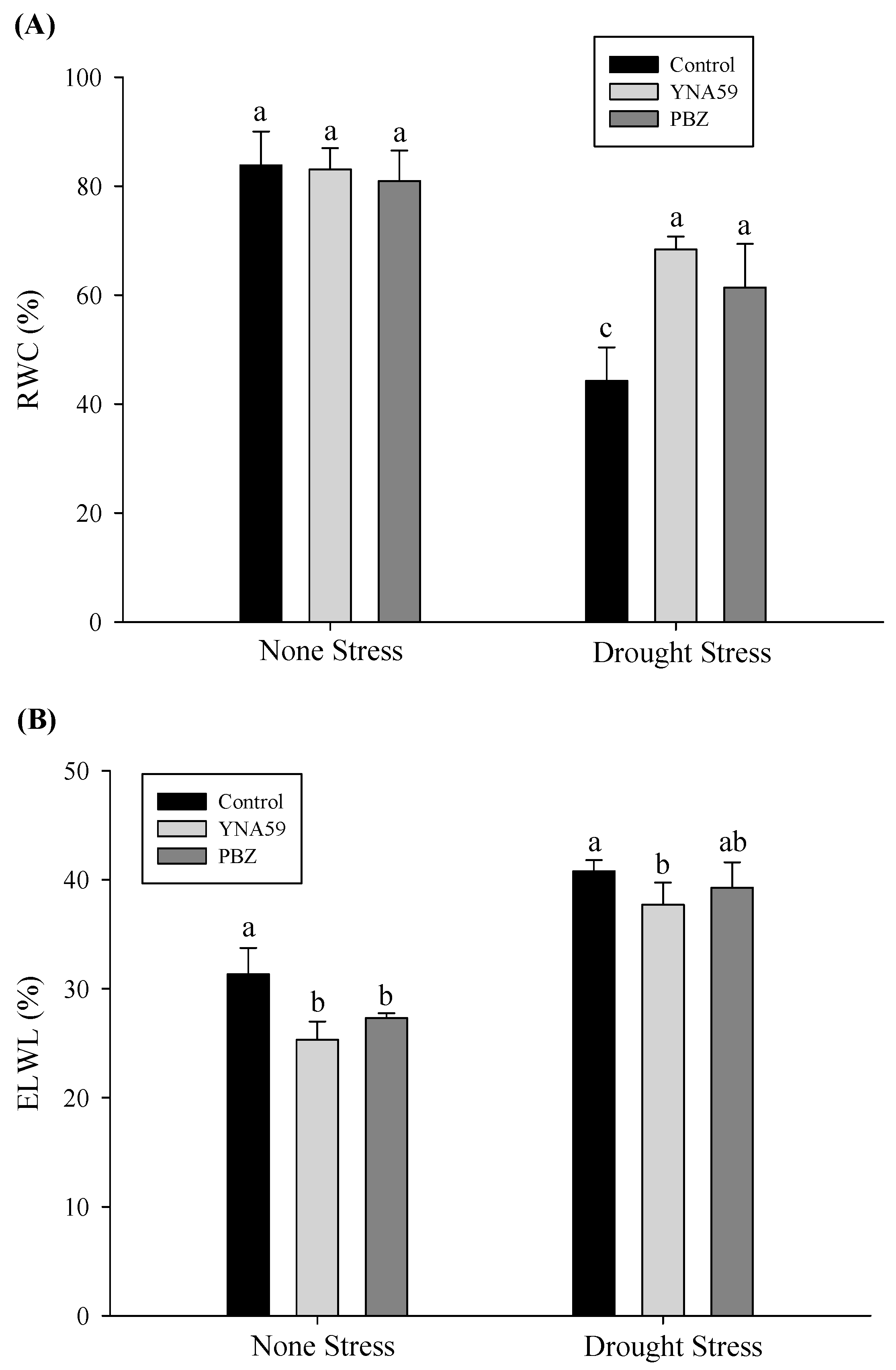
3.3. Relative Water Content Measurement
3.4. Relative Water Loss (ELWL) Measurement
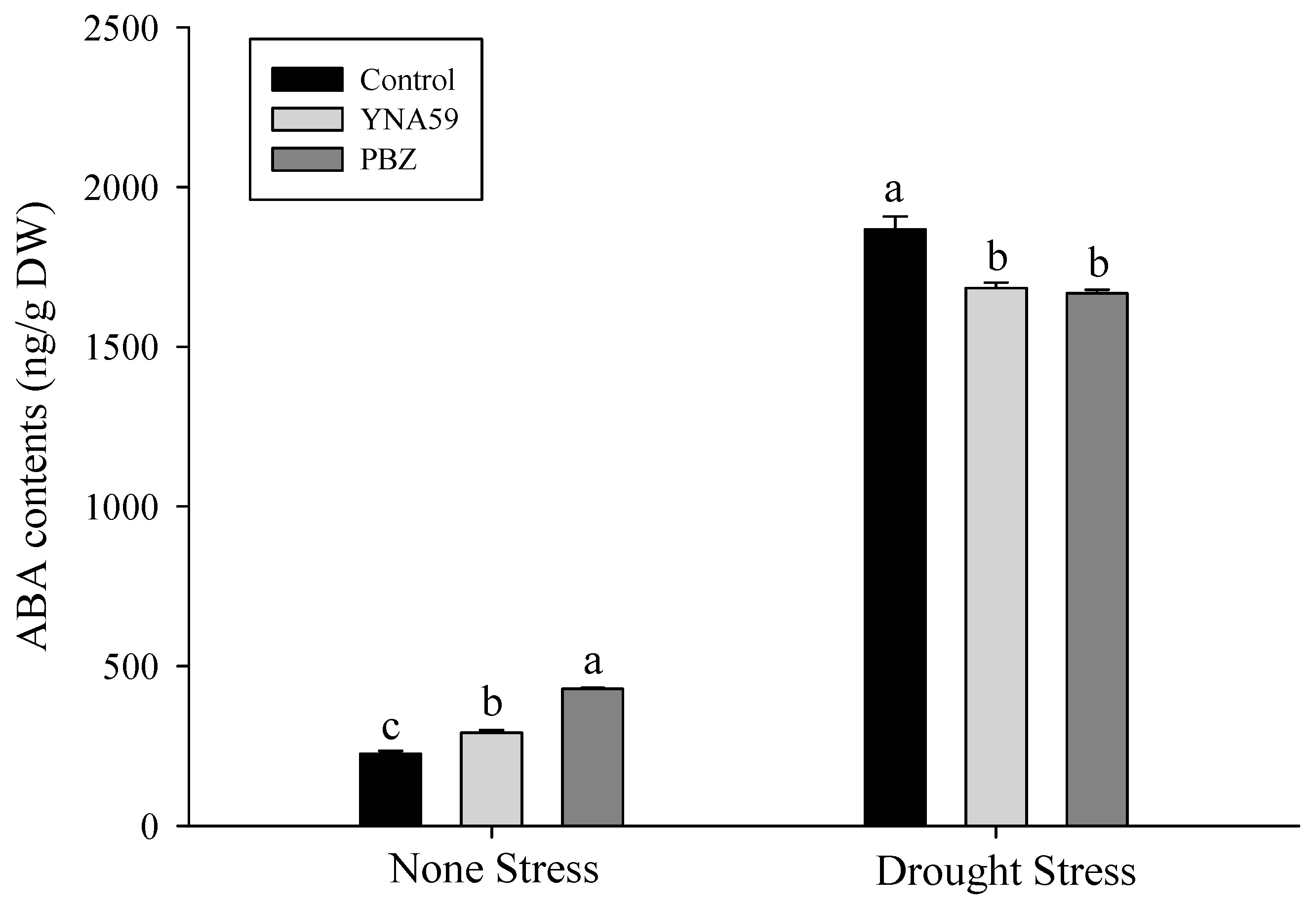
3.5. Quantification of Endogenous Phytohormone Abscisic Acid (ABA)
3.6. Analysis of Antioxidant Enzymes
3.7. Measurement of Polyphenol Oxidase (PPO) and Peroxidase (POD)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| NT | No treatment |
| YNA59 | Rhizobacterium Variovorax sp. Inoculum |
| PBZ | Paclobutrazol |
| NS | No stress |
| DS | Drought stress |
Appendix A
References
- Matiu, M.; Ankerst, D.P.; Menzel, A. Interactions between temperature and drought in global and regional crop yield variability during 1961–2014. PLoS ONE 2017, 12, e0178339. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Iizumi, T.; Nishimori, M. Global patterns of crop production losses associated with droughts from 1983 to 2009. J. Appl. Meteorol. Climatol. 2019, 58, 1233–1244. [Google Scholar] [CrossRef]
- Premanandh, J. Factors affecting food security and contribution of modern technologies in food sustainability. J. Sci. Food Agric. 2011, 91, 2707–2714. [Google Scholar] [CrossRef] [PubMed]
- Fariduddin, Q.; Zaid, A.; Mohammad, F. Plant growth regulators and salt stress: Mechanism of tolerance trade-off. Salt Stress Microbes Plant Interact. Causes Solut. 2019, 1, 91–111. [Google Scholar]
- Adhikari, A.; Khan, M.A.; Imran, M.; Lee, K.-E.; Kang, S.-M.; Shin, J.Y.; Joo, G.-J.; Khan, M.; Yun, B.-W.; Lee, I.-J. The Combined Inoculation of Curvularia lunata AR11 and Biochar Stimulates Synthetic Silicon and Potassium Phosphate Use Efficiency, and Mitigates Salt and Drought Stresses in Rice. Front. Plant Sci. 2022, 13, 816858. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H.M. Drought stress in plants: An overview. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–33. [Google Scholar]
- Munné-Bosch, S.; Villadangos, S. Cheap, cost-effective, and quick stress biomarkers for drought stress detection and monitoring in plants. Trends Plant Sci. 2023, 28, 527–536. [Google Scholar] [CrossRef]
- Harb, A.; Krishnan, A.; Ambavaram, M.M.R.; Pereira, A. Molecular and physiological analysis of drought stress in Arabidopsis reveals early responses leading to acclimation in plant growth. Plant Physiol. 2010, 154, 1254–1271. [Google Scholar] [CrossRef]
- Wang, Y.-T.; Chen, Z.-Y.; Jiang, Y.; Duan, B.-B.; Xi, Z.-M. Involvement of ABA and antioxidant system in brassinosteroid-induced water stress tolerance of grapevine (Vitis vinifera L.). Sci. Hortic. 2019, 256, 108596. [Google Scholar] [CrossRef]
