
Interaction of KLAKLAK-NH2 and Analogs with Biomimetic Membrane Models
 , ,
, ,  , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Peptide Synthesis
2.2.2. Vesicle Preparation
2.2.3. Formation of Bilayer Lipid Membranes (BLMs)
2.2.4. Thermal Shape Fluctuation Analysis (TSFA) of GUVs
2.2.5. Fast Fourier-Transform Electrochemical Impedance Spectroscopy (FFT-EIS) of Lipid Bilayers
2.2.6. Fourier-Transform Infrared Spectroscopy by Attenuated Total Reflectance (FTIR-ATR)
3. Results and Discussion
3.1. FTIR-ATR Spectra Analysis
3.1.1. Spectral Region 3600–2800 cm−1
3.1.2. Spectral Region 1750–1700 cm−1
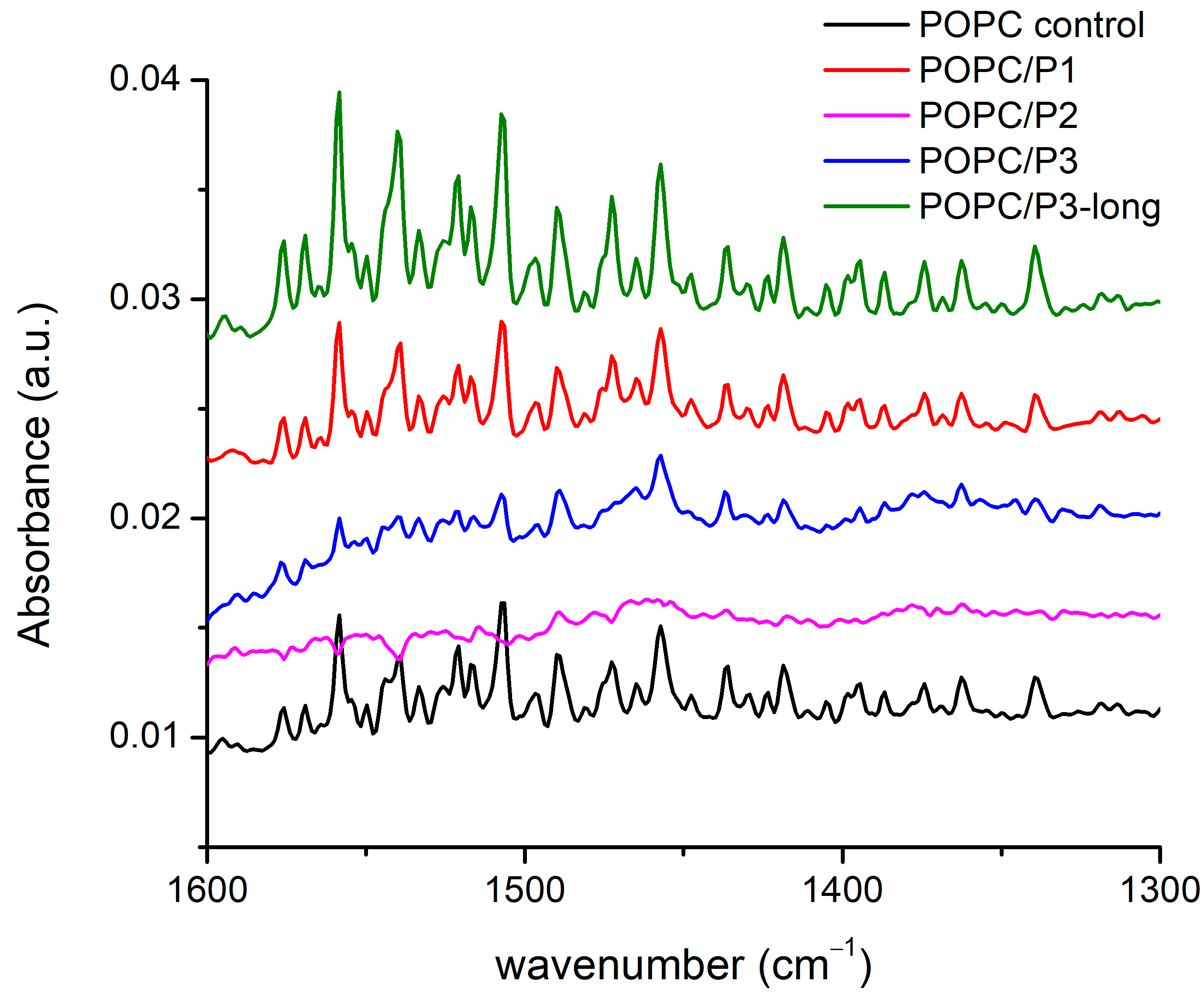
3.1.3. Spectral Region 1600–1300 cm−1
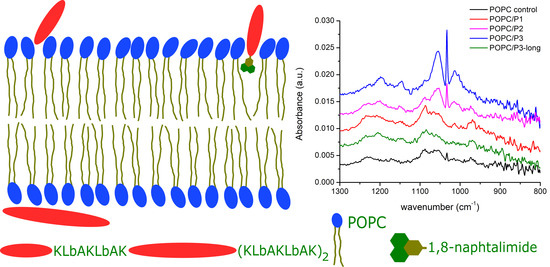
3.1.4. Spectral Region 1300–800 cm−1
3.1.5. Spectral Region 1600–1670 cm−1
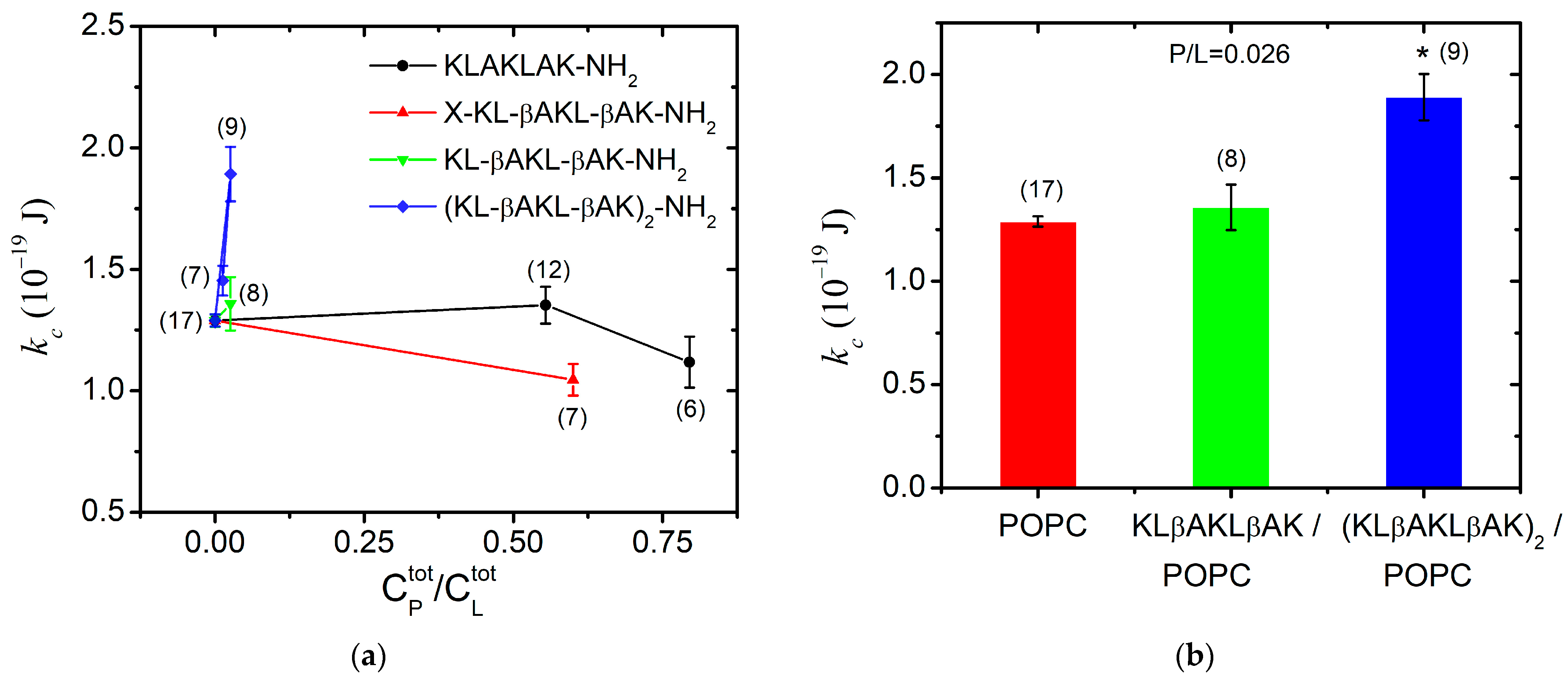
3.2. Bending Elasticity of POPC Membranes in the Presence of KLAKLAK-NH2 and Its Analogs
3.3. Impedance Characteristics of Lipid Bilayers in the Presence of KLAKLAK-NH2 and Analogues
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Frieri, M.; Kumar, K.; Boutin, A. Antibiotic resistance. J. Infect. Public Health 2017, 10, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Samreen; Ahmad, I.; Malak, H.A.; Abulreesh, H.H. Environmental antimicrobial resistance and its drivers: A potential threat to public health. J. Glob. Antimicrob. Resist. 2021, 27, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Thayer, A.M. Improving Peptides. Chem. Eng. News 2011, 89, 13–20. [Google Scholar] [CrossRef]
- Zasloff, M. Magainins, a class of antimicrobial peptides from Xenopus skin: Isolation, characterization of two active forms, and partial cDNA sequence of a precursor. Proc. Natl. Acad. Sci. USA 1987, 84, 5449–5453. [Google Scholar] [CrossRef] [PubMed]
- Boman, H.G. Peptide Antibiotics and Their Role in Innate Immunity. Annu. Rev. Immunol. 1995, 13, 61–92. [Google Scholar] [CrossRef] [PubMed]
- Boman, H.G. Innate immunity and the normal microflora. Immunol. Rev. 2000, 173, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Pushpanathan, M.; Gunasekaran, P.; Rajendhran, J. Antimicrobial Peptides: Versatile Biological Properties. Int. J. Pept. 2013, 2013, 675391. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Barra, D.; Simmaco, M. Amphibian skin: A promising resource for antimicrobial peptides. Trends Biotechnol. 1995, 13, 205–209. [Google Scholar] [CrossRef]
