Clinical Implications of 20-Hydroxyeicosatetraenoic Acid in the Kidney, Liver, Lung and Brain: An Emerging Therapeutic Target

Abstract
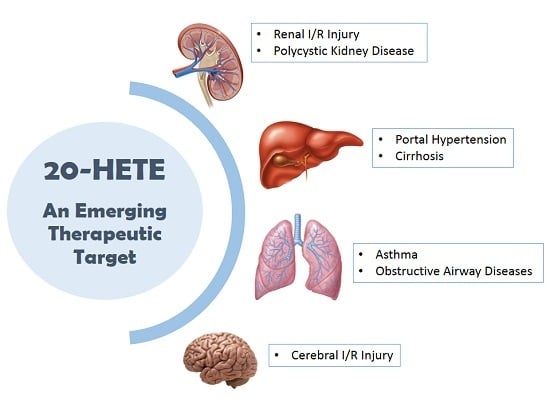
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
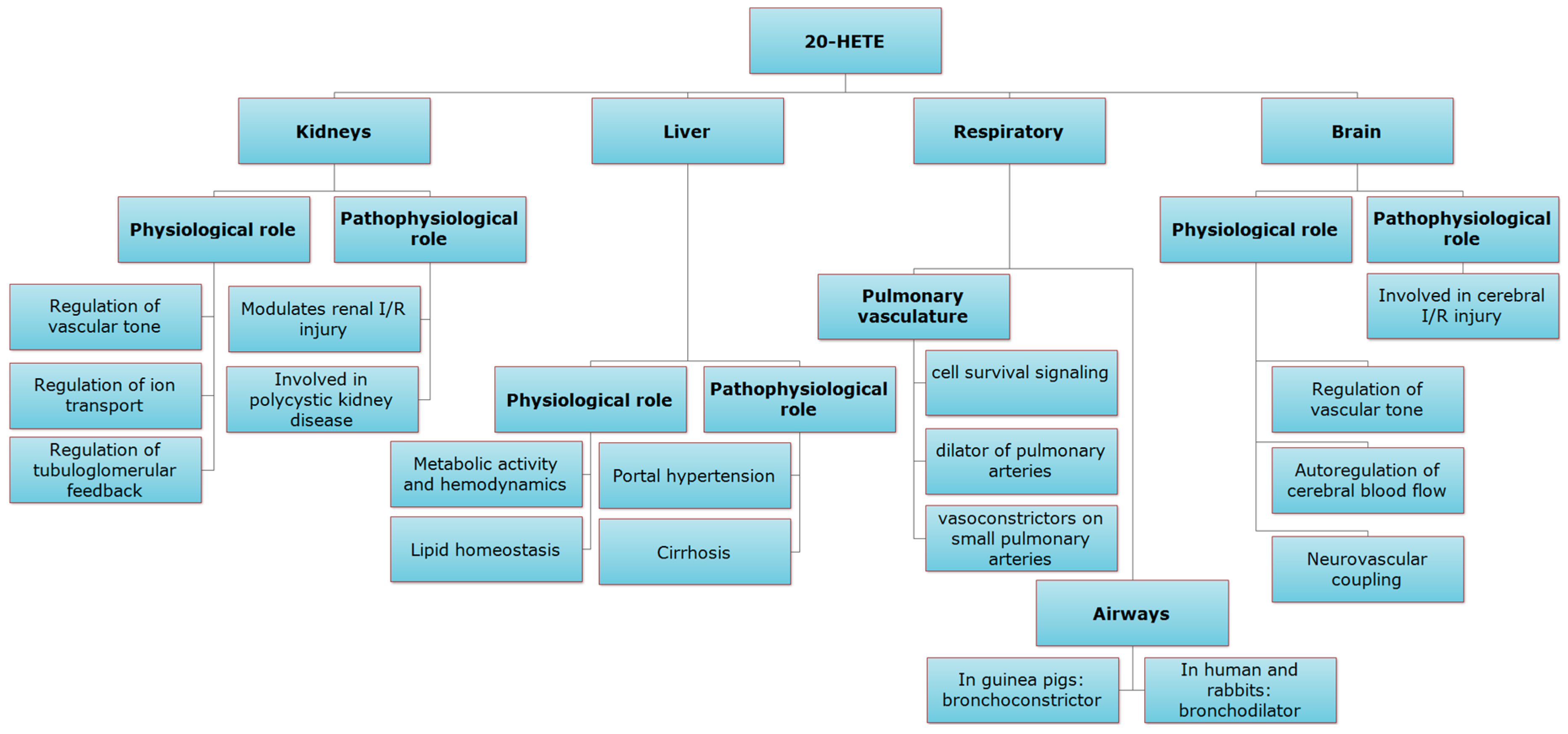
2. Role of 20-HETE in the Kidney
2.1. Biosynthesis of 20-HETE in the Kidney
2.2. Metabolism and Regulation of 20-HETE in the Kidney
2.3. Role of 20-HETE in the Kidney
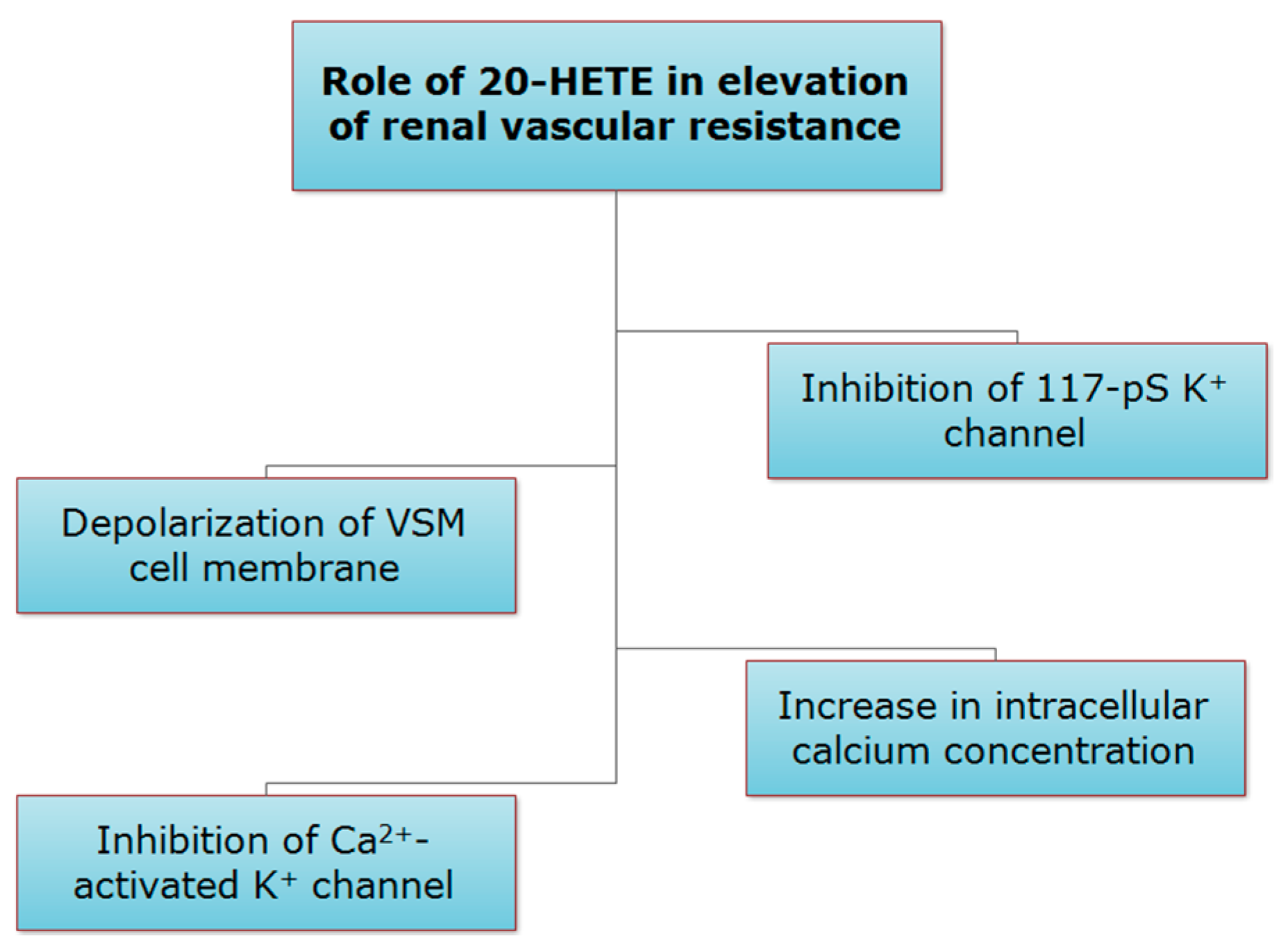
2.3.1. Role of 20-HETE in Preglomerular Vascular Tone Regulation
2.3.2. Role of 20-HETE in Tubular Ion Transport
2.4. Role of 20-HETE in Pathophysiology of the Kidney
2.4.1. Role of 20-HETE in Renal Ischemia/Reperfusion (I/R) Injury
2.4.2. Role of 20-HETE in Polycystic Kidney Diseases
2.5. 20-HETE Mediation of Drug Induced Toxicity in the Kidney
2.6. Role of 20-HETE in the Renal System during Pregnancy
3. Role of 20-HETE in the Liver
3.1. Formation of 20-HETE in the Liver
3.2. Role of 20-HETE in the Liver
4. Role of 20-HETE in the Respiratory System
4.1. Distribution of 20-HETE in Pulmonary Tissues
4.2. Role of 20-HETE in Pulmonary Cell Survival
4.3. Role of 20-HETE in Pulmonary Vascular Tone
4.4. Role of 20-HETE in the Airways
5. Role of 20-HETE in the Brain
5.1. Formation, Metabolism and Regulation of 20-HETE in the Brain
5.2. Role of 20-HETE in Regulating Vascular Tone in Brain
5.3. Role of 20-HETE in Cerebral Ischemia/Reperfusion (I/R) Injury
6. Role of 20-HETE in Tumors
7. Role of 20-HETE in Metabolic Syndrome and Diabetes
8. Summary and Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| 10-SUYS | acetylenic fatty acid sodium 10-undecynyl sulfate |
| 17-ODYA | 17-octadecynoic acid |
| 20-COOH-AA | 20-carboxyeicosatetraenoic acid |
| 20-HETE | 20-hydroxyeicosatetraenoic acid |
| 20-OH PGH2 | hydroxyl analogue of vasoconstrictor prostaglandin H2 |
| 5,14,20-HEDE; WIT003 | 20-hydroxyeicosa-5(Z),14(Z)-dienoic acid |
| 5,14,20-HEDGE | N-[20-hydroxyeicosa-5(Z),14(Z)-dienoyl]glycine |
| 6,15,20-HEDE; WIT002 | 20-hydroxyeicosa-6(Z),15(Z)-dienoic acid |
| 6,15,20-HEDGE | 20-hydroxyeicosa-6(Z),15(Z)-dienoyl]glycine |
| AA | Arachidonic acid |
| ABT | 1-Aminobenzotriazole |
| ADH | alcohol dehydrogenase |
| AngII | vasoconstrictors Angiotensin II |
| aSAH | aneurysmal subarachnoid hemorrhage model |
| ASM | airway smooth muscle cells |
| BPAECs | bovine pulmonary artery endothelial cells |
| CMV | cyto-megalovirus |
| CO | carbon monoxide |
| CoCl2 | Cobalt(II) chloride |
| COX | cyclooxygenase |
| CSF | cerebrospinal fluid |
| CYP | cytochrome P450 |
| DBDD | dibromododec-11-enoic acid |
| DDMS | N-methylsulfonyl-12,12-dibromododec-11-enamide |
| EETs | epoxyeicosatrienoic acids |
| eNOS | Endothelial NOS |
| ET | endothelin |
| EGFR | epidermal growth factor receptor |
| HET0016 | N-hydroxy-N’-(4-butyl-2methylphenyl)formamidine |
| HETEs | hydroxyeicosatetraenoic acids |
| HO | heme oxygenase enzymes |
| I/R | Ischemia/Reperfusion injury |
| KAP | kidney androgen-regulated protein |
| KCa | Ca2+-activated K+ channels |
| l-NA | nitro-l-arginine |
| l-NAME | N(ω)-nitro-l-arginine-methyl ester |
| LOX | lipoxygenase |
| LTs | leukotrienes |
| LXs | lipoxins |
| mGluR | Group I metabotropic glutamate receptors |
| NOS | NO synthase |
| NSCLC | non-small cell lung cancer cells |
| OGD | oxygen-glucose deprivation |
| Palb | albumin permeability |
| PAs | pulmonary arteries |
| PGs | prostaglandins |
| PKC | protein kinase C |
| PPARα | Peroxisome proliferator-activated receptor α |
| ROS | reactive oxygen species |
| RUPP | reductions in uterine perfusion pressure |
| RVR | renal vascular resistance |
| SAH | subarachnoid hemorrhage model |
| SD | Sprague Dawley |
| sEH | soluble epoxide hydrolase |
| siRNA | small interfering RNA |
| SPPC | Scolopendra Polysaccharide-Protein Complex |
| TAL | thick ascending limb of the loop of Henle |
| TGF-β | transforming growth factor-beta |
| TS011 | N-(3-Chloro-4-morpholin-4-yl)Phenyl-N’-hydroxyimido formamide |
| VEGF | vascular endothelial growth factor |
| VSM | vascular smooth muscle |
References
- Imig, J.D. Epoxides and soluble epoxide hydrolase in cardiovascular physiology. Physiol. Rev. 2012, 92, 101–130. [Google Scholar] [CrossRef]
- Elshenawy, O.H.; Anwar-Mohamed, A.; Abdelhamid, G.; El-Kadi, A.O. Murine atrial hl-1 cell line is a reliable model to study drug metabolizing enzymes in the heart. Vascul. Pharmacol. 2012, 58, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Elshenawy, O.H.; Anwar-Mohamed, A.; El-Kadi, A.O. 20-Hydroxyeicosatetraenoic acid is a potential therapeutic target in cardiovascular diseases. Curr. Drug Metab. 2013, 14, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Maayah, Z.H.; El-Kadi, A.O. The role of mid-chain hydroxyeicosatetraenoic acids in the pathogenesis of hypertension and cardiac hypertrophy. Arch. Toxicol. 2016, 90, 119–136. [Google Scholar] [CrossRef] [PubMed]
- Kroetz, D.L.; Xu, F. Regulation and inhibition of arachidonic acid omega-hydroxylases and 20-hete formation. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 413–438. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Falck, J.R.; Ortiz de Montellano, P.R.; Kroetz, D.L. Catalytic activity and isoform-specific inhibition of rat cytochrome P450 4f enzymes. J. Pharmacol. Exp. Ther. 2004, 308, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.; Ge, Y.; Lv, W.; Elliott, M.R.; Muroya, Y.; Hirata, T.; Booz, G.W.; Roman, R.J. Molecular mechanisms and cell signaling of 20-hydroxyeicosatetraenoic acid in vascular pathophysiology. Front Biosci. 2016, 21, 1427–1463. [Google Scholar]
- McGiff, J.C.; Quilley, J. 20-HETE and the kidney: Resolution of old problems and new beginnings. Am. J. Physiol. 1999, 277, 607–623. [Google Scholar]
- Carroll, M.A.; McGiff, J.C. A new class of lipid mediators: Cytochrome P450 arachidonate metabolites. Thorax 2000, 55 (Suppl. 2), 13–16. [Google Scholar] [CrossRef]
- Oyekan, A.O.; McGiff, J.C. Functional response of the rat kidney to inhibition of nitric oxide synthesis: Role of cytochrome P450-derived arachidonate metabolites. Br. J. Pharmacol. 1998, 125, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Askari, B.; Bell-Quilley, C.P.; Fulton, D.; Quilley, J.; McGiff, J.C. Analysis of eicosanoid mediation of the renal functional effects of hyperchloremia. J. Pharmacol. Exp. Ther. 1997, 282, 101–107. [Google Scholar] [PubMed]
- Xu, F.; Straub, W.O.; Pak, W.; Su, P.; Maier, K.G.; Yu, M.; Roman, R.J.; Ortiz De Montellano, P.R.; Kroetz, D.L. Antihypertensive effect of mechanism-based inhibition of renal arachidonic acid omega-hydroxylase activity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.M.; Murphy, S.; Burke, M.; Roman, R.J. 20-hydroxyeicosatetraeonic acid: A new target for the treatment of hypertension. J. Cardiovasc. Pharmacol. 2010, 56, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Tunctan, B.; Korkmaz, B.; Buharalioglu, C.K.; Firat, S.S.; Anjaiah, S.; Falck, J.; Roman, R.J.; Malik, K.U. A 20-hydroxyeicosatetraenoic acid agonist, n-[20-hydroxyeicosa-5(z),14(z)-dienoyl]glycine, opposes the fall in blood pressure and vascular reactivity in endotoxin-treated rats. Shock 2008, 30, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Regner, K.R.; Zuk, A.; Van Why, S.K.; Shames, B.D.; Ryan, R.P.; Falck, J.R.; Manthati, V.L.; McMullen, M.E.; Ledbetter, S.R.; Roman, R.J. Protective effect of 20-HETE analogues in experimental renal ischemia reperfusion injury. Kidney Int. 2009, 75, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Ishii, T.; Miyata, N.; Taniguchi, K.; Tomishima, Y.; Ueki, T.; Sato, M. Design and synthesis of 1-(4-benzoylphenyl)imidazole derivatives as new potent 20-HETE synthase inhibitors. Bioorg. Med. Chem. Lett. 2004, 14, 5305–5308. [Google Scholar] [CrossRef] [PubMed]
- Yanes, L.L.; Lima, R.; Moulana, M.; Romero, D.G.; Yuan, K.; Ryan, M.J.; Baker, R.; Zhang, H.; Fan, F.; Davis, D.D. Postmenopausal hypertension: Role of 20-HETE. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, 1543–1548. [Google Scholar] [CrossRef] [PubMed]
- Chabova, V.C.; Kramer, H.J.; Vaneckova, I.; Vernerova, Z.; Eis, V.; Tesar, V.; Skaroupkova, P.; Thumova, M.; Schejbalova, S.; Huskova, Z. Effects of chronic cytochrome P-450 inhibition on the course of hypertension and end-organ damage in ren-2 transgenic rats. Vascul. Pharmacol. 2007, 47, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.J. Eicosanoid receptors: Targets for the treatment of disrupted intestinal epithelial homeostasis. Eur. J. Pharmacol. 2016, 796, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Garcia, V.; Schwartzman, M.L. Recent developments on the vascular effects of 20-hydroxyeicosatetraenoic acid. Curr. Opin. Nephrol. Hypertens. 2017, 26, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Quigley, R.; Baum, M.; Reddy, K.M.; Griener, J.C.; Falck, J.R. Effects of 20-HETE and 19(s)-HETE on rabbit proximal straight tubule volume transport. Am. J. Physiol. Renal Physiol. 2000, 278, 949–953. [Google Scholar]
- Omata, K.; Abraham, N.G.; Schwartzman, M.L. Renal cytochrome P-450-arachidonic acid metabolism: Localization and hormonal regulation in shr. Am. J. Physiol. 1992, 262, 591–599. [Google Scholar]
- Oliw, E.H. Biosynthesis of 20-hydroxyeicosatetraenoic acid (20-HETE) and 12 (s)-HETE by renal cortical microsomes of the cynomolgus monkey. Eicosanoids 1990, 3, 161–164. [Google Scholar] [PubMed]
- Lu, M.; Zhu, Y.; Balazy, M.; Reddy, K.M.; Falck, J.R.; Wang, W. Effect of angiotensin ii on the apical K+ channel in the thick ascending limb of the rat kidney. J. Gen. Physiol. 1996, 108, 537–547. [Google Scholar] [CrossRef]
- Escalante, B.; Erlij, D.; Falck, J.R.; McGiff, J.C. Cytochrome P450-dependent arachidonate metabolites affect renal transport in the rabbit. J. Cardiovasc. Pharmacol. 1993, 22 (Suppl. 2), 106–108. [Google Scholar] [CrossRef]
- Marji, J.S.; Wang, M.H.; Laniado-Schwartzman, M. Cytochrome P-450 4a isoform expression and 20-HETE synthesis in renal preglomerular arteries. Am. J. Physiol. Renal Physiol. 2002, 283, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.H.; Guan, H.; Nguyen, X.; Zand, B.A.; Nasjletti, A.; Laniado-Schwartzman, M. Contribution of cytochrome P-450 4a1 and 4a2 to vascular 20-hydroxyeicosatetraenoic acid synthesis in rat kidneys. Am. J. Physiol. 1999, 276, 246–253. [Google Scholar]
- Zhao, X.; Imig, J.D. Kidney cyP450 enzymes: Biological actions beyond drug metabolism. Curr. Drug Metab. 2003, 4, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Ward, N.C.; Tsai, I.J.; Barden, A.; van Bockxmeer, F.M.; Puddey, I.B.; Hodgson, J.M.; Croft, K.D. A single nucleotide polymorphism in the cyp4f2 but not cyp4a11 gene is associated with increased 20-HETE excretion and blood pressure. Hypertension 2008, 51, 1393–1398. [Google Scholar] [CrossRef] [PubMed]
- Harder, D.R.; Campbell, W.B.; Roman, R.J. Role of cytochrome P-450 enzymes and metabolites of arachidonic acid in the control of vascular tone. J. Vasc. Res. 1995, 32, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.S.; Zhang, F.; Jiang, M.; Wang, M.H.; Zand, B.A.; Abraham, N.G.; Nasjletti, A.; Laniado-Schwartzman, M. Transfection and functional expression of CYP4A1 and CYP4A2 using bicistronic vectors in vascular cells and tissues. J. Pharmacol. Exp. Ther. 2004, 311, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Kaide, J.; Wang, M.H.; Wang, J.S.; Zhang, F.; Gopal, V.R.; Falck, J.R.; Nasjletti, A.; Laniado-Schwartzman, M. Transfection of CYP4A1 cDNA increases vascular reactivity in renal interlobar arteries. Am. J. Physiol. Renal Physiol. 2003, 284, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Murphy, S.R.; Lu, Y.; Falck, J.; Liu, R.; Roman, R.J. Endogenously produced 20-HETE modulates myogenic and TGF response in microperfused afferent arterioles. Prostaglandin Other Lipid Mediat. 2013, 102, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Minuz, P.; Jiang, H.; Fava, C.; Turolo, L.; Tacconelli, S.; Ricci, M.; Patrignani, P.; Morganti, A.; Lechi, A.; McGiff, J.C. Altered release of cytochrome P450 metabolites of arachidonic acid in renovascular disease. Hypertension 2008, 51, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Dahly-Vernon, A.J.; Sharma, M.; McCarthy, E.T.; Savin, V.J.; Ledbetter, S.R.; Roman, R.J. Transforming growth factor-β, 20-HETE interaction, and glomerular injury in DAHL salt-sensitive rats. Hypertension 2005, 45, 643–648. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, E.T.; Sharma, R.; Sharma, M. Protective effect of 20-hydroxyeicosatetraenoic acid (20-HETE) on glomerular protein permeability barrier. Kidney Int. 2005, 67, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Zhou, Y.; Chang, H.H.; Zhang, J.; Seki, T.; Wang, C.Y.; Inscho, E.W.; Wang, M.H. Glomerular 20-HETE, EETs, and TGF-β1 in diabetic nephropathy. Am. J. Physiol. Renal Physiol. 2009, 296, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Stec, D.E.; Flasch, A.; Roman, R.J.; White, J.A. Distribution of cytochrome P-450 4A and 4F isoforms along the nephron in mice. Am. J. Physiol. Renal Physiol. 2003, 284, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Ito, O.; Alonso-Galicia, M.; Hopp, K.A.; Roman, R.J. Localization of cytochrome P-450 4A isoforms along the rat nephron. Am. J. Physiol. 1998, 274, 395–404. [Google Scholar]
- Harder, D.R.; Narayanan, J.; Birks, E.K.; Liard, J.F.; Imig, J.D.; Lombard, J.H.; Lange, A.R.; Roman, R.J. Identification of a putative microvascular oxygen sensor. Circ. Res. 1996, 79, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Kirita, S.; Hasegawa, H.; Aoyama, J.; Imaoka, S.; Minamiyama, S.; Funae, Y.; Baba, T.; Matsubara, T. Contribution of CYP4A8 to the formation of 20-hydroxyeicosatetraenoic acid from arachidonic acid in rat kidney. Drug Metab. Pharmacokinet. 2002, 17, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Hercule, H.C.; Wang, M.H.; Oyekan, A.O. Contribution of cytochrome P450 4a isoforms to renal functional response to inhibition of nitric oxide production in the rat. J. Physiol. 2003, 551, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Mezentsev, A.; Kemp, R.; Byun, K.; Falck, J.R.; Miano, J.M.; Nasjletti, A.; Abraham, N.G.; Laniado-Schwartzman, M. Smooth muscle-specific expression of CYP4A1 induces endothelial sprouting in renal arterial microvessels. Circ. Res. 2004, 94, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, X.; Wang, M.H.; Reddy, K.M.; Falck, J.R.; Schwartzman, M.L. Kinetic profile of the rat CYP4A isoforms: Arachidonic acid metabolism and isoform-specific inhibitors. Am. J. Physiol. 1999, 276, 1691–1700. [Google Scholar]
- Botros, F.T.; Laniado-Schwartzman, M.; Abraham, N.G. Regulation of cyclooxygenase and cytochrome P450 derived eicosanoids by heme oxygenase in the rat kidney. Hypertension 2002, 39, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Kalsotra, A.; Cui, X.; Anakk, S.; Hinojos, C.A.; Doris, P.A.; Strobel, H.W. Renal localization, expression, and developmental regulation of P450 4f cytochromes in three substrains of spontaneously hypertensive rats. Biochem. Biophys. Res. Commun. 2005, 338, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Fidelis, P.; Wilson, L.; Thomas, K.; Villalobos, M.; Oyekan, A.O. Renal function and vasomotor activity in mice lacking the CYP4A14 gene. Exp. Biol. Med. 2010, 235, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- Muller, D.N.; Schmidt, C.; Barbosa-Sicard, E.; Wellner, M.; Gross, V.; Hercule, H.; Markovic, M.; Honeck, H.; Luft, F.C.; Schunck, W.H. Mouse CYP4A isoforms: Enzymatic properties, gender- and strain-specific expression, and role in renal 20-hydroxyeicosatetraenoic acid formation. Biochem. J. 2007, 403, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Lasker, J.M.; Chen, W.B.; Wolf, I.; Bloswick, B.P.; Wilson, P.D.; Powell, P.K. Formation of 20-hydroxyeicosatetraenoic acid, a vasoactive and natriuretic eicosanoid, in human kidney. Role of cyp4f2 and cyp4A11. J. Biol. Chem. 2000, 275, 4118–4126. [Google Scholar] [CrossRef] [PubMed]