- Lata, R.; Gond, S.K. 6—Plant growth-promoting microbes for abiotic stress tolerance in plants. In Role of Plant Growth Promoting Microorganisms in Sustainable Agriculture and Nanotechnology; Kumar, A., Singh, A.K., Choudhary, K.K., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 89–105. [Google Scholar]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and salinity stress responses and microbe-induced tolerance in plants. Front. Plant Sci. 2020, 11, 591911. [Google Scholar] [CrossRef]
- Coleman-Derr, D.; Tringe, S.G. Building the crops of tomorrow: Advantages of symbiont-based approaches to improving abiotic stress tolerance. Front. Microbiol. 2014, 5, 283. [Google Scholar] [CrossRef]
- Ha-Tran, D.M.; Nguyen, T.T.M.; Hung, S.-H.; Huang, E.; Huang, C.-C. Roles of plant growth-promoting rhizobacteria (PGPR) in stimulating salinity stress defense in plants: A review. Int. J. Mol. Sci. 2021, 22, 3154. [Google Scholar] [CrossRef] [PubMed]
- Bal, H.B.; Nayak, L.; Das, S.; Adhya, T.K. Isolation of ACC deaminase producing PGPR from rice rhizosphere and evaluating their plant growth promoting activity under salt stress. Plant Soil 2013, 366, 93–105. [Google Scholar] [CrossRef]
- Batool, T.; Ali, S.; Seleiman, M.F.; Naveed, N.H.; Ali, A.; Ahmed, K.; Abid, M.; Rizwan, M.; Shahid, M.R.; Alotaibi, M. Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci. Rep. 2020, 10, 16975. [Google Scholar] [CrossRef] [PubMed]
- Notununu, I.; Moleleki, L.; Roopnarain, A.; Adeleke, R. Effects of plant growth-promoting rhizobacteria on the molecular responses of maize under drought and heat stresses: A review. Pedosphere 2022, 32, 90–106. [Google Scholar] [CrossRef]
- Ashraf, A.; Bano, A.; Ali, S.A. Characterisation of plant growth-promoting rhizobacteria from rhizosphere soil of heat-stressed and unstressed wheat and their use as bio-inoculant. Plant Biol. 2019, 21, 762–769. [Google Scholar] [CrossRef]
- Khanna, K.; Jamwal, V.L.; Gandhi, S.G.; Ohri, P.; Bhardwaj, R. Metal resistant PGPR lowered Cd uptake and expression of metal transporter genes with improved growth and photosynthetic pigments in Lycopersicon esculentum under metal toxicity. Sci. Rep. 2019, 9, 5855. [Google Scholar] [CrossRef]
- Cordero, I.; Balaguer, L.; Rincón, A.; Pueyo, J.J. Inoculation of tomato plants with selected PGPR represents a feasible alternative to chemical fertilization under salt stress. J. Plant Nutr. Soil Sci. 2018, 181, 694–703. [Google Scholar] [CrossRef]
