
Bacillus Metabolites: Compounds, Identification and Anti-Candida albicans Mechanisms

Abstract
:1. Introduction
2. Inhibition of Bacillus Metabolites on C. albicans
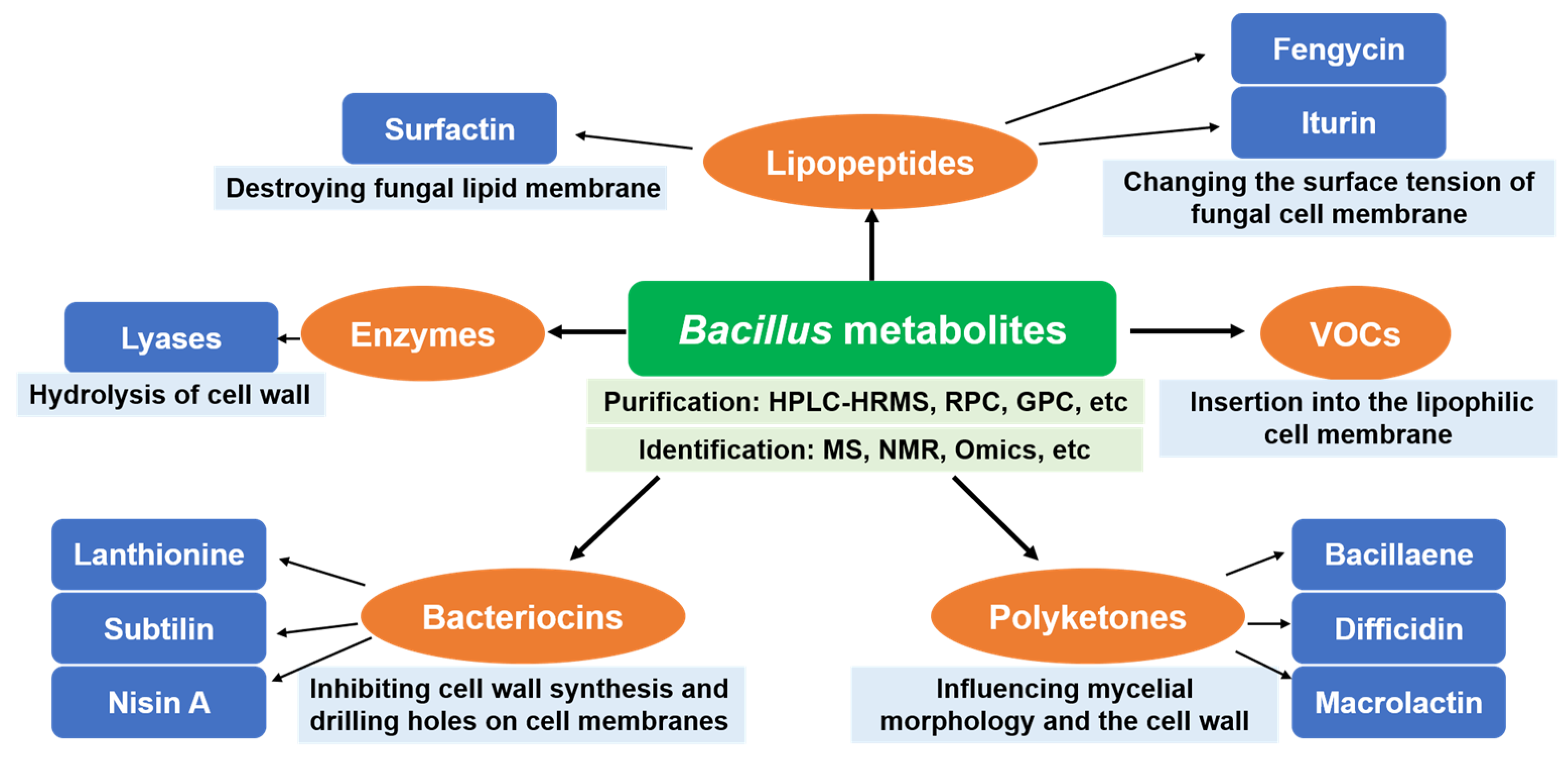
2.1. Lipopeptides
2.2. Enzymes
2.3. Polyketones
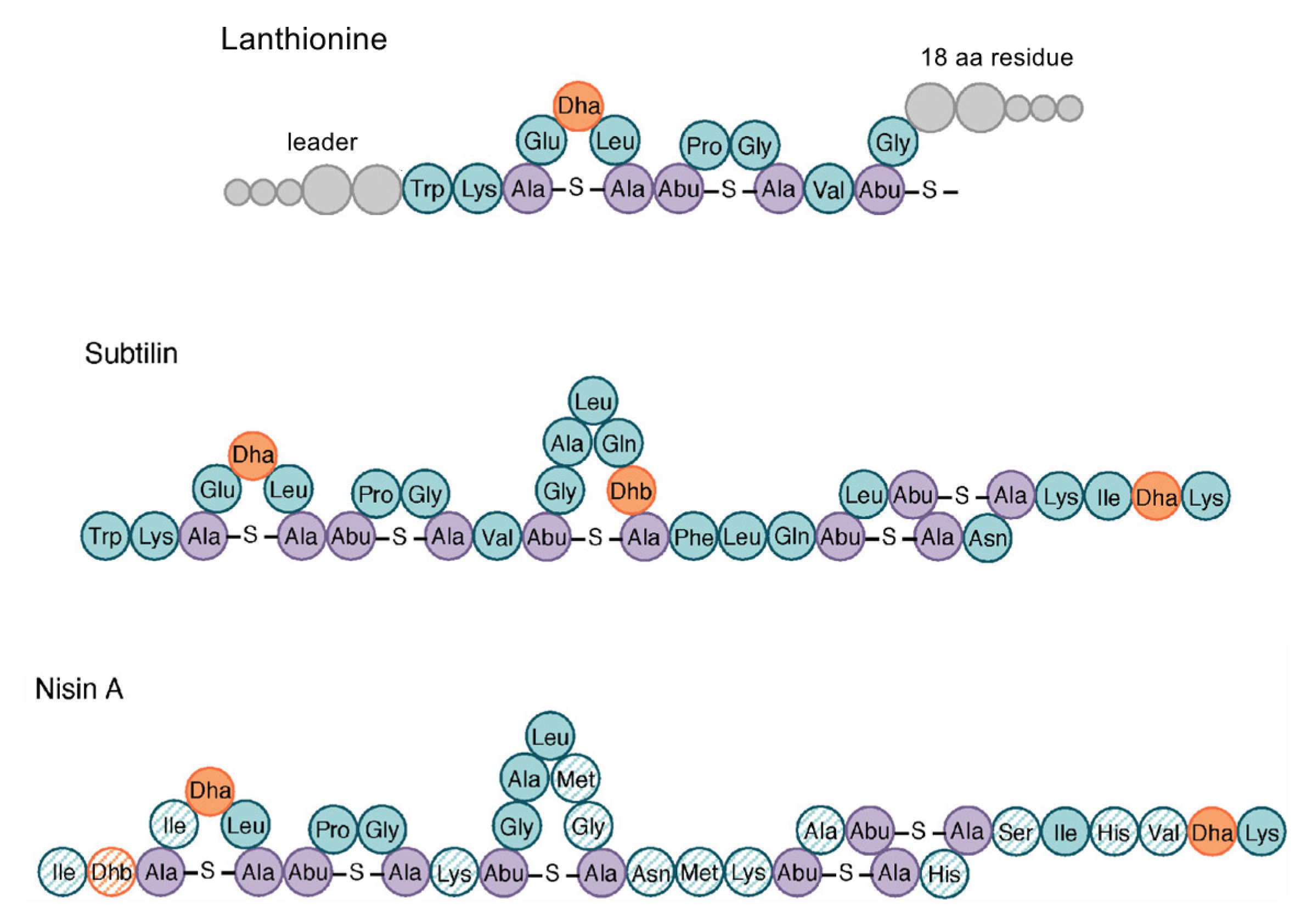
2.4. Bacteriocins
2.5. Volatile Compounds
3. Purification and Identification of Bacillus Metabolites
3.1. Purification and Identification Technology of Bacillus Metabolites
3.2. Combination of Purification and Identification Methods
3.3. Application of Omics and Other Methods in Purification and Identification of Bacillus Metabolites
4. Mechanism of Bacillus Metabolites Inhibiting C. albicans
4.1. Inhibition of Pathogenic Fungi
4.1.1. Damage of Cell Wall and Cell Membrane
4.1.2. Intracellular Damage
4.2. Inhibition of Virulence Factors
4.2.1. Inhibition of Yeast-Hyphae Biphasic Transition
4.2.2. Inhibition of Adhesion
4.2.3. Inhibition of Pathogenic Fungal Invasion
4.2.4. Inhibition of Hydrolase Secretion
4.2.5. Inhibition of Biofilm Formation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kim, J.Y. Human fungal pathogens: Why should we learn? J. Microbiol. 2016, 54, 145–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, M.; Fan, X.; Chen, S.C.; Wang, H.; Sun, Z.Y.; Liao, K.; Chen, S.L.; Yan, Y.; Kang, M.; Hu, Z.D.; et al. Antifungal susceptibilities of Candida glabrata species complex, Candida krusei, Candida parapsilosis species complex and Candida tropicalis causing invasive candidiasis in China: 3 year national surveillance. J. Antimicrob. Chemother. 2015, 70, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Arias, M.J.; Hernández-Chávez, M.J.; García-Carnero, L.C.; Amezcua-Hernández, D.G.; Lozoya-Pérez, N.E.; Estrada-Mata, E.; Martínez-Duncker, I.; Franco, B.; Mora-Montes, H.M. Differential recognition of Candida tropicalis, Candida guilliermondii, Candida krusei, and Candida auris by human innate immune cells. Infect. Drug Resist. 2019, 12, 783–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vila, T.; Romo, J.A.; Pierce, C.G.; Mchardy, S.F.; Saville, S.P.; Lopez-Ribot, J.L. Targeting Candida albicans filamentation for antifungal drug development. Virulence 2017, 8, 150–158. [Google Scholar] [CrossRef] [Green Version]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole antifungal resistance in Candida albicans and emerging non-albicans Candida species. Front. Microbiol. 2016, 7, 2173. [Google Scholar] [CrossRef] [Green Version]
