1. Introduction
Dental caries is the most prevalent infectious disease of the oral cavity. Biofilms formed by microorganisms over teeth and gum surfaces play an important causative role.
It is a well-known fact that biofilm prevents antibiotics from inhibiting bacterial growth and, thus, may contribute to the development of antibiotic resistance. Moreover, the widespread use of antibiotics results in an increase in more virulent and resistant bacteria. The emergence of antimicrobial agents’ resistant pathogenic bacteria has become a severe health concern [
1]; hence, new antibacterial agents are always beneficial.
One of the main etiological factors of dental caries is
Streptococcus mutans (S. mutans) [
2,
3,
4,
5,
6], a Gram-positive facultative anaerobic bacterium [
7,
8].
The pathogenicity of
S. mutans is inseparably associated with its ability to form biofilms on solid surfaces, such as implants and catheters [
2].
S. mutans has the ability to form extracellular polysaccharides (EPS), essential components in dental biofilms [
9].
S. mutans also generates glucan-binding proteins, which are assumed to encourage adherence to matrix glucans and influence the biofilm’s general structure [
10,
11]. Moreover, it has the ability to thrive in an acidic environment (aciduric) and metabolize carbohydrates, which produce lactic acids, leading to the development of dental tissue demineralization (acidogenic) [
2,
7].
Natural materials, herbs, spices, essential oils, etc., have been used in traditional medicine for many years [
12]. The cinnamon plant is one of these natural components. The main constituents of cinnamon essential oils and extracts are cinnamaldehyde, eugenol, phenol, and linalool [
13]. Trans-cinnamaldehyde (TC) is a phytochemical from the cinnamon plant.
TC, an α- and β-unsaturated aromatic aldehyde, offers numerous advantages. It can be obtained from a variety of sources, such as various vegetables and fruits, and it has a low cost and low cytotoxicity [
14].
Many studies have shown that cinnamaldehyde has medical and agricultural benefits. Gomez et al. [
15] proved that it inhibited fungal growth and aflatoxin production in food. Kim et al. [
16] showed how TC induces anti-inflammatory effects on the activation of macrophages stimulated via LPS. Lu et al. [
17] summarized cinnamaldehyde’s therapeutic effects and its related cardiovascular protective mechanisms.
In nature, cinnamaldehyde occurs as a trans-stereoisomer, namely, (2E)-3-phenylprop-2-enal or trans-cinnamaldehyde (TC) [
18]. However, when TC is exposed to air, it is quite unstable because the reactive unsaturated aldehyde is oxidized to cinnamic acid [
19]. This is due to its chemical structure of unstable bonds and being a member of the aldehyde group [
20].
This unsaturated aldehyde is the component of cinnamon that has established antibacterial properties for Gram-positive and -negative bacteria [
21]. TC has been shown to effectively inhibit many microorganisms’ growth and has been reported to inhibit toxin production by micro-organisms [
21].
Mu et al. recently reported their attempts to use chitosan-based nanocapsules loaded with TC against an
S. mutans biofilm [
22]. The nanoparticles adsorb to the bacterial membrane and act against
S. mutans.
Utilizing plants for antimicrobial agents is a reasonable strategy due to their natural production of a wide range of secondary metabolites that provide defenses against microbial invasions of the host. Additionally, unique chemical entities that exist in plants, such as complex structures, can limit the attempt to artificially produce them.
This capacity could enable these natural antibacterials to activate different modes of action in comparison to conventional antibacterial agents, thereby helping to reduce microbial resistance [
23].
In the present study, our general objective was to assess and establish the antibacterial effect of trans-cinnamaldehyde on S. mutans and its in vitro antibiofilm activity. We hypothesized that the TC tested would possess a potent antibacterial effect against S. mutans.
