
Multidrug Resistance Dissemination in Escherichia coli Isolated from Wild Animals: Bacterial Clones and Plasmid Complicity


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Antibiotic Susceptibility Testing
2.3. Whole-Genome Sequencing
Genomic Data Analysis
2.4. Plasmid Analysis
3. Results
3.1. Resistance Phenotype and Resistome
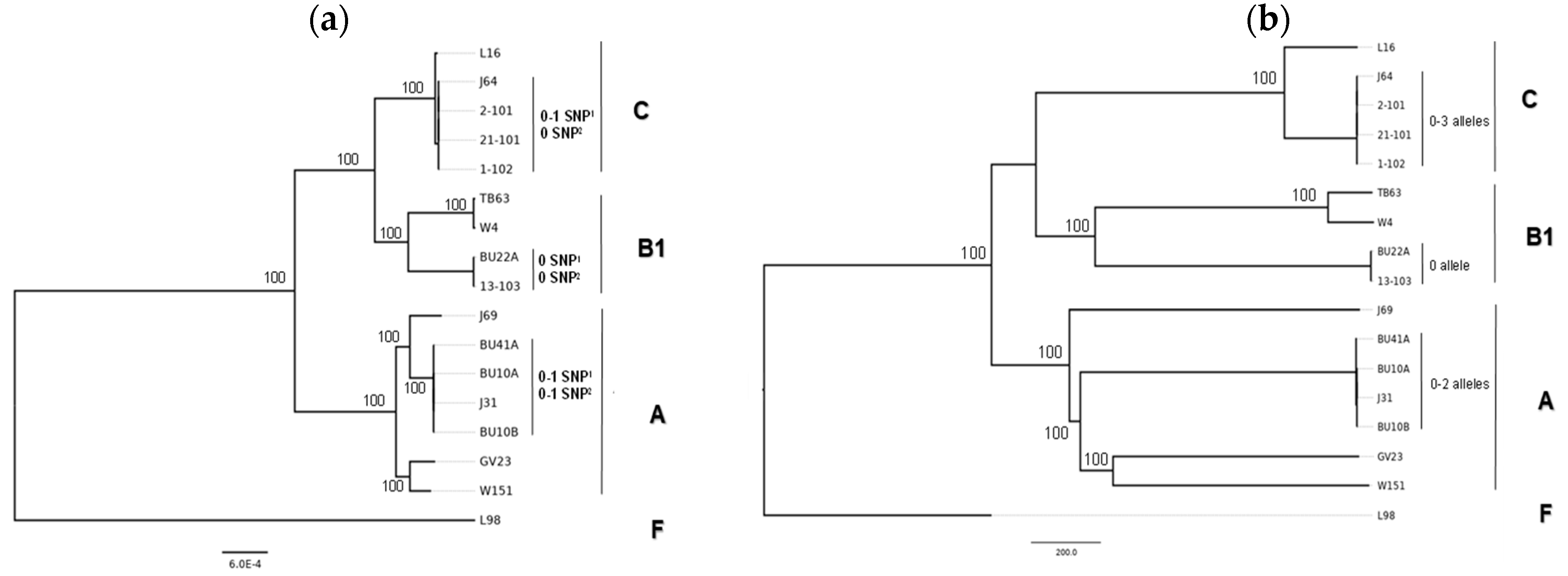
3.2. Whole-Genome Typing of E. coli Isolates
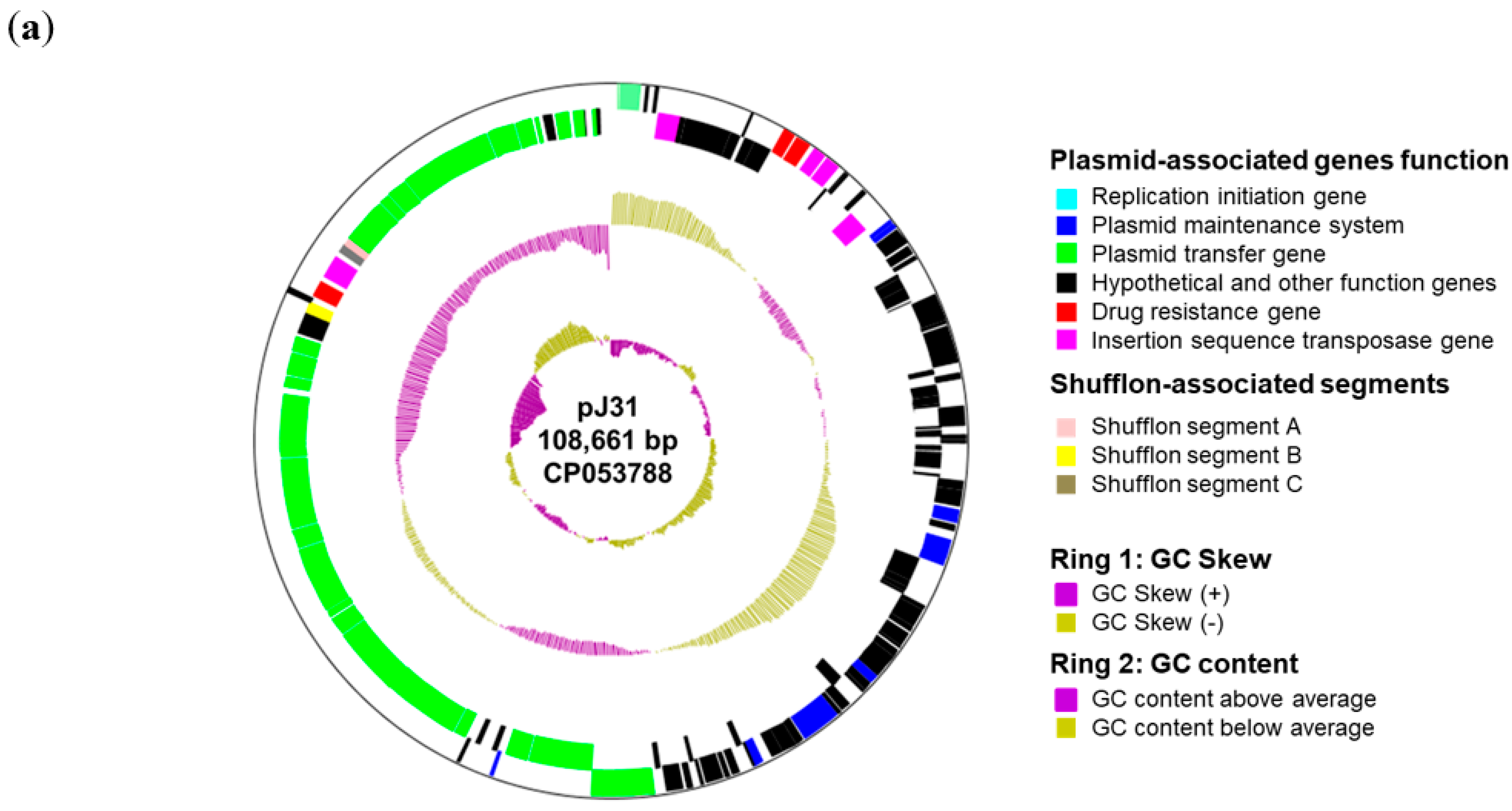
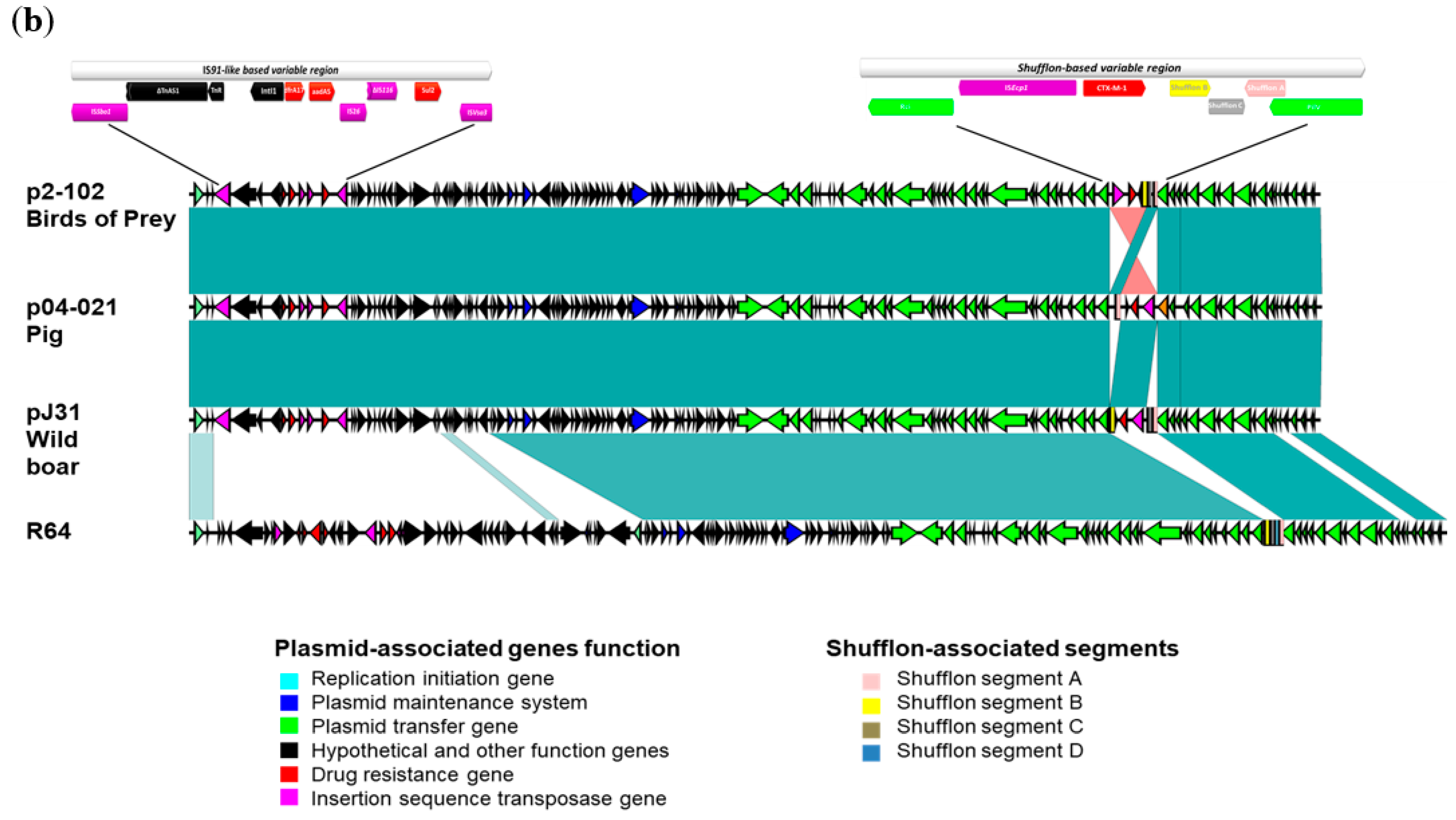
3.3. CTX-M-1-Encoding Plasmids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roca, I.; Akova, M.; Baquero, F.; Carlet, J.; Cavaleri, M.; Coenen, S.; Cohen, J.; Findlay, D.; Gyssens, I.; Heure, O.E.; et al. The global threat of antimicrobial resistance: Science for intervention. New Microbes New Infect. 2015, 6, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. WHO List of Critically Important Antimicrobials for Human Medicine (WHO CIA List); 5th revision; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martínez, J.L. Defining and combating antibiotic resistance from One Health and Global Health perspectives. Nat. Microbiol. 2019, 4, 1432–1442. [Google Scholar] [CrossRef]
- Huijbers, P.M.C.; Graat, E.A.M.; Haenen, A.P.J.; van Santen, M.G.; van Essen-Zandbergen, A.; Mevius, D.J.; van Duijkeren, E.; van Hoek, A.H.A.M. Extended-spectrum and AmpC β-lactamase-producing Escherichia coli in broilers and people living and/or working on broiler farms: Prevalence, risk factors and molecular characteristics. J. Antimicrob. Chemother. 2014, 69, 2669–2675. [Google Scholar] [CrossRef] [PubMed]
- Sato, G.; Oka, C.; Asagi, M.; Ishiguro, N. Detection of conjugative R plasmids conferring chloramphenicol resistance in Escherichia coli isolated from domestic and feral pigeons and crows. Zent. Bakteriol. Parasitenkd. Infekt. Hyg. 1978, 241, 407–417. [Google Scholar] [PubMed]
- Wang, J.; Ma, Z.B.; Zeng, Z.L.; Yang, X.W.; Huang, Y.; Liu, J.H. The role of wildlife (wild birds) in the global transmission of antimicrobial resistance genes. Zool. Res. 2017, 38, 55–80. [Google Scholar] [CrossRef] [Green Version]
- Costa, D.; Poeta, P.; Sáenz, Y.; Vinué, L.; Rojo-Bezares, B.; Jouini, A.; Zarazaga, M.; Rodrigues, J.; Torres, C. Detection of Escherichia coli harbouring extended-spectrum β-lactamases of the CTX-M, TEM and SHV classes in faecal samples of wild animals in Portugal. J. Antimicrob. Chemother. 2006, 58, 1311–1312. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Potron, A.; De La Cuesta, C.; Cleary, T.; Nordmann, P.; Munoz-Price, L.S. Wild coastline birds as reservoirs of broad-spectrum-β-lactamase- producing Enterobacteriaceae in Miami Beach, Florida. Antimicrob. Agents Chemother. 2012, 56, 2756–2758. [Google Scholar] [CrossRef] [Green Version]
- Stedt, J.; Bonnedahl, J.; Hernandez, J.; Waldenström, J.; McMahon, B.J.; Tolf, C.; Olsen, B.; Drobni, M. Carriage of CTX-M type extended spectrum β-lactamases (ESBLs) in gulls across Europe. Acta Vet. Scand. 2015, 57, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Dolejska, M.; Papagiannitsis, C.C. Plasmid-mediated resistance is going wild. Plasmid 2018, 99, 99–111. [Google Scholar] [CrossRef]
