
Advances in Antitumor Effects Using Liposomal Citrinin in Induced Breast Cancer Model
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Reagents
2.2. Liposomal Citrinin (LP-CIT)
2.3. Animals and Treated Groups
2.4. Breast Cancer Induced by 7,12-Dimethylbenzanthracene (DMBA)
2.5. Toxicological Monitoring during Breast Cancer Induction and Therapy
2.5.1. Hippocratic Analysis
2.5.2. Open Field
2.5.3. Rota Rod
2.5.4. Biochemical and Hematological Analysis
2.5.5. Euthanasia and Organ Collection
2.5.6. Histopathological Evaluation
2.6. Genotoxic Damage, DNA Repair, and Mutagenic Profile in Tumor and Healthy Cells
2.6.1. Comet Alkaline Assay
2.6.2. Micronucleus Test and Erythrocyte Evaluation
2.7. Statistical Analysis
3. Results
3.1. Toxicological Evaluation
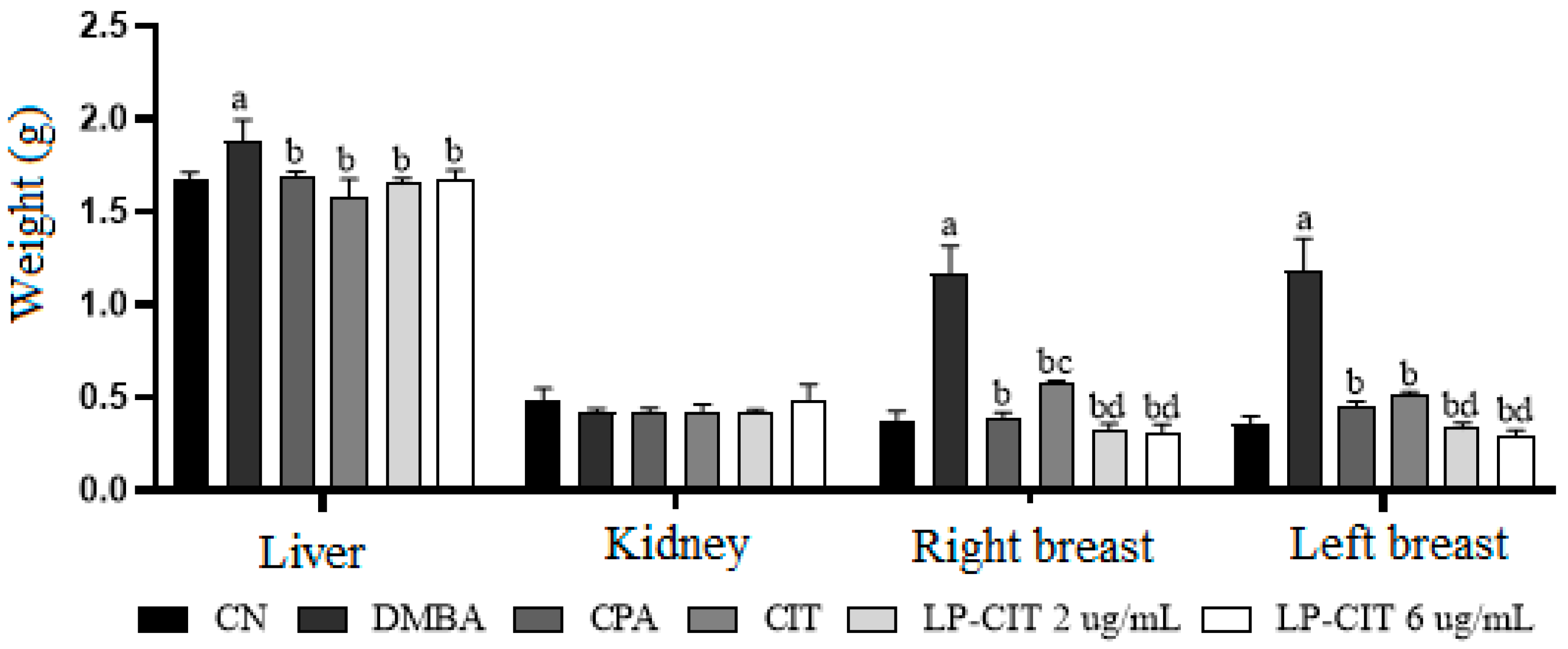
3.1.1. Hippocratic, Behavioral, and Organ Weight Monitoring
3.1.2. Biochemical and Hematological Profile
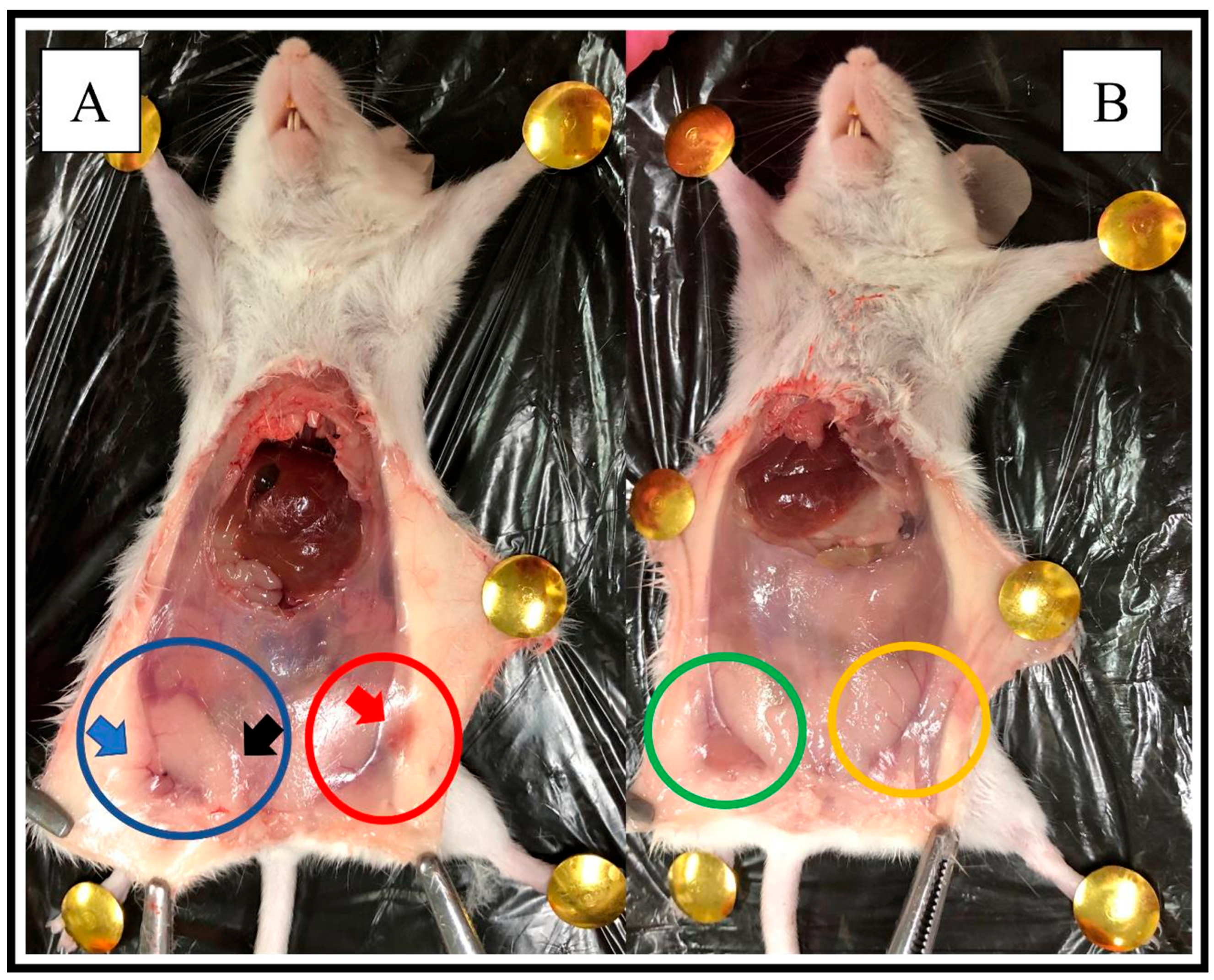
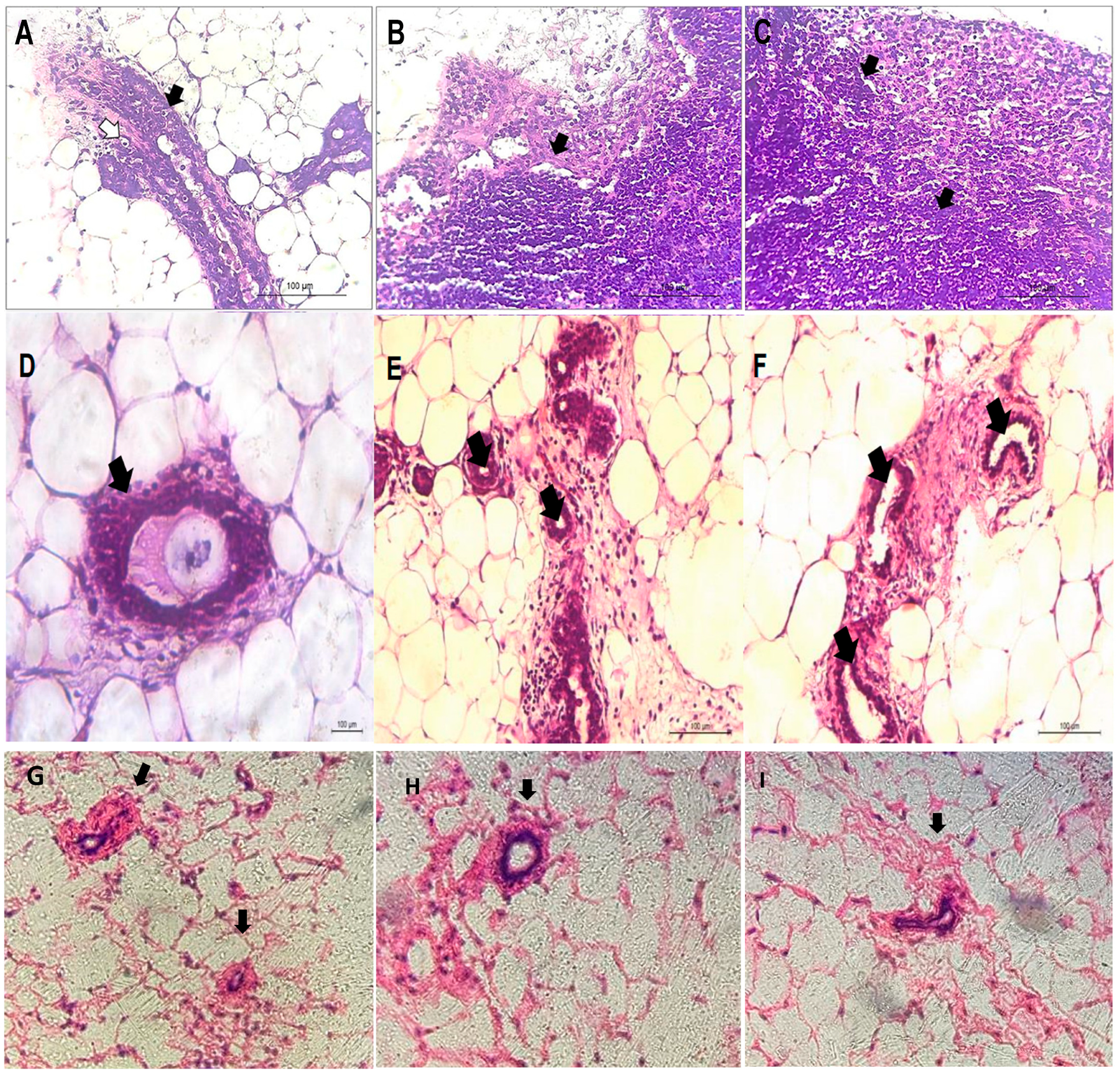
3.1.3. Macroscopic and Histopathological Analysis in Mice with DMBA-Induced Breast Cancer
3.2. Toxicogenetic Profile of DMBA, CPA, CIT, and LP-CIT
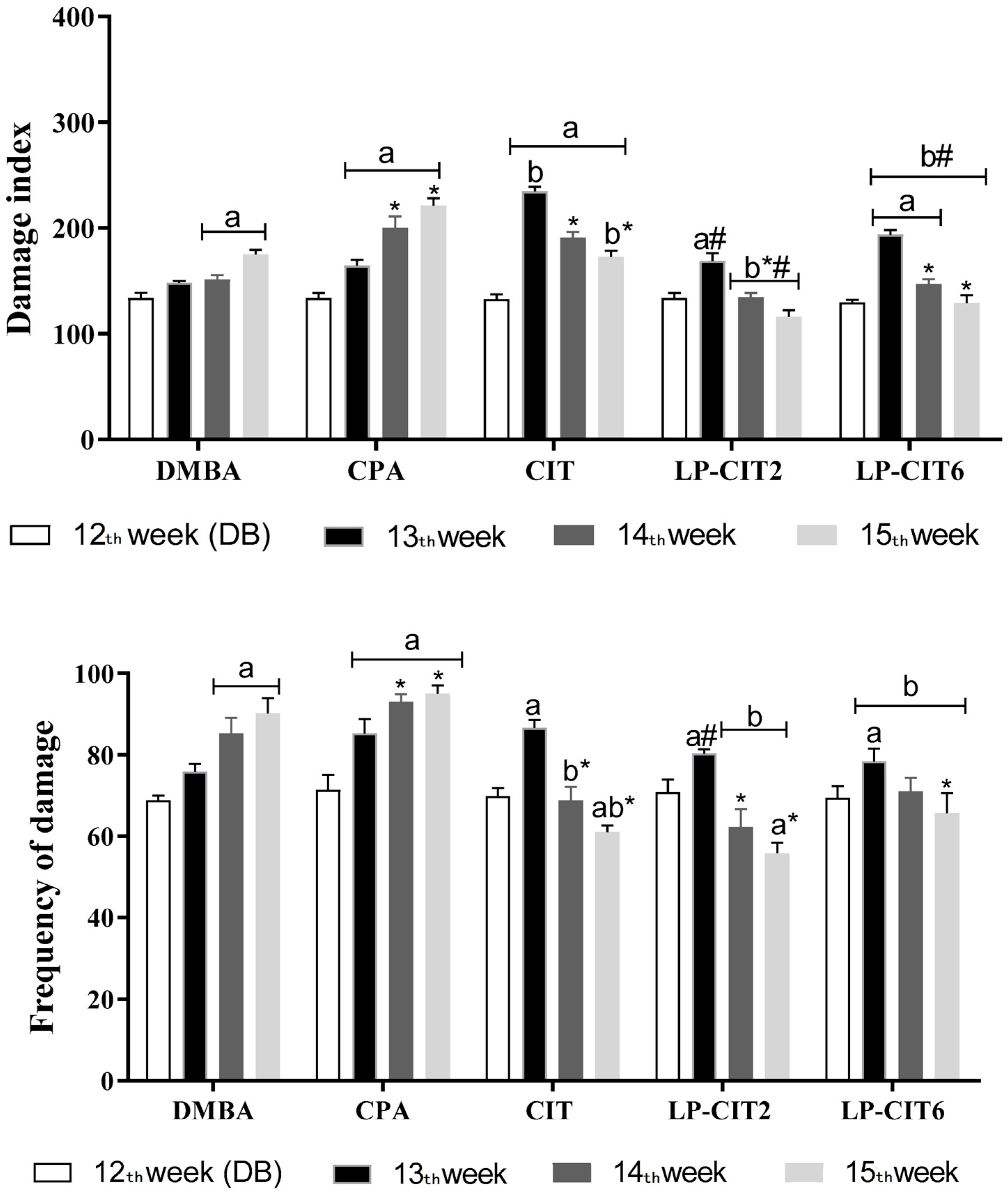
3.2.1. Genotoxicity Induced by DMBA, CPA, CIT, and LP-CIT in Tumor Tissues and Healthy Cells of Female Mice
3.2.2. Evaluation of DNA Damage Repair Capacity in Peripheral Blood Cells during and after Treatment with DMBA and Therapy with CPA, CIT, and LP-CIT
3.2.3. Cytotoxicity and Mutagenicity of CPA, CIT, and Nanoformulations in Breast, Liver, and Bone Marrow
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Montemurro, F.; Nuzzolese, I.; Ponzone, R. Neoadjuvant or adjuvant chemotherapy in early breast cancer? Expert Opin. Pharmacother. 2020, 21, 1071–1082. [Google Scholar] [CrossRef] [PubMed]
- Yeo, S.K.; Guan, J.L. Breast Cancer: Multiple Subtypes within a Tumor? Trends Cancer 2017, 3, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Tsang, J.Y.S.; Tse, G.M. Molecular Classification of Breast Cancer. Adv. Anat. Pathol. 2020, 27, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Wang, H.; Zhang, M. Nanoparticles for imaging and treatment of metastatic breast cancer. Expert. Opin. Drug Deliv. 2017, 14, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Mundhe, K.; Jain, V.; Pruthi, G.; Shah, N. Clinical study to evaluate the wear of natural enamel antagonist to zirconia and metal ceramic crowns. J. Prosthet. Dent. 2015, 114, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, S.; Meng, X.; Gan, R.Y.; Zhang, J.J.; Li, H.B. Dietary Natural Products for Prevention and Treatment of Breast Cancer. Nutrients 2017, 9, 728. [Google Scholar] [CrossRef] [PubMed]
- Haider, M.; Abdin, S.M.; Kamal, L.; Orive, G. Nanostructured Lipid Carriers for Delivery of Chemotherapeutics: A Review. Pharmaceutics 2020, 12, 288. [Google Scholar] [CrossRef]
- Barzaman, K.; Karami, J.; Zarei, Z.; Hosseinzadeh, A.; Kazemi, M.H.; Moradi-Kalbolandi, S.; Safari, E.; Farahmand, L. Breast cancer: Biology, biomarkers, and treatments. Int. Immunopharmacol. 2020, 84, 106535. [Google Scholar] [CrossRef]
- García-Pinel, B.; Porras-Alcalá, C.; Ortega-Rodríguez, A.; Sarabia, F.; Prados, J.; Melguizo, C.; López-Romero, J.M. Lipid-Based Nanoparticles: Application and Recent Advances in Cancer Treatment. Nanomaterials 2019, 9, 638. [Google Scholar] [CrossRef]
- Zhang, J.; Du, Q.; Song, X.; Gao, S.; Pang, X.; Li, Y.; Zhang, R.; Abliz, Z.; He, J. Evaluation of the tumor-targeting efficiency and intratumor heterogeneity of anticancer drugs using quantitative mass spectrometry imaging. Theranostics 2020, 10, 2621–2630. [Google Scholar] [CrossRef]
- Doi, K.; Uetshuka, K. Mechanisms of Mycotoxin-induced Dermal Toxicity and Tumorigenesis Throung Oxidative Stress-related Pathways. J. Toxicol. Pathol. 2014, 27, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Ho, C.C.; Chen, C.H.D.; Wang, W.C.; Ding, S.J. In Vitro Bioactivity and Biocompatibility of Dicalcium Silicate Cements for Endodontic Use. J. Endod. 2009, 35, 1554–1557. [Google Scholar] [CrossRef] [PubMed]
- Klarić, S.M.; Medić, N.; Hulina, A.; Zanić Grubišić, T.; Rumora, L. Disturbed Hsp70 and Hsp27 expression and thiol redox status in porcine kidney PK15 cells provoked by individual and combined ochratoxin A and citrinin treatments. Food Chem. Toxicol. 2014, 71, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Su, M.; Song, S.J.; Jung, J.H. Marine-Derived Penicillium Species as Producers of Cytotoxic Metabolites. Mar. Drugs 2017, 15, 329. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Mishra, S.K.; Tripathi, S.; de Alencar, M.V.O.B.; de Castro e Sousa, J.M.; Rolim, H.M.L.; de Medeiros, M.D.G.F.; Ferreira, P.M.P.; Rouf, R.; Uddin, S.J. Mycotoxin-assisted mitochondrial dysfunction and cytotoxicity: Unexploited tools against proliferative disorders. IUBMB Life 2018, 70, 1084–1092. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.Y.; Liao, Y.C.; Chang, C.H.; Liu, B.H. Citrinin induces apoptosis in HL-60 cells via activation of the mitochondrial pathway. Toxicol. Lett. 2006, 161, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Amselem, S.; Cohen, R.; Barenholz, Y. In vitro tests to predict in vivo performance of liposomal dosage forms. Chem. Phys. Lipids 1993, 64, 219–237. [Google Scholar] [CrossRef]
- de Oliveira Filho, J.W.G.; de Jesus Aguiar dos Santos Andrade, T.; de Lima, R.M.T.; Reis, A.C.D.; Silva, D.H.S.; de Oliveira Santos, J.V.; de Menezes, A.-A.P.M.; da Mata, A.M.O.; Dias, A.C.S.; de Alencar, M.V.O.B.; et al. Citrinin against breast cancer: A cytogenotoxicological study. Phytother. Res. 2021, 35, 504–516. [Google Scholar] [CrossRef]
- de Alencar, M.V.O.B.; Islam, M.T.; de Lima, R.M.T.; Paz, M.F.C.J.; Reis, A.C.D.; da Mata, A.M.O.F.; de Oliveira Filho, J.W.G.; Cerqueira, G.S.; Ferreira, P.M.P.; de Castro e Sousa, J.M.; et al. Phytol as an anticarcinogenic and antitumoral agent: An in vivo study in swiss mice with DMBA-Induced breast cancer. IUBMB Life 2019, 71, 200–212. [Google Scholar] [CrossRef]
- Barros, A.C.S.D.; Muranaka, E.N.K.; Mori, L.J.; Pelizon, C.H.T.; Iriy, K.; Giocondo, G.; Pinotti, J.A. Induction of experimental mammary carcinogenesis in rats with 7,12-dimethylbenz(a)anthracene. Rev. Hosp. Clínicas Fac. Med. São Paulo 2004, 59, 257–261. [Google Scholar] [CrossRef]
- Archer, J. Tests for emotionality in rats and mice: A review. Anim. Behav. 1973, 21, 205–235. [Google Scholar] [CrossRef] [PubMed]
- Machado, K.C.; Oliveira, G.L.S.; Machado, K.C.; Islam, M.T.; Gomes-Junior, A.L.; Sousa, D.P.; Freitas, R.M. Anticonvulsant and behavioral effects observed in mice following treatment with an ester derivative of ferulic acid: Isopentyl ferulate. Chem.-Biol. Interact. 2015, 42, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Behmer, O.A.; Tolosa, E.M. Manual de Técnicas para Histologia Normal e Patológica, 2nd ed.; Manole: São Paulo, Brazil, 2003; 331p. [Google Scholar]