- Hajihashemi, S.; Kiarostami, K.; Saboora, A.; Enteshari, S. Exogenously applied paclobutrazol modulates growth in salt-stressed wheat plants. Plant Growth Regul. 2007, 53, 117–128. [Google Scholar] [CrossRef]
- Desta, B.; Amare, G. Paclobutrazol as a plant growth regulator. Chem. Biol. Technol. Agric. 2021, 8, 1. [Google Scholar] [CrossRef]
- Fan, Z.X.; Li, S.C.; Sun, H.L. Paclobutrazol modulates physiological and hormonal changes in Amorpha fruticosa under drought stress. Russ. J. Plant Physiol. 2020, 67, 122–130. [Google Scholar] [CrossRef]
- Shalaby, T.A.; Taha, N.A.; Taher, D.I.; Metwaly, M.M.; El-Beltagi, H.S.; Rezk, A.A.; El-Ganainy, S.M.; Shehata, W.F.; El-Ramady, H.R.; Bayoumi, Y.A. Paclobutrazol Improves the Quality of Tomato Seedlings to Be Resistant to Alternaria solani Blight Disease: Biochemical and Histological Perspectives. Plants 2022, 11, 425. [Google Scholar] [CrossRef]
- Cho, J.; Lee, D.; Yang, C.; Jeon, J.; Kim, J.; Han, H. Microbial population dynamics of kimchi, a fermented cabbage product. FEMS Microbiol. Lett. 2006, 257, 262–267. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, J.H.; Lee, S.G.; An, S.; Lee, H.S.; Choi, C.K.; Kim, S.K. Foliar application of biostimulants affects physiological responses and improves heat stress tolerance in Kimchi cabbage. Hortic. Environ. Biotechnol. 2019, 60, 841–851. [Google Scholar] [CrossRef]
- Jang, D.-J.; Chung, K.R.; Yang, H.J.; Kim, K.-s.; Kwon, D.Y. Discussion on the origin of kimchi, representative of Korean unique fermented vegetables. J. Ethn. Foods 2015, 2, 126–136. [Google Scholar] [CrossRef]
- Lee, S.G.; Seo, T.C.; Jang, Y.A.; Lee, J.G.; Nam, C.W.; Choi, C.S.; Yeo, K.-H.; Um, Y.C. Prediction of Chinese cabbage yield as affected by planting date and nitrogen fertilization for spring production. J. Bio-Environ. Control 2012, 21, 271–275. [Google Scholar]
- Oh, S.; Moon, K.H.; Son, I.-C.; Song, E.Y.; Moon, Y.E.; Koh, S.C. Growth, photosynthesis and chlorophyll fluorescence of Chinese cabbage in response to high temperature. Hortic. Sci. Technol. 2014, 32, 318–329. [Google Scholar]
- Hussain, A.; Audira, G.; Siregar, P.; Lin, Y.-C.; Villalobos, O.; Villaflores, O.; Wang, W.-D.; Hsiao, C.-D. Waterborne Exposure of Paclobutrazol at Environmental Relevant Concentration Induce Locomotion Hyperactivity in Larvae and Anxiolytic Exploratory Behavior in Adult Zebrafish. Int. J. Environ. Res. Public Health 2020, 17, 4632. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-D.; Wu, C.-Y.; Lonameo, B.K. Toxic Effects of Paclobutrazol on Developing Organs at Different Exposure Times in Zebrafish. Toxics 2019, 7, 62. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, Y.; Xie, H.; Li, R.; Wei, J.; Liu, Y. Environmental behavior of paclobutrazol in soil and its toxicity on potato and taro plants. Environ. Sci. Pollut. Res. 2019, 26, 27385–27395. [Google Scholar] [CrossRef]