- Yeung, A.T.; Gellatly, S.L.; Hancock, R.E. Multifunctional cationic host defence peptides and their clinical applications. Cell. Mol. Life Sci. CMLS 2011, 68, 2161–2176. [Google Scholar] [CrossRef]
- Medina, L.; Guzmán, F.; Álvarez, C.; Delgado, J.P.; Carbonell-M, B. Ramosin: The First Antibacterial Peptide Identified on Bolitoglossa ramosi Colombian Salamander. Pharmaceutics 2022, 14, 2579. [Google Scholar] [CrossRef] [PubMed]
- Michael Conlon, J.; Mechkarska, M.; King, J.D. Host-defense peptides in skin secretions of African clawed frogs (Xenopodinae, Pipidae). Gen. Comp. Endocrinol. 2012, 176, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.E.; Howard, A.; Kasprzak, A.B.; Gordon, K.H.; East, P.D. A peptidomics study reveals the impressive antimicrobial peptide arsenal of the wax moth Galleria mellonella. Insect Biochem. Mol. Biol. 2009, 39, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E.W. Cationic peptides: Effectors in innate immunity and novel antimicrobials. Lancet Infect. Dis. 2001, 1, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of Antimicrobial Peptide Action and Resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed]
- Hale, J.D.F.; Hancock, R.E.W. Alternative mechanisms of action of cationic antimicrobial peptides on bacteria. Expert Rev. Anti-Infect. Ther. 2007, 5, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Javadpour, M.M.; Juban, M.M.; Lo, W.-C.J.; Bishop, S.M.; Alberty, J.B.; Cowell, S.M.; Becker, C.L.; McLaughlin, M.L. De Novo Antimicrobial Peptides with Low Mammalian Cell Toxicity. J. Med. Chem. 1996, 39, 3107–3113. [Google Scholar] [CrossRef] [PubMed]
- Jaber, S.; Iliev, I.; Angelova, T.; Nemska, V.; Sulikovska, I.; Naydenova, E.; Georgieva, N.; Givechev, I.; Grabchev, I.; Danalev, D. Synthesis, Antitumor and Antibacterial Studies of New Shortened Analogues of (KLAKLAK)2-NH2 and Their Conjugates Containing Unnatural Amino Acids. Molecules 2021, 26, 898. [Google Scholar] [CrossRef]
- Jaber, S.; Nemska, V.; Iliev, I.; Ivanova, E.; Foteva, T.; Georgieva, N.; Givechev, I.; Naydenova, E.; Karadjova, V.; Danalev, D. Synthesis and Biological Studies on (KLAKLAK)2-NH2 Analog Containing Unnatural Amino Acid β-Ala and Conjugates with Second Pharmacophore. Molecules 2021, 26, 7321. [Google Scholar] [CrossRef]