2. Materials and Methods
Trans-cinnamaldehyde (TC) (C
9H
80; C
6H
5CH=CHCHO), also called trans-3-Phenyl-2-propenal (trans-cinnamaldehyde 99%, Sigma-Aldrich, Jerusalem, Israel), was purchased. TC’s molecular structure is shown in
Figure 1 (molecular weight, 132.16 g/mol). The TC was diluted progressively to the following concentrations: 156.25 μg/mL, 312.5 μg/mL, 625 μg/mL, 1250 μg/mL, 2500 μg/mL, and 5000 μg/mL in dimethyl sulfoxide (DMSO) 0.03–1% (an organic solvent).
2.1. Bacterial Growth and Culture Conditions
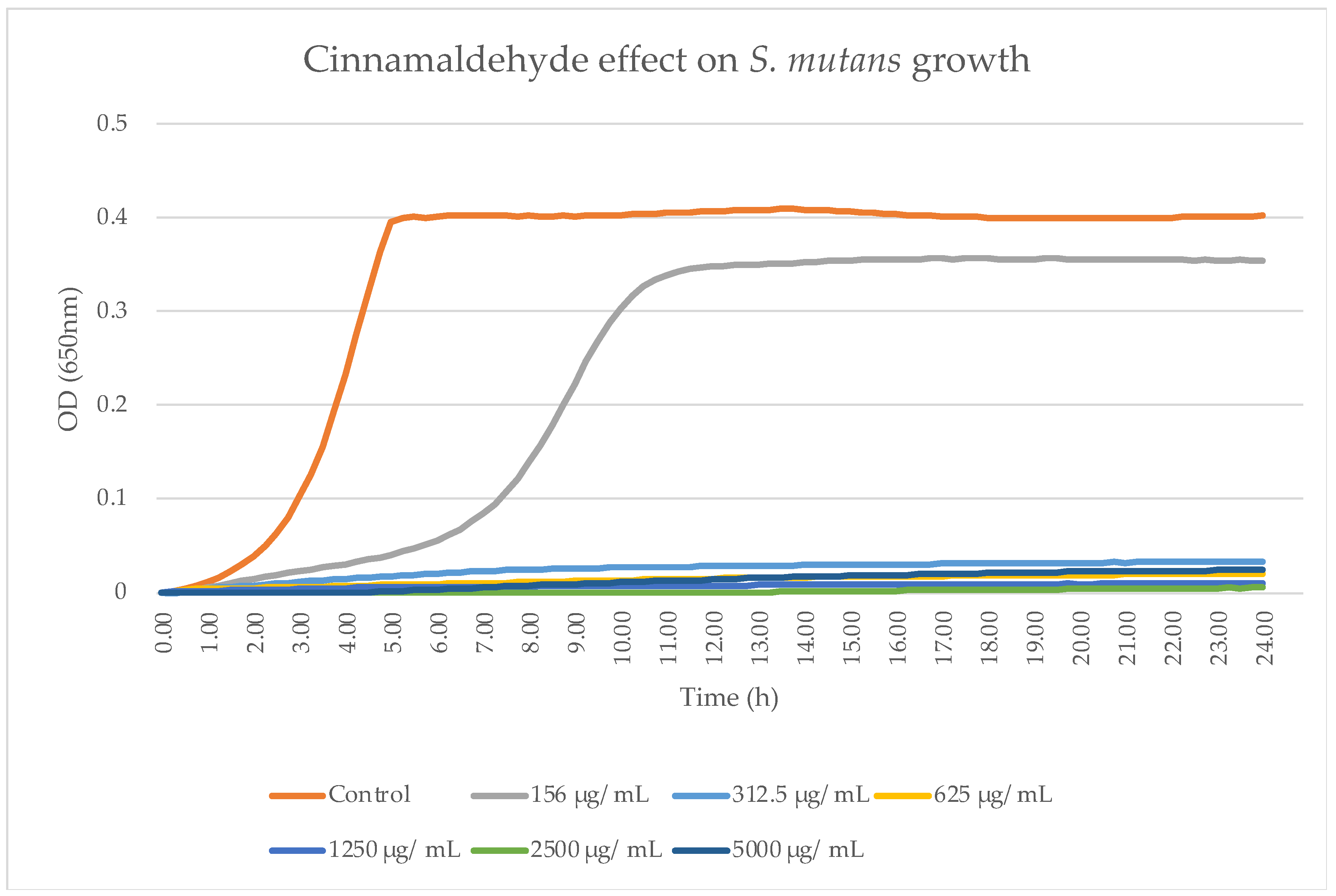
Planktonic S. mutans UA159 was grown overnight at 37 °C in brain–heart infusion broth (BHI, Acumedia, Lansing, MI, USA) in 95% air/5% CO2. An OD600 nm of 0.1 was determined as the initial concentration. The bacterial cultures were treated with the concentrations of TC mentioned above. Untreated bacteria served as the control, while chlorhexidine served as a positive control and BHI alone as a clear control. For kinetic studies, samples of S. mutans were treated with increasing concentrations of TC (156.25–5000 μg/mL) and the OD650 nm was measured every 15 min for 24 h using a microplate reader (Tecan M200, Tecan Trading AG, Männedorf, Switzerland) at 37 °C.