- Kim, Y.-N.; Khan, M.A.; Kang, S.-M.; Hamayun, M.; Lee, I.-J. Enhancement of Drought-Stress Tolerance of Brassica oleracea var. italica L. by Newly Isolated Variovorax sp. YNA59. J. Microbiol. Biotechnol. 2020, 30, 1500–1509. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1. [Google Scholar] [CrossRef] [PubMed]
- Lubna; Asaf, S.; Hamayun, M.; Khan, A.L.; Waqas, M.; Khan, M.A.; Jan, R.; Lee, I.-J.; Hussain, A. Salt tolerance of Glycine max L. induced by endophytic fungus Aspergillus flavus CSH1, via regulating its endogenous hormones and antioxidative system. Plant Physiol. Biochem. 2018, 128, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.-R.; Adhikari, A.; Kang, Y.; Gam, H.-J.; Kang, S.-M.; Kim, K.-Y.; Lee, I.-J. Investigation of Solanum carolinense Dominance and Phytotoxic Effect in Festuca arundinacea with Special Reference to Allelochemical Identification, Analysis of Phytohormones and Antioxidant Mechanisms. Agronomy 2022, 12, 1954. [Google Scholar] [CrossRef]
- Khan, A.L.; Waqas, M.; Hussain, J.; Al-Harrasi, A.; Hamayun, M.; Lee, I.-J. Phytohormones enabled endophytic fungal symbiosis improve aluminum phytoextraction in tolerant Solanum lycopersicum: An examples of Penicillium janthinellum LK5 and comparison with exogenous GA3. J. Hazard. Mater. 2015, 295, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase. In Methods of Enzymatic Analysis; Elsevier: Amsterdam, The Netherlands, 1974; pp. 673–684. [Google Scholar]
- Khan, A.R.; Ullah, I.; Waqas, M.; Park, G.-S.; Khan, A.L.; Hong, S.-J.; Ullah, R.; Jung, B.K.; Park, C.E.; Ur-Rehman, S.; et al. Host plant growth promotion and cadmium detoxification in Solanum nigrum, mediated by endophytic fungi. Ecotoxicol. Environ. Saf. 2017, 136, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Pütter, J. Peroxidases. In Methods of Enzymatic Analysis; Elsevier: Amsterdam, The Netherlands, 1974; pp. 685–690. [Google Scholar]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef]
- Arif, M.; Jan, T.; Riaz, M.; Fahad, S.; Arif, M.S.; Shakoor, M.B.; Rasul, F. Advances in rice research for abiotic stress. In Advances in Rice Research for Abiotic Stress Tolerance; Elsevier: Amsterdam, The Netherlands, 2019; pp. 585–614. [Google Scholar]