- Jaber, S.; Nemska, V.; Iliev, I.; Ivanova, E.; Foteva, T.; Georgieva, N.; Givechev, I.; Tanev, D.; Naydenova, E.; Danalev, D. Synthesis, antiproliferative and antimicrobial activities of (KLAKLAK)2-NH2 analogue containing nor-Leu and its conjugates with a second pharmacophore. Biotechnol. Biotechnol. Equip. 2023, 37, 151–158. [Google Scholar] [CrossRef]
- Mäe, M.; Langel, U. Cell-penetrating peptides as vectors for peptide, protein and oligonucleotide delivery. Curr. Opin. Pharmacol. 2006, 6, 509–514. [Google Scholar] [CrossRef]
- Deb, P.K.; Al-Attraqchi, O.; Chandrasekaran, B.; Paradkar, A.; Tekade, R.K. Chapter 16—Protein/Peptide Drug Delivery Systems: Practical Considerations in Pharmaceutical Product Development. In Basic Fundamentals of Drug Delivery; Tekade, R.K., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 651–684. [Google Scholar] [CrossRef]
- Marinov, M.N.; Naydenova, E.D.; Momekov, G.T.; Prodanova, R.Y.; Markova, N.V.; Voynikov, Y.T.; Stoyanov, N.M. Synthesis, Characterization, Quantum-Chemical Calculations and Cytotoxic Activity of 1,8-Naphthalimide Derivatives with Non-Protein Amino Acids. Anti-Cancer Agents Med. Chem. 2019, 19, 1276–1284. [Google Scholar] [CrossRef]
- Luisi, P.L.; Walde, P. (Eds.) Giant Vesicles; John Wiley & Sons, Ltd.: Chichester, UK, 2000. [Google Scholar]
- Dimova, R.; Aranda, S.; Bezlyepkina, N.; Nikolov, V.; Riske, K.A.; Lipowsky, R. A practical guide to giant vesicles: Probing the membrane nanoregime via optical microscopy. J. Phys. Condens. Matter 2006, 18, S1151–S1176. [Google Scholar] [CrossRef]
- Dimova, R.; Marques, C. (Eds.) The Giant Vesicle Book, 1st ed.; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar]
- Bagatolli, L.A. To see or not to see: Lateral organization of biological membranes and fluorescence microscopy. Biochim. Biophys. Acta 2006, 1758, 1541–1556. [Google Scholar] [CrossRef]
- Peng, B.; Ding, X.-Y.; Sun, C.; Yang, Y.-N.; Gao, Y.-J.; Zhao, X. The chain order of binary unsaturated lipid bilayers modulated by aromatic-residue-containing peptides: An ATR-FTIR spectroscopy study. RSC Adv. 2017, 7, 29386–29394. [Google Scholar] [CrossRef]
- Korkmaz, F.; Köster, S.; Yildiz, Ö.; Mäntele, W. The Role of Lipids for the Functional Integrity of Porin: An FTIR Study Using Lipid and Protein Reporter Groups. Biochemistry 2008, 47, 12126–12134. [Google Scholar] [CrossRef] [PubMed]
- Ergun, S.; Demir, P.; Uzbay, T.; Severcan, F. Agomelatine strongly interacts with zwitterionic DPPC and charged DPPG membranes. Biochim. Biophys. Acta (BBA) Biomembr. 2014, 1838, 2798–2806. [Google Scholar] [CrossRef] [PubMed]