- Elgengaihi, S.; Mossa, A.T.; Refaie, A.A.; Aboubaker, D. Hepatoprotective Efficacy of Cichorium intybus L. Extract against Carbon. Tetrachloride-induced Liver Damage in Rats. J. Diet. Suppl. 2016, 13, 570–584. [Google Scholar] [PubMed]
- Speit, G.; Hartmann, A. The comet assay: A sensitive genotoxicity test for the detection of DNA damage and repair. Methods Mol. Biol. 2006, 314, 275–286. [Google Scholar]
- Moraes, G.P.; Alencar, M.V.; Islam, T.; Araújo, L.D.; Sobral, A.L.; Machado, K.D.; Aguiar, R.P.; Gomes Júnior, A.L.; Corrêa, D.; Paz, M.F.; et al. Cytogenotoxic and oxidative status evaluation of Morinda citrifolia. Int. Arch. Med. 2016, 9, 1–13. [Google Scholar]
- Li, K.; Du, Y.; Li, L.; Wei, D.Q. Bioinformatics Approaches for Anti-cancer Drug Discovery. Curr. Drug Targets 2020, 21, 3–17. [Google Scholar] [CrossRef]
- Davison, C.A.; Durbin, S.M.; Thau, M.R.; Zellmer, V.R.; Chapman, S.E.; Diener, J.; Wathen, C.; Leevy, W.M.; Schafer, Z.T. Antioxidant enzymes mediate survival of breast cancer cells deprived of extracellular matrix. Cancer Res. 2013, 73, 3704–3715. [Google Scholar] [CrossRef]
- Sancho-Garnier, H.; Colonna, M. Épidémiologie des cancers du sein [Breast cancer epidemiology]. Presse Med. 2019, 48, 1076–1084. [Google Scholar] [CrossRef]
- Vurusaner, B.; Poli, G.; Basaga, H. Tumor suppressor genes and ROS: Complex networks of interactions. Free Radic. Biol. Med. 2012, 52, 7–18. [Google Scholar] [CrossRef]
- Semenza, G.L. Hypoxia-inducible factors: Coupling glucose metabolism and redox regulation with induction of the breast cancer stem cell phenotype. EMBO J. 2017, 36, 252–259. [Google Scholar] [CrossRef]
- Pantavos, A.; Ruiter, R.; Feskens, E.F.; de Keyser, C.E.; Hofman, A.; Stricker, B.H.; Franco, O.H.; Kiefte-de Jong, J.C. Total dietary antioxidant capacity, individual antioxidant intake and breast cancer risk: The Rotterdam Study. Int. J. Cancer 2015, 136, 2178–2186. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Xing, X.; Wang, X.; Wu, D.; Wu, W.; Guo, J.; Mitragotri, S. Nanocarrier-mediated cytosolic delivery of biopharmaceuticals. Adv. Funct. Mater. 2014, 30, 1910566. [Google Scholar] [CrossRef]
- Ozdemir, I.; Selamoglu, Z.; Ates, B.; Gok, Y.; Yilmaz, I. Modulation of DMBA-induced biochemical changes by organoselenium compounds in blood of rats. Indian J. Biochem. Biophys. 2007, 44, 257–259. [Google Scholar] [PubMed]
- Alvarado, A.; Faustino-Rocha, A.I.; Colaco, B.; Oliveira, P.A. Experimental mammary carcinogenesis-Rat models. Life Sci. 2017, 173, 116–134. [Google Scholar] [CrossRef] [PubMed]
- Salamone, S.; Colin, C.; Grillier-Vuissoz, I.; Kuntz, S.; Mazerbourg, S.; Flament, S.; Martin, H.; Richert, L.; Chapleur, Y.; Boisbrun, M. Synthesis of new troglitazone derivatives: Anti-proliferative activity in breast cancer cell lines and preliminary toxicological study. Eur. J. Med. Chem. 2012, 51, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.J.; Qi, M.; Li, N.; Lei, Y.H.; Zhang, D.M.; Chen, J.X. Natural products and their derivatives: Promising modulators of tumor immunotherapy. J. Leukoc. Biol. 2020, 108, 493–508. [Google Scholar] [CrossRef]