- Hossain, M.M.; Sultana, F.; Islam, S. Plant growth-promoting fungi (PGPF): Phytostimulation and induced systemic resistance. Plant-Microbe Interact. Agro-Ecol. Perspect. Microb. Interact. Agro-Ecol. Impacts 2017, 2, 135–191. [Google Scholar]
- Zou, Y.N.; Wu, Q.S.; Kuča, K. Unravelling the role of arbuscular mycorrhizal fungi in mitigating the oxidative burst of plants under drought stress. Plant Biol. 2021, 23, 50–57. [Google Scholar] [CrossRef]
- Sheikh Mohammadi, M.H.; Etemadi, N.; Arab, M.M.; Aalifar, M.; Arab, M.; Pessarakli, M. Molecular and physiological responses of Iranian Perennial ryegrass as affected by Trinexapac ethyl, Paclobutrazol and Abscisic acid under drought stress. Plant Physiol. Biochem. 2017, 111, 129–143. [Google Scholar] [CrossRef]
- Davari, K.; Rokhzadi, A.; Mohammadi, K.; Pasari, B. Paclobutrazol and Amino Acid-Based Biostimulant as Beneficial Compounds in Alleviating the Drought Stress Effects on Safflower (Carthamus tinctorius L.). J. Soil Sci. Plant Nutr. 2022, 22, 674–690. [Google Scholar] [CrossRef]
- Still, J.R.; Pill, W.G. Growth and stress tolerance of tomato seedlings (Lycopersicon esculentum Mill.) in response to seed treatment with paclobutrazol. J. Hortic. Sci. Biotechnol. 2004, 79, 197–203. [Google Scholar] [CrossRef]
- Ferioun, M.; Bouhraoua, S.; Srhiouar, N.; Tirry, N.; Belahcen, D.; Siang, T.C.; Louahlia, S.; El Ghachtouli, N. Optimized drought tolerance in barley (Hordeum vulgare L.) using plant growth-promoting rhizobacteria (PGPR). Biocatal. Agric. Biotechnol. 2023, 50, 102691. [Google Scholar] [CrossRef]
- Gul, L.; Ullah, S.; Nafees, M.; Ahmed, I. Resistance induction in Brassica napus L. against water deficit stress through application of biochar and plant growth promoting rhizobacteria. J. Saudi Soc. Agric. Sci. 2023, in press. [Google Scholar] [CrossRef]
- Burgess, P.; Huang, B. Mechanisms of hormone regulation for drought tolerance in plants. Drought Stress Toler. Plants Physiol. Biochem. 2016, 1, 45–75. [Google Scholar]
- Singroha, G.; Sharma, P.; Sunkur, R. Current status of microRNA-mediated regulation of drought stress responses in cereals. Physiol. Plant. 2021, 172, 1808–1821. [Google Scholar] [CrossRef]
- Shahzad, R.; Khan, A.L.; Bilal, S.; Waqas, M.; Kang, S.-M.; Lee, I.-J. Inoculation of abscisic acid-producing endophytic bacteria enhances salinity stress tolerance in Oryza sativa. Environ. Exp. Bot. 2017, 136, 68–77. [Google Scholar] [CrossRef]
- Kang, S.M.; Adhikari, A.; Lee, K.E.; Khan, M.A.; Khan, A.L.; Shahzad, R.; Dhungana, S.K.; Lee, I.J. Inoculation with Indole-3-Acetic Acid-Producing Rhizospheric Rhodobacter sphaeroides KE149 Augments Growth of Adzuki Bean Plants under Water Stress. J. Microbiol. Biotechnol. 2020, 30, 717–725. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar]