- Sikorska, E.; Dawgul, M.; Greber, K.; Iłowska, E.; Pogorzelska, A.; Kamysz, W. Self-assembly and interactions of short antimicrobial cationic lipopeptides with membrane lipids: ITC, FTIR and molecular dynamics studies. Biochim. Biophys. Acta (BBA) Biomembr. 2014, 1838, 2625–2634. [Google Scholar] [CrossRef]
- Kaiser, E.; Colescott, R.L.; Bossinger, C.D.; Cook, P.I. Color test for detection of free terminal amino groups in the solid-phase synthesis of peptides. Anal. Biochem. 1970, 34, 595–598. [Google Scholar] [CrossRef] [PubMed]
- Angelova, M.; Dimitrov, D. Liposome electroformation. Faraday Discuss. Chem. Soc. 1986, 81, 303–311. [Google Scholar] [CrossRef]
- Vitkova, V.; Yordanova, V.; Staneva, G.; Petkov, O.; Stoyanova-Ivanova, A.; Antonova, K.; Popkirov, G. Dielectric Properties of Phosphatidylcholine Membranes and the Effect of Sugars. Membranes 2021, 11, 847. [Google Scholar] [CrossRef]
- Szoka, F., Jr.; Papahadjopoulos, D. Comparative properties and methods of preparation of lipid vesicles (liposomes). Annu. Rev. Biophys. Bioeng. 1980, 9, 467–508. [Google Scholar] [CrossRef]
- Lasic, D.D. The mechanism of vesicle formation. Biochem. J. 1988, 256, 1–11. [Google Scholar] [CrossRef]
- Akbarzadeh, A.; Rezaei-Sadabady, R.; Davaran, S.; Joo, S.W.; Zarghami, N.; Hanifehpour, Y.; Samiei, M.; Kouhi, M.; Nejati-Koshki, K. Liposome: Classification, preparation, and applications. Nanoscale Res. Lett. 2013, 8, 102. [Google Scholar] [CrossRef]
- Gutsmann, T.; Heimburg, T.; Keyser, U.; Mahendran, K.R.; Winterhalter, M. Protein reconstitution into freestanding planar lipid membranes for electrophysiological characterization. Nat. Protoc. 2015, 10, 188–198. [Google Scholar] [CrossRef]
- Montal, M.; Mueller, P. Formation of Bimolecular Membranes from Lipid Monolayers and a Study of Their Electrical Properties. Proc. Natl. Acad. Sci. USA 1972, 69, 3561–3566. [Google Scholar] [CrossRef]
- Helfrich, W. Elastic properties of lipid bilayers: Theory and possible experiments. Z. Naturforsch. 1973, 28, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Genova, J.; Vitkova, V.; Bivas, I. Registration and analysis of the shape fluctuations of nearly spherical lipid vesicles. Phys. Rev. E 2013, 88, 022707. [Google Scholar] [CrossRef] [PubMed]