- Muroya, Y.; Fan, F.; Regner, K.R.; Falck, J.R.; Garrett, M.R.; Juncos, L.A.; Roman, R.J. Deficiency in the formation of 20-hydroxyeicosatetraenoic acid enhances renal ischemia-reperfusion injury. J. Am. Soc. Nephrol. 2015, 26, 2460–2469. [Google Scholar] [CrossRef] [PubMed]
- Stec, D.E.; Roman, R.J.; Flasch, A.; Rieder, M.J. Functional polymorphism in human CYP4F2 decreases 20-HETE production. Physiol. Genomics 2007, 30, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.K.; McGiff, J.C.; Carroll, M.A. Renal arterial 20-hydroxyeicosatetraenoic acid levels: Regulation by cyclooxygenase. Am. J. Physiol. Renal Physiol. 2003, 284, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Carroll, M.A.; Kemp, R.; Cheng, M.K.; McGiff, J.C. Regulation of preglomerular microvascular 20-hydroxyeicosatetraenoic acid levels by salt depletion. Med. Sci. Monit. 2001, 7, 567–572. [Google Scholar] [PubMed]
- Alonso-Galicia, M.; Drummond, H.A.; Reddy, K.K.; Falck, J.R.; Roman, R.J. Inhibition of 20-HETE production contributes to the vascular responses to nitric oxide. Hypertension 1997, 29, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Oyekan, A.O.; Youseff, T.; Fulton, D.; Quilley, J.; McGiff, J.C. Renal cytochrome P450 ϖ-hydroxylase and epoxygenase activity are differentially modified by nitric oxide and sodium chloride. J. Clin. Invest. 1999, 104, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Quilley, J.; Qiu, Y.; Hirt, J. Inhibitors of 20-hydroxyeicosatetraenoic acid reduce renal vasoconstrictor responsiveness. J. Pharmacol. Exp. Ther. 2003, 307, 223–229. [Google Scholar] [CrossRef]
- Ito, O.; Roman, R.J. Regulation of P-450 4a activity in the glomerulus of the rat. Am. J. Physiol. 1999, 276, 1749–1757. [Google Scholar]
- Alonso-Galicia, M.; Sun, C.W.; Falck, J.R.; Harder, D.R.; Roman, R.J. Contribution of 20-HETE to the vasodilator actions of nitric oxide in renal arteries. Am. J. Physiol. 1998, 275, 370–378. [Google Scholar]
- Sarkis, A.; Roman, R.J. Role of cytochrome P450 metabolites of arachidonic acid in hypertension. Curr. Drug Metab. 2004, 5, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, J.L.; Zand, B.A.; Yang, L.M.; Sabaawy, H.E.; Lianos, E.; Abraham, N.G. Heme oxygenase isoform-specific expression and distribution in the rat kidney. Kidney Int. 2001, 59, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Abraham, N.G.; Schwartzman, M.L. Cytochrome P450 arachidonic acid omega-hydroxylation in the proximal tubule of the rat kidney. Ann. N. Y. Acad. Sci. 1994, 744, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, T.; Ito, O.; Tan, L.; Ogawa, S.; Kohzuki, M.; Omata, K.; Takeuchi, K.; Ito, S. Regulation of cytochrome P-450 4a activity by peroxisome proliferator-activated receptors in the rat kidney. Hypertens. Res. 2003, 26, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, D.; Tie, C.; Wu, J.; Wu, Q.; Li, Q. Cisplatin-mediated cytotoxicity through inducing CYP4A 11 expression in human renal tubular epithelial cells. J. Toxicol. Sci. 2015, 40, 895–900. [Google Scholar] [CrossRef] [PubMed]
- Sarkis, A.; Ito, O.; Mori, T.; Kohzuki, M.; Ito, S.; Verbalis, J.; Cowley, A.W., Jr.; Roman, R.J. Cytochrome P-450-dependent metabolism of arachidonic acid in the kidney of rats with diabetes insipidus. Am. J. Physiol. Renal Physiol. 2005, 289, 1333–1340. [Google Scholar] [CrossRef] [PubMed]
- Maier, K.G.; Roman, R.J. Cytochrome P450 metabolites of arachidonic acid in the control of renal function. Curr. Opin. Nephrol. Hypertens. 2001, 10, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Zou, A.P.; Imig, J.D.; Kaldunski, M.; Ortiz de Montellano, P.R.; Sui, Z.; Roman, R.J. Inhibition of renal vascular 20-HETE production impairs autoregulation of renal blood flow. Am. J. Physiol. 1994, 266, 275–282. [Google Scholar]
- Imaoka, S.; Funae, Y. The physiological role of P450-derived arachidonic acid metabolites. Nihon Yakurigaku Zasshi 1998, 112, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Zou, A.P.; Imig, J.D.; Ortiz de Montellano, P.R.; Sui, Z.; Falck, J.R.; Roman, R.J. Effect of P-450 ϖ-hydroxylase metabolites of arachidonic acid on tubuloglomerular feedback. Am. J. Physiol. 1994, 266, 934–941. [Google Scholar]
- Schwartzman, M.L.; Omata, K.; Lin, F.M.; Bhatt, R.K.; Falck, J.R.; Abraham, N.G. Detection of 20-hydroxyeicosatetraenoic acid in rat urine. Biochem. Biophys. Res. Commun. 1991, 180, 445–449. [Google Scholar] [CrossRef]
- Roman, R.J.; Alonso-Galicia, M. P-450 eicosanoids: A novel signaling pathway regulating renal function. News Physiol. Sci. 1999, 14, 238–242. [Google Scholar] [PubMed]
- Kauser, K.; Clark, J.E.; Masters, B.S.; Ortiz de Montellano, P.R.; Ma, Y.H.; Harder, D.R.; Roman, R.J. Inhibitors of cytochrome P-450 attenuate the myogenic response of dog renal arcuate arteries. Circ. Res. 1991, 68, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Mei, S.; Cheng, J.; Ding, Y.; Weidenhammer, A.; Garcia, V.; Zhang, F.; Gotlinger, K.; Manthati, V.L.; Falck, J.R.; et al. Androgen-sensitive hypertension associates with upregulated vascular CYP4A12-20-HETE synthase. J. Am. Soc. Nephrol. 2013, 24, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Falck, J.R.; Gopal, V.R.; Inscho, E.W.; Imig, J.D. P2x receptor-stimulated calcium responses in preglomerular vascular smooth muscle cells involves 20-hydroxyeicosatetraenoic acid. J. Pharmacol. Exp. Ther. 2004, 311, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.H.; Gebremedhin, D.; Schwartzman, M.L.; Falck, J.R.; Clark, J.E.; Masters, B.S.; Harder, D.R.; Roman, R.J. 20-hydroxyeicosatetraenoic acid is an endogenous vasoconstrictor of canine renal arcuate arteries. Circ. Res. 1993, 72, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Arima, S.; Omata, K.; Ito, S.; Tsunoda, K.; Abe, K. 20-HETE requires increased vascular tone to constrict rabbit afferent arterioles. Hypertension 1996, 27, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Gupta, T.; Garcia, V.; Ding, Y.; Schwartzman, M.L. 20-HETE and blood pressure regulation: Clinical implications. Cardiol. Rev. 2014, 22, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zou, A.P.; Fleming, J.T.; Falck, J.R.; Jacobs, E.R.; Gebremedhin, D.; Harder, D.R.; Roman, R.J. 20-HETE is an endogenous inhibitor of the large-conductance Ca2+-activated K+ channel in renal arterioles. Am. J. Physiol. 1996, 270, 228–237. [Google Scholar]
- Harder, D.R.; Lange, A.R.; Gebremedhin, D.; Birks, E.K.; Roman, R.J. Cytochrome P450 metabolites of arachidonic acid as intracellular signaling molecules in vascular tissue. J. Vasc. Res. 1997, 34, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Roman, R.J.; Harder, D.R. Cellular and ionic signal transduction mechanisms for the mechanical activation of renal arterial vascular smooth muscle. J. Am. Soc. Nephrol. 1993, 4, 986–996. [Google Scholar] [PubMed]
- Imig, J.D.; Zou, A.P.; Stec, D.E.; Harder, D.R.; Falck, J.R.; Roman, R.J. Formation and actions of 20-hydroxyeicosatetraenoic acid in rat renal arterioles. Am. J. Physiol. 1996, 270, 217–227. [Google Scholar]
- Sun, C.W.; Falck, J.R.; Harder, D.R.; Roman, R.J. Role of tyrosine kinase and PKC in the vasoconstrictor response to 20-HETE in renal arterioles. Hypertension 1999, 33, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.W.; Alonso-Galicia, M.; Taheri, M.R.; Falck, J.R.; Harder, D.R.; Roman, R.J. Nitric oxide-20-hydroxyeicosatetraenoic acid interaction in the regulation of K+ channel activity and vascular tone in renal arterioles. Circ. Res. 1998, 83, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Kuczeriszka, M.; Badzynska, B.; Kompanowska-Jezierska, E. Cytochrome P-450 monooxygenases in control of renal haemodynamics and arterial pressure in anaesthetized rats. J. Physiol. Pharmacol. 2006, 57 (Suppl. 11), 179–185. [Google Scholar] [PubMed]
- Oyekan, A.O. Differential effects of 20-hydroxyeicosatetraenoic acid on intrarenal blood flow in the rat. J. Pharmacol. Exp. Ther. 2005, 313, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- Hercule, H.C.; Oyekan, A.O. Role of no and cytochrome P-450-derived eicosanoids in et-1-induced changes in intrarenal hemodynamics in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, 2132–2141. [Google Scholar]
- Hercule, H.C.; Oyekan, A.O. Cytochrome P450 ϖ/ϖ-1 hydroxylase-derived eicosanoids contribute to endothelin(a) and endothelin(b) receptor-mediated vasoconstriction to endothelin-1 in the rat preglomerular arteriole. J. Pharmacol. Exp. Ther. 2000, 292, 1153–1160. [Google Scholar] [PubMed]
- Oyekan, A.O.; McAward, K.; McGiff, J.C. Renal functional effects of endothelins: Dependency on cytochrome P450-derived arachidonate metabolites. Biol. Res. 1998, 31, 209–215. [Google Scholar] [PubMed]
- Lai, G.; Liu, X.; Wu, J.; Liu, H.; Zhao, Y. Evaluation of CMV and KAP promoters for driving the expression of human CYP4F2 in transgenic mice. Int. J. Mol. Med. 2012, 29, 107–112. [Google Scholar] [PubMed]
- Zhang, B.; Lai, G.; Liu, X.; Zhao, Y. Alteration of epoxyeicosatrienoic acids in the liver and kidney of cytochrome P450 4f2 transgenic mice. Mol. Med. Rep. 2016, 14, 5739–5745. [Google Scholar] [CrossRef] [PubMed]
- Roman, R.J. P-450 metabolites of arachidonic acid in the control of cardiovascular function. Physiol. Rev. 2002, 82, 131–185. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Belusa, R.; Nowicki, S.; Aperia, A. Arachidonic acid metabolic pathways regulating activity of renal Na+-K+-atpase are age dependent. Am. J. Physiol. Renal Physiol. 2000, 278, 823–829. [Google Scholar]
- Nowicki, S.; Chen, S.L.; Aizman, O.; Cheng, X.J.; Li, D.; Nowicki, C.; Nairn, A.; Greengard, P.; Aperia, A. 20-hydroxyeicosa-tetraenoic acid (20 HETE) activates protein kinase c: Role in regulation of rat renal Na+-K+-atpase. J. Clin. Invest. 1997, 99, 1224–1230. [Google Scholar] [CrossRef] [PubMed]
- Sacerdoti, D.; Gatta, A.; McGiff, J.C. Role of cytochrome P450-dependent arachidonic acid metabolites in liver physiology and pathophysiology. Prostaglandins Other Lipid Mediat. 2003, 72, 51–71. [Google Scholar] [CrossRef]
- Cloutier, M.; Campbell, S.; Basora, N.; Proteau, S.; Payet, M.D.; Rousseau, E. 20-HETE inotropic effects involve the activation of a nonselective cationic current in airway smooth muscle. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 285, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.; Muroya, Y.; Roman, R.J. Cytochrome P450 eicosanoids in hypertension and renal disease. Curr. Opin. Nephrol. Hypertens. 2015, 24, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Quigley, R.; Chakravarty, S.; Zhao, X.; Imig, J.D.; Capdevila, J.H. Increased renal proximal convoluted tubule transport contributes to hypertension in CYP4A14 knockout mice. Nephron Physiol. 2009, 113, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Schwartzman, M.L. The role of 20-HETE in androgen-mediated hypertension. Prostaglandins Other Lipid Mediat. 2011, 96, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Holla, V.R.; Adas, F.; Imig, J.D.; Zhao, X.; Price, E., Jr.; Olsen, N.; Kovacs, W.J.; Magnuson, M.A.; Keeney, D.S.; Breyer, M.D. Alterations in the regulation of androgen-sensitive Cyp 4a monooxygenases cause hypertension. Proc. Natl. Acad. Sci. USA 2001, 98, 5211–5216. [Google Scholar] [CrossRef] [PubMed]
- Beltowski, J.; Marciniak, A.; Wojcicka, G.; Gorny, D. The opposite effects of cyclic amp-protein kinase a signal transduction pathway on renal cortical and medullary Na+,K+-atpase activity. J. Physiol. Pharmacol. 2002, 53, 211–231. [Google Scholar] [PubMed]
- Derrickson, B.H.; Mandel, L.J. Parathyroid hormone inhibits Na+-K+-atpase through Gq/G11 and the calcium-independent phospholipase A2. Am. J. Physiol. 1997, 272, F781–F788. [Google Scholar] [PubMed]
- Kirchheimer, C.; Mendez, C.F.; Acquier, A.; Nowicki, S. Role of 20-HETE in D1/D2 dopamine receptor synergism resulting in the inhibition of Na+-K+-ATPase activity in the proximal tubule. Am. J. Physiol. Renal Physiol. 2007, 292, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Ominato, M.; Satoh, T.; Katz, A.I. Regulation of Na+-K+-atpase activity in the proximal tubule: Role of the protein kinase c pathway and of eicosanoids. J. Membr. Biol. 1996, 152, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, D.M.; Barac-Nieto, M.; Falck, J.R.; Spitzer, A. 20-HETE mediates the effect of parathyroid hormone and protein kinase c on renal phosphate transport. Prostaglandins Leukot. Essent. Fatty Acids 1998, 58, 209–213. [Google Scholar] [CrossRef]
- Silverstein, D.M.; Barac-Nieto, M.; Spitzer, A. Multiple arachidonic acid metabolites inhibit sodium-dependent phosphate transport in ok cells. Prostaglandins Leukot. Essent. Fatty Acids 1999, 61, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, D.M.; Spitzer, A.; Barac-Nieto, M. Hormonal regulation of sodium-dependent phosphate transport in opossum kidney cells. Horm. Res. 2000, 54, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, D.M.; Spitzer, A.; Barac-Nieto, M. Parathormone sensitivity and responses to protein kinases in subclones of opossum kidney cells. Pediatr. Nephrol. 2005, 20, 721–724. [Google Scholar] [CrossRef] [PubMed]
- Amlal, H.; Legoff, C.; Vernimmen, C.; Paillard, M.; Bichara, M. Na+-K+-Nh4+-2Cl− cotransport in medullary thick ascending limb: Control by pKa, pKc, and 20-HETE. Am. J. Physiol. 1996, 271, 455–463. [Google Scholar]
- Grider, J.S.; Falcone, J.C.; Kilpatrick, E.L.; Ott, C.E.; Jackson, B.A. P450 arachidonate metabolites mediate bradykinin-dependent inhibition of NACl transport in the rat thick ascending limb. Can. J. Physiol. Pharmacol. 1997, 75, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lu, M. Effect of arachidonic acid on activity of the apical K+ channel in the thick ascending limb of the rat kidney. J. Gen. Physiol. 1995, 106, 727–743. [Google Scholar] [CrossRef] [PubMed]
- Gu, R.M.; Wei, Y.; Jiang, H.L.; Lin, D.H.; Sterling, H.; Bloom, P.; Balazy, M.; Wang, W.H. K depletion enhances the extracellular Ca2+-induced inhibition of the apical k channels in the mTAL of rat kidney. J. Gen. Physiol. 2002, 119, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wei, Y.; Wang, W.H. Dietary K intake regulates the response of apical K channels to adenosine in the thick ascending limb. Am. J. Physiol. Renal Physiol. 2004, 287, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Ikari, A.; Nakajima, K.; Suketa, Y.; Harada, H.; Takagi, K. Activation of Na+-independent Mg2+ efflux by 20-hydroxyeicosatetraenoic acid in rat renal epithelial cells. Jpn. J. Physiol. 2004, 54, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Savas, U.; Wei, S.; Hsu, M.H.; Falck, J.R.; Guengerich, F.P.; Capdevila, J.H.; Johnson, E.F. 20-hydroxyeicosatetraenoic acid (HETE)-dependent hypertension in human cytochrome P450 (CYP) 4A11 transgenic mice: Normalization of blood pressure by sodium restriction, hydrochlorothiazide, or blockade of the type 1 angiotensin ii receptor. J. Biol. Chem. 2016, 291, 16904–16919. [Google Scholar] [CrossRef] [PubMed]
- Coca, S.G.; Yusuf, B.; Shlipak, M.G.; Garg, A.X.; Parikh, C.R. Long-term risk of mortality and other adverse outcomes after acute kidney injury: A systematic review and meta-analysis. Am. J. Kidney Dis. 2009, 53, 961–973. [Google Scholar] [CrossRef] [PubMed]
- Bonventre, J.V.; Yang, L. Cellular pathophysiology of ischemic acute kidney injury. J. Clin. Invest. 2011, 121, 4210–4221. [Google Scholar] [CrossRef] [PubMed]
- Siew, E.D.; Davenport, A. The growth of acute kidney injury: A rising tide or just closer attention to detail? Kidney Int. 2015, 87, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Dolegowska, B.; Blogowski, W.; Domanski, L. Is it possible to predict the early post-transplant allograft function using 20-HETE measurements? A preliminary report. Transpl. Int. 2009, 22, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Roman, R.J.; Akbulut, T.; Park, F.; Regner, K.R. 20-HETE in acute kidney injury. Kidney Int. 2011, 79, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Nilakantan, V.; Maenpaa, C.; Jia, G.; Roman, R.J.; Park, F. 20-HETE-mediated cytotoxicity and apoptosis in ischemic kidney epithelial cells. Am. J. Physiol. Renal Physiol. 2008, 294, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Hoff, U.; Lukitsch, I.; Chaykovska, L.; Ladwig, M.; Arnold, C.; Manthati, V.L.; Fuller, T.F.; Schneider, W.; Gollasch, M.; Muller, D.N. Inhibition of 20-HETE synthesis and action protects the kidney from ischemia/reperfusion injury. Kidney Int. 2011, 79, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Regner, K.R.; Roman, R.J. Role of medullary blood flow in the pathogenesis of renal ischemia-reperfusion injury. Curr. Opin. Nephrol. Hypertens. 2012, 21, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Park, F.; Sweeney, W.E.; Jia, G.; Roman, R.J.; Avner, E.D. 20-HETE mediates proliferation of renal epithelial cells in polycystic kidney disease. J. Am. Soc. Nephrol. 2008, 19, 1929–1939. [Google Scholar] [CrossRef] [PubMed]
- Park, F.; Sweeney, W.E., Jr.; Jia, G.; Akbulut, T.; Mueller, B.; Falck, J.R.; Birudaraju, S.; Roman, R.J.; Avner, E.D. Chronic blockade of 20-HETE synthesis reduces polycystic kidney disease in an orthologous rat model of arpkd. Am. J. Physiol. Renal Physiol. 2009, 296, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Klawitter, J.; Klawitter, J.; McFann, K.; Pennington, A.T.; Abebe, K.Z.; Brosnahan, G.; Cadnapaphornchai, M.A.; Chonchol, M.; Gitomer, B.; Christians, U.; et al. Bioactive lipid mediators in polycystic kidney disease. J. Lipid Res. 2014, 55, 1139–1149. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Li, J.; Wang, M.Y.; Xie, X.F.; Li, Q.X. Protective effect of 20-hydroxyeicosatetraenoic acid (20-HETE) on adriamycin-induced toxicity of human renal tubular epithelial cell (HK-2). Eur. J. Pharmacol. 2012, 683, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Blanton, A.; Nsaif, R.; Hercule, H.; Oyekan, A. Nitric oxide/cytochrome P450 interactions in cyclosporin a-induced effects in the rat. J. Hypertens. 2006, 24, 1865–1872. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Ishimoto, T.; Sakurai, T.; Yasuda, Y.; Taniguchi, K.; Doi, M.; Sato, M.; Roman, R.J.; Miyata, N. Increased excretion of urinary 20-HETE in rats with cyclosporine-induced nephrotoxicity. J. Pharmacol. Sci. 2005, 97, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Vickers, A.E.; Alegret, M.; Meyer, E.; Smiley, S.; Guertler, J. Hydroxyethyl cyclosporin a induces and decreases P4503a and p-glycoprotein levels in rat liver. Xenobiotica 1996, 26, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.H.; Zand, B.A.; Nasjletti, A.; Laniado-Schwartzman, M. Renal 20-hydroxyeicosatetraenoic acid synthesis during pregnancy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 282, 383–389. [Google Scholar]
- Wang, M.H.; Wang, J.; Chang, H.H.; Zand, B.A.; Jiang, M.; Nasjletti, A.; Laniado-Schwartzman, M. Regulation of renal CYP4A expression and 20-HETE synthesis by nitric oxide in pregnant rats. Am. J. Physiol. Renal Physiol. 2003, 285, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Zhou, Y.; Raju, V.T.; Du, J.; Chang, H.H.; Wang, C.Y.; Brands, M.W.; Falck, J.R.; Wang, M.H. Renal 20-HETE inhibition attenuates changes in renal hemodynamics induced by l-name treatment in pregnant rats. Am. J. Physiol. Renal Physiol. 2005, 289, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Llinas, M.T.; Alexander, B.T.; Capparelli, M.F.; Carroll, M.A.; Granger, J.P. Cytochrome P-450 inhibition attenuates hypertension induced by reductions in uterine perfusion pressure in pregnant rats. Hypertension 2004, 43, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Rifkind, A.B.; Lee, C.; Chang, T.K.; Waxman, D.J. Arachidonic acid metabolism by human cytochrome P450s 2c8, 2c9, 2e1, and 1a2: Regioselective oxygenation and evidence for a role for CYP2C enzymes in arachidonic acid epoxygenation in human liver microsomes. Arch. Biochem. Biophys. 1995, 320, 380–389. [Google Scholar] [CrossRef]
- Powell, P.K.; Wolf, I.; Jin, R.; Lasker, J.M. Metabolism of arachidonic acid to 20-hydroxy-5,8,11,14-eicosatetraenoic acid by P450 enzymes in human liver: Involvement of cyp4f2 and CYP4A11. J. Pharmacol. Exp. Ther. 1998, 285, 1327–1336. [Google Scholar] [PubMed]
- Antoun, J.; Goulitquer, S.; Amet, Y.; Dreano, Y.; Salaun, J.P.; Corcos, L.; Plee-Gautier, E. CYP4F3B is induced by PGA1 in human liver cells: A regulation of the 20-HETE synthesis. J. Lipid Res. 2008, 49, 2135–2141. [Google Scholar] [CrossRef] [PubMed]
- Plee-Gautier, E.; Antoun, J.; Goulitquer, S.; Le Jossic-Corcos, C.; Simon, B.; Amet, Y.; Salaun, J.P.; Corcos, L. Statins increase cytochrome P450 4F3-mediated eicosanoids production in human liver cells: A PXR dependent mechanism. Biochem. Pharmacol. 2012, 84, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Poloyac, S.M.; Tortorici, M.A.; Przychodzin, D.I.; Reynolds, R.B.; Xie, W.; Frye, R.F.; Zemaitis, M.A. The effect of isoniazid on CYP2E1- and CYP4A-mediated hydroxylation of arachidonic acid in the rat liver and kidney. Drug Metab. Dispos. 2004, 32, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Sacerdoti, D.; Jiang, H.; Gaiani, S.; McGiff, J.C.; Gatta, A.; Bolognesi, M. 11,12-EET increases porto-sinusoidal resistance and may play a role in endothelial dysfunction of portal hypertension. Prostaglandins Other Lipid Mediat. 2011, 96, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Sacerdoti, D.; Balazy, M.; Angeli, P.; Gatta, A.; McGiff, J.C. Eicosanoid excretion in hepatic cirrhosis: Predominance of 20-HETE. J. Clin. Invest. 1997, 100, 1264–1270. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.D.; Hammock, B.D. Soluble epoxide hydrolase as a therapeutic target for cardiovascular diseases. Nat. Rev. Drug Discov. 2009, 8, 794–805. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Zhang, C.; Medhora, M.; Jacobs, E.R. CYP4A mrna, protein, and product in rat lungs: Novel localization in vascular endothelium. J. Appl. Physiol. 2002, 93, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Lakhkar, A.; Dhagia, V.; Joshi, S.R.; Gotlinger, K.; Patel, D.; Sun, D.; Wolin, M.S.; Schwartzman, M.L.; Gupte, S.A. 20-HETE-induced mitochondrial superoxide production and inflammatory phenotype in vascular smooth muscle is prevented by glucose-6-phosphate dehydrogenase inhibition. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, 1107–1117. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Effros, R.M.; Harder, D.R.; Roman, R.J.; Jacobs, E.R. Tissue sources of cytochrome P450 4a and 20-HETE synthesis in rabbit lungs. Am. J. Respir. Cell Mol. Biol. 1998, 19, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Yaghi, A.; Bradbury, J.A.; Zeldin, D.C.; Mehta, S.; Bend, J.R.; McCormack, D.G. Pulmonary cytochrome P-450 2j4 is reduced in a rat model of acute pseudomonas pneumonia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 285, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Eiserich, J.P.; Cross, C.E.; Morrissey, B.M.; Hammock, B.D. Metabolomic profiling of regulatory lipid mediators in sputum from adult cystic fibrosis patients. Free Radic. Biol. Med. 2012, 53, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, E.R.; Bodiga, S.; Ali, I.; Falck, A.M.; Falck, J.R.; Medhora, M.; Dhanasekaran, A. Tissue protection and endothelial cell signaling by 20-HETE analogs in intact ex vivo lung slices. Exp. Cell Res. 2012, 16, 2143–2152. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Birks, E.K.; Dawson, C.A.; Patel, M.; Falck, J.R.; Presberg, K.; Roman, R.J.; Jacobs, E.R. Hypoxic pulmonary vasoconstriction is modified by P-450 metabolites. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, 1526–1533. [Google Scholar]
- Dhanasekaran, A.; Bodiga, S.; Gruenloh, S.; Gao, Y.; Dunn, L.; Falck, J.R.; Buonaccorsi, J.N.; Medhora, M.; Jacobs, E.R. 20-HETE increases survival and decreases apoptosis in pulmonary arteries and pulmonary artery endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tang, X.; Li, Y.; Leu, C.; Guo, L.; Zheng, X.; Zhu, D. 20-Hydroxyeicosatetraenoic acid inhibits the apoptotic responses in pulmonary artery smooth muscle cells. Eur. J. Pharmacol. 2008, 588, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Medhora, M.; Chen, Y.; Gruenloh, S.; Harland, D.; Bodiga, S.; Zielonka, J.; Gebremedhin, D.; Gao, Y.; Falck, J.R.; Anjaiah, S.; et al. 20-HETE increases superoxide production and activates NAPDH oxidase in pulmonary artery endothelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 294, 902–911. [Google Scholar] [CrossRef] [PubMed]
- Bodiga, S.; Gruenloh, S.K.; Gao, Y.; Manthati, V.L.; Dubasi, N.; Falck, J.R.; Medhora, M.; Jacobs, E.R. 20-HETE-induced nitric oxide production in pulmonary artery endothelial cells is mediated by NADPH oxidase, H2O2, and PI3-kinase/Akt. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 298, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; McAndrew, R.P.; Al-Saghir, R.; Maier, K.G.; Medhora, M.; Roman, R.J.; Jacobs, E.R. Nitric oxide contributes to 20-HETE-induced relaxation of pulmonary arteries. J. Appl. Physiol. 2002, 93, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Fuloria, M.; Eckman, D.M.; Leach, D.A.; Aschner, J.L. 20-hydroxyeicosatetraenoic acid is a vasoconstrictor in the newborn piglet pulmonary microcirculation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 287, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.; Guibert, C.; Sirois, M.; Echave, V.; Gomes, M.M.; Rousseau, E. Effects of ϖ-hydroxylase product on distal human pulmonary arteries. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, 1435–1443. [Google Scholar] [CrossRef] [PubMed]
- Birks, E.K.; Bousamra, M.; Presberg, K.; Marsh, J.A.; Effros, R.M.; Jacobs, E.R. Human pulmonary arteries dilate to 20-HETE, an endogenous eicosanoid of lung tissue. Am. J. Physiol. 1997, 272, 823–829. [Google Scholar]
- Yaghi, A.; Webb, C.D.; Scott, J.A.; Mehta, S.; Bend, J.R.; McCormack, D.G. Cytochrome P450 metabolites of arachidonic acid but not cyclooxygenase-2 metabolites contribute to the pulmonary vascular hyporeactivity in rats with acute pseudomonas pneumonia. J. Pharmacol. Exp. Ther. 2001, 297, 479–488. [Google Scholar] [PubMed]
- Jacobs, E.R.; Zhu, D.; Gruenloh, S.; Lopez, B.; Medhora, M. VEGF-induced relaxation of pulmonary arteries is mediated by endothelial cytochrome P-450 hydroxylase. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 291, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Medhora, M.; Falck, J.R.; Pritchard, K.A., Jr.; Jacobs, E.R. Mechanisms of activation of eNOS by 20-HETE and VEGF in bovine pulmonary artery endothelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 291, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Kizub, I.V.; Lakhkar, A.; Dhagia, V.; Joshi, S.R.; Jiang, H.; Wolin, M.S.; Falck, J.R.; Koduru, S.R.; Errabelli, R.; Jacobs, E.R. Involvement of gap junctions between smooth muscle cells in sustained hypoxic pulmonary vasoconstriction development: A potential role for 15-HETE and 20-HETE. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 310, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.U.; Choudhury, S.; Parida, S.; Maruti, B.S.; Mishra, S.K. Arachidonic acid inhibits Na+-K+-atpase via cytochrome P-450, lipoxygenase and protein kinase c-dependent pathways in sheep pulmonary artery. Vascul. Pharmacol. 2012, 56, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Parker, T.A.; Grover, T.R.; Kinsella, J.P.; Falck, J.R.; Abman, S.H. Inhibition of 20-HETE abolishes the myogenic response during NOS antagonism in the ovine fetal pulmonary circulation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 289, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, E.; Cloutier, M.; Morin, C.; Proteau, S. Capsazepine, a vanilloid antagonist, abolishes tonic responses induced by 20-HETE on guinea pig airway smooth muscle. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 288, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.; Sirois, M.; Echave, V.; Gomes, M.M.; Rousseau, E. Functional effects of 20-HETE on human bronchi: Hyperpolarization and relaxation due to BKCa channel activation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 293, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, E.R.; Effros, R.M.; Falck, J.R.; Reddy, K.M.; Campbell, W.B.; Zhu, D. Airway synthesis of 20-hydroxyeicosatetraenoic acid: Metabolism by cyclooxygenase to a bronchodilator. Am. J. Physiol. 1999, 276, 280–288. [Google Scholar]
- Cooper, P.R.; Mesaros, A.C.; Zhang, J.; Christmas, P.; Stark, C.M.; Douaidy, K.; Mittelman, M.A.; Soberman, R.J.; Blair, I.A.; Panettieri, R.A. 20-HETE mediates ozone-induced, neutrophil-independent airway hyper-responsiveness in mice. PLoS ONE 2010, 5, 10235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokalari, B.; Koziol-White, C.; Jester, W.; Panettieri, R.A.; Haczku, A.; Jiang, Z. 20-hydroxyeicosatetraenoic acid (20-HETE) induces airway inflammation and hyperresponsiveness in mice. In Proceedings of C101. Allergic airway inflammation and hyperresponsiveness: Novel mechanisms and therapy, Washington, DC, USA, 19–24 May 2015; Am Thoracic Soc: New York, NY, USA, 2015; p. 5165. [Google Scholar]
- Chuang, S.S.; Helvig, C.; Taimi, M.; Ramshaw, H.A.; Collop, A.H.; Amad, M.; White, J.A.; Petkovich, M.; Jones, G.; Korczak, B. Cyp2u1, a novel human thymus- and brain-specific cytochrome P450, catalyzes ϖ- and (ϖ -1)-hydroxylation of fatty acids. J. Biol. Chem. 2004, 279, 6305–6314. [Google Scholar] [CrossRef] [PubMed]
- Gebremedhin, D.; Zhang, D.X.; Carver, K.A.; Rau, N.; Rarick, K.R.; Roman, R.J.; Harder, D.R. Expression of CYP 4A omega-hydroxylase and formation of 20-hydroxyeicosatetreanoic acid (20-HETE) in cultured rat brain astrocytes. Prostaglandins Other Lipid Mediat. 2016, 124, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Collins, X.H.; Harmon, S.D.; Kaduce, T.L.; Berst, K.B.; Fang, X.; Moore, S.A.; Raju, T.V.; Falck, J.R.; Weintraub, N.L.; Duester, G.; et al. Omega-oxidation of 20-hydroxyeicosatetraenoic acid (20-HETE) in cerebral microvascular smooth muscle and endothelium by alcohol dehydrogenase 4. J. Biol. Chem. 2005, 280, 33157–33164. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Meng, L.; Li, Y.; Qu, Y. Arterial carbon dioxide partial pressure influences CYP4A distribution in the rat brain. Histol. Histopathol. 2012, 27, 897–903. [Google Scholar] [PubMed]
- Liu, X.; Li, C.; Falck, J.R.; Roman, R.J.; Harder, D.R.; Koehler, R.C. Interaction of nitric oxide, 20-HETE, and eets during functional hyperemia in whisker barrel cortex. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, 619–631. [Google Scholar] [CrossRef] [PubMed]
- Pratt, P.F.; Medhora, M.; Harder, D.R. Mechanisms regulating cerebral blood flow as therapeutic targets. Curr. Opin. Investig. Drugs 2004, 5, 952–956. [Google Scholar] [PubMed]