- Lakshminarayanan, R.; Sridhar, R.; Xian, J.L.; Nandhakumar, M.; Barathi, V.A.; Kalaipriya, M.; Jia, L.K.; Liu, S.P.; Beuerman, R.W.; Ramakrishna, S. Interaction of gelatin with polyenes modulates antifungal activity and biocompatibility of electrospun fiber mats. Int. J. Nanomed. 2014, 2014, 2439–2458. [Google Scholar] [CrossRef] [Green Version]
- Perlin, D.S. Mechanisms of echinocandin antifungal drug resistance. Ann. NY Acad. Sci. 2015, 1354, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Muhialdin, B.J.; Hassan, Z.; Bakar, F.A.; Saari, N. Identification of antifungal peptides produced by Lactobacillus plantarum IS10 grown in the MRS broth. Food Control 2016, 59, 27–30. [Google Scholar] [CrossRef]
- Zida, A.; Bamba, S.; Yacouba, A.; Ouedraogo-Traore, R.; Guiguemdé, R.T. Anti-Candida albicans natural products, sources of new antifungal drugs: A review. J. Mycol. Méd. 2017, 27, 1–19. [Google Scholar] [CrossRef]
- Roemer, T.; Krysan, D.J. Antifungal drug development: Challenges, unmet clinical needs, and new approaches. CSH Perspect. Med. 2014, 4, a019703. [Google Scholar] [CrossRef]
- Jasim, B.; Sreelakshmi, K.S.; Mathew, J.; Radhakrishnan, E.K. Surfactin, Iturin, and Fengycin biosynthesis by endophytic Bacillus sp. from Bacopa monnieri. Microb. Ecol. 2016, 72, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the Antimicrobial Compounds Produced by Members of the Bacillus subtilis Group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, W.G.; Parreira, A.; Nascimento, L.A.; Leonel, C.A.; Ferreira, J.M.S. Absence of antibacterial, anti-Candida, and anti-dengue activities of a surfactin isolated from Bacillus subtilis. J. Pharm. Negat. Result. 2018, 9, 27–32. [Google Scholar]
- Lei, S.; Zhao, H.; Pang, B.; Qu, R.; Lian, Z.; Jiang, C.; Shao, D.; Huang, Q.; Jin, M.; Shi, J. Capability of iturin from Bacillus subtilis to inhibit Candida albicans in vitro and in vivo. Appl. Microbiol. Biotechnol. 2019, 103, 4377–4392. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Sen, S.; Bhakat, A.; Bhowmick, A.; Sarkar, K. The lipopeptides fengycin and iturin are involved in the anticandidal activity of endophytic Bacillus sp. as determined by experimental and in silico analysis. Lett. Appl. Microbiol. 2022, 75, 450–459. [Google Scholar] [CrossRef]
- Yuliani, H.; Perdani, M.S.; Savitri, I.; Manurung, M.; Sahlan, M.; Wijanarko, A.; Hermansyah, H. Antimicrobial activity of biosurfactant derived from Bacillus subtilis C19. Energy Procedia 2018, 153, 274–278. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, J.; Sun, J.; Zhu, X.; Zhou, L.; Lu, Z.; Lu, Y. C16-Fengycin A affect the growth of Candida albicans by destroying its cell wall and accumulating reactive oxygen species. Appl. Microbiol. Biotechnol. 2019, 103, 8963–8975. [Google Scholar] [CrossRef]