2.2. Viable Count Evaluations
The colony-forming unit (CFU) assay was carried out following various incubation times (0, 4 h, 8 h, 12 h, 16 h, 20 h, and 24 h) with TC in numerous concentrations (156.25–5000 μg/mL). Progressive serial dilutions were conducted on both untreated and treated samples. This process entailed transferring 100 μL from one sample to another tube containing 900 μL of phosphate-buffered saline (PBS). Following thorough vortexing, 10 μL of the bacterial suspension was subsequently plated in triplicate on brain–heart infusion (BHI) agar plates. The plates were then incubated overnight at 37 °C under conditions of 5% CO2.
2.3. Agar Diffusion Test (ADT)
The agar diffusion test is also known as the agar contact method. Following the inoculation of 100 μL 0.1 OD650 nm S. mutans on BHI–agar plates, we placed 10 μL of our different TC concentrations (156.25–5000 μg/mL) onto sterilized circular chromatography papers. This method involved the transfer, through diffusion, of TC to the plates inoculated with S. mutans and placed in a closed jar with a CO2 bag in the incubator at 37 °C. After 24 h, we evaluated the growth inhibition halo around the circular papers.
2.4. Biofilm Biomass Evaluation Using Crystal Violet (CV) Staining
For biofilm growth, S. mutans UA159 was grown overnight at 37 °C in 95% air/5% CO2 in brain–heart infusion and diluted to an OD600 nm of 0.1 in BHI containing 2.5% sucrose (BHIS), and then treated with increasing concentrations of TC (156.25–5000 μg/mL) with a positive control (chlorhexidine) and an untreated control for 24 h.
The resultant biofilms underwent staining with 200 μL of 0.1% crystal violet (CV), prepared from a 0.4% Gram’s crystal violet solution (Merck, EMD Millipore Corporation, Billerica, MA, USA) and diluted with double-distilled water (DDW). Following a 15 min incubation at room temperature, the CV solution was aspirated, and the wells were subjected to two washes with DDW and subsequently air-dried overnight. The extraction of the CV stain ensued through the addition of 150 μL of 33% acetic acid to the wells, accompanied by a 20 min incubation on an orbital shaker. The quantification of biofilm biomass was determined by measuring the absorbance at 595 nm using the M200 Tecan plate reader (Tecan Trading AG, Männedorf, Switzerland).
2.5. Biofilm Biomass Evaluation via Tetrazolium Reduction Assay (MTT Metabolic Assay)
The MTT assay is a colorimetric assay measuring bacterial metabolic activity. Following the S. mutans UA159 biofilm growth and incubation in the presence or absence of TC as described above, the MTT metabolic assay was performed. A 0.5 mg/mL MTT solution (Calbiochem, Darmstadt, Germany) in phosphate-buffered saline (PBS) was added to the biofilms in 96-well plates at a volume of 50 μL. After a 1 h incubation period at 37 °C, the wells underwent PBS washing. After an additional 1 h incubation period, absorbance readings were taken at 570 nm using the M200 Tecan plate reader (Tecan Trading AG, Männedorf, Switzerland) for quantitative assessment.