- Bivas, I.; Hanusse, P.; Bothorel, P.; Lalanne, J.; Aguerre-Chariol, O. An application of the optical microscopy to the determination of the curvature elastic modulus of biological and model membranes. J. Phys. 1987, 48, 855–867. [Google Scholar] [CrossRef]
- Popkirov, G.S.; Schindler, R.N. Validation of experimental data in electrochemical impedance spectroscopy. Electrochim. Acta 1993, 38, 861–867. [Google Scholar] [CrossRef]
- Popkirov, G.S.; Schindler, R.N. A new approach to the problem of “good” and “bad” impedance data in electrochemical impedance spectroscopy. Electrochim. Acta 1994, 39, 2025–2030. [Google Scholar] [CrossRef]
- Casal, H.L.; Mantsch, H.H. Polymorphic phase behaviour of phospholipid membranes studied by infrared spectroscopy. Biochim. Biophys. Acta (BBA) Rev. Biomembr. 1984, 779, 381–401. [Google Scholar] [CrossRef]
- Güler, G.; Gärtner, R.M.; Ziegler, C.; Mäntele, W. Lipid-Protein Interactions in the Regulated Betaine Symporter BetP Probed by Infrared Spectroscopy. J. Biol. Chem. 2016, 291, 4295–4307. [Google Scholar] [CrossRef] [PubMed]
- Arrondo, J.L.R.; Goñi, F.M.; Macarulla, J.M. Infrared spectroscopy of phosphatidylcholines in aqueous suspension a study of the phosphate group vibrations. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1984, 794, 165–168. [Google Scholar] [CrossRef]
- Nagata, Y.; Mukamel, S. Vibrational Sum-Frequency Generation Spectroscopy at the Water/Lipid Interface: Molecular Dynamics Simulation Study. J. Am. Chem. Soc. 2010, 132, 6434–6442. [Google Scholar] [CrossRef]
- Mondal, J.A.; Nihonyanagi, S.; Yamaguchi, S.; Tahara, T. Three Distinct Water Structures at a Zwitterionic Lipid/Water Interface Revealed by Heterodyne-Detected Vibrational Sum Frequency Generation. J. Am. Chem. Soc. 2012, 134, 7842–7850. [Google Scholar] [CrossRef] [PubMed]
- Ohto, T.; Backus, E.H.G.; Hsieh, C.-S.; Sulpizi, M.; Bonn, M.; Nagata, Y. Lipid Carbonyl Groups Terminate the Hydrogen Bond Network of Membrane-Bound Water. J. Phys. Chem. Lett. 2015, 6, 4499–4503. [Google Scholar] [CrossRef]
- Fringeli, U.P.; Günthard, H.H. Infrared Membrane Spectroscopy. In Membrane Spectroscopy; Grell, E., Ed.; Springer: Berlin/Heidelberg, Germany, 1981; pp. 270–332. [Google Scholar] [CrossRef]
- Schultz, Z.D.; Levin, I.W. Vibrational Spectroscopy of Biomembranes. Annu. Rev. Anal. Chem. 2011, 4, 343–366. [Google Scholar] [CrossRef]