- Ribeiro, S.M.; Chagas, G.M.; Campello, A.P.; Klüppel, M.L. Mechanism of citrinin-induced dysfunction of mitochondria. V. Effect on the homeostasis of the reactive oxygen species. Cell Biochem. Funct. 1997, 15, 203–209. [Google Scholar] [CrossRef]
- Abdal, D.A.; Choi, H.Y.; Yang, G.M.; Kim, K.; Saha, S.K.; Cho, S.G. The Anti-Cancer Effect of Polyphenols against Breast Cancer and Cancer Stem Cells: Molecular Mechanisms. Nutrients 2016, 8, 581. [Google Scholar] [CrossRef]
- Sharath Babu, G.R.; Anand, T.; Ilaiyaraja, N.; Khanum, F.; Gopalan, N. Pelargonidin Modulates Keap1/Nrf2 Pathway Gene Expression and Ameliorates Citrinin-Induced Oxidative Stress in HepG2 Cells. Front. Pharmacol. 2017, 8, 868. [Google Scholar] [CrossRef]
- Gayathri, L.; Karthikeyan, B.S.; Rajalakshmi, M.; Dhanasekaran, D.; Li, A.P.; Akbarsha, M.A. Metabolism-dependent cytotoxicity of citrinin and ochratoxin A alone and in combination as assessed adopting integrated discrete multiple organ co-culture (IdMOC). Toxicol. In Vitro 2018, 46, 166–177. [Google Scholar] [CrossRef]
- Piccart, M.; Procter, M.; Fumagalli, D.; de Azambuja, E.; Clark, E.; Ewer, M.S.; Restuccia, E.; Jerusalem, G.; Dent, S.; Reaby, L.; et al. Adjuvant Pertuzumab and Trastuzumab in Early HER2-Positive Breast Cancer in the APHINITY Trial: 6 Years’ Follow-Up. J. Clin. Oncol. 2021, 39, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Magar, K.T.; Boafo, G.F.; Li, X.; Chen, Z.; He, W. Liposome-based delivery of biological drugs. Chin. Chem. Lett. 2022, 33, 587–596. [Google Scholar] [CrossRef]
- Hu, M.L. Dietary polyphenols as antioxidants and anticancer agents: More questions than answers. Chang. Gung Med. J. 2011, 34, 449–460. [Google Scholar] [PubMed]
- Li, W.; Guo, L.; Bi, X.; Ma, J.; Zheng, S. Immunohistochemistry of p53 and Ki-67 and p53 mutation analysis in renal epithelioid angiomyolipoma. Int. J. Clin. Exp. Pathol. 2015, 8, 9446–9451. [Google Scholar]
- Thorat, M.A.; Balasubramanian, R. Breast cancer prevention in high-risk women. Best. Pract. Res. Clin. Obstet. Gynaecol. 2020, 65, 18–31. [Google Scholar] [CrossRef]
- Bouslimi, A.; Ouannes, Z.; Golli, E.E.; Bouaziz, C.; Hassen, W.; Bacha, H. Citotoxicity and oxidative damage in kidney cells exposed to the mycotoxins ochratoxin and citrinin: Individual and combined effects. Toxicol. Mech. Methods 2008, 18, 341–349. [Google Scholar] [CrossRef]
- Sun, T.; Gao, J.; Han, D.; Shi, H.; Liu, X. Fabrication and characterization of solid lipid nano-formulation of astraxanthin against DMBA-induced breast cancer via Nrf-2-Keap1 and NF-kB and mTOR/Maf-1/PTEN pathway. Drug Deliv. 2019, 26, 975–988. [Google Scholar] [CrossRef]
- Chang, C.H.; Yu, F.Y.; Wang, L.T.; Lin, Y.S.; Li, B.H. Activation of ERK and JNK signaling pathways by mycotoxin citrinin in human cells. Toxicol. Appl. Pharmacol. 2009, 237, 281–287. [Google Scholar] [CrossRef]
- Chan, W.H. Citrinin induces apoptosis via a mitochondria-dependent pathway and inhibition of survival signals in embryonic stem cells, and causes developmental injury in blastocysts. Biochem. J. 2007, 404, 317–326. [Google Scholar] [CrossRef]
- Anbuselvam, C.; Vijayavel, K.; Balasubramanian, M.P. Protective effect of Operculina turpethum against 7,12-dimethyl benz(a)anthracene induced oxidative stress with reference to breast cancer in experimental rats. Chem. Biol. Interact. 2007, 168, 229–236. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Kamendulis, L.M.; Hocevar, B.A. Oxidative stress and oxidative damage in carcinogenesis. Toxicol. Pathol. 2010, 38, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Akyıl, D.; Konuk, M. Detection of genotoxicity and mutagenicity of chlorthiophos using micronucleus, chromosome aberration, sister chromatid exchange, and Ames tests. Environ. Toxicol. 2015, 30, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Koul, M.; Meena, S.; Kumar, A.; Sharma, P.R.; Singamaneni, V.; Riyaz-Ul-Hassan, S.; Hamid, A.; Chaubey, A.; Prabhakar, A.; Gupta, P.; et al. Secondary Metabolites from Endophytic Fungus Penicillium pinophilum Induce ROS-Mediated Apoptosis through Mitochondrial Pathway in Pancreatic Cancer Cells. Planta Med. 2016, 82, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Li, S.J.; Zhang, X.; Wang, X.H.; Zhao, C.Q. Novel natural compounds from endophytic fungi with anticancer activity. Eur. J. Med. Chem. 2018, 156, 316–343. [Google Scholar] [CrossRef] [PubMed]
- Jollow, D.J.; Mitchell, J.R.; Zampaglione, N.; Gillette, J.R. Bromobenzene-induced liver necrosis. Protective role of glutathione and evidence for 3,4-bromobenzene oxide as the hepatotoxic metabolite. Pharmacology 1974, 11, 151–169. [Google Scholar] [CrossRef]
- de Oliveira Filho, J.W.; Dos Santos, T.D.; Tí, R.M.; Dos Reis, A.C.; Hameed, A.; de Oliveira Santos, J.V.; Afzal, M.I.; De Menezes, A.A.; De Alencar, M.V.; Silva, D.H.; et al. Antitumor effects of citrinin in an animal model of Sarcoma 180 via cytogenetic mechanisms. Cell. Mol. Biol. 2020, 66, 120–126. [Google Scholar] [CrossRef]
- Rašić, D.; Micek, V.; Klarić, M.S.; Peraica, M. Oxidative stress as a mechanism of combined OTA and CTN toxicity in rat plasma, liver and kidney. Hum. Exp. Toxicol. 2019, 38, 434–445. [Google Scholar] [CrossRef]
- Richetti, A.; Cavallaro, A.; Ainis, T.; Fimiani, V. Effect of mycotoxins on some activities of isolated human neutrophils. Immunopharmacol. Immunotoxicol. 2005, 27, 433–446. [Google Scholar] [CrossRef]
- Singh, N.D.; Sharma, A.K.; Dwivedi, P.; Patil, R.D.; Kumar, M. Citrinin and endosulfan induced teratogenic effects in Wistar rats. J. Appl. Toxicol. 2007, 27, 143–151. [Google Scholar] [CrossRef]
- Ferguson, L.R.; Chen, H.; Collins, A.R.; Connell, M.; Damia, G.; Dasgupta, S.; Malhotra, M.; Meeker, A.K.; Amedei, A.; Amin, A.; et al. Genomic instability in human cancer: Molecular insights and opportunities for therapeutic attack and prevention through diet and nutrition. Semin. Cancer Biol. 2015, 35, S5–S24. [Google Scholar] [CrossRef]
- Sharma, M.; Sharma, R.; Jain, D.K. Nanotechnology Based Approaches for Enhancing Oral Bioavailability of Poorly Water Soluble Antihypertensive Drugs. Scientifica 2016, 2016, 8525679. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.P.; Pereira, F.C.; Almeida, M.A.P.; Mello, F.M.S.; Pires, W.C.; Pinto, T.M. Cytoxicity and apoptotic mechanism of ruthenium (II) amino acid complexes in sarcoma-180 tumor cells. PLoS ONE 2014, 9, e105865. [Google Scholar] [CrossRef] [PubMed]
- Vanacloig-Pedros, E.; Proft, M.; Pascual-Ahuir, A. Different Toxicity Mechanisms for Citrinin and Ochratoxin A Revealed by Transcriptomic Analysis in Yeast. Toxins 2016, 8, 273. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Filho, J.W.; Andrade, T.D.; de Lima, R.M.; Silva, D.H.; Dos Reis, A.C.; Santos, J.V.; de Meneses, A.A.; de Carvalho, R.M.; da Mata, A.M.; de Alencar, M.V.; et al. Cytogenotoxic evaluation of the acetonitrile extract, citrinin and dicitrinin-A from Penicillium citrinum. Drug Chem. Toxicol. 2022, 45, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Chan, W.H. Protective Effects of Liquiritigenin against Citrinin-Triggered, Oxidative-Stress-Mediated Apoptosis and Disruption of Embryonic Development in Mouse Blastocysts. Int. J. Mol. Sci. 2017, 18, 2538. [Google Scholar] [CrossRef] [PubMed]
- Yard, B.D.; Adams, D.J.; Chie, E.K.; Tamayo, P.; Battaglia, J.S.; Gopal, P.; Rogacki, K.; Pearson, B.E.; Phillips, J.; Raymond, D.P.; et al. A genetic basis for the variation in the vulnerability of cancer to DNA damage. Nat. Commun. 2016, 7, 11428. [Google Scholar] [CrossRef] [PubMed]
- Salah, A.; Bouaziz, C.; Prola, A.; Pires Da Silva, J.; Bacha, H.; Abid-Essefi, S.; Lemaire, C. Citrinin induces apoptosis in human HCT116 colon cancer cells through endoplasmic reticulum stress. J. Toxicol. Environ. Health A 2017, 80, 1230–1241. [Google Scholar] [CrossRef]
- Minamisakamoto, T.; Nishiguchi, S.; Hashimoto, K.; Ogawara, K.I.; Maruyama, M.; Higaki, K. Sequential administration of PEG-Span 80 niosome enhances anti-tumor effect of doxorubicin-containing PEG liposome. Eur. J. Pharm. Biopharm. 2021, 169, 20–28. [Google Scholar] [CrossRef]
- Nandi, U.; Onyesom, I.; Douroumis, D. Anti-cancer activity of sirolimus loaded liposomes in prostate cancer cell lines. J. Drug Deliv. Sci. Technol. 2021, 61, 102200. [Google Scholar] [CrossRef]
- Shi, Z.; Li, Q.; Mei, L. pH-Sensitive nanoscale materials as robust drug delivery systems for cancer therapy. Chin. Chem. Lett. 2020, 31, 1345–1356. [Google Scholar] [CrossRef]
- Zhou, X.; Hao, Y.; Yuan, L.; Pradhan, S.; Shrestha, K.; Pradhan, O.; Li, W. Nano-formulations for transdermal drug delivery: A review. Chin. Chem. Lett. 2018, 29, 1713–1724. [Google Scholar] [CrossRef]
- Powell, D.; Chandra, S.; Dodson, K.; Shaheen, F.; Wiltz, K.; Ireland, S.; Syed, M.; Dash, S.; Wiese, T.; Mandal, T.; et al. Aptamer-functionalized hybrid nanoparticle for the treatment of breast cancer. Eur. J. Pharm. Biopharm. 2017, 114, 108–118. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Groups and Weeks | Behavioral Parameters and Locomotor Activity | |||||
|---|---|---|---|---|---|---|
| Open Field | Rota Rod | |||||
| Crossings | Self-Cleaning | Elevation | TP (s) | NQ | ||
| Olive oil | S 01 | 83.20 ± 20.60 | 5.00 ± 2.14 | 12.00 ± 6.16 | 180.00 ± 0.01 | 0.40 ± 0.02 |
| S 01 | 83.20 ± 20.60 | 5.00 ± 2.14 | 12.00 ± 6.16 | 180.00 ± 0.01 | 0.40 ± 0.02 | |
| S 02 | 81.00 ± 16.80 | 14.40 ± 3.05 | 24.00 ± 7.48 | 174.40 ± 7.43 | 1.40 ± 0.89 | |
| S 03 | 83.60 ± 33.40 | 11.40 ± 5.12 | 24.00 ± 19.5 | 180.00 ± 0.01 | 0.00 ± 0.00 | |
| S 04 | 57.20 ± 9.44 | 14.00 ± 2.91 | 16.80 ± 2.16 | 176.80 ± 7.15 | 0.00 ± 0.00 | |
| S 05 | 55.00 ± 14.80 | 12.60 ± 6.10 | 12.80 ± 6.26 | 180.00 ± 0.10 | 0.00 ± 0.00 | |
| S 06 | 52.20 ± 21.40 | 9.80 ± 4.08 | 13.60± 8.05 | 180.00 ± 0.05 | 0.00 ± 0.00 | |
| S 07 | 37.00 ± 26.40 | 8.40 ± 11.3 | 14.60 ± 4.55 | 180.00 ± 0.20 | 0.00 ± 0.00 | |
| S 08 | 73.10 ± 15.00 | 7.70 ± 3.45 | 23.90 ± 5.46 | 180.00 ± 0.01 | 0.00 ± 0.00 | |
| S 09 | 80.00 ±16.80 | 12.35 ± 5.23 | 27.74 ± 7.5 | 180.00 ± 0.03 | 0.00 ± 0.00 | |
| S 10 | 82.30 ± 31.40 | 13.00 ± 4.42 | 22.41 ± 5.54 | 180.00 ± 0.02 | 0.00 ± 0.00 | |
| S 11 | 56.50 ± 12.00 | 8.99 ± 2.34 | 23.60 ± 7.55 | 180.00 ± 0.03 | 0.00 ± 0.00 | |
| S 12 | 55.00 ± 14.00 | 9.13 ± 3.51 | 21.80 ± 4.78 | 180.00 ± 0.10 | 0.00 ± 0.00 | |
| S 13 | 55.20 ± 20.10 | 13.24 ± 5.1 | 18.59 ± 5.55 | 180.00 ± 0.10 | 0.00 ± 0.00 | |
| S 14 | 39.00 ± 27.00 | 11.25 ± 5.12 | 14.00 ± 4.55 | 180.00 ± 0.02 | 0.00 ± 0.00 | |
| DMBA 6 mg/kg | S 01 | 68.20 ± 23.0 | 9.60 ± 4.82 | 20.60 ± 5.94 | 169.00 ± 11.44 a | 1.40 ± 0.67 |
| S 02 | 82.00 ± 30.0 | 15.80 ± 6.14 | 27.20 ± 4.60 | 176.00 ± 6.92 | 0.40 ± 0.28 | |
| S 03 | 65.00 ± 14.6 | 11.20 ± 1.30 | 19.60 ± 3.43 | 180.00 ± 0.00 | 0.00 ± 0.00 | |
| S 04 | 76.80 ± 11.3 a | 15.80 ± 7.59 | 19.40 ± 10.6 | 176.60 ± 7.60 | 0.60 ± 0.12 | |
| S 05 | 76.40 ± 18.9 | 15.40 ± 13.9 | 20.40 ± 4.03 | 169.40 ± 7.87 a | 0.80 ± 0.32 | |
| S 06 | 62.20 ± 24.3 | 11.40 ± 4.72 | 26.60 ± 12.1 | 168.87 ± 7.15 a | 0.00 ± 0.00 | |
| S 07 | 60.80 ± 7.19 | 14.70 ± 10.3 | 19.80 ± 7.08 | 168.57 ± 7.14 a | 0.00 ± 0.00 | |
| S 08 | 63.00 ± 5.80 | 10.30 ± 4.50 | 23.70 ± 7.30 | 180.00 ± 0.10 | 0.00 ± 0.00 | |
| S 09 | 70.50 ± 6.20 | 12.34 ± 7.00 | 18.50 ± 5.20 | 180.00 ± 0.20 | 0.00 ± 0.00 | |
| S 10 | 69.40 ± 3.90 | 13.43 ± 3.70 | 19.35 ± 4.30 | 180.00 ± 0.10 | 0.00 ± 0.00 | |
| S 11 | 80.20 ± 14.00 | 15.37 ± 5.35 | 20.30 ± 5.34 | 165.00 ± 8.03 a | 1.85 ± 1.51 a | |
| CPA 25 mg/kg | S 12 | 54.67 ± 18.23 | 18.26 ± 6.24 | 22.6 ± 5.55 | 165.0 ± 15.00 | 4.30 ± 1.12 |
| S 13 | 78.00 ± 22.39 | 17.78 ± 4.78 | 21.8 ± 4.13 | 164.9 ± 13.50 | 1.80 ± 0.21 | |
| S 14 | 68.28 ± 18.34 | 20.34 ± 9.73 | 32.00 ± 6.32 a | 177.73 ± 12.24 | 1.80 ± 0.19 | |
| CIT 2 μg/kg | S 12 | 50.28 ± 15.51 | 14.0 ± 4.96 | 24.28 ± 6.77 | 175.57 ± 9.14 | 0.71 ± 1.46 |
| S 13 | 51.14 ± 13.65 | 15.14 ± 4.87 | 23.71 ± 6.52 | 179.14 ± 2.23 | 0.42 ± 1.11 | |
| S 14 | 47.14 ± 31.60 | 13.42 ± 8.59 | 17.42 ± 10.81 | 178.14 ± 3.07 | 0.28 ± 0.47 | |
| LP-CIT 2 μg/kg | S 12 | 82.30 ± 21.70 | 13.00 ± 4.42 | 22.41 ± 5.54 | 180.00 ± 0.02 | 0.00 ± 0.00 |
| S 13 | 73.10 ± 11.00 | 7.70 ± 3.45 | 24.90 ± 5.46 | 180.00 ± 0.20 | 0.00 ± 0.00 | |
| S 14 | 80.00 ±13.80 | 12.35 ± 5.23 | 27.74 ± 7.5 | 180.00 ± 0.10 | 0.00 ± 0.00 | |
| LP-CIT 6 μg/kg | S 12 | 78.50 ± 7.40 | 12.54 ± 7.00 | 18.30 ± 5.30 | 180.00 ± 0.10 | 0.00 ± 0.00 |
| S 13 | 79.40 ± 2.90 | 14.43 ± 5.70 | 20.35 ± 4.30 | 180.00 ± 0.03 | 0.00 ± 0.00 | |
| S 14 | 79.20 ± 4.00 | 16.34 ± 3.37 | 22.50 ± 6.34 | 180.00 ± 0.20 | 0.00 ± 0.00 | |
| Groups | ||||||
|---|---|---|---|---|---|---|
| Hematological Profile | Vehicle 10 mg/kg | DMBA 6 mg/kg | CPA 25 mg/kg | CIT 2 mg/kg | LP-CIT 2 mg/kg | LP-CIT 6 mg/kg |
| Red series | ||||||
| HEM (106/uL) | 7.5 ± 0.63 | 9.65 ± 0.32 | 9.97 ± 0.54 | 9.0 ± 0.70 | 12.21 ± 0.91 | 8.75 ± 2.17 |
| HGB (g/dL) | 14.12 ± 0.67 | 14.8 ± 0.42 | 15.50 ± 1.16 | 15.30 ± 1.40 | 15.52 ± 0.88 | 14.1 ± 0.48 |
| HCT (%) | 37.7 ± 2.95 | 54.1 ± 0.03 | 56.50 ± 2.86 | 51.56 ± 3.10 | 52.87 ± 1.54 | 45.0 ± 2.03 |
| White series | ||||||
| Leukocytes (103/uL) | 5.000 ± 1894.72 | 2.600 ± 424.26 | 3398.0 ± 840.70 | 2834 ± 427.20 | 2.908 ± 378.18 | 5.137 ± 1876.51 |
| Neutrophils (%) | 9.0 ± 1.87 | 4 ± 4.25 | 8.76 ± 2.51 | 11.80 ± 0.70 | 3.5 ± 2.64 | 17.25 ± 5.73 |
| Lymphocytes (%) | 83.0 ± 2.23 | 87.5 ± 1.92 | 77.8 ± 4.62 | 86.00 ± 5.90 | 85.75 ± 5.11 | 79.0 ± 7.87 |
| Monocytes (%) | 8.0 ± 1.87 | 4.5 ± 6.36 | 12.8 ± 4.21 | 1.20 ± 2.20 * | 4.75 ± 3.59 | 2.25 ± 2.5 |
| Eosinophils (%) | 0 ± 0 | 0 ± 0 | 0.14 ± 0.05 | 0.10 ± 0.04 | 0 ± 0 | 0.5 ± 0.57 |