- Parveen, A.; Liu, W.; Hussain, S.; Asghar, J.; Perveen, S.; Xiong, Y. Silicon Priming Regulates Morpho-Physiological Growth and Oxidative Metabolism in Maize under Drought Stress. Plants 2019, 8, 431. [Google Scholar] [CrossRef]
- Koffler, B.E.; Luschin-Ebengreuth, N.; Stabentheiner, E.; Müller, M.; Zechmann, B. Compartment specific response of antioxidants to drought stress in Arabidopsis. Plant Sci. 2014, 227, 133–144. [Google Scholar] [PubMed]
- Kocsy, G.; Szalai, G.; Galiba, G. Induction of glutathione synthesis and glutathione reductase activity by abiotic stresses in maize and wheat. Sci. World J. 2002, 2, 1699–1705. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, Y.; Yao, H.; Wu, J.; Sun, H.; Gong, H. Silicon improves seed germination and alleviates oxidative stress of bud seedlings in tomato under water deficit stress. Plant Physiol. Biochem. 2014, 78, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Staszek, P.; Krasuska, U.; Ciacka, K.; Gniazdowska, A. ROS Metabolism Perturbation as an Element of Mode of Action of Allelochemicals. Antioxidants 2021, 10, 1648. [Google Scholar] [CrossRef]
- Chen, J.-H.; Jiang, H.-W.; Hsieh, E.-J.; Chen, H.-Y.; Chien, C.-T.; Hsieh, H.-L.; Lin, T.-P. Drought and salt stress tolerance of an Arabidopsis glutathione S-transferase U17 knockout mutant are attributed to the combined effect of glutathione and abscisic acid. Plant Physiol. 2012, 158, 340–351. [Google Scholar] [PubMed]
- Saruhan, N.; Terzi, R.; Saglam, A.; Kadioglu, A. The relationship between leaf rolling and ascorbate-glutathione cycle enzymes in apoplastic and symplastic areas of Ctenanthe setosa subjected to drought stress. Biol. Res. 2009, 42, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Hernández, I.; Cela, J.; Alegre, L.; Munné-Bosch, S. Antioxidant defenses against drought stress. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Springer: Berlin/Heidelberg, Germany, 2012; pp. 231–258. [Google Scholar]
- Bhargava, S.; Sawant, K. Drought stress adaptation: Metabolic adjustment and regulation of gene expression. Plant Breed. 2013, 132, 21–32. [Google Scholar]
- Li, J.; Guo, X.; Zhang, M.; Wang, X.; Zhao, Y.; Yin, Z.; Zhang, Z.; Wang, Y.; Xiong, H.; Zhang, H.; et al. OsERF71 confers drought tolerance via modulating ABA signaling and proline biosynthesis. Plant Sci. 2018, 270, 131–139. [Google Scholar] [CrossRef]
- Saeedipour, S. Relationship of Grain Yield, ABA and Proline Accumulation in Tolerant and Sensitive Wheat Cultivars as Affected by Water Stress. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2013, 83, 311–315. [Google Scholar] [CrossRef]
- Yuan, X.K.; Yang, Z.Q.; Li, Y.X.; Liu, Q.; Han, W. Effects of different levels of water stress on leaf photosynthetic characteristics and antioxidant enzyme activities of greenhouse tomato. Photosynthetica 2016, 54, 28–39. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Glutathione in plants: Biosynthesis and physiological role in environmental stress tolerance. Physiol. Mol. Biol. Plants 2017, 23, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Soumya, P.R.; Kumar, P.; Pal, M. Paclobutrazol: A novel plant growth regulator and multi-stress ameliorant. Indian J. Plant Physiol. 2017, 22, 267–278. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shoot Length (cm) | Root Length (cm) | Total Fresh Biomass (g) | Leaf Length (cm) | Leaf Width (cm) | |
|---|---|---|---|---|---|
| None Stress | |||||
| NT | 20.9 ± 1.22 a | 15.2 ± 0.87 b | 6.9 ± 0.31 b | 19.4 ± 1.23 a | 8.8 ± 0.74 a |
| YNA59 | 21.2 ± 1.53 a | 18.4 ± 1.38 a | 7.2 ± 0.27 a | 20.7 ± 1.04 a | 9.6 ± 0.56 a |
| PBZ | 14.4 ± 0.98 b | 14.4 ± 20.95 b | 6.1 ± 0.12 c | 13.0 ± 0.78 b | 7.3 ± 0.33 b |
| Drought Stress | |||||
| NT | 15.1 ± 2.42 b | 13.1 ± 0.65 b | 5.0 ± 0.12 c | 14.4 ± 0.78 b | 6.7 ± 0.54 b |
| YNA59 | 18.8 ± 1.01 a | 17.1 ± 1.33 a | 6.4 ± 0.25 a | 17.7 ± 0.69 a | 8.3 ± 0.36 a |
| PBZ | 13.4 ± 0.87 b | 11.3 ± 0.93 a | 5.4 ± 0.21 b | 10.1 ± 0.56 c | 6.1 ± 0.57 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, S.-M.; Adhikari, A.; Kim, Y.-N.; Lee, I.-J. Comparative Evaluation of Paclobutrazol and Rhizobacterium Variovorax sp. YNA59 Effects on Mitigating Drought Stress in Chinese Cabbage (Brassica rapa ssp. Pekinensis). Microbiol. Res. 2023, 14, 1210-1224. https://doi.org/10.3390/microbiolres14030081
Kang S-M, Adhikari A, Kim Y-N, Lee I-J. Comparative Evaluation of Paclobutrazol and Rhizobacterium Variovorax sp. YNA59 Effects on Mitigating Drought Stress in Chinese Cabbage (Brassica rapa ssp. Pekinensis). Microbiology Research. 2023; 14(3):1210-1224. https://doi.org/10.3390/microbiolres14030081
Chicago/Turabian StyleKang, Sang-Mo, Arjun Adhikari, Yu-Na Kim, and In-Jung Lee. 2023. "Comparative Evaluation of Paclobutrazol and Rhizobacterium Variovorax sp. YNA59 Effects on Mitigating Drought Stress in Chinese Cabbage (Brassica rapa ssp. Pekinensis)" Microbiology Research 14, no. 3: 1210-1224. https://doi.org/10.3390/microbiolres14030081