- Gomaa, E.Z.; El-Mahdy, O.M. Improvement of chitinase production by Bacillus thuringiensis NM101-19 for antifungal biocontrol through physical mutation. Microbiology 2018, 87, 472–485. [Google Scholar] [CrossRef]
- Subramani, A.K.; Raval, R.; Sundareshan, S.; Sivasengh, R.; Raval, K. A marine chitinase from Bacillus aryabhattai with antifungal activity and broad specificity toward crystalline chitin degradation. Prep. Biochem. Biotechnol. 2022, 52, 1160–1172. [Google Scholar] [CrossRef]
- Abassi, S.; Emtiazi, G.; Hosseini-Abari, A.; Kim, B.G. Chitooligosaccharides and thermostable chitinase against vulvovaginal candidiasis and saprophyte fungi: LC mass studies of shrimp shell fermentation by Bacillus altitudinis. Curr. Microbiol. 2020, 77, 40–48. [Google Scholar] [CrossRef]
- Allonsius, C.N.; Vandenheuvel, D.; Oerlemans, E.F.M.; Petrova, M.I.; Donders, G.G.G.; Cos, P.; Delputte, P.; Lebeer, S. Inhibition of Candida albicans morphogenesis by chitinase from Lactobacillus rhamnosus GG. Sci. Rep. 2019, 9, 2900. [Google Scholar] [CrossRef] [PubMed]
- Mayer, F.L.; Kronstad, J.W. Disarming Fungal Pathogens: Bacillus safensis Inhibits virulence factor production and biofilm formation by Cryptococcus neoformans and Candida albicans. Mbio 2017, 8, e01537-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, F.; Ortiz, A.; Sansinenea, E. A strong antifungal activity of 7-O-succinyl macrolactin A vs macrolactin A from Bacillus amyloliquefaciens ELI149. Curr. Microbiol. 2020, 77, 3409–3413. [Google Scholar] [CrossRef] [PubMed]
- Daas, M.S.; Acedo, J.Z.; Rosana, A.R.R.; Orata, F.D.; Reiz, B.; Zheng, J.; Nateche, F.; Case, R.J.; Kebbouche-Gana, S.; Vederas, J.C. Bacillus amyloliquefaciens ssp plantarum F11 isolated from Algerian salty lake as a source of biosurfactants and bioactive lipopeptides. Fems Microbiol. Lett. 2018, 365, fnx248. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.C.P.; Sousa, R.C.S.; Otoni, C.G.; Moraes, A.R.F.; Souza, V.G.L.; Medeiros, E.A.A.; Espitia, P.J.P.; Pires, A.C.S.; Coimbra, J.S.R.; Soares, N.F.F. Nisin and other antimicrobial peptides: Production, mechanisms of action, and application in active food packaging. Innov. Food Sci. Emerg. 2018, 48, 179–194. [Google Scholar] [CrossRef]
- Shayesteh, F.; Ahmad, A.; Usup, G. Purification and partial characterisation of an antifungal bacteriocin from Bacillus sp. Sh10 associated with marine carpet clam (Paphia textile). J. Microb. Biotechnol. Food 2021, 10, e2513. [Google Scholar] [CrossRef]
- Brown, A.O.; Graham, C.E.; Cruz, M.R.; Singh, K.V.; Murray, B.E.; Lorenz, M.C.; Garsin, D.A. Antifungal activity of the Enterococcus faecalis peptide EntV requires protease cleavage and disulfide bond formation. Mbio 2019, 10, e01334-19. [Google Scholar] [CrossRef] [Green Version]
- Calvo, H.; Mendiara, I.; Arias, E.; Gracia, A.P.; Blanco, D.; Venturini, M.E. Antifungal activity of the volatile organic compounds produced by Bacillus velezensis strains against postharvest fungal pathogens. Postharvest Biol. Technol. 2020, 166, 111208. [Google Scholar] [CrossRef]
- Hamiche, S.; Badis, A.; Jouadi, B.; Bouzidi, N.; Daghbouche, Y.; Utczás, M.; Mondello, L.; El Hattab, M. Identification of antimicrobial volatile compounds produced by the marine bacterium Bacillus amyloliquefaciens strain S13 newly isolated from brown alga Zonaria tournefortii. J. Essent. Oil Res. 2019, 31, 203–210. [Google Scholar] [CrossRef]
- Xie, C.L.; Xia, J.M.; Su, R.Q.; Li, J.; Liu, Y.H.; Yang, X.W.; Yang, Q. Bacilsubteramide A, a new indole alkaloid, from the deep-sea-derived Bacillus subterraneus 11593. Nat. Prod. Res. 2018, 32, 2553–2557. [Google Scholar] [CrossRef]
- Ashitha, A.; Radhakrishnan, E.K.; Mathew, J. Characterization of biosurfactant produced by the endophyte Burkholderia sp. WYAT7 and evaluation of its antibacterial and antibiofilm potentials. J. Biotechnol. 2020, 313, 1–10. [Google Scholar]
- Subramenium, G.A.; Swetha, T.K.; Iyer, P.M.; Balamurugan, K.; Pandian, S.K. 5-hydroxymethyl-2-furaldehyde from marine bacterium Bacillus subtilis inhibits biofilm and virulence of Candida albicans. Microbiol. Res. 2018, 207, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Al-Thubiania, A.S.A.; Maher, Y.A.; Fathi, A.; Abourehab, M.A.S.; Alarjah, M.; Khan, M.S.A.; Al-Ghamdi, S.B. Identification and characterization of a novel antimicrobial peptide compound produced by Bacillus megaterium strain isolated from oral microflora. Saudi Pharm. J. 2018, 26, 1089–1097. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Mohammed Breig, S.J.; Gómez, A.; Sánchez-Arévalo, I.; González-Faune, P.; Sarkar, S.; Bandopadhyay, R.; Vuree, S.; Cornejo, J.; Tapia, J.; et al. Optimization and characterization of a novel exopolysaccharide from Bacillus haynesii CamB6 for food applications. Biomolecules 2022, 12, 834. [Google Scholar] [CrossRef] [PubMed]
- Devi, S.; Kiesewalter, H.T.; Kovacs, R.; Frisvad, J.C.; Weber, T.; Larsen, T.O.; Kovacs, A.T.; Ding, L. Depiction of secondary metabolites and antifungal activity of Bacillus velezensis DTU001. Synth. Syst. Biotechnol. 2019, 4, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Du, C.; Gubbens, J.; Choi, Y.H.; Van Wezel, G.P. Metabolomics-driven discovery of a prenylated isatin antibiotic produced by Streptomyces species MBT28. J. Nat. Prod. 2015, 78, 2355–2363. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Tian, X.; Shao, M.; Xie, Y.; Huang, H.; Hu, J.; Ju, J. Genome mining and metabolic profiling illuminate the chemistry driving diverse biological activities of Bacillus siamensis SCSIO 05746. Appl. Microbiol. Biotechnol. 2019, 103, 4153–4165. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, W.; Shi, X.; Zheng, H.; Zheng, Z.; Lu, X.; Xing, Y.; Ji, K.; Liu, M.; Dong, Y. Inducing secondary metabolite production of Aspergillus sydowii through microbial co-culture with Bacillus subtilis. Microb. Cell Factories 2021, 20, 42. [Google Scholar] [CrossRef]
- Swidergall, M.; Ernst, J.F. Interplay between Candida albicans and the antimicrobial peptide armory. Eukaryot. Cell 2014, 13, 950–957. [Google Scholar] [CrossRef] [Green Version]
- Tsai, P.W.; Cheng, Y.L.; Hsieh, W.P.; Lan, C.Y. Responses of Candida albicans to the human antimicrobial peptide LL-37. J. Microbiol. 2014, 52, 581–589. [Google Scholar] [CrossRef]
- Avitabile, C.; D’andrea, L.D.; Saviano, M.; Olivieri, M.; Cimmino, A.; Romanelli, A. Binding studies of antimicrobial peptides to Escherichia coli cells. Biochem. Biophys. Res. Commun. 2016, 478, 149–153. [Google Scholar] [CrossRef] [PubMed]
- De Medeiros, L.N.; Domitrovic, T.; De Andrade, P.C.; Faria, J.; Bergter, E.B.; Weissmuller, G.; Kurtenbach, E. Psd1 binding affinity toward fungal membrane components as assessed by SPR: The role of glucosylceramide in fungal recognition and entry. Biopolymers 2014, 102, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Jakab, A.; Kovacs, F.; Balla, N.; Toth, Z.; Ragyak, A.; Sajtos, Z.; Csillag, K.; Nagy-Koteles, C.; Nemes, D.; Bacskay, I.; et al. Physiological and transcriptional profiling of surfactin exerted antifungal effect against Candida albicans. Biomed. Pharmacother. 2022, 152, 113220. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Zhao, X.; Yang, L.; Su, P.; Fu, P.; Peng, J.; Yang, N.; Guo, G. Antimicrobial peptide AMP-17 affects Candida albicans by disrupting its cell wall and cell membrane integrity. Infect. Drug Resist. 2020, 13, 2509–2520. [Google Scholar] [CrossRef]
- Fischer, D.; Gessner, G.; Fill, T.P.; Barnett, R.; Tron, K.; Dornblut, K.; Kloss, F.; Stallforth, P.; Hube, B.; Heinemann, S.H.; et al. Disruption of membrane integrity by the bacterium-derived antifungal Jagaricin. Antimicrob. Agents Chemother. 2019, 63, e00707-19. [Google Scholar] [CrossRef] [Green Version]
- Vieira, M.E.B.; Gomes, V.M.; Carvalho, A.d.O.; Vasconcelos, I.M.; Machado, O.L.T. Isolation, characterization and mechanism of action of an antimicrobial peptide from Lecythis pisonis seeds with inhibitory activity against Candida albicans. Acta Biochim. Biophys. Sin. 2015, 47, 716–729. [Google Scholar] [CrossRef] [Green Version]
- Pooja; Prasher, P.; Singh, P.; Pawar, K.; Vikramdeo, K.S.; Mondal, N.; Komath, S.S. Synthesis of amino acid appended indoles: Appreciable anti-fungal activity and inhibition of ergosterol biosynthesis as their probable mode of action. Eur. J. Med. Chem. 2014, 80, 325–339. [Google Scholar] [CrossRef]
- Li, L.; Sun, J.; Xia, S.; Tian, X.; Cheserek, M.J.; Le, G. Mechanism of antifungal activity of antimicrobial peptide APP, a cell-penetrating peptide derivative, against Candida albicans: Intracellular DNA binding and cell cycle arrest. Appl. Microbiol. Biotechnol. 2016, 100, 3245–3253. [Google Scholar] [CrossRef]
- Lu, Y.; Su, C.; Liu, H. Candida albicans hyphal initiation and elongation. Trends Microbiol. 2014, 22, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Silva-Dias, A.; Miranda, I.M.; Branco, J.; Monteiro-Soares, M.; Pina-Vaz, C.; Rodrigues, A.G. Adhesion, biofilm formation, cell surface hydrophobicity, and antifungal planktonic susceptibility: Relationship among Candida spp. Front. Microbiol. 2015, 6, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, C.; Yu, J.; Lu, Y. Hyphal development in Candida albicans from different cell states. Curr. Genet. 2018, 64, 1239–1243. [Google Scholar] [CrossRef] [PubMed]
- Childers, D.S.; Kadosh, D. Filament condition-specific response elements control the expression of NRG1 and UME6, key transcriptional regulators of morphology and virulence in Candida albicans. PLoS ONE 2015, 10, e0122775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, M.P.; De Barros, P.P.; Jorjão, A.L.; Rossoni, R.D.; Junqueira, J.C.; Jorge, A.O.C. Effects of Bacillus subtilis on Candida albicans: Biofilm formation, filamentation and gene expression. Brazilian Dental Sci. 2019, 22, 252–259. [Google Scholar] [CrossRef] [Green Version]
- De Barros, P.P.; Scorzoni, L.; Ribeiro, F.d.C.; De Oliveira Fugisaki, L.R.; Fuchs, B.B.; Mylonakis, E.; Cardoso Jorge, A.O.; Junqueira, J.C.; Rossoni, R.D. Lactobacillus paracasei 28.4 reduces in vitro hyphae formation of Candida albicans and prevents the filamentation in an experimental model of Caenorhabditis elegans. Microb. Pathog. 2018, 117, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Martin, H.; Kavanagh, K.; Velasco-Torrijos, T. Targeting adhesion in fungal pathogen Candida albicans. Future Med. Chem. 2021, 13, 313–334. [Google Scholar] [CrossRef]