| Others | ||||||
| Platelets (109/uL) | 6672.00 ± 693.88 | 7850.00 ± 108.07 | 1034.00 ± 100.7 * | 8531.0 ± 109.1 | 6390.0 ± 378.15 | 7985.0 ± 937.00 |
| Biochemical Profile | Vehicle 10 mg/kg | DMBA 6 mg/kg | CPA 25 mg/kg | CIT 2 mg/kg | LP-CIT 2 mg/kg | LP-CIT 6 mg/kg |
|---|---|---|---|---|---|---|
| Urea (mg/dL) | 28.2 ± 4.44 | 40 ± 1.41 | 36.0 ± 8.71 | 38.7 ± 9.40 | 28.25 ± 6.99 | 24.25 ± 1.25 |
| Creatinine (mg/dL) | 0.42 ± 0.13 | 0.25 ± 0.07 | 0.46 ± 0.05 | 0.50 ± 0.05 | 0.5 ± 0.18 | 0.475 ± 0.24 |
| TGO (IU/L) | 102.2 ± 13.63 | 142 ± 0.03 | 101.6 ± 13.6 | 231.5 ± 22.00 * | 138 ± 18.97 | 105.75 ± 27.84 |
| TGP (IU/L) | 85.8 ± 9.52 | 101 ± 0.04 | 156.6 ± 6.65 * | 186.5 ± 24.60 * | 125.75 ± 9.53 | 120 ± 20.54 |
| Treatment | Dose | Damage Index | Frequency of Damage | ||||
|---|---|---|---|---|---|---|---|
| Breast | Liver | Bone Marrow | Breast | Liver | Bone Marrow | ||
| Vehicle | 10 mL/kg | 24.60 ± 3.1 | 25.1 ± 10.9 | 34 ± 13.4 | 27.7 ± 3.9 | 32.2 ± 9.1 | 38.1 ± 10.2 |
| DMBA | 6 mg/kg | 131.2 ± 9.2 a | 191.1 ± 18.7 a | 268.6 ± 18.7 a | 89.1 ± 2.2 a | 86.7 ± 3.5 a | 94.7± 2.4 a |
| CPA | 25 mg/kg | 187.6 ± 6.8 ab | 166.4 ± 18.9 a | 155.9 ± 17.6 ab | 98.8 ± 2.9 ab | 96.3 ± 3.6 a | 96.4 ± 3.2 a |
| CIT | 2.0 µg/kg | 135.4 ± 10.1 ac | 130.3 ± 22.7 ab | 93.8 ± 25.6 abc | 85.5 ± 4.2 ac | 81.3 ± 7.1 a | 82.1 ± 5.8 abc |
| LP-CIT | 2.0 µg/kg | 150.1 ± 14.1 ac | 64.2 ± 10.2 abcd | 42.9 ± 8.9 bcd | 95.4 ± 4.2 a | 69.1 ± 4.9 abc | 46.8 ± 5.4 bcd |
| 6.0 µg/kg | 183.8 ± 10.2 abd | 82.9 ± 9.9 abcd | 55.9 ± 10.2 bc | 96.3 ± 3.6 ad | 76.9 ± 5.8 ac | 69.8 ± 5.9 abc | |
| Group | Dose | Breast | Bone Marrow | Liver | PCE/EPC + NCE |
|---|---|---|---|---|---|
| Vehicle | 10 mL/kg | 6.1 ± 2.6 | 6.0 ± 1.3 | 5.5 ± 1.5 | 0.78 ± 0.05 |
| CPA | 25 mg/kg | 39.4 ± 5.4 a | 28.4 ± 3.9 a | 34.6 ± 4.6 a | 0.46 ± 0.02 a |
| CIT | 2.0 µg/kg | 12.0 ± 4.5 b | 10.0 ± 1.5 b | 9.2 ± 3.1 b | 0.73 ± 0.04 b |
| LP-CIT | 2.0 µg/kg | 13.1 ± 4.8 b | 8.4 ± 3.5 b | 7.9 ± 3.7 b | 0.72 ± 0.03 b |
| 6.0 µg/kg | 24.4 ± 3.5 abc | 10.2 ± 4.7 b | 9.2 ± 4.0 b | 0.71 ± 0.06 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moura, M.L.V.; de Menezes, A.-A.P.M.; de Oliveira Filho, J.W.G.; do Nascimento, M.L.L.B.; dos Reis, A.C.; Ribeiro, A.B.; da Silva, F.C.C.; Nunes, A.M.V.; Rolim, H.M.L.; de Carvalho Melo Cavalcante, A.A.; et al. Advances in Antitumor Effects Using Liposomal Citrinin in Induced Breast Cancer Model. Pharmaceutics 2024, 16, 174. https://doi.org/10.3390/pharmaceutics16020174
Moura MLV, de Menezes A-APM, de Oliveira Filho JWG, do Nascimento MLLB, dos Reis AC, Ribeiro AB, da Silva FCC, Nunes AMV, Rolim HML, de Carvalho Melo Cavalcante AA, et al. Advances in Antitumor Effects Using Liposomal Citrinin in Induced Breast Cancer Model. Pharmaceutics. 2024; 16(2):174. https://doi.org/10.3390/pharmaceutics16020174
Chicago/Turabian StyleMoura, Michely Laiany Vieira, Ag-Anne Pereira Melo de Menezes, José Williams Gomes de Oliveira Filho, Maria Luiza Lima Barreto do Nascimento, Antonielly Campinho dos Reis, Alessandra Braga Ribeiro, Felipe Cavalcanti Carneiro da Silva, Adriana Maria Viana Nunes, Hercília Maria Lins Rolim, Ana Amélia de Carvalho Melo Cavalcante, and et al. 2024. "Advances in Antitumor Effects Using Liposomal Citrinin in Induced Breast Cancer Model" Pharmaceutics 16, no. 2: 174. https://doi.org/10.3390/pharmaceutics16020174