

Anti-Inflammatory Effects of the LK5 Herbal Complex on LPS- and IL-4/IL-13-Stimulated HaCaT Cells and a DNCB-Induced Animal Model of Atopic Dermatitis in BALB/c Mice
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Materials
2.3. High-Performance Liquid Chromatography (HPLC) Analysis
2.4. Cell Culture
2.5. Cell Viability and Nitrite Determination
2.6. Reverse Transcription Quantitative Polymerase Chain Reaction (RT-qPCR)
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Western Blot Analysis
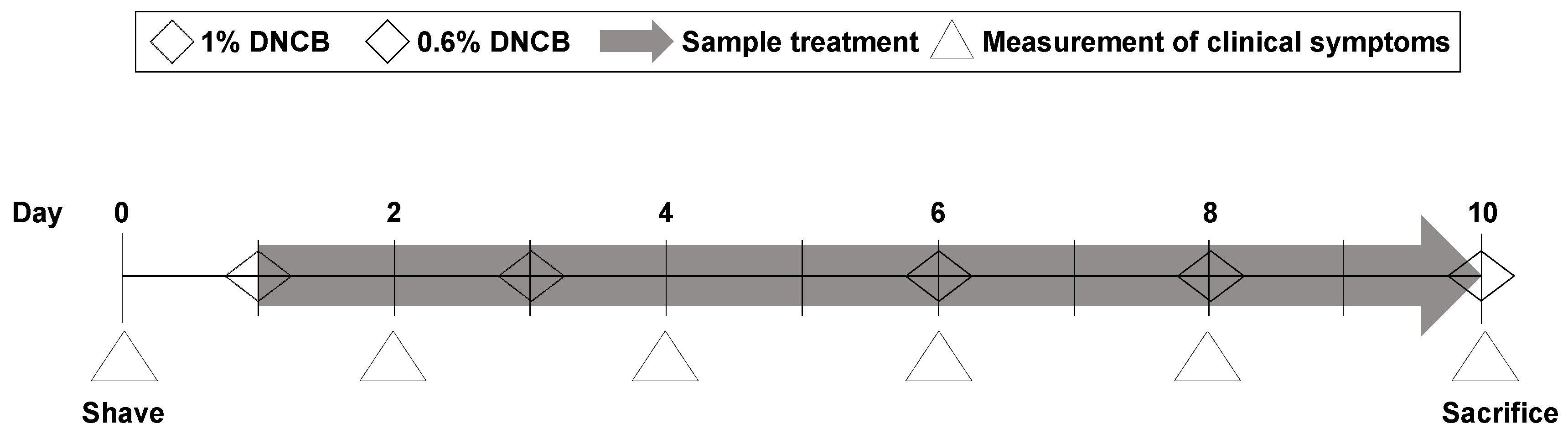
2.9. DNCB-Induced Animal Model of Atopic Dermatitis
2.10. Measurement of Clinical Symptoms Including Dermatitis Score and Ear Thickness
2.11. Transepidermal Water Loss
2.12. Histological Analysis
2.13. Pruritus
2.14. Statistic
3. Results
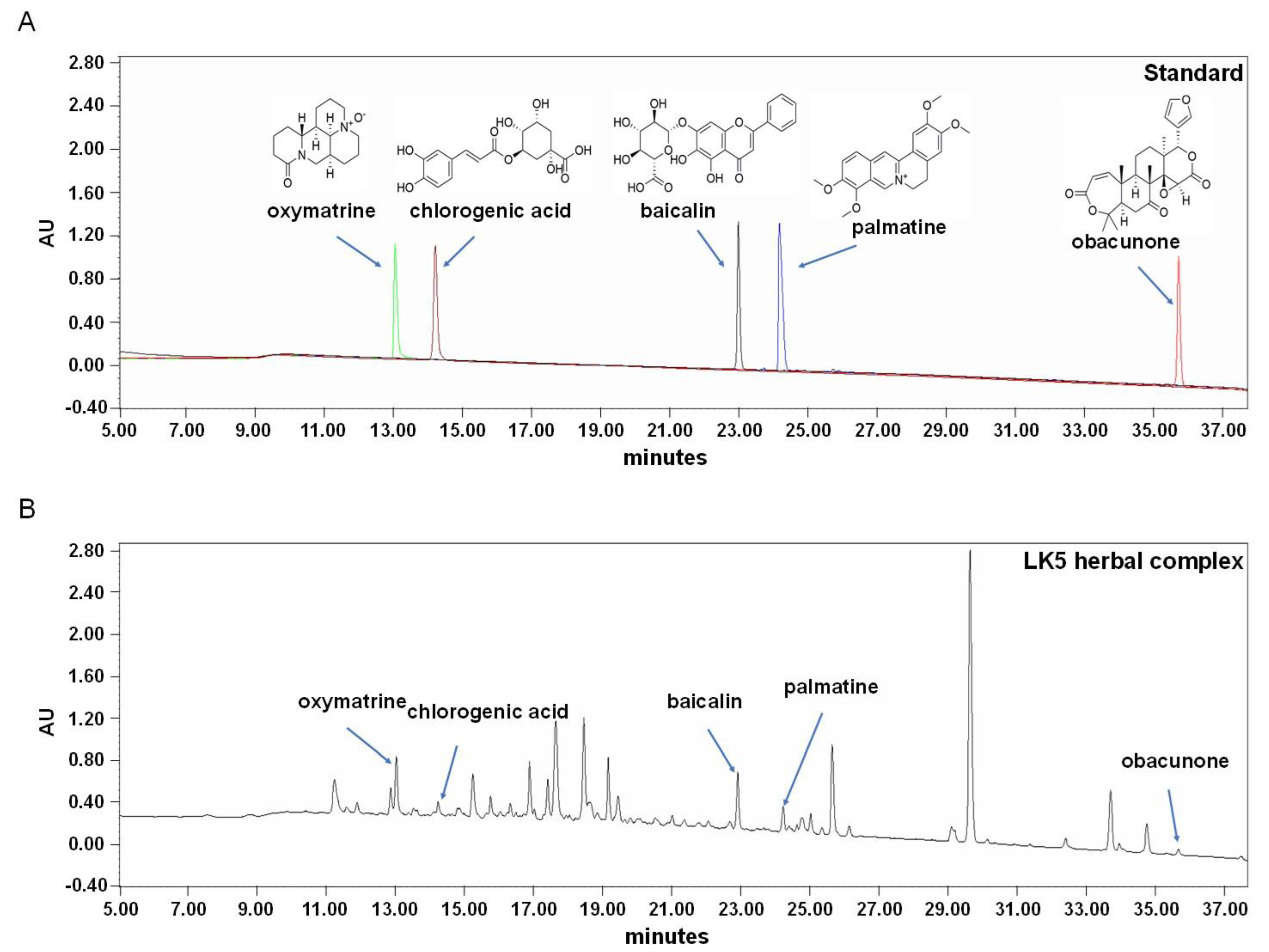
3.1. Quantification of the LK5 Herbal Complex Using HPLC
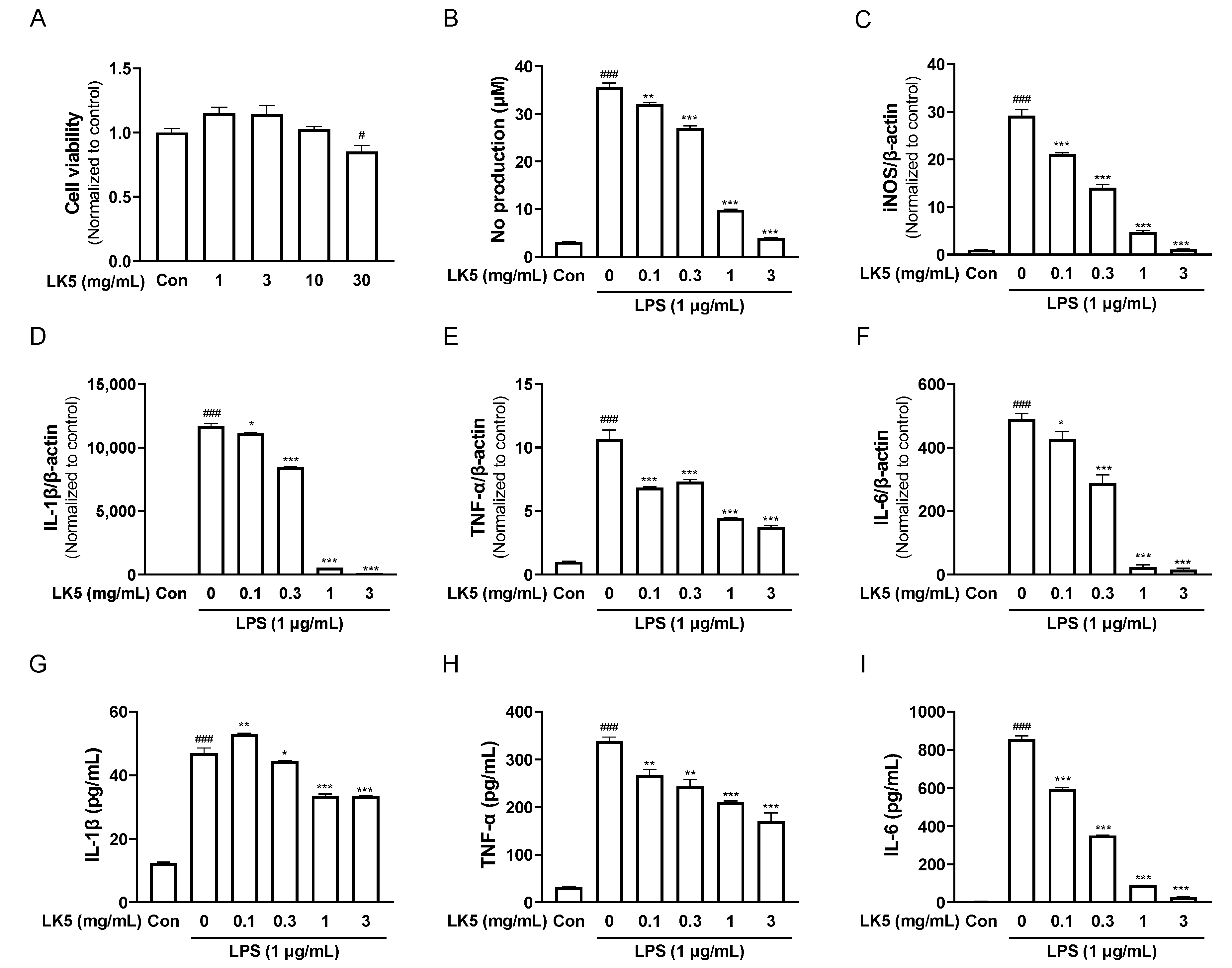
3.2. The LK5 Herbal Complex Inhibited LPS-Stimulated Inflammatory Mediators and Inflammatory Cytokines in HaCaT Cells
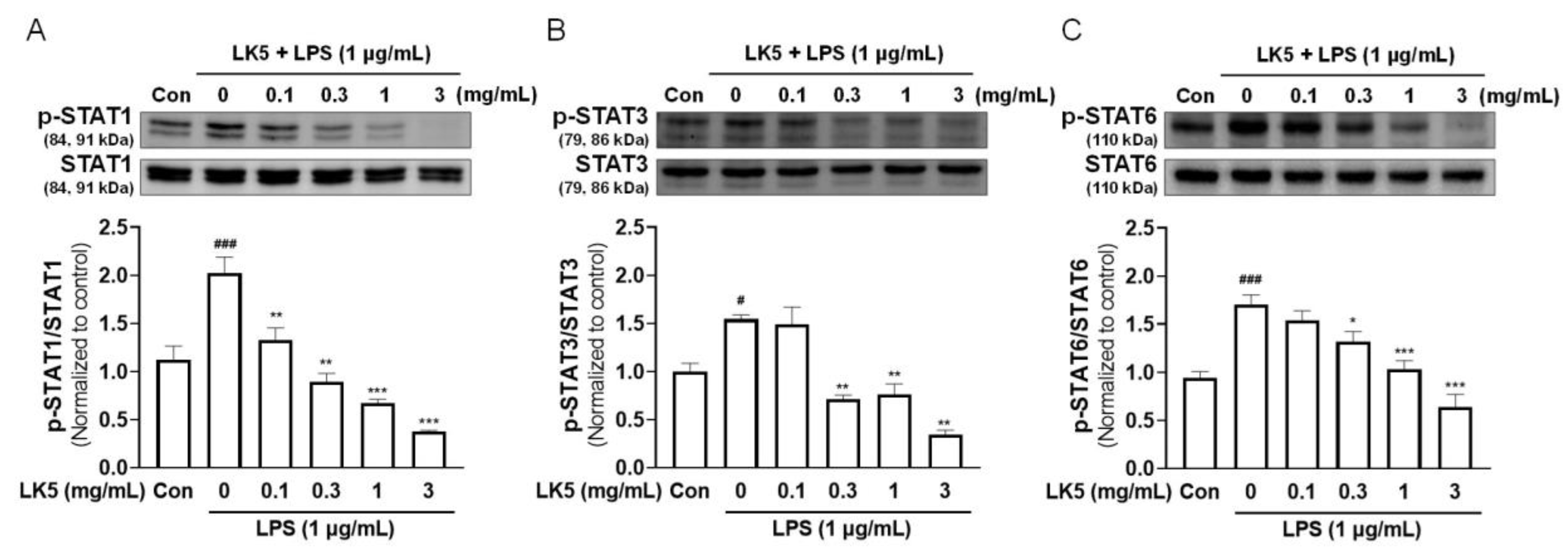
3.3. The LK5 Herbal Complex Downregulated the Phosphorylation of STAT in LPS-Stimulated HaCaT Cells
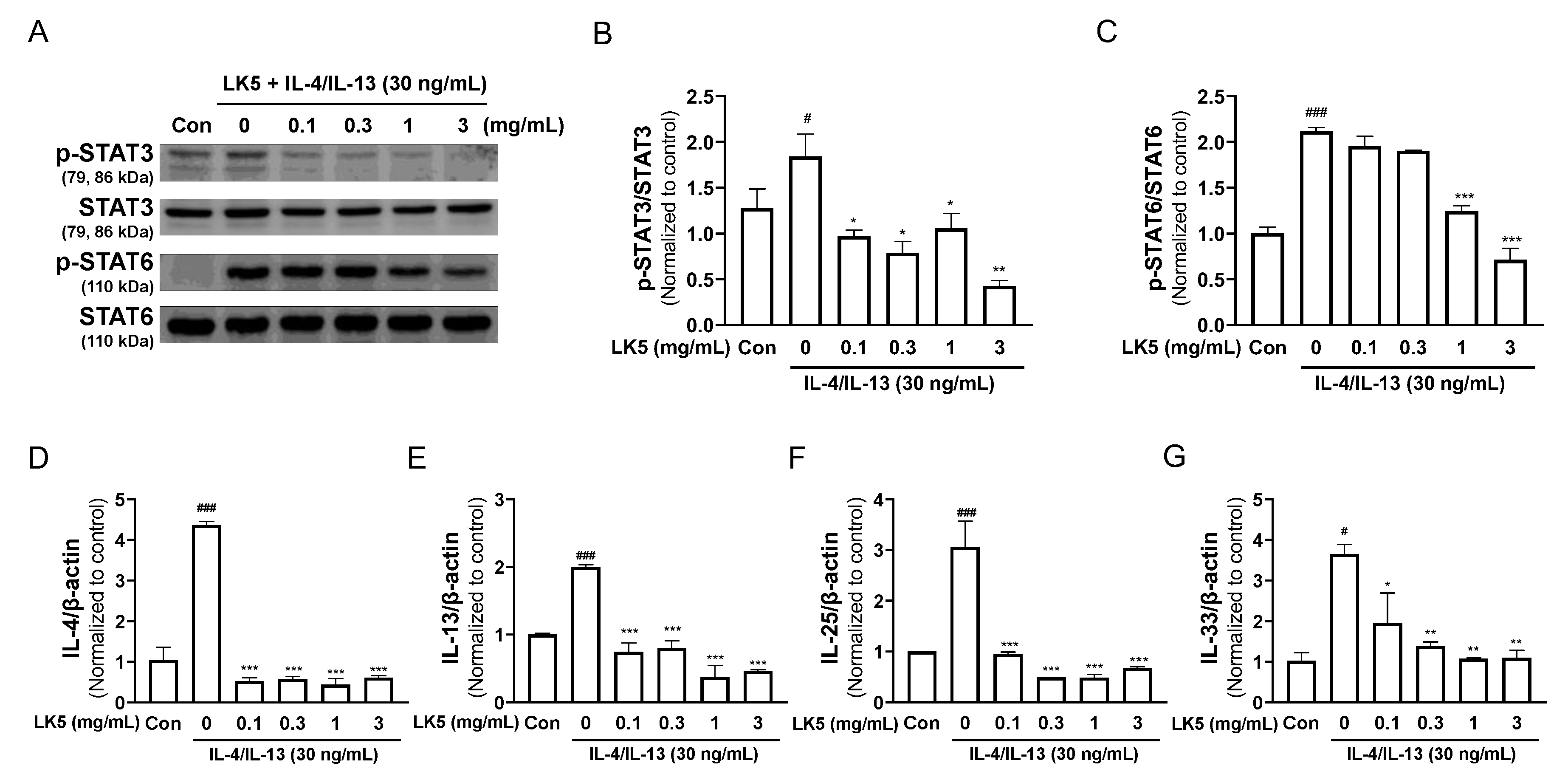
3.4. The LK5 Herbal Complex Regulated Inflammatory Cytokines by Inhibiting STAT Phosphorylation in IL-4/IL-13-Stimulated HaCaT Cells
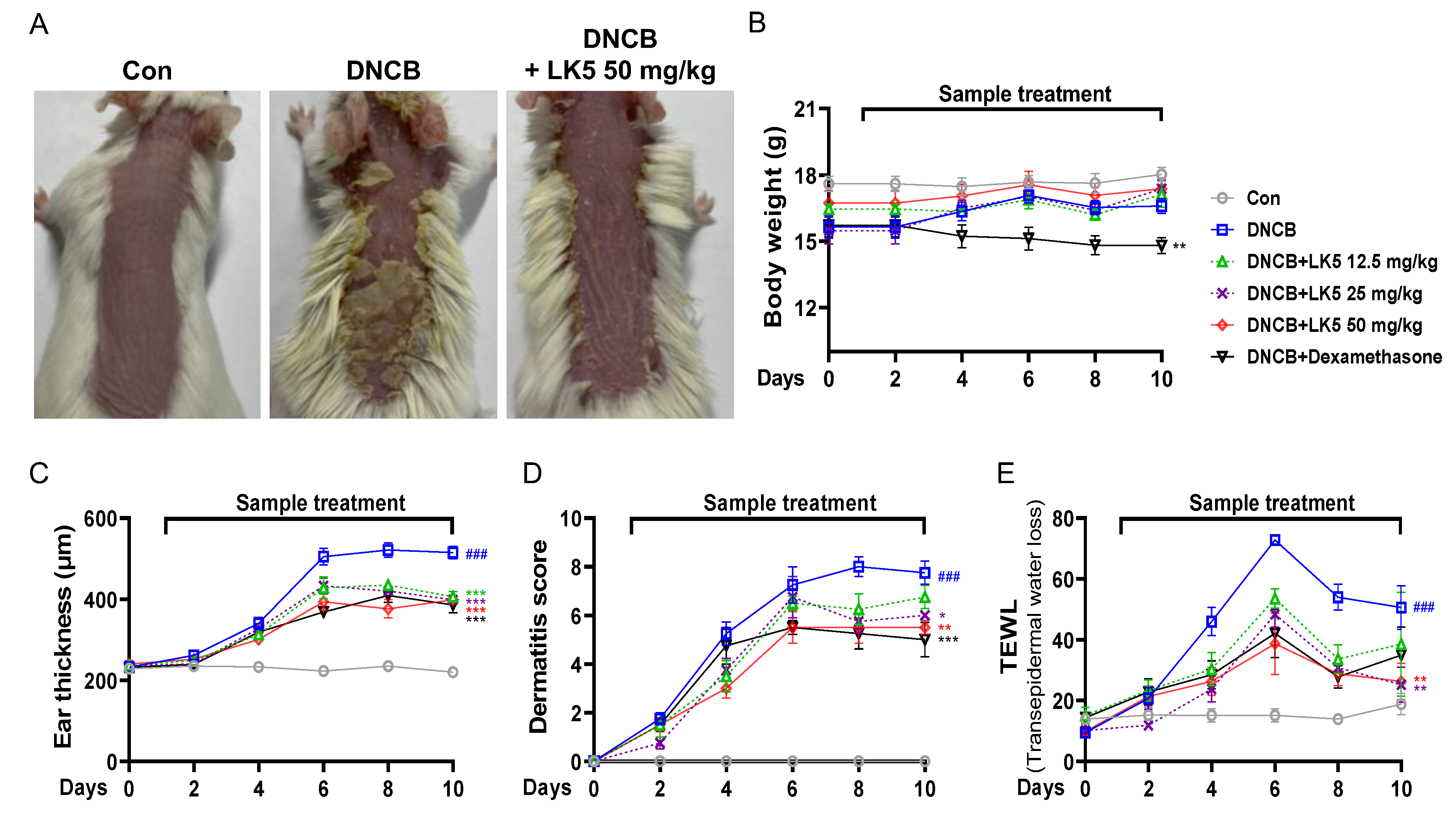
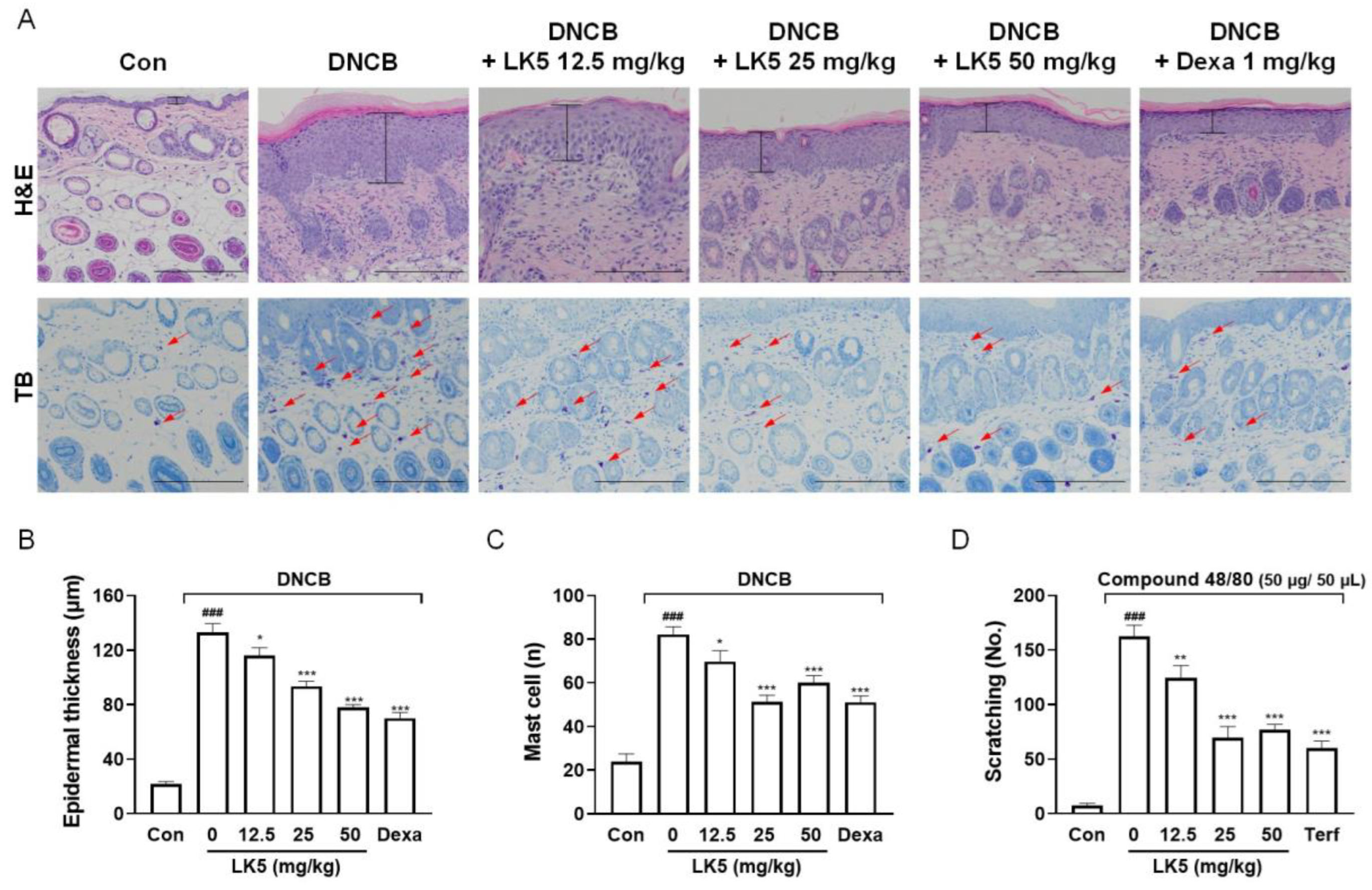
3.5. The LK5 Herbal Complex Ameliorated Clinical Symptoms and Histological Analysis in DNCB-Induced AD and Compound 48/80-Induced Pruritis Animal Models
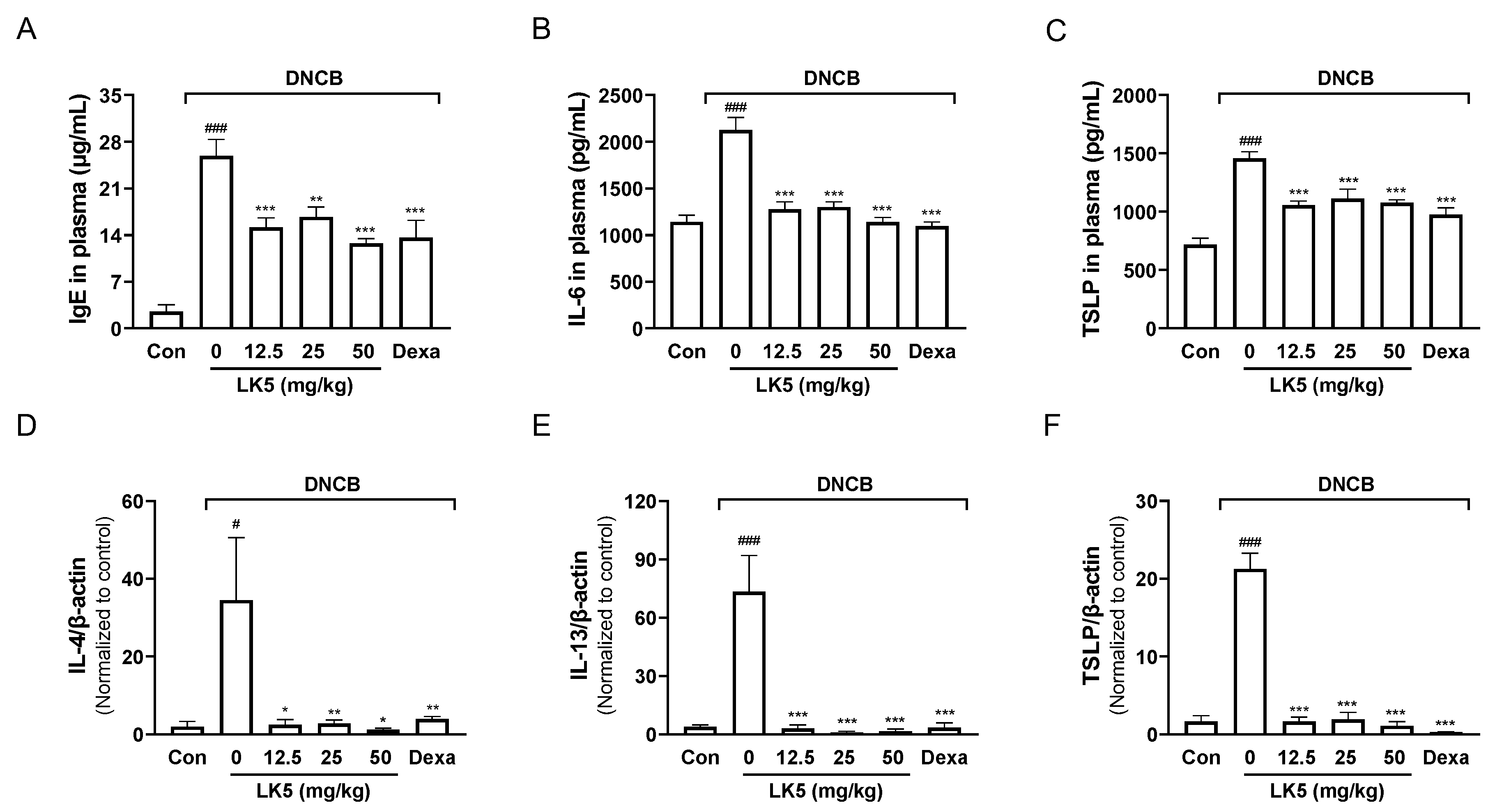
3.6. The LK5 Herbal Complex Inhibited IgE and Inflammatory Cytokines in the DNCB-Induced AD Animal Model
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | Atopic dermatitis |
| DNCB | 2,4-Dinitrochlorobenzene |
| ELISA | Enzyme-linked immunosorbent assay |
| H&E | Hematoxylin and eosin |
| IL | Interleukin |
| IgE iNOS | Immunoglobulin E Inducible nitric oxide synthase |
| LPS MAPK NO | Lipopolysaccharide Mitogen-Activated Protein Kinase Nitric oxide |
| RT-qPCR | Reverse transcription-quantitative polymerase chain reaction |
| STAT | Signal transducers and activators of transcription |
| TB TEWL TNF-α TSLP | Toluidine blue Transepidermal water loss Tumor necrosis factor-α Thymic stromal lymphopoietin |
References
- Boguniewicz, M.; Leung, D.Y. Atopic dermatitis: A disease of altered skin barrier and immune dysregulation. Immunol. Rev. 2011, 242, 233–246. [Google Scholar] [CrossRef]
- Gur Cetinkaya, P.; Sahiner, U.M. Childhood atopic dermatitis: Current developments, treatment approaches, and future expectations. Turk. J. Med. Sci. 2019, 49, 963–984. [Google Scholar] [CrossRef] [PubMed]
- Silvestre Salvador, J.F.; Romero-Perez, D.; Encabo-Duran, B. Atopic Dermatitis in Adults: A Diagnostic Challenge. J. Investig. Allergol. Clin. Immunol. 2017, 27, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, B.E.; Leung, D.Y.M. Pathophysiology of atopic dermatitis: Clinical implications. Allergy Asthma Proc. 2019, 40, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Vakharia, P.P.; Silverberg, J.I. Adult-Onset Atopic Dermatitis: Characteristics and Management. Am. J. Clin. Dermatol. 2019, 20, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Gittler, J.K.; Krueger, J.G.; Guttman-Yassky, E. Atopic dermatitis results in intrinsic barrier and immune abnormalities: Implications for contact dermatitis. J. Allergy Clin. Immunol. 2013, 131, 300–313. [Google Scholar] [CrossRef]
- Wallmeyer, L.; Dietert, K.; Sochorova, M.; Gruber, A.D.; Kleuser, B.; Vavrova, K.; Hedtrich, S. TSLP is a direct trigger for T cell migration in filaggrin-deficient skin equivalents. Sci. Rep. 2017, 7, 774. [Google Scholar] [CrossRef]
- Dong, C.; Flavell, R.A. Cell fate decision: T-helper 1 and 2 subsets in immune responses. Arthritis Res. 2000, 2, 179–188. [Google Scholar] [CrossRef]
- Iwasaki, N.; Terawaki, S.; Shimizu, K.; Oikawa, D.; Sakamoto, H.; Sunami, K.; Tokunaga, F. Th2 cells and macrophages cooperatively induce allergic inflammation through histamine signaling. PLoS ONE 2021, 16, e0248158. [Google Scholar] [CrossRef]
- Jiang, H.; Harris, M.B.; Rothman, P. IL-4/IL-13 signaling beyond JAK/STAT. J. Allergy Clin. Immunol. 2000, 105, 1063–1070. [Google Scholar] [CrossRef]
- Shao, F.; Pang, X.; Baeg, G.H. Targeting the JAK/STAT Signaling Pathway for Breast Cancer. Curr. Med. Chem. 2021, 28, 5137–5151. [Google Scholar] [CrossRef] [PubMed]
- Szalus, K.; Trzeciak, M.; Nowicki, R.J. JAK-STAT Inhibitors in Atopic Dermatitis from Pathogenesis to Clinical Trials Results. Microorganisms 2020, 8, 1743. [Google Scholar] [CrossRef]
- Di Benedetto, P.; Ruscitti, P.; Berardicurti, O.; Panzera, N.; Grazia, N.; Di Vito Nolfi, M.; Di Francesco, B.; Navarini, L.; Maurizi, A.; Rucci, N.; et al. Blocking Jak/STAT signalling using tofacitinib inhibits angiogenesis in experimental arthritis. Arthritis Res. Ther. 2021, 23, 213. [Google Scholar] [CrossRef] [PubMed]
- Bridgewood, C.; Wittmann, M.; Macleod, T.; Watad, A.; Newton, D.; Bhan, K.; Amital, H.; Damiani, G.; Giryes, S.; Bragazzi, N.L.; et al. T Helper 2 IL-4/IL-13 Dual Blockade with Dupilumab Is Linked to Some Emergent T Helper 17–Type Diseases, Including Seronegative Arthritis and Enthesitis/Enthesopathy, but Not to Humoral Autoimmune Diseases. J. Investig. Dermatol. 2022, 142, 2660–2667. [Google Scholar] [CrossRef] [PubMed]
- Bath-Hextall, F.; Delamere, F.M.; Williams, H.C. Dietary exclusions for established atopic eczema. Cochrane Database Syst. Rev. 2008, 2008. 1, CD005203. [Google Scholar] [CrossRef]
- Abuabara, K.; Margolis, D.J.; Langan, S.M. The Long-Term Course of Atopic Dermatitis. Dermatol. Clin. 2017, 35, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Takamori, K. Peripheral itch sensitization in atopic dermatitis. Allergol. Int. 2022, 71, 265–277. [Google Scholar] [CrossRef]
- Yasir, M.; Goyal, A.; Sonthalia, S. Corticosteroid Adverse Effects; StatPearls: Treasure Island, FL, USA, 2023. [Google Scholar]
- Herrero, M.; Cifuentes, A.; Ibanez, E. Sub- and supercritical fluid extraction of functional ingredients from different natural sources: Plants, food-by-products, algae and microalgae—A review. Food Chem. 2006, 98, 136–148. [Google Scholar] [CrossRef]
- Zhao, Q.; Chen, X.Y.; Martin, C. Scutellaria baicalensis, the golden herb from the garden of Chinese medicinal plants. Sci. Bull. 2016, 61, 1391–1398. [Google Scholar] [CrossRef]
- He, X.; Fang, J.; Huang, L.; Wang, J.; Huang, X. Sophora flavescens Ait.: Traditional usage, phytochemistry and pharmacology of an important traditional Chinese medicine. J. Ethnopharmacol. 2015, 172, 10–29. [Google Scholar] [CrossRef]
- Park, C.H.; Morgan, A.M.A.; Park, B.B.; Lee, S.Y.; Lee, S.; Kim, J.K.; Park, S.U. Metabolic Analysis of Four Cultivars of Liriope platyphylla. Metabolites 2019, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Sun, J.; Xu, J.; Yan, Q.; Gao, E.; Qu, W.; Zhao, Y.; Yu, Z. Pharmacokinetics, tissue distribution and excretion study of dictamnine, a major bioactive component from the root bark of Dictamnus dasycarpus Turcz. (Rutaceae). J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2013, 942–943, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Nakashima, K.; Nishino, K.; Kotani, K.; Tomida, J.; Inoue, M.; Kawamura, Y. Berberine Is a Novel Type Efflux Inhibitor Which Attenuates the MexXY-Mediated Aminoglycoside Resistance in Pseudomonas aeruginosa. Front. Microbiol. 2016, 7, 1223. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Kim, S.Y.; Mony, T.J.; Bae, H.J.; Choi, S.H.; Choi, Y.Y.; An, J.Y.; Kim, H.J.; Cho, Y.E.; Sowndhararajan, K.; et al. Moringa concanensis L. Alleviates DNCB-Induced Atopic Dermatitis-like Symptoms by Inhibiting NLRP3 Inflammasome-Mediated IL-1beta in BALB/c Mice. Pharmaceuticals 2022, 15, 1217. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Han, S.D.; Kim, M.; Mony, T.J.; Lee, E.S.; Kim, K.M.; Choi, S.H.; Hong, S.H.; Choi, J.W.; Park, S.J. Mentha arvensis Essential Oil Exerts Anti-Inflammatory in LPS-Stimulated Inflammatory Responses via Inhibition of ERK/NF-kappaB Signaling Pathway and Anti-Atopic Dermatitis-like Effects in 2,4-Dinitrochlorobezene-Induced BALB/c Mice. Antioxidants 2021, 10, 1941. [Google Scholar] [CrossRef] [PubMed]
- Oranje, A.P.; Glazenburg, E.J.; Wolkerstorfer, A.; de Waard-van der Spek, F.B. Practical issues on interpretation of scoring atopic dermatitis: The SCORAD index, objective SCORAD and the three-item severity score. Br. J. Dermatol. 2007, 157, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, Y.; Umakoshi, K.; Nojiri, N.; Kamei, C. Effects of histamine H1 receptor antagonists on compound 48/80-induced scratching behavior in mice. Eur. J. Pharmacol. 1998, 351, 1–5. [Google Scholar] [CrossRef]
- Liao, H.; Ye, J.; Gao, L.; Liu, Y. The main bioactive compounds of Scutellaria baicalensis Georgi. for alleviation of inflammatory cytokines: A comprehensive review. Biomed. Pharmacother. 2021, 133, 110917. [Google Scholar] [CrossRef]
- Yuan, X.; Sun, Y.; Miao, N.; Sun, S.; Wang, Y.; Hu, Z.; Yuan, J.; Xu, M.; Liu, Z. The synergistic anti-inflammatory effect of the combination of sodium ferulate and oxymatrine and its modulation on inflammation-associated mediators in RAW 264.7 cells. J. Ethnopharmacol. 2011, 137, 1477–1485. [Google Scholar] [CrossRef]
- Luo, X.; Yu, Z.; Yue, B.; Ren, J.; Zhang, J.; Mani, S.; Wang, Z.; Dou, W. Obacunone reduces inflammatory signalling and tumour occurrence in mice with chronic inflammation-induced colorectal cancer. Pharm. Biol. 2020, 58, 886–897. [Google Scholar] [CrossRef]
- Ryuk, J.A.; Zheng, M.S.; Lee, M.Y.; Seo, C.S.; Li, Y.; Lee, S.H.; Moon, D.C.; Lee, H.W.; Lee, J.H.; Park, J.Y.; et al. Discrimination of Phellodendron amurense and P. chinense based on DNA analysis and the simultaneous analysis of alkaloids. Arch. Pharm. Res. 2012, 35, 1045–1054. [Google Scholar] [CrossRef]
- Lan, X.; Zhao, J.; Zhang, Y.; Chen, Y.; Liu, Y.; Xu, F. Oxymatrine exerts organ- and tissue-protective effects by regulating inflammation, oxidative stress, apoptosis, and fibrosis: From bench to bedside. Pharmacol. Res. 2020, 151, 104541. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.J.; Xu, B.; Huang, S.W.; Luo, X.; Deng, X.L.; Luo, S.; Liu, C.; Wang, Q.; Chen, J.Y.; Zhou, L. Baicalin prevents LPS-induced activation of TLR4/NF-kappaB p65 pathway and inflammation in mice via inhibiting the expression of CD14. Acta Pharmacol. Sin. 2021, 42, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Chen, F.; Fang, H.; Mi, J.; Qi, Q.; Yang, M. Daphnetin inhibits proliferation and inflammatory response in human HaCaT keratinocytes and ameliorates imiquimod-induced psoriasis-like skin lesion in mice. Biol. Res. 2020, 53, 48. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Siddiqi, M.H.; Aceituno, V.C.; Simu, S.Y.; Zhang, J.; Jimenez Perez, Z.E.; Kim, Y.J.; Yang, D.C. Ginsenoside Rg5:Rk1 attenuates TNF-alpha/IFN-gamma-induced production of thymus- and activation-regulated chemokine (TARC/CCL17) and LPS-induced NO production via downregulation of NF-kappaB/p38 MAPK/STAT1 signaling in human keratinocytes and macrophages. In Vitro Cell Dev. Biol. Anim. 2016, 52, 287–295. [Google Scholar] [CrossRef]
- Hirano, T. Interleukin 6 in autoimmune and inflammatory diseases: A personal memoir. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 717–730. [Google Scholar] [CrossRef] [PubMed]
- Van Den Eeckhout, B.; Huyghe, L.; Van Lint, S.; Burg, E.; Plaisance, S.; Peelman, F.; Cauwels, A.; Uze, G.; Kley, N.; Gerlo, S.; et al. Selective IL-1 activity on CD8(+) T cells empowers antitumor immunity and synergizes with neovasculature-targeted TNF for full tumor eradication. J. Immunother. Cancer 2021, 9, e003293. [Google Scholar] [CrossRef] [PubMed]
- Zelova, H.; Hosek, J. TNF-alpha signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. 2013, 62, 641–651. [Google Scholar] [CrossRef]
- Ma, Y.F.; Tang, T.; Sheng, L.L.; Wang, Z.Q.; Tao, H.; Zhang, Q.; Zhang, Y.; Qi, Z.L. Aloin suppresses lipopolysaccharide-induced inflammation by inhibiting JAK1-STAT1/3 activation and ROS production in RAW264.7 cells. Int. J. Mol. Med. 2018, 42, 1925–1934. [Google Scholar] [CrossRef]
- Tang, X.; Marciano, D.L.; Leeman, S.E.; Amar, S. LPS induces the interaction of a transcription factor, LPS-induced TNF-alpha factor, and STAT6(B) with effects on multiple cytokines. Proc. Natl. Acad. Sci. USA 2005, 102, 5132–5137. [Google Scholar] [CrossRef]
- Banerjee, S.; Biehl, A.; Gadina, M.; Hasni, S.; Schwartz, D.M. JAK-STAT Signaling as a Target for Inflammatory and Autoimmune Diseases: Current and Future Prospects. Drugs 2017, 77, 1261. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Shin, J.S.; Chung, K.S.; Lee, Y.G.; Baek, N.I.; Lee, K.T. Anti-Inflammatory Mechanisms of Koreanaside A, a Lignan Isolated from the Flower of Forsythia koreana, against LPS-Induced Macrophage Activation and DSS-Induced Colitis Mice: The Crucial Role of AP-1, NF-kappaB, and JAK/STAT Signaling. Cells 2019, 8, 1163. [Google Scholar] [CrossRef] [PubMed]
- Huang, I.H.; Chung, W.H.; Wu, P.C.; Chen, C.B. JAK-STAT signaling pathway in the pathogenesis of atopic dermatitis: An updated review. Front. Immunol. 2022, 13, 1068260. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Oh, G.S.; Lee, J.H.; Lyu, A.R.; Ji, H.M.; Lee, S.H.; Song, J.; Park, S.J.; You, Y.O.; Sul, J.D.; et al. Cisplatin ototoxicity involves cytokines and STAT6 signaling network. Cell Res. 2011, 21, 944–956. [Google Scholar] [CrossRef] [PubMed]
- Heneghan, A.F.; Pierre, J.F.; Kudsk, K.A. IL-25 Improves IgA Levels During Parenteral Nutrition Through the JAK-STAT Pathway. Ann. Surg. 2013, 258, 1065–1071. [Google Scholar] [CrossRef]
- Hong, H.; Liao, S.; Chen, F.; Yang, Q.; Wang, D.Y. Role of IL-25, IL-33, and TSLP in triggering united airway diseases toward type 2 inflammation. Allergy 2020, 75, 2794–2804. [Google Scholar] [CrossRef]
- Salimi, M.; Barlow, J.L.; Saunders, S.P.; Xue, L.; Gutowska-Owsiak, D.; Wang, X.; Huang, L.C.; Johnson, D.; Scanlon, S.T.; McKenzie, A.N.; et al. A role for IL-25 and IL-33-driven type-2 innate lymphoid cells in atopic dermatitis. J. Exp. Med. 2013, 210, 2939–2950. [Google Scholar] [CrossRef]
- Lee, Y.; Choi, H.K.; N’Deh, K.P.; Choi, Y.J.; Fan, M.; Kim, E.K.; Chung, K.H.; An, A.J.H. Inhibitory Effect of Centella asiatica Extract on DNCB-Induced Atopic Dermatitis in HaCaT Cells and BALB/c Mice. Nutrients 2020, 12, 411. [Google Scholar] [CrossRef]
- Chrostowska-Plak, D.; Reich, A.; Szepietowski, J.C. Relationship between itch and psychological status of patients with atopic dermatitis. J. Eur. Acad. Dermatol. 2013, 27, E239–E242. [Google Scholar] [CrossRef]
- Moniaga, C.S.; Jeong, S.K.; Egawa, G.; Nakajima, S.; Hara-Chikuma, M.; Jeon, J.E.; Lee, S.H.; Hibino, T.; Miyachi, Y.; Kabashima, K. Protease activity enhances production of thymic stromal lymphopoietin and basophil accumulation in flaky tail mice. Am. J. Pathol. 2013, 182, 841–851. [Google Scholar] [CrossRef]
- Chen, L.Q.; Grabowski, K.A.; Xin, J.P.; Coleman, J.; Huang, Z.; Espiritu, B.; Alkan, S.; Xie, H.B.; Zhu, Y.C.; White, F.A.; et al. IL-4 induces differentiation and expansion of Th2 cytokine-producing eosinophils. J. Immunol. 2004, 172, 2059–2066. [Google Scholar] [CrossRef] [PubMed]
- Pope, S.M.; Brandt, E.B.; Mishra, A.; Hogan, S.P.; Zimmermann, N.; Matthaei, K.I.; Foster, P.S.; Rothenberg, M.E. IL-13 induces eosinophil recruitment into the lung by an IL-5- and eotaxin-dependent mechanism. J. Allergy Clin. Immunol. 2001, 108, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xian, Y.F.; Loo, S.K.F.; Ip, S.P.; Yang, W.; Chan, W.Y.; Lin, Z.X.; Wu, J.C.Y. Baicalin ameliorates 2,4-dinitrochlorobenzene-induced atopic dermatitis-like skin lesions in mice through modulating skin barrier function, gut microbiota and JAK/STAT pathway. Bioorg. Chem. 2022, 119, 105538. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, S.; Novak, N. Atopic dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Jung, H.S.; Kim, M.H.; Gwak, N.G.; Im, Y.S.; Lee, K.Y.; Sohn, Y.; Choi, H.; Yang, W.M. Antiallergic effects of Scutellaria baicalensis on inflammation in vivo and in vitro. J. Ethnopharmacol. 2012, 141, 345–349. [Google Scholar] [CrossRef]
- Song, J.; Zheng, H.; Seo, H.J.; Ji, G.E. Effect of Oral Administration of Root Extract on Atopic Dermatitis-like Skin Lesion Induced by Oxazolone in Hairless Mice. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 175–181. [Google Scholar] [CrossRef]
- Kwak, M.H.; Kim, J.E.; Hwang, I.S.; Lee, Y.J.; An, B.S.; Hong, J.T.; Lee, S.H.; Hwang, D.Y. Quantitative evaluation of therapeutic effect of Liriope platyphylla on phthalic anhydride-induced atopic dermatitis in IL-4/Luc/CNS-1 Tg mice. J. Ethnopharmacol. 2013, 148, 880–889. [Google Scholar] [CrossRef]
- Kim, M.J.; Yoo, Y.C.; Sung, N.Y.; Lee, J.; Park, S.R.; Shon, E.J.; Lee, B.D.; Kim, M.R. Anti-Inflammatory Effects of in LPS-Stimulated Macrophages and Endotoxemic Mice. Am. J. Chin. Med. 2016, 44, 1127–1143. [Google Scholar] [CrossRef]
- Yamaguchi-Miyamoto, T.; Kawasuji, T.; Kuraishi, Y.; Suzuki, H. Antipruritic effects of Sophora flavescens on acute and chronic itch-related responses in mice. Biol. Pharm. Bull. 2003, 26, 722–724. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.C.; Huang, W.C.; Chen, L.C.; Yeh, K.W.; Lin, C.F.; Liou, C.J. Sophoraflavanone G from Ameliorates Allergic Airway Inflammation by Suppressing Th2 Response and Oxidative Stress in a Murine Asthma Model. Int. J. Mol. Sci. 2022, 23, 6104. [Google Scholar] [CrossRef]
- Choi, M.; Yi, J.K.; Kim, S.Y.; Ryu, J.H.; Lee, J.; Kwon, W.; Jang, S.; Kim, D.; Kim, M.; Kim, H.; et al. Anti-inflammatory effects of a methanol extract of Dictamnus dasycarpus Turcz. root bark on imiquimod-induced psoriasis. BMC Complement. Altern. Med. 2019, 19, 347. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Nakano, Y.; Rahman, M.A.; Yatsuzuka, R.; Kamel, C. Effects of a T. extract on allergic models in mice. Biosci. Biotechnol. Biochem. 2008, 72, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Gong, Q.; Aoki, D.; Matsushita, Y.; Yoshida, M.; Taniguchi, T.; Endoh, K.; Fukushima, K. Microscopic distribution of alkaloids in freeze-fixed stems of Phellodendron amurense. Front. Plant Sci. 2023, 14, 1203768. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Shieh, D.E. The anti-inflammatory activity of Scutellaria rivularis extracts and its active components, baicalin, baicalein and wogonin. Am. J. Chin. Med. 1996, 24, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Huang, C.Q.; Wang, F.Q.; Dong, J.L.; Zhang, D.; Jiang, J.; Feng, Y.; Wu, B.; Xie, T.T.; Cheng, L. Qingxue jiedu formulation ameliorated DNFB-induced atopic dermatitis by inhibiting STAT3/MAPK/NF-κB signaling pathways. J. Ethnopharmacol. 2021, 270, 113773. [Google Scholar] [CrossRef] [PubMed]
- Inami, Y.; Andoh, T.; Kuraishi, Y. Prevention of Topical Surfactant Induced Itch-Related Responses by Chlorogenic Acid Through the Inhibition of Increased Histamine Production in the Epidermis. J. Pharmacol. Sci. 2013, 121, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.L.; Tan, J.; Zheng, Y. Chlorogenic Acid Alleviates Allergic Inflammatory Responses Through Regulating Th1/Th2 Balance in Ovalbumin-Induced Allergic Rhinitis Mice. Med. Sci. Monit. 2020, 26, e923358-1–e923358-10. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.J.; Kim, Y.W.; Park, Y.; Lee, H.J.; Kim, K.W. Anti-inflammatory effects of chlorogenic acid in lipopolysaccharide-stimulated RAW 264.7 cells. Inflamm. Res. 2014, 63, 81–90. [Google Scholar] [CrossRef]
- Han, X.W.; Ma, T.M.; Wang, Q.; Jin, C.L.; Han, Y.S.; Liu, G.J.; Li, H. The mechanism of oxymatrine on atopic dermatitis in mice based on SOCS1/JAK-STAT3 pathway. Front. Pharmacol. 2023, 13, 1091090. [Google Scholar] [CrossRef]
- Bai, Y.Q.; Wang, W.W.; Wang, L.; Ma, L.R.; Zhai, D.S.; Wang, F.R.; Shi, R.; Liu, C.Y.; Xu, Q.; Chen, G.; et al. Obacunone Attenuates Liver Fibrosis with Enhancing Anti-Oxidant Effects of GPx-4 and Inhibition of EMT. Molecules 2021, 26, 318. [Google Scholar] [CrossRef]
- Xiao, T.; Yu, X.; Yang, L.; Duan, X. Palmatine treats urticaria by reducing inflammation and increasing autophagy. Front. Immunol. 2023, 14, 1268467. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.J.; Ma, X.D.; Ai, G.X.; Yu, Q.X.; Chen, X.Y.; Yan, F.; Li, Y.C.; Xie, J.H.; Su, Z.R.; Xie, Q.F. Palmatine Protects Against MSU-Induced Gouty Arthritis via Regulating the NF-kappaB/NLRP3 and Nrf2 Pathways. Drug Des. Dev. Ther. 2022, 16, 2119–2132. [Google Scholar] [CrossRef] [PubMed]
- Garbers, C.; Aparicio-Siegmund, S.; Rose-John, S. The IL-6/gp130/STAT3 signaling axis: Recent advances towards specific inhibition. Curr. Opin. Immunol. 2015, 34, 75–82. [Google Scholar] [CrossRef]
- Kandhaya-Pillai, R.; Yang, X.; Tchkonia, T.; Martin, G.M.; Kirkland, J.L.; Oshima, J. TNF-alpha/IFN-gamma synergy amplifies senescence-associated inflammation and SARS-CoV-2 receptor expression via hyper-activated JAK/STAT1. Aging Cell 2022, 21, e13646. [Google Scholar] [CrossRef] [PubMed]
- Baldini, C.; Moriconi, F.R.; Galimberti, S.; Libby, P.; De Caterina, R. The JAK-STAT pathway: An emerging target for cardiovascular disease in rheumatoid arthritis and myeloproliferative neoplasms. Eur. Heart J. 2021, 42, 4389–4400. [Google Scholar] [CrossRef] [PubMed]
- Conde, E.; Bertrand, R.; Balbino, B.; Bonnefoy, J.; Stackowicz, J.; Caillot, N.; Colaone, F.; Hamdi, S.; Houmadi, R.; Loste, A.; et al. Dual vaccination against IL-4 and IL-13 protects against chronic allergic asthma in mice. Nat. Commun. 2021, 12, 2574. [Google Scholar] [CrossRef]
- Zhao, P.; Hiramoto, T.; Asano, Y.; Kubo, C.; Sudo, N. Chronic psychological stress exaggerates the compound 48/80-induced scratching behavior of mice. Pharmacol. Biochem. Behav. 2013, 105, 173–176. [Google Scholar] [CrossRef]
- Becerra-Diaz, M.; Valderrama-Carvajal, H.; Terrazas, L.I. Signal Transducers and Activators of Transcription (STAT) Family Members in Helminth Infections. Int. J. Biol. Sci. 2011, 7, 1371–1381. [Google Scholar] [CrossRef]
- Cardoso, C.R.; Provinciatto, P.R.; Godoi, D.F.; Fonseca, M.T.; Ferreira, B.R.; Teixeira, G.; Cunha, F.Q.; Pinzan, C.F.; da Silva, J.S. The signal transducer and activator of transcription 6 (STAT-6) mediates Th2 inflammation and tissue damage in a murine model of peanut-induced food allergy. Allergol. Immunopathol. 2019, 47, 535–543. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence |
|---|---|
| IL-1β | F 5′-ACCT GCT GGT GTG TGA CGT T-3′ R 5′-TCG TTG CTT GGT TCT CCT TG-3′ |
| IL-6 | F 5′-GAG GAT ACC ACT CCC AAC AGA CC-3′ R 5′-AAG TGC ATC ATC GTT GTT CAT ACA-3′ |
| TNF-α | F 5′-AAATGGGCTCCCTCTCATCAGTTC-3′ R 5′-TCTGCTTGGTGGTTTGCTACGAC-3′ |
| iNOS | F 5′-CAT GCT ACT GGA GGT GGG TG-3′ R 5′-CAT TGA TCT CCG TGA CAG CC-3′ |
| TSLP | F 5′-GGA CCA CTG GTG TTT ATT CT-3′ R 5′-CGA GGT TTA GAT GCT GTC AT-3′ |
| IL-4 | F 5′-AGA TGG ATG TGC CAA ACG TCC TCA-3′ R 5′-AAT ATG CGA AGC TTG GAA GCC-3′ |
| IL-13 | F 5′-GCA ACG GCA GCA TGG TAT GGA-3′ R 5′-TGG TAT AGG GGA GGC TGG AGA C-3′ |
| IL-25 | F 5′-ACA GGG ACT TGA ATC GGG TC-3′ R 5′-TGG TAA AGT GGG ACG GAG TTG-3′ |
| IL-33 | F 5′-CAC ATT GAG CAT CCA AGG AA-3′ R 5′-AAC AGA TTG GTC ATT GTA TGT ACT CAG-3′ |
| β-actin | F 5′-ATC ACT ATT GGC AAC GAG CG-3′ R 5′-TCA GCA ATG CCT GGG TAC AT-3′ |
| Standard Compounds | Oxymatrine | Chlorogenic Acid | Baicalin | Palmatine | Obacunone |
|---|---|---|---|---|---|
| Content (mg/g) | 2.52 ± 0.00 | 0.40 ± 0.00 | 2.61 ± 0.01 | 0.75 ± 0.00 | 0.36 ± 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-J.; Kim, S.-Y.; Bae, H.J.; Choi, Y.-Y.; An, J.-Y.; Cho, Y.E.; Cho, S.-Y.; Lee, S.-J.; Lee, S.; Sin, M.; et al. Anti-Inflammatory Effects of the LK5 Herbal Complex on LPS- and IL-4/IL-13-Stimulated HaCaT Cells and a DNCB-Induced Animal Model of Atopic Dermatitis in BALB/c Mice. Pharmaceutics 2024, 16, 40. https://doi.org/10.3390/pharmaceutics16010040
Kim H-J, Kim S-Y, Bae HJ, Choi Y-Y, An J-Y, Cho YE, Cho S-Y, Lee S-J, Lee S, Sin M, et al. Anti-Inflammatory Effects of the LK5 Herbal Complex on LPS- and IL-4/IL-13-Stimulated HaCaT Cells and a DNCB-Induced Animal Model of Atopic Dermatitis in BALB/c Mice. Pharmaceutics. 2024; 16(1):40. https://doi.org/10.3390/pharmaceutics16010040
Chicago/Turabian StyleKim, Hyun-Jeong, So-Yeon Kim, Ho Jung Bae, Yu-Yeong Choi, Ju-Yeon An, Ye Eun Cho, So-Young Cho, Su-Jung Lee, Sanghyun Lee, MinSub Sin, and et al. 2024. "Anti-Inflammatory Effects of the LK5 Herbal Complex on LPS- and IL-4/IL-13-Stimulated HaCaT Cells and a DNCB-Induced Animal Model of Atopic Dermatitis in BALB/c Mice" Pharmaceutics 16, no. 1: 40. https://doi.org/10.3390/pharmaceutics16010040