- Mantsch, H.H.; McElhaney, R.N. Phospholipid phase transitions in model and biological membranes as studied by infrared spectroscopy. Chem. Phys. Lipids 1991, 57, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.N.A.H.; McElhaney, R.N. Membrane lipid phase transitions and phase organization studied by Fourier transform infrared spectroscopy. Biochim. Biophys. Acta (BBA) Biomembr. 2013, 1828, 2347–2358. [Google Scholar] [CrossRef]
- Mushayakarara, E.; Levin, I.W. Determination of acyl chain conformation at the lipid interface region: Raman spectroscopic study of the carbonyl stretching mode region of dipalmitoyl phosphatidylcholine and structurally related molecules. J. Phys. Chem. 1982, 86, 2324–2327. [Google Scholar] [CrossRef]
- Fox, C.B.; Harris, J.M. Confocal Raman microscopy for simultaneous monitoring of partitioning and disordering of tricyclic antidepressants in phospholipid vesicle membranes. J. Raman Spectrosc. 2010, 41, 498–507. [Google Scholar] [CrossRef]
- Lohner, K. Effects of small organic molecules on phospholidpid phase transitions. Chem. Phys. Lipids 1991, 57, 341–362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-P.; Lewis, R.N.A.H.; Hodges, R.S.; McElhaney, R.N. Interaction of a Peptide Model of a Hydrophobic Transmembrane alpha-Helical Segment of a Membrane Protein with Phosphatidylethanolamine Bilayers: Differential Scanning Calorimetric and Fourier Transform Infrared Spectroscopic Studies. Biophys. J. 1995, 68, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Deo, N.; Somasundaran, T.; Somasundaran, P. Solution properties of amitriptyline and its partitioning into lipid bilayers. Colloids Surf. B Biointerfaces 2004, 34, 155–159. [Google Scholar] [CrossRef]
- Fišar, Z.; Fuksová, K.; Velenovská, M. Binding of Imipramine to Phospholipid Bilayers Using Radioligand Binding Assay. Gen. Physiol. Biophys. 2004, 23, 77–99. [Google Scholar]
- Qiao, Z.-Y.; Hou, C.-Y.; Zhang, D.; Liu, Y.; Lin, Y.-X.; An, H.-W.; Li, X.-J.; Wang, H. Self-assembly of cytotoxic peptide conjugated poly(β-amino ester)s for synergistic cancer chemotherapy. J. Mater. Chem. B 2015, 3, 2943–2953. [Google Scholar] [CrossRef]
- Strandberg, E.; Tiltak, D.; Ehni, S.; Wadhwani, P.; Ulrich, A.S. Lipid shape is a key factor for membrane interactions of amphipathic helical peptides. Biochim. Biophys. Acta (BBA) Biomembr. 2012, 1818, 1764–1776. [Google Scholar] [CrossRef]