- Greig, J.; Rajić, A.; Young, I.; Mascarenhas, M.; Waddell, L.; Lejeune, J. A scoping review of the role of wildlife in the transmission of bacterial pathogens and antimicrobial resistance to the food chain. Zoonoses Public Health 2015, 62, 269–284. [Google Scholar] [CrossRef]
- Haenni, M.; Beyrouthy, R.; Lupo, A.; Châtre, P.; Madec, J.Y.; Bonnet, R. Epidemic spread of Escherichia coli ST744 isolates carrying mcr-3 and blaCTX-M-55 in cattle in France. J. Antimicrob. Chemother. 2018, 73, 533–536. [Google Scholar] [CrossRef]
- Robin, F.; Beyrouthy, R.; Bonacorsi, S.; Aissa, N.; Bret, L.; Brieu, N.; Cattoir, V.; Chapuis, A.; Chardon, H.; Degand, N.; et al. Inventory of Extended-Spectrum-B- Lactamase-Producing Enterobacteriaceae in France as Assessed by a Multicenter Study. Antimicrob. Agents Chemother. 2017, 61, e01911-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellington, E.M.H.; Boxall, A.B.A.; Cross, P.; Feil, E.J.; Gaze, W.H.; Hawkey, P.M.; Johnson-Rollings, A.S.; Jones, D.L.; Lee, N.M.; Otten, W.; et al. The role of the natural environment in the emergence of antibiotic resistance in Gram-negative bacteria. Lancet Infect. Dis. 2013, 13, 155–165. [Google Scholar] [CrossRef]
- Pietsch, M.; Eller, C.; Wendt, C.; Holfelder, M.; Falgenhauer, L.; Fruth, A.; Grössl, T.; Leistner, R.; Valenza, G.; Werner, G.; et al. Molecular characterisation of extended-spectrum β-lactamase (ESBL)-producing Escherichia coli isolates from hospital and ambulatory patients in Germany. Vet. Microbiol. 2017, 200, 130–137. [Google Scholar] [CrossRef]
- Liu, X.; Thungrat, K.; Boothe, D.M. Occurrence of oxa-48 carbapenemase and other β-lactamase genes in esbl-producing multidrug resistant: Escherichia coli from dogs and cats in the united states, 2009–2013. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Madec, J.Y.; Haenni, M.; Métayer, V.; Saras, E.; Nicolas-Chanoine, M.H. High prevalence of the animal-associated blaCTX-M-1 IncI1/ST3 plasmid in human Escherichia coli isolates. Antimicrob. Agents Chemother. 2015, 59, 5860–5861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsen, L.; Bortolaia, V.; Bielak, E.; Moodley, A.; Olsen, S.S.; Hansen, D.S.; Frimodt-Møller, N.; Guardabassi, L.; Hasman, H. Limited similarity between plasmids encoding CTX-M-1 β-lactamase in Escherichia coli from humans, pigs, cattle, organic poultry layers and horses in Denmark. J. Glob. Antimicrob. Resist. 2015, 3, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A. Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [Green Version]
- Literak, I.; Dolejska, M.; Janoszowska, D.; Hrusakova, J.; Meissner, W.; Rzyska, H.; Bzoma, S.; Cizek, A. Antibiotic-resistant Escherichia coli bacteria, including strains with genes encoding the extended-spectrum beta-lactamase and QnrS, in waterbirds on the baltic sea coast of Poland. Appl. Environ. Microbiol. 2010, 76, 8126–8134. [Google Scholar] [CrossRef] [Green Version]
- Veldman, K.; van Tulden, P.; Kant, A.; Testerink, J.; Mevius, D. Characteristics of cefotaxime-resistant Escherichia coli from wild birds in the Netherlands. Appl. Environ. Microbiol. 2013, 79, 7556–7561. [Google Scholar] [CrossRef] [Green Version]
- Schaufler, K.; Nowak, K.; Düx, A.; Semmler, T.; Villa, L.; Kourouma, L.; Bangoura, K.; Wieler, L.H.; Leendertz, F.H.; Guenther, S. Clinically relevant ESBL-producing K. pneumoniae ST307 and E. coli ST38 in an urban West African rat population. Front. Microbiol. 2018, 9, 150. [Google Scholar] [CrossRef] [PubMed]
- Guyomard-Rabenirina, S.; Reynaud, Y.; Pot, M.; Albina, E.; Couvin, D.; Ducat, C.; Gruel, G.; Ferdinand, S.; Legreneur, P.; Le Hello, S.; et al. Antimicrobial Resistance in Wildlife in Guadeloupe (French West Indies): Distribution of a Single blaCTX–M–1/IncI1/ST3 Plasmid Among Humans and Wild Animals. Front. Microbiol. 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Poeta, P.; Radhouani, H.; Igrejas, G.; Gonçalves, A.; Carvalho, C.; Rodrigues, J.; Vinué, L.; Somalo, S.; Torres, C. Seagulls of the Berlengas natural reserve of Portugal as carriers of fecal Escherichia coli harboring CTX-M and TEM extended-spectrum beta-lactamases. Appl. Environ. Microbiol. 2008, 74, 7439–7441. [Google Scholar] [CrossRef] [Green Version]
- Radhouani, H.; Pinto, L.; Coelho, C.; Gonçalves, A.; Sargo, R.; Torres, C.; Igrejas, G.; Poeta, P. Detection of Escherichia coli harbouring extended-spectrum β-lactamases of the CTX-M classes in faecal samples of common buzzards (Buteo buteo). J. Antimicrob. Chemother. 2010, 65, 171–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, A.; Igrejas, G.; Radhouani, H.; Estepa, V.; Alcaide, E.; Zorrilla, I.; Serra, R.; Torres, C.; Poeta, P. Detection of extended-spectrum beta-lactamase-producing Escherichia coli isolates in faecal samples of Iberian lynx. Lett. Appl. Microbiol. 2012, 54, 73–77. [Google Scholar] [CrossRef]
- Pinto, L.; Radhouani, H.; Coelho, C.; Da Costa, P.M.; Simões, R.; Brandão, R.M.L.; Torres, C.; Igrejas, G.; Poeta, P. Genetic detection of extended-spectrum β-lactamase-containing Escherichia coli isolates from birds of prey from Serra da Estrela natural reserve in Portugal. Appl. Environ. Microbiol. 2010, 76, 4118–4120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, A.; Igrejas, G.; Radhouani, H.; Estepa, V.; Pacheco, R.; Monteiro, R.; Brito, F.; Guerra, A.; Petrucci-Fonseca, F.; Torres, C.; et al. Iberian Wolf as a Reservoir of Extended-Spectrum β-Lactamase-Producing Escherichia coli of the TEM, SHV, and CTX-M Groups. Microb. Drug Resist. 2012, 18, 215–219. [Google Scholar] [CrossRef]
- Poeta, P.; Radhouani, H.; Pinto, L.; Martinho, A.; Rego, V.; Rodrigues, R.; Gonçalves, A.; Rodrigues, J.; Estepa, V.; Torres, C.; et al. Wild boars as reservoirs of extended-spectrum beta-lactamase (ESBL) producing Escherichia coli of different phylogenetic groups. J. Basic Microbiol. 2009, 49, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Garcês, A.; Correia, S.; Amorim, F.; Pereira, J.E.; Igrejas, G.; Poeta, P. First report on extended-spectrum beta-lactamase (ESBL) producing Escherichia coli from European free-tailed bats (Tadarida teniotis) in Portugal: A one-health approach of a hidden contamination problem. J. Hazard. Mater. 2017, 370, 219–224. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.A.; Korobeynikov, A.; Lapidus, A.; Prjibelski, A.D.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling single-cell genomes and mini-metagenomes from chimeric MDA products. J. Comput. Biol. J. Comput. Mol. Cell Biol. 2013, 20, 714–737. [Google Scholar] [CrossRef] [Green Version]
- Beghain, J.; Bridier-Nahmias, A.; Le Nagard, H.; Denamur, E.; Clermont, O. ClermonTyping: An easy-to-use and accurate in silico method for Escherichia genus strain phylotyping. Microb. Genom. 2018, 4. [Google Scholar] [CrossRef] [PubMed]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.J.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Totsika, M.; Beatson, S.A.; Sarkar, S.; Phan, M.D.; Petty, N.K.; Bachmann, N.; Szubert, M.; Sidjabat, H.E.; Paterson, D.L.; Upton, M.; et al. Insights into a multidrug resistant Escherichia coli pathogen of the globally disseminated ST131 lineage: Genome analysis and virulence mechanisms. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, M.D.; Peters, K.M.; Sarkar, S.; Lukowski, S.W.; Allsopp, L.P.; Moriel, D.G.; Achard, M.E.S.; Totsika, M.; Marshall, V.M.; Upton, M.; et al. The Serum Resistome of a Globally Disseminated Multidrug Resistant Uropathogenic Escherichia coli Clone. PLoS Genet. 2013, 9. [Google Scholar] [CrossRef] [Green Version]
- Ben Zakour, N.L.; Alsheikh-Hussain, A.S.; Ashcroft, M.M.; Khanh Nhu, N.T.; Roberts, L.W.; Stanton-Cook, M.; Schembri, M.A.; Beatsona, S.A. Sequential acquisition of virulence and fluoroquinolone resistance has shaped the evolution of Escherichia coli ST131. mBio 2016, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croucher, N.J.; Page, A.J.; Connor, T.R.; Delaney, A.J.; Keane, J.A.; Bentley, S.D.; Parkhill, J.; Harris, S.R. Rapid phylogenetic analysis of large samples of recombinant bacterial whole genome sequences using Gubbins. Nucleic Acids Res. 2015, 43, e15. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bionformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Machado, M.P.; Silva, D.N.; Rossi, M.; Moran-Gilad, J.; Santos, S.; Ramirez, M.; Carriço, J.A. chewBBACA: A complete suite for gene-by-gene schema creation and strain identification. Microb. Genom. 2018, 4, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017, 45, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Beyrouthy, R.; Barets, M.; Marion, E.; Dananché, C.; Dauwalder, O.; Robin, F.; Gauthier, L.; Jousset, A.; Dortet, L.; Guérin, F.; et al. Novel enterobacter lineage as leading cause of nosocomial outbreak involving carbapenemase-producing strains. Emerg. Infect. Dis. 2018, 24, 1505–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kado, C.I.; Liu, S.T. Rapid procedure for detection and isolation of large and small plasmids. J. Bacteriol. 1981, 145, 1365–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyrouthy, R.; Robin, F.; Fouad, D.; Mallat, H.; Hamze, M.; Bonnet, R. Carbapenemase and virulence factors of Enterobacteriaceae in North Lebanon between 2008 and 2012: Evolution via endemic spread of OXA-48. J. Antimicrob. Chemother. 2014, 69, 2699–2705. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; García-fernández, A.; Larsen, V.; Lund, O.; Villa, L.; Aarestrup, M.; Hasman, H. In Silico Detection and Typing of Plasmids using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.F.; Ali, M.M.M.; Mohamed, Z.K.; Moussa, T.A.; Klena, J.D. Fecal carriage of extended-spectrum β-lactamases and AmpC-producing Escherichia coli in a Libyan community. Ann. Clin. Microbiol. Antimicrob. 2014, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nicolas-chanoine, M.H.; Gruson, C.; Bialek-Davenet, S.; Bertrand, X.; Thomas-Jean, F.; Bert, F.; Moyat, M.; Meiller, E.; Marcon, E.; Danchin, N.; et al. 10-fold increase (2006-11) in the rate of healthy subjects with extended-spectrum β-lactamase-producing Escherichia coli faecal carriage in a parisian check-up centre. J. Antimicrob. Chemother. 2013, 68, 562–568. [Google Scholar] [CrossRef]
- Aibinu, I.; Odugbemi, T.; Koenig, W.; Ghebremedhin, B. Sequence Type ST131 and ST10 Complex (ST617) predominant among CTX-M-15-producing Escherichia coli isolates from Nigeria. Clin. Microbiol. Infect. 2012, 18, E49–E51. [Google Scholar] [CrossRef] [Green Version]
- Mshana, S.E.; Imirzalioglu, C.; Hain, T.; Domann, E.; Lyamuya, E.F.; Chakraborty, T. Multiple ST clonal complexes, with a predominance of ST131, of Escherichia coli harbouring blaCTX-M-15 in a tertiary hospital in Tanzania. Clin. Microbiol. Infect. 2011, 17, 1279–1282. [Google Scholar] [CrossRef] [Green Version]
- Van Der Bij, A.K.; Peirano, G.; Goessens, W.H.F.; Van Der Vorm, E.R.; Van Westreenen, M.; Pitout, J.D.D. Clinical and molecular characteristics of extended-spectrum-β- lactamase-producing Escherichia coli causing bacteremia in the Rotterdam Area, Netherlands. Antimicrob. Agents Chemother. 2011, 55, 3576–3578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correia, S.; Pacheco, R.; Radhouani, H.; Diniz, J.C.; Ponce, P.; Jones-Dias, D.; Caniça, M.; Igrejas, G.; Poeta, P. High prevalence of ESBL-producing Escherichia coli isolates among hemodialysis patients in Portugal: Appearance of ST410 with the blaCTX-M-14 gene. Diagn. Microbiol. Infect. Dis. 2012, 74, 423–425. [Google Scholar] [CrossRef]
- Horner, C.; Fawley, W.; Morris, K.; Parnell, P.; Denton, M.; Wilcox, M. Escherichia coli bacteraemia: 2 years of prospective regional surveillance (2010-12). J. Antimicrob. Chemother. 2014, 69, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Day, M.J.; Mafura, M.T.; Nunez-Garcia, J.; Fenner, J.J.; Sharma, M.; van Essen-Zandbergen, A.; Rodríguez, I.; Dierikx, C.; Kadlec, K.; et al. Comparative Analysis of ESBL-Positive Escherichia coli Isolates from Animals and Humans from the UK, The Netherlands and Germany. PLoS ONE 2013, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Coque, T.M.; Novais, A.; Carattoli, A.; Poirel, L.; Pitout, J.; Peixe, L.; Baquero, F.; Canton, R.; Nordmann, P. Dissemination of clonally related E.coli strains expressing ESBL CTX-M-15. Emerg. Infect. Dis. 2008, 14, 195–200. [Google Scholar] [CrossRef]
- Doi, Y.; Park, Y.S.; Rivera, J.I.; Adams-Haduch, J.M.; Hingwe, A.; Sordillo, E.M.; Lewis, J.S.; Howard, W.J.; Johnson, L.E.; Polsky, B.; et al. Community-associated extended-spectrum -lactamase-producing Escherichia coli infection in the United States. Clin. Infect. Dis. 2013, 56, 641–648. [Google Scholar] [CrossRef]
- Cao, X.; Cavaco, L.M.; Lv, Y.; Li, Y.; Zheng, B.; Wang, P.; Hasman, H.; Liu, Y.; Aarestrup, F.M. Molecular characterization and antimicrobial susceptibility testing of Escherichia coli isolates from patients with urinary tract infections in 20 Chinese Hospitals. J. Clin. Microbiol. 2011, 49, 2496–2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, Y.; Yamamoto, M.; Nagao, M.; Hotta, G.; Matsushima, A.; Ito, Y.; Takakura, S.; Ichiyama, S. Emergence and spread of B2-st131-o25b, B2-st131-o16 and d-st405 clonal groups among extended-spectrum-β-lactamase-producing escherichia coli in Japan. J. Antimicrob. Chemother. 2012, 67, 2612–2620. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, C.; Machado, E.; Fernandes, S.; Peixe, L.; Novais, Â. An update on faecal carriage of ESBL-producing Enterobacteriaceae by Portuguese healthy humans: Detection of the H30 subclone of B2-ST131 Escherichia coli producing CTX-M-27. J. Antimicrob. Chemother. 2016, 71, 1120–1122. [Google Scholar] [CrossRef] [Green Version]
- Blanco, J.; Mora, A.; Mamani, R.; López, C.; Blanco, M.; Dahbi, G.; Herrera, A.; Blanco, J.E.; Alonso, M.P.; García-Garrote, F.; et al. National survey of Escherichia coli causing extraintestinal infections reveals the spread of drug-resistant clonal groups O25b:H4-B2-ST131, O15:H1-D-ST393 and CGA-D-ST69 with high virulence gene content in Spain. J. Antimicrob. Chemother. 2011, 66, 2011–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojer-Usoz, E.; González, D.; Vitas, A.I. Clonal diversity of ESBL-producing Escherichia coli isolated from environmental, human and food samples. Int. J. Environ. Res. Public Health 2017, 14, 10–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, R.; Johnston, B.; Lohse, C.; Chattopadhyay, S.; Tchesnokova, V.; Sokurenko, E.V.; Johnson, J.R. The clonal distribution and diversity of extraintestinal Escherichia coli isolates vary according to patient characteristics. Antimicrob. Agents Chemother. 2013, 57, 5912–5917. [Google Scholar] [CrossRef] [Green Version]
- Giufrè, M.; Graziani, C.; Accogli, M.; Luzzi, I.; Busani, L.; Cerquetti, M. Escherichia coli of human and avian origin: Detection of clonal groups associated with fluoroquinolone and multidrug resistance in Italy. J. Antimicrob. Chemother. 2012, 67, 860–867. [Google Scholar] [CrossRef] [Green Version]
- Ishiwa, A.; Komano, T. Thin pilus PilV adhesins of plasmid R64 recognize specific structures of the lipopolysaccharide molecules of recipient cells. J. Bacteriol. 2003, 185, 5192–5199. [Google Scholar] [CrossRef] [Green Version]
- Ishiwa, A.; Komano, T. PilV adhesins of plasmid R64 thin pili specifically bind to the lipopolysaccharides of recipient cells. J. Mol. Biol. 2004, 343, 615–625. [Google Scholar] [CrossRef]
- Abraham, S.; Kirkwood, R.N.; Laird, T.; Saputra, S.; Mitchell, T.; Singh, M.; Linn, B.; Abraham, R.J.; Pang, S.; Gordon, D.M.; et al. Dissemination and persistence of extended-spectrum cephalosporin-resistance encoding IncI1-bla CTXM-1 plasmid among Escherichia coli in pigs. ISME J. 2018, 12, 2352–2362. [Google Scholar] [CrossRef] [Green Version]
- Valcek, A.; Roer, L.; Overballe-Petersen, S.; Hansen, F.; Bortolaia, V.; Leekitcharoenphon, P.; Korsgaard, H.B.; Seyfarth, A.M.; Hendriksen, R.S.; Hasman, H.; et al. IncI1 ST3 and IncI1 ST7 plasmids from CTX-M-1-producing Escherichia coli obtained from patients with bloodstream infections are closely related to plasmids from E. coli of animal origin. J. Antimicrob. Chemother. 2019, 74, 2171–2175. [Google Scholar] [CrossRef]
- Brouwer, M.S.M.; Tagg, K.A.; Mevius, D.J.; Iredell, J.R.; Bossers, A.; Smith, H.E.; Partridge, S.R. IncI shufflons: Assembly issues in the next-generation sequencing era. Plasmid 2015, 80, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Lovari, S.; Franco, A.; Cordaro, G.; Di Matteo, P.; Battisti, A. Extended-spectrum β-lactamases in Escherichia coli isolated from dogs and cats in Rome, Italy, from 2001 to 2003. Antimicrob. Agents Chemother. 2005, 49, 833–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smet, A.; Martel, A.; Persoons, D.; Dewulf, J.; Heyndrickx, M.; Herman, L.; Haesebrouck, F.; Butaye, P. Broad-spectrum β-lactamases among Enterobacteriaceae of animal origin: Molecular aspects, mobility and impact on public health. FEMS Microbiol. Rev. 2010, 34, 295–316. [Google Scholar] [CrossRef] [Green Version]
- Hordijk, J.; Schoormans, A.; Kwakernaak, M.; Duim, B.; Broens, E.; Dierikx, C.; Mevius, D.; Wagenaar, J.A. High prevalence of fecal carriage of extended-spectrum β-lactamase/AmpC-producing Enterobacteriaceae in cats and dogs. Front. Microbiol. 2013, 4, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, J.; Mora, A.; Mamani, R.; López, C.; Blanco, M.; Dahbi, G.; Herrera, A.; Marzoa, J.; Fernández, V.; De La Cruz, F.; et al. Four main virotypes among extended-spectrum-β-lactamase-producing isolates of Escherichia coli O25b:H4-B2-ST131: Bacterial, epidemiological, and clinical characteristics. J. Clin. Microbiol. 2013, 51, 3358–3367. [Google Scholar] [CrossRef] [Green Version]
- Bachiri, T.; Bakour, S.; Ladjouzi, R.; Thongpan, L.; Rolain, J.M.; Touati, A. High rates of CTX-M-15-producing Escherichia coli and Klebsiella pneumoniae in wild boars and Barbary macaques in Algeria. J. Glob. Antimicrob. Resist. 2017. [Google Scholar] [CrossRef] [PubMed]
- Bonnedahl, J.; Järhult, J.D. Antibiotic resistance in wild birds. Upsala J. Med. Sci. 2014, 119, 113–116. [Google Scholar] [CrossRef] [Green Version]
- Peirano, G.; Pitout, J.D.D. Molecular epidemiology of Escherichia coli producing CTX-M β-lactamases: The worldwide emergence of clone ST131 O25:H4. Int. J. Antimicrob. Agents 2010, 35, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Naseer, U.; Sundsfjord, A. The CTX-M Conundrum: Dissemination of Plasmids and Escherichia coli Clones. Microb. Drug Resist. 2011, 17, 83–97. [Google Scholar] [CrossRef]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Zurfluh, K.; Jakobi, G.; Stephan, R.; Hächler, H.; Nüesch-Inderbinen, M. Replicon typing of plasmids carrying bla CTX-M-1 in Enterobacteriaceae of animal, environmental and human origin. Front. Microbiol. 2014, 5, 555. [Google Scholar] [CrossRef]
- Carattoli, A.; Villa, L.; Fortini, D.; García-Fernández, A. Contemporary IncI1 plasmids involved in the transmission and spread of antimicrobial resistance in Enterobacteriaceae. Plasmid 2018. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Strain ID | Source | Country/Year | Phylogroup/ST | ESBL Gene/Replicon a | Plasmid-Mediated Resistance Genes b | Chromosome-Mediated Resistance Genes b |
|---|---|---|---|---|---|---|
| TB63 | Bat | Portugal/2014 | B1/ST155 | blaCTX-M-1/IncI1-ST3 | aadA5, dfrA17, sul2 | gyrA-S83L |
| 1-102 | Bird of prey | Portugal/2008 | C/ST1998 | blaCTX-M-1/IncI1-ST3 | blaTEM-1, aadA5, dfrA17, sul2, tet(A), tet(R), | gyrA-S83L-D87N, parC-S80I |
| 13-103 | Bird of prey | Portugal/2008 | B1/ST1800 | blaCTX-M-1/IncI1-ST3 | blaTEM-1, aadA5, dfrA17, sul2, tet(A), tet(R) | gyrA-S83L-D87N, parC-S80I |
| 2-101 | Bird of prey | Portugal/2008 | C/ST1998 | blaCTX-M-1/IncI1-ST3 | blaTEM-1, aadA5, dfrA17, sul2, tet(A), tet(R), | gyrA-S83L-D87N, parC-S80I |
| 21-101 | Bird of prey | Portugal/2008 | C/ST1998 | blaCTX-M-1/IncI1-ST3 | blaTEM-1, aadA5, dfrA17, sul2, tet(A), tet(R) | gyrA-S83L-D87N, parC-S80I |
| BU10A | Buzzard | Portugal/2007 | A/ST48 | blaCTX-M-1/IncI1-ST3 | blaTEM-1, aadA5, dfrA17, sul2, tet(A), tet(R), qnrS1, strAB, aadA1, dfrA1, sul1 | |
| BU10B | Buzzard | Portugal/2007 | A/ST48 | blaCTX-M-1/IncI1-ST3 | blaTEM-1, aadA5, dfrA17, sul2, tet(A), tet(R), qnrS1, strAB, aadA1, dfrA1, sul1 | |
| BU22A | Buzzard | Portugal/2007 | B1/ST1800 | blaCTX-M-1/IncI1-ST3 | blaTEM-1, aadA5, dfrA17, sul2, tet(A), tet(R) | gyrA-S83L-D87N, parC-S80I |
| BU41A | Buzzard | Portugal/2007 | A/ST48 | blaCTX-M-1/IncI1-ST3 | blaTEM-1, aadA5, dfrA17, sul2, tet(A), tet(R), qnrS1, strAB, aadA1, dfrA1, sul1 | |
| L16 | Lynx | Spain/2010 | C/ST23 | blaCTX-M-14/IncK | aadA1, aac(3)-VIa, tet(A), tet(R), sul1 | |
| L98 | Lynx | Spain/2010 | F/ST117 | blaSHV-12/IncI1-ST3 | aadA1, aadA2, cmlA1, sul3, strA, dfrA5 | gyrA-S83L |
| GV23 | Seagull | Portugal/2007 | A/ST710 | blaCTX-M-1/IncN-ST1 | blaOXA-1, aadA1, catA1, sul1, tet(B), tet(R) | |
| J31 | Wild boar | Portugal/2006 | A/ST48 | blaCTX-M-1/IncI1-ST3 | blaTEM-1, aadA5, dfrA17, sul2, tet(A), tet(R), qnrS1, strAB, aadA1, dfrA1, sul1 | |
| J64 | Wild boar | Portugal/2006 | C/ST1998 | blaCTX-M-1/IncI1-ST3 | blaTEM-1, aadA5, dfrA17, sul2, tet(A), tet(R) | gyrA-S83L-D87N, parC-S80I |
| J69 | Wild boar | Portugal/2006 | A/ST10430 | blaCTX-M-1/IncI1-ST3 | aadA5, dfrA17, sul2 | gyrA-S83L, parC-S80R, tet(B), tet(R) |
| W151 | Wolf | Portugal/2008 | A/ST361 | blaCTX-M-32/IncN-ST1 | ||
| W4 | Wolf | Portugal/2008 | B1/ST155 | blaCTX-M-1/IncI1-ST3 | tet(A), tet(R) | gyrA-S83L |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabença, C.; Igrejas, G.; Poeta, P.; Robin, F.; Bonnet, R.; Beyrouthy, R. Multidrug Resistance Dissemination in Escherichia coli Isolated from Wild Animals: Bacterial Clones and Plasmid Complicity. Microbiol. Res. 2021, 12, 123-137. https://doi.org/10.3390/microbiolres12010009
Sabença C, Igrejas G, Poeta P, Robin F, Bonnet R, Beyrouthy R. Multidrug Resistance Dissemination in Escherichia coli Isolated from Wild Animals: Bacterial Clones and Plasmid Complicity. Microbiology Research. 2021; 12(1):123-137. https://doi.org/10.3390/microbiolres12010009
Chicago/Turabian StyleSabença, Carolina, Gilberto Igrejas, Patrícia Poeta, Frédéric Robin, Richard Bonnet, and Racha Beyrouthy. 2021. "Multidrug Resistance Dissemination in Escherichia coli Isolated from Wild Animals: Bacterial Clones and Plasmid Complicity" Microbiology Research 12, no. 1: 123-137. https://doi.org/10.3390/microbiolres12010009