- Ceresa, C.; Tessarolo, F.; Maniglio, D.; Caola, I.; Nollo, G.; Rinaldi, M.; Fracchia, L. Inhibition of Candida albicans biofilm by lipopeptide AC7 coated medical-grade silicone in combination with farnesol. Aims Bioeng. 2018, 5, 192–208. [Google Scholar] [CrossRef]
- Jemil, N.; Hmidet, N.; Manresa, A.; Rabanal, F.; Nasri, M. Isolation and characterization of kurstakin and surfactin isoforms produced by Enterobacter cloacae C3 strain. J. Mass Spectrom. 2019, 54, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Bucci, A.R.; Marcelino, L.; Mendes, R.K.; Etchegaray, A. The antimicrobial and antiadhesion activities of micellar solutions of surfactin, CTAB and CPCl with terpinen-4-ol: Applications to control oral pathogens. World J. Microb. Biotechnol. 2018, 34, 86. [Google Scholar] [CrossRef]
- Behzadi, P.; Behzadi, E.; Ranjbar, R. Urinary tract infections and Candida albicans. Cent. Eur. J. Urol. 2015, 68, 96–101. [Google Scholar] [CrossRef] [Green Version]
- Rautela, R.; Singh, A.K.; Shukla, A.; Cameotra, S.S. Lipopeptides from Bacillus strain AR2 inhibits biofilm formation by Candida albicans. Antonie Leeuwenhoek 2014, 105, 809–821. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Wang, X.; Zhao, C.; Yan, Z. Immunomodulatory mechanism of Bacillus subtilis R0179 in RAW 264.7 cells against Candida albicans challenge. Microb. Pathog. 2021, 157, 104988. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibition of Pathogenic Fungi | Inhibition of Virulence Factors |
|---|---|
| (1) Damage of cell wall and cell membrane | (1) Inhibition of yeast to hypha biphasic transition |
| (2) Intracellular damage (Inhibition of target cell wall synthesis; Inhibition of cellular respiration and protein synthesis; Binding with fungal DNA) | (2) Inhibition of adhesion |
| (3) Inhibition of pathogenic fungal invasion | |
| (4) Inhibition of hydrolase secretion | |
| (5) Inhibition of biofilm formation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Zhao, J.; Zhang, Z. Bacillus Metabolites: Compounds, Identification and Anti-Candida albicans Mechanisms. Microbiol. Res. 2022, 13, 972-984. https://doi.org/10.3390/microbiolres13040070
Wang W, Zhao J, Zhang Z. Bacillus Metabolites: Compounds, Identification and Anti-Candida albicans Mechanisms. Microbiology Research. 2022; 13(4):972-984. https://doi.org/10.3390/microbiolres13040070
Chicago/Turabian StyleWang, Weichen, Jin Zhao, and Zhizi Zhang. 2022. "Bacillus Metabolites: Compounds, Identification and Anti-Candida albicans Mechanisms" Microbiology Research 13, no. 4: 972-984. https://doi.org/10.3390/microbiolres13040070