- Lee, J.; Oh, E.-T.; Lee, H.-J.; Lee, E.; Kim, H.G.; Park, H.J.; Kim, C. Tuning of Peptide Cytotoxicity with Cell Penetrating Motif Activatable by Matrix Metalloproteinase-2. ACS Omega 2022, 7, 29684–29691. [Google Scholar] [CrossRef]
- Malina, A.; Shai, Y. Conjugation of fatty acids with different lengths modulates the antibacterial and antifungal activity of a cationic biologically inactive peptide. Biochem. J. 2005, 390, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Vitkova, V.; Misbah, C. Dynamics of lipid vesicles—From thermal fluctuations to rheology. In Advances in Planar Lipid Bilayers and Liposomes; Iglic, A., Ed.; Academic Press: Burlington, NJ, USA, 2011; Volume 14, pp. 258–292. [Google Scholar]
- Dreier, L.B.; Wolde-Kidan, A.; Bonthuis, D.J.; Netz, R.R.; Backus, E.H.G.; Bonn, M. Unraveling the Origin of the Apparent Charge of Zwitterionic Lipid Layers. J. Phys. Chem. Lett. 2019, 10, 6355–6359. [Google Scholar] [CrossRef]
- Vitkova, V.; Meleard, P.; Pott, T.; Bivas, I. Alamethicin influence on the membrane bending elasticity. Eur. Biophys. J. 2006, 35, 281–286. [Google Scholar] [CrossRef]
- Vitkova, V.; Stoyanova-Ivanova, A.; Jaber, S.; Naydenova, E.; Danalev, D. Bending Elasticity of Phospholipid Bilayers Containing an Amphipathic Peptide with Low Mammalian Cytotoxicity. Proc. Bulg. Acad. Sci. 2022, 75, 1428–1436. [Google Scholar] [CrossRef]
- Ben-Shaul, A.; Ben-Tal, N.; Honig, B. Statistical thermodynamic analysis of peptide and protein insertion into lipid membranes. Biophys. J. 1996, 71, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Kondrashov, O.V.; Akimov, S.A. Regulation of Antimicrobial Peptide Activity via Tuning Deformation Fields by Membrane-Deforming Inclusions. Int. J. Mol. Sci. 2022, 23, 326. [Google Scholar] [CrossRef] [PubMed]
- Kondrashov, O.V.; Rokitskaya, T.I.; Batishchev, O.V.; Kotova, E.A.; Antonenko, Y.N.; Akimov, S.A. Peptide-induced membrane elastic deformations decelerate gramicidin dimer-monomer equilibration. Biophys. J. 2021, 120, 5309–5321. [Google Scholar] [CrossRef] [PubMed]
- Bivas, I.; Meleard, P. Bending elasticity of a lipid bilayer containing an additive. Phys. Rev. E 2003, 67, 012901. [Google Scholar] [CrossRef] [PubMed]
- Shchelokovskyy, P.; Tristram-Nagle, S.; Dimova, R. Effect of the HIV-1 fusion peptide on the mechanical properties and leaflet coupling of lipid bilayers. New J. Phys. 2011, 13, 025004. [Google Scholar] [CrossRef] [PubMed]
- Dimova, R. Recent developments in the field of bending rigidity measurements on membranes. Adv. Colloid Interface Sci. 2014, 208, 225–234. [Google Scholar] [CrossRef]