2.6. Metabolic Activity Assay (MTT) with Preformed Biofilms
S. mutans biofilm was grown in the manner previously mentioned, and then incubated for 4 h at 37 °C before being treated with increasing concentrations of TC (156.25–5000 μg/mL) with a positive control (chlorhexidine) and an untreated control for 24 h.
Then, the biofilms were washed twice with PBS, and MTT assays were performed. The wells were washed with PBS after 1 h of incubation at 37 °C. After another 1 h of incubation, the absorbance was measured at 570 nm using the M200 Tecan plate reader (Tecan Trading AG, Männedorf, Switzerland).
2.7. Biofilm Analysis via Spinning-Disk Confocal Microscopy (SDCM)
Spinning-disk confocal microscopy (SDCM) was employed to examine the biofilm architecture post-treatment with the assessed material, discerning live/dead bacterial presence and extracellular polysaccharides (EPS). The biofilms, cultivated on hydroxyapatite discs in 24-well tissue culture plates for 24 h in the absence or existence of varying TC concentrations, underwent double PBS washes.
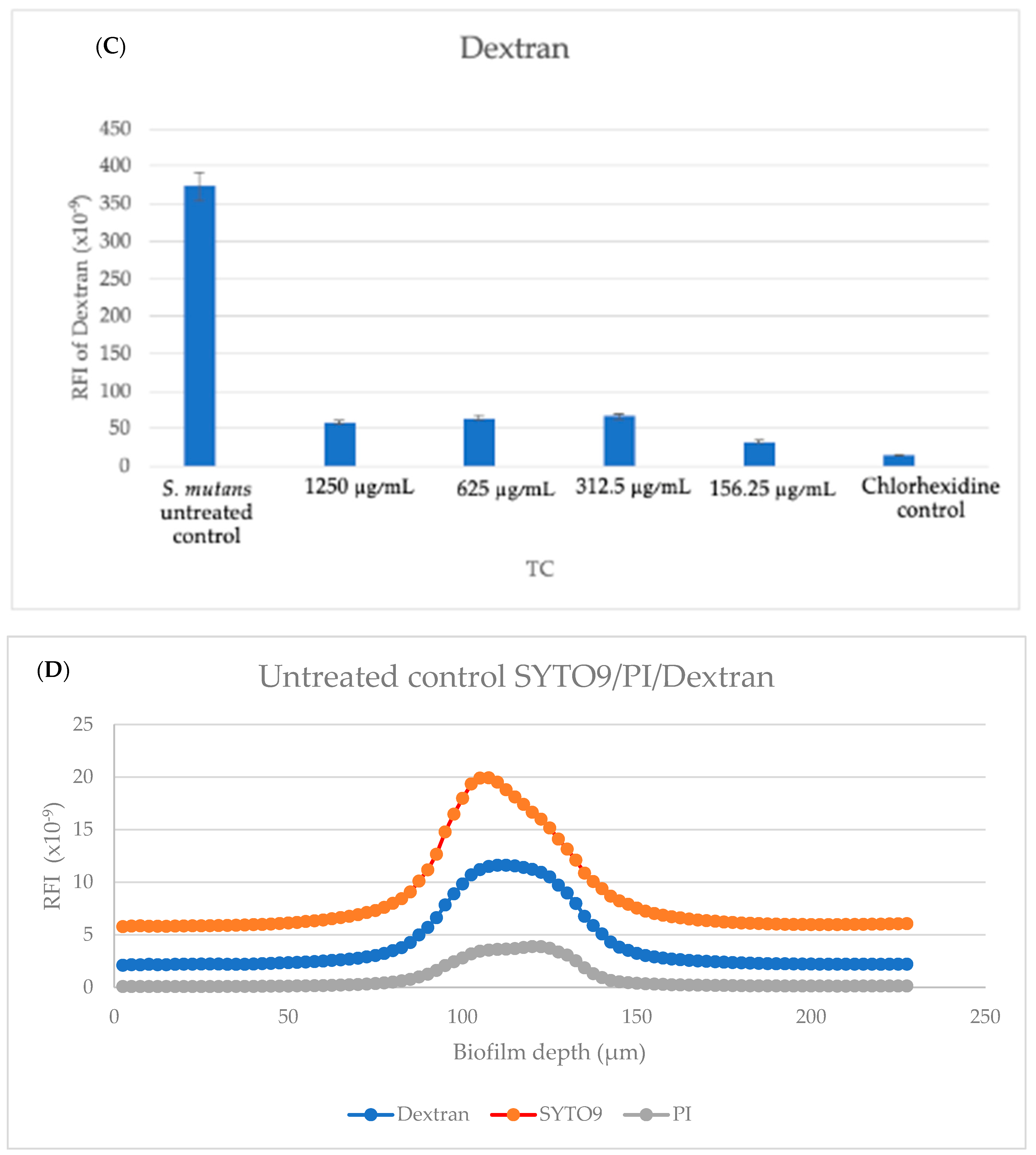
Subsequently, staining ensued with 3.3 μM SYTO 9, 10 μg/mL propidium iodide (PI), and 10 μg/mL Alexafluor647-conjugated dextran 10,000 for 20 min at room temperature.
SYTO 9’s green fluorescence (488 nm excitation, 515 nm emission) visualized both live and dead bacteria, while PI’s red fluorescence (543 nm excitation, 570 nm emission) specifically detected dead bacteria. Dextran’s blue fluorescence highlighted extracellular polysaccharides (EPS), presenting a fluorescence wavelength of 640 nm excitation and 665 nm emission for Alexafluor647. This resulted in a dual fluorescence, with live bacteria emitting green light and dead bacteria emitting both green and red light.
Utilizing the Nikon Yokogawa W1 spinning-disk confocal microscope, the samples were observed for thickness and bacterial vitality, capturing optical cross-sections at 2.5 μm intervals from the biofilm’s bottom to its apex.
2.8. High-Resolution Scanning Electron Microscopy (HR-SEM)