- Gordon, G.R.; Mulligan, S.J.; MacVicar, B.A. Astrocyte control of the cerebrovasculature. Glia 2007, 55, 1214–1221. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.D.; Simpkins, A.N.; Renic, M.; Harder, D.R. Cytochrome P450 eicosanoids and cerebral vascular function. Expert Rev. Mol. Med. 2011, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Gebremedhin, D.; Lange, A.R.; Lowry, T.F.; Taheri, M.R.; Birks, E.K.; Hudetz, A.G.; Narayanan, J.; Falck, J.R.; Okamoto, H.; Roman, R.J. Production of 20-HETE and its role in autoregulation of cerebral blood flow. Circ. Res. 2000, 87, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Nakayama, T.; Sato, N.; Izumi, Y.; Kasamaki, Y.; Shindo, A.; Ohta, M.; Soma, M.; Aoi, N.; Sato, M.; et al. A haplotype of the CYP4F2 gene is associated with cerebral infarction in Japanese men. Am. J. Hypertens. 2008, 21, 1216–1223. [Google Scholar] [CrossRef] [PubMed]
- Berg, R.M. Myogenic and metabolic feedback in cerebral autoregulation: Putative involvement of arachidonic acid-dependent pathways. Med. Hypotheses 2016, 92, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Metea, M.R.; Newman, E.A. Signalling within the neurovascular unit in the mammalian retina. Exp. Physiol. 2007, 92, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Metea, M.R.; Newman, E.A. Glial cells dilate and constrict blood vessels: A mechanism of neurovascular coupling. J. Neurosci. 2006, 26, 2862–2870. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, C.; Gebremedhin, D.; Hwang, S.H.; Hammock, B.D.; Falck, J.R.; Roman, R.J.; Harder, D.R.; Koehler, R.C. Epoxyeicosatrienoic acid-dependent cerebral vasodilation evoked by metabotropic glutamate receptor activation in vivo. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Lange, A.; Gebremedhin, D.; Narayanan, J.; Harder, D. 20-Hydroxyeicosatetraenoic acid-induced vasoconstriction and inhibition of potassium current in cerebral vascular smooth muscle is dependent on activation of protein kinase c. J. Biol. Chem. 1997, 272, 27345–27352. [Google Scholar] [CrossRef] [PubMed]
- Ohata, H.; Gebremedhin, D.; Narayanan, J.; Harder, D.R.; Koehler, R.C. Onset of pulmonary ventilation in fetal sheep produces pial arteriolar constriction dependent on cytochrome P450 omega-hydroxylase activity. J. Appl. Physiol. 2010, 109, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Mapelli, L.; Gagliano, G.; Soda, T.; Laforenza, U.; Moccia, F.; D’Angelo, E.U. Granular layer neurons control cerebellar neurovascular coupling through an NMDA receptor/no-dependent system. J. Neurosci. 2017, 37, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Hama-Tomioka, K.; Kinoshita, H.; Azma, T.; Nakahata, K.; Matsuda, N.; Hatakeyama, N.; Kikuchi, H.; Hatano, Y. The role of 20-hydroxyeicosatetraenoic acid in cerebral arteriolar constriction and the inhibitory effect of propofol. Anesth. Analg. 2009, 109, 1935–1942. [Google Scholar] [CrossRef] [PubMed]
- Renic, M.; Kumar, S.N.; Gebremedhin, D.; Florence, M.A.; Gerges, N.Z.; Falck, J.R.; Harder, D.R.; Roman, R.J. Protective effect of 20-HETE inhibition in a model of oxygen-glucose deprivation in hippocampal slice cultures. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.N.; Reynell, C.; Gesslein, B.; Hamilton, N.B.; Mishra, A.; Sutherland, B.A.; O’Farrell, F.M.; Buchan, A.M.; Lauritzen, M.; Attwell, D. Capillary pericytes regulate cerebral blood flow in health and disease. Nature 2014, 508, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Faraci, F.M.; Kaduce, T.L.; Harmon, S.; Modrick, M.L.; Hu, S.; Moore, S.A.; Falck, J.R.; Weintraub, N.L.; Spector, A.A. 20-Hydroxyeicosatetraenoic acid is a potent dilator of mouse basilar artery: Role of cyclooxygenase. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, 2301–2307. [Google Scholar] [CrossRef] [PubMed]
- Marumo, T.; Eto, K.; Wake, H.; Omura, T.; Nabekura, J. The inhibitor of 20-HETE synthesis, ts-011, improves cerebral microcirculatory autoregulation impaired by middle cerebral artery occlusion in mice. Br. J. Pharmacol. 2010, 161, 1391–1402. [Google Scholar] [CrossRef] [PubMed]
- Poloyac, S.M.; Reynolds, R.B.; Yonas, H.; Kerr, M.E. Identification and quantification of the hydroxyeicosatetraenoic acids, 20-HETE and 12-HETE, in the cerebrospinal fluid after subarachnoid hemorrhage. J. Neurosci. Methods 2005, 144, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Marumo, T.; Shirakami, K.; Mori, T.; Doi, H.; Suzuki, M.; Watanabe, Y.; Chaki, S.; Nakazato, A.; Ago, Y. Increase of 20-HETE synthase after brain ischemia in rats revealed by pet study with (11)c-labeled 20-HETE synthase-specific inhibitor. J. Cereb. Blood Flow Metab. 2012, 9, 1737–1746. [Google Scholar] [CrossRef] [PubMed]
- Poloyac, S.M.; Zhang, Y.; Bies, R.R.; Kochanek, P.M.; Graham, S.H. Protective effect of the 20-HETE inhibitor het0016 on brain damage after temporary focal ischemia. J. Cereb. Blood Flow Metab. 2006, 26, 1551–1561. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Omura, T.; Fukasawa, M.; Horiuchi, N.; Miyata, N.; Minagawa, T.; Yoshida, S.; Nakaike, S. Continuous inhibition of 20-HETE synthesis by ts-011 improves neurological and functional outcomes after transient focal cerebral ischemia in rats. Neurosci. Res. 2007, 59, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.J.; Carter, E.L.; Kibler, K.K.; Kwansa, H.; Crafa, D.A.; Martin, L.J.; Roman, R.J.; Harder, D.R.; Koehler, R.C. Attenuation of neonatal ischemic brain damage using a 20-HETE synthesis inhibitor. J. Neurochem. 2012, 121, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Cambj-Sapunar, L.; Yu, M.; Harder, D.R.; Roman, R.J. Contribution of 5-hydroxytryptamine1b receptors and 20-hydroxyeiscosatetraenoic acid to fall in cerebral blood flow after subarachnoid hemorrhage. Stroke 2003, 34, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, K.; Miyata, N.; Renic, M.; Harder, D.R.; Roman, R.J. Hemoglobin, no, and 20-HETE interactions in mediating cerebral vasoconstriction following sah. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Crago, E.A.; Thampatty, B.P.; Sherwood, P.R.; Kuo, C.W.; Bender, C.; Balzer, J.; Horowitz, M.; Poloyac, S.M. Cerebrospinal fluid 20-HETE is associated with delayed cerebral ischemia and poor outcomes after aneurysmal subarachnoid hemorrhage. Stroke 2011, 42, 1872–1877. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Zhu, G.; Liu, F.; Zhang, H.; Qin, X.; Li, L.; Zhiyi, H. CYP4F2 gene V433M polymorphism is associated with ischemic stroke in the male northern chinese han population. Prog. Neuropsychopharmacol. Biol. Psychiatry 2010, 34, 664–668. [Google Scholar] [CrossRef] [PubMed]
- Renic, M.; Klaus, J.A.; Omura, T.; Kawashima, N.; Onishi, M.; Miyata, N.; Koehler, R.C.; Harder, D.R.; Roman, R.J. Effect of 20-HETE inhibition on infarct volume and cerebral blood flow after transient middle cerebral artery occlusion. J. Cereb. Blood Flow Metab. 2009, 29, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Klamerus, M.M.; Miller, T.M.; Rohan, L.C.; Graham, S.H.; Poloyac, S.M. Intravenous formulation of n-hydroxy-n′-(4-n-butyl-2-methylphenyl)formamidine (het0016) for inhibition of rat brain 20-hydroxyeicosatetraenoic acid formation. Drug Metab. Dispos. 2008, 36, 2324–2330. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, D.; Wang, H.; Qu, Y.; Xiao, X.; Zhu, Y. The protective effect of HET0016 on brain edema and blood-brain barrier dysfunction after cerebral ischemia/reperfusion. Brain Res. 2014, 1544, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Miyata, N.; Seki, T.; Tanaka, Y.; Omura, T.; Taniguchi, K.; Doi, M.; Bandou, K.; Kametani, S.; Sato, M.; Okuyama, S. Beneficial effects of a new 20-hydroxyeicosatetraenoic acid synthesis inhibitor, ts-011 [n-(3-chloro-4-morpholin-4-yl) phenyl-n′-hydroxyimido formamide], on hemorrhagic and ischemic stroke. J. Pharmacol. Exp. Ther. 2005, 314, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Omura, T.; Tanaka, Y.; Miyata, N.; Koizumi, C.; Sakurai, T.; Fukasawa, M.; Hachiuma, K.; Minagawa, T.; Susumu, T.; Yoshida, S.; et al. Effect of a new inhibitor of the synthesis of 20-HETE on cerebral ischemia reperfusion injury. Stroke 2006, 37, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Goodman, A.I.; Choudhury, M.; da Silva, J.L.; Schwartzman, M.L.; Abraham, N.G. Overexpression of the heme oxygenase gene in renal cell carcinoma. Proc. Soc. Exp. Biol. Med. 1997, 214, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Alexanian, A.; Miller, B.; Roman, R.J.; Sorokin, A. 20-HETE-producing enzymes are up-regulated in human cancers. Cancer Genomics Proteomics 2012, 9, 163–169. [Google Scholar] [PubMed]
- Chen, P.; Guo, M.; Wygle, D.; Edwards, P.A.; Falck, J.R.; Roman, R.J.; Scicli, A.G. Inhibitors of cytochrome P450 4a suppress angiogenic responses. Am. J. Pathol. 2005, 166, 615–624. [Google Scholar] [CrossRef]
- Yu, W.; Chen, L.; Yang, Y.Q.; Falck, J.R.; Guo, A.M.; Li, Y.; Yang, J. Cytochrome P450 ϖ-hydroxylase promotes angiogenesis and metastasis by upregulation of vegf and mmp-9 in non-small cell lung cancer. Cancer Chemother. Pharmacol. 2011, 68, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Alexanian, A.; Rufanova, V.A.; Miller, B.; Flasch, A.; Roman, R.J.; Sorokin, A. Down-regulation of 20-HETE synthesis and signaling inhibits renal adenocarcinoma cell proliferation and tumor growth. Anticancer Res. 2009, 29, 3819–3824. [Google Scholar] [PubMed]
- Akbulut, T.; Regner, K.R.; Roman, R.J.; Avner, E.D.; Falck, J.R.; Park, F. 20-HETE activates the RAF/MEK/ERK pathway in renal epithelial cells through an EGFR- and C-SRC-dependent mechanism. Am. J. Physiol. Renal Physiol. 2009, 297, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Rios, A.; Falck, J.R.; Belosludtsev, Y.; Schwartzman, M.L. 20-Hydroxyeicosatetraenoic acid is formed in response to EGF and is a mitogen in rat proximal tubule. Am. J. Physiol. 1995, 269, 806–816. [Google Scholar]
- Chen, J.K.; Falck, J.R.; Reddy, K.M.; Capdevila, J.; Harris, R.C. Epoxyeicosatrienoic acids and their sulfonimide derivatives stimulate tyrosine phosphorylation and induce mitogenesis in renal epithelial cells. J. Biol. Chem. 1998, 273, 29254–29261. [Google Scholar] [CrossRef] [PubMed]
- Nithipatikom, K.; Isbell, M.A.; See, W.A.; Campbell, W.B. Elevated 12- and 20-hydroxyeicosatetraenoic acid in urine of patients with prostatic diseases. Cancer Lett. 2006, 233, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Vanella, L.; Di Giacomo, C.; Acquaviva, R.; Barbagallo, I.; Li Volti, G.; Cardile, V.; Abraham, N.G.; Sorrenti, V. Effects of ellagic acid on angiogenic factors in prostate cancer cells. Cancers 2013, 5, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Chai, H.; Li, Y.; Zhao, H.; Xie, X.; Zheng, H.; Wang, C.; Wang, X.; Yang, G.; Cai, X.; et al. Increased expression of CYP4Z1 promotes tumor angiogenesis and growth in human breast cancer. Toxicol. Appl. Pharmacol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Borin, T.F.; Zuccari, D.A.; Jardim-Perassi, B.V.; Ferreira, L.C.; Iskander, A.S.; Varma, N.R.; Shankar, A.; Guo, A.M.; Scicli, G.; Arbab, A.S. Het0016, a selective inhibitor of 20-HETE synthesis, decreases pro-angiogenic factors and inhibits growth of triple negative breast cancer in mice. PLoS ONE 2014, 9, 116247. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.M.; Sheng, J.; Scicli, G.M.; Arbab, A.S.; Lehman, N.L.; Edwards, P.A.; Falck, J.R.; Roman, R.J.; Scicli, A.G. Expression of CYP4A1 in U251 human glioma cell induces hyperproliferative phenotype in vitro and rapidly growing tumors in vivo. J. Pharmacol. Exp. Ther. 2008, 327, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Roman, R.J.; Falck, J.R.; Edwards, P.A.; Scicli, A.G. Human u251 glioma cell proliferation is suppressed by het0016 [n-hydroxy-n′-(4-butyl-2-methylphenyl)formamidine], a selective inhibitor of CYP4A. J. Pharmacol. Exp. Ther. 2005, 315, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Roman, R.J.; Fenstermacher, J.D.; Brown, S.L.; Falck, J.R.; Arbab, A.S.; Edwards, P.A.; Scicli, A.G. 9l gliosarcoma cell proliferation and tumor growth in rats are suppressed by n-hydroxy-n′-(4-butyl-2-methylphenol) formamidine (het0016), a selective inhibitor of CYP4A. J. Pharmacol. Exp. Ther. 2006, 317, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Shankar, A.; Borin, T.F.; Iskander, A.; Varma, N.R.; Achyut, B.R.; Jain, M.; Mikkelsen, T.; Guo, A.M.; Chwang, W.B.; Ewing, J.R. Combination of vatalanib and a 20-HETE synthesis inhibitor results in decreased tumor growth in an animal model of human glioma. OncoTarget Ther. 2016, 9, 1205–1219. [Google Scholar]
- Zhao, H.; Li, Y.; Wang, Y.; Zhang, J.; Ouyang, X.; Peng, R.; Yang, J. Antitumor and immunostimulatory activity of a polysaccharide-protein complex from scolopendra subspinipes mutilans l: Koch in tumor-bearing mice. Food Chem. Toxicol. 2012, 50, 2648–2655. [Google Scholar] [CrossRef] [PubMed]
- Theken, K.N.; Deng, Y.; Schuck, R.N.; Oni-Orisan, A.; Miller, T.M.; Kannon, M.A.; Poloyac, S.M.; Lee, C.R. Enalapril reverses high-fat diet-induced alterations in cytochrome P450-mediated eicosanoid metabolism. Am. J. Physiol. Endocrinol. Metab. 2012, 302, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Joseph, G.; Soler, A.; Hutcheson, R.; Hunter, I.; Bradford, C.; Hutcheson, B.; Gotlinger, K.H.; Jiang, H.; Falck, J.R.; Proctor, S.; et al. Elevated 20-HETE impairs coronary collateral growth in metabolic syndrome via endothelial dysfunction. Am. J. Physiol. Heart Circ. Physiol. 2016. ajpheart 00561. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Puri, N.; Sodhi, K.; Falck, J.R.; Abraham, N.G.; Shapiro, J.; Schwartzman, M.L. Cyclooxygenase-2 dependent metabolism of 20-HETE increases adiposity and adipocyte enlargement in mesenchymal stem cell-derived adipocytes. J. Lipid Res. 2013, 54, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Yousif, M.H.; Benter, I.F.; Dunn, K.M.; Dahly-Vernon, A.J.; Akhtar, S.; Roman, R.J. Role of 20-hydroxyeicosatetraenoic acid in altering vascular reactivity in diabetes. Auton. Autacoid Pharmacol. 2009, 29, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yadav, A.S.; Leskova, W.; Harris, N.R. Inhibition of 20-HETE attenuates diabetes-induced decreases in retinal hemodynamics. Exp. Eye Res. 2011, 93, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Eid, S.; Abou-Kheir, W.; Sabra, R.; Daoud, G.; Jaffa, A.; Ziyadeh, F.N.; Roman, L.; Eid, A.A. Involvement of renal cytochromes P450 and arachidonic acid metabolites in diabetic nephropathy. J. Biol. Regul. Homeost. Agents 2013, 27, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Li, J.; Quilley, J. Deficient renal 20-HETE release in the diabetic rat is not the result of oxidative stress. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, 2305–2312. [Google Scholar] [CrossRef] [PubMed]
- Tse, M.M.; Aboutabl, M.E.; Althurwi, H.N.; Elshenawy, O.H.; Abdelhamid, G.; El-Kadi, A.O. Cytochrome P450 epoxygenase metabolite, 14,15-eet, protects against isoproterenol-induced cellular hypertrophy in h9c2 rat cell line. Vascul. Pharmacol. 2013, 5, 363–373. [Google Scholar] [CrossRef] [PubMed]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elshenawy, O.H.; Shoieb, S.M.; Mohamed, A.; El-Kadi, A.O.S. Clinical Implications of 20-Hydroxyeicosatetraenoic Acid in the Kidney, Liver, Lung and Brain: An Emerging Therapeutic Target. Pharmaceutics 2017, 9, 9. https://doi.org/10.3390/pharmaceutics9010009
Elshenawy OH, Shoieb SM, Mohamed A, El-Kadi AOS. Clinical Implications of 20-Hydroxyeicosatetraenoic Acid in the Kidney, Liver, Lung and Brain: An Emerging Therapeutic Target. Pharmaceutics. 2017; 9(1):9. https://doi.org/10.3390/pharmaceutics9010009
Chicago/Turabian StyleElshenawy, Osama H., Sherif M. Shoieb, Anwar Mohamed, and Ayman O.S. El-Kadi. 2017. "Clinical Implications of 20-Hydroxyeicosatetraenoic Acid in the Kidney, Liver, Lung and Brain: An Emerging Therapeutic Target" Pharmaceutics 9, no. 1: 9. https://doi.org/10.3390/pharmaceutics9010009