- Dimova, R. Giant Vesicles and Their Use in Assays for Assessing Membrane Phase State, Curvature, Mechanics, and Electrical Properties. Annu. Rev. Biophys. 2019, 48, 93–119. [Google Scholar] [CrossRef]
- Naumowicz, M.; Zając, M.; Kusaczuk, M.; Gál, M.; Kotyńska, J. Electrophoretic Light Scattering and Electrochemical Impedance Spectroscopy Studies of Lipid Bilayers Modified by Cinnamic Acid and Its Hydroxyl Derivatives. Membranes 2020, 10, 343. [Google Scholar] [CrossRef] [PubMed]
- Velikonja, A.; Kramar, P.; Miklavčič, D.; Maček Lebar, A. Specific electrical capacitance and voltage breakdown as a function of temperature for different planar lipid bilayers. Bioelectrochemistry 2016, 112, 132–137. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Structure | Molecular Mass (g/mol) |
|---|---|---|
| P1 | KLAKLAK-NH2 | 769.52 |
| P2 | * NphtG-KLβAKLβAK-NH2 | 1006.56 |
| P3 | KLβAKLβAK-NH2 | 769.52 |
| P3-long | KLβAKLβAKKLβAKLβAK-NH2 | 1522.05 |
| Sample | POPC | POPC/P1 | POPC/P2 | POPC/P3 | POPC/P3-Long | |
|---|---|---|---|---|---|---|
| Parameter | ||||||
| Peak wavenumber, cm−1 | 2922 asym CH2 | 2922 asym CH2 | 2922 + 2954 asym CH2 + CH3 | 2922 + 2954 asym CH2 + CH3 | 2922 asym CH2 | |
| Half-width, cm−1 | 19 | 20 | 31 | 30 | 24 | |
| Peak wavenumber, cm−1 | 2853 sym CH2 | 2853 sym CH2 | 2853 + 2865 sym CH2 + CH3 | 2853 + 2865 sym CH2 + CH3 | 2853 sym CH2 | |
| Half-width, cm−1 | 16 | 13 | 17 (24) | 19 (30) | 14 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vitkova, V.; Antonova, K.; Petkov, O.; Stoyanova-Ivanova, A.; Jaber, S.; Ivanova, V.; Naydenova, E.; Danalev, D. Interaction of KLAKLAK-NH2 and Analogs with Biomimetic Membrane Models. Pharmaceutics 2024, 16, 340. https://doi.org/10.3390/pharmaceutics16030340
Vitkova V, Antonova K, Petkov O, Stoyanova-Ivanova A, Jaber S, Ivanova V, Naydenova E, Danalev D. Interaction of KLAKLAK-NH2 and Analogs with Biomimetic Membrane Models. Pharmaceutics. 2024; 16(3):340. https://doi.org/10.3390/pharmaceutics16030340
Chicago/Turabian StyleVitkova, Victoria, Krassimira Antonova, Ognyan Petkov, Angelina Stoyanova-Ivanova, Sirine Jaber, Vladislava Ivanova, Emilia Naydenova, and Dancho Danalev. 2024. "Interaction of KLAKLAK-NH2 and Analogs with Biomimetic Membrane Models" Pharmaceutics 16, no. 3: 340. https://doi.org/10.3390/pharmaceutics16030340