The biofilms were cultured on sterile hydroxyapatite discs, both with and without varying concentrations of TC. Following 24 h of incubation, the hydroxyapatite discs underwent rinsing with double-distilled water (DDW) and were subsequently fixed in 4% glutaraldehyde in DDW for 40 min. After an additional DDW wash, the discs were air-dried at room temperature. Subsequently, the hydroxyapatite discs were affixed to a metal stub, subjected to iridium sputter coating, and examined through a Magellan XHR 400 L high-resolution scanning electron microscope (Magellan XHR 400 L, FEI Company, Hillsboro, OR, USA). Three specimens from each treatment group were prepared and analyzed under SEM to assess the impact of TC on biofilm formation.
2.9. Statistical Analysis
Absorbance measurements were graphically represented, yielding bacterial growth curves for each well. The statistical examination concentrated on the linear segment of the logarithmic growth phase, where the slope denoted the bacterial growth rate and the intercept reflected the total viable bacterial count. The data analysis employed one-way ANOVA followed by Tukey’s post hoc test. The level of significance was p < 0.05.
4. Discussion
Trans-cinnamaldehyde is an effective and fast-acting antibacterial agent that can cause total bacterial growth inhibition using high concentrations. Partial bacterial growth inhibition was observed using all of the tested concentrations.
S. mutans is a Gram-positive bacterium that is a key contributor to dental caries and the formation of dental biofilms. It plays an important role in the production of pathogenic dental biofilms owing to its ability to produce extracellular polysaccharides [
9]. Gram-positive bacteria are more susceptible to phytochemicals and their constituents than Gram-negative bacteria due to the less complex cell walls of Gram-positive bacteria [
24]. Moreover, Gram-positive bacteria allow hydrophobic molecules, including TC, to easily pass through the cell membrane and operate on both the cell wall and the cytoplasm [
19].
To evaluate the antibacterial and antibiofilm activity of TC on S. mutans, we used our obtained sub-MBC doses. This is because the use of higher TC concentrations will lead to a bacteriocidic effect, which may cause dramatic changes in natural flora, leaving no bacteria to evaluate.
Our MBC against planktonic
S. mutans was 2500 μg/mL, which was higher than the 2000 μg/mL value obtained by He et al. [
14]. This could be due to technical differences such as the OD and TC concentrations used.
In terms of the antibiofilm activity, the 625 μg/mL TC dose was the lowest effective concentration, and this dose can be used for future translational studies.
We also observed a significant antibiofilm activity with the preformed (treatment) biofilm using our sub-MBC doses ≥ 312.5 μg/mL, similar to the results obtained in the metabolic assays. The ability of the TC to affect the biofilm after it was established was proven. These findings set the basis for future in vivo studies.
Additionally, biofilm formation is associated with quorum-sensing activity [
25]; hence, it may be reasonable to assume that this biofilm inhibition may lead to the downregulation of quorum-sensing systems (an intercellular communication system), making
S. mutans more prone to antibacterial agents and less resistant.
We observed no visible inhibition halo around disks with our TC concentrations; this could be due to the low solubility and hydrophobic nature of TC, which inhibits diffusion on sterilized circular chromatography papers.
Hydroxyapatite, a calcium phosphate compound with a calcium-to-phosphorus ratio of 1:67 and the chemical formula Ca
10 (PO
4) OH
2, is the main inorganic component of teeth and bone [
26]. The microscopic assays HR SEM and SDCM were conducted on hydroxyapatite disks, an essential component of tooth enamel, in order to simulate the effect in the oral cavity as much as possible. Through HR SEM and SDCM, morphological cell wall changes were observed, as well as bacterial aggregation and syncytium-like cell-wall fusion. No visible cell division was observed.
The use of 156.25 μg/mL and 312.5 μg/mL concentrations resulted in the partial inhibition of
S. mutans growth, as can be seen in
Figure 2,
Figure 3 and
Figure 7. Hence, the dramatic increase in the number of dead cells stained with PI was due to an overall higher number of dead cells. When using concentrations of 625 μg/mL and 1250 μg/mL, as expected, we observed more cell death and some (albeit less) biofilm activity. The 2500 μg/mL and 5000 μg/mL concentrations caused total bacterial kill after 24 h, and, thus, were not included in the following imaging experiments due to the fact that these concentrations resulted in almost no live bacteria, causing 99.9% of cell death at ≥ the MBC dose. When thinking of human clinical applications, we need to use the lowest TC concentration possible.
In
Figure 10f, an increase in EPS staining was observed, which was higher compared to the lower TC concentrations. This higher fluorescence observed in
Figure 10f is a three-dimensional representation of just one well, and the relative fluorescent intensities of the area under the curve are a better representation of our quantitative EPS measurement for the 1250 μg/mL concentration (
Figure 11C), which showed reduced EPS formation for all treatment groups, including 1250 μg/mL, compared to our untreated control.
EPS constitutes a diverse matrix of polymers, encompassing polysaccharides, proteins, glycoproteins, nucleic acids, phospholipids, and humic acids. Its documented role involves the establishment of a gel-like network, promoting bacterial cohesion within biofilms, facilitating biofilm adhesion to surfaces, and providing protection against adverse environmental conditions. Despite these recognized functions, the precise factors governing the augmentation of EPS formation remain incompletely elucidated. Polysaccharides can also provide protection from a wide range of stresses and it may be suggested that the stress formed by the TC can cause a temporary increase in EPS.
Balasubramanian et al. [
27] suggested a synergistic effect against cariogenic bacteria when combining TC with chlorohexidine or fluoride.
As reported in the literature, cinnamon essential oils may also be beneficial in the treatment of halitosis, while reducing biofilm formation [
13]. The use of the sub-MBC that was evaluated in this research may also be useful for the treatment of halitosis and caries, with a possible minimal effect on the microbial oral population. This assumption should be further investigated clinically.
Previous study found that TC decreased
S. mutans activity and affected biofilm formation. TC increased the cell surface hydrophobicity and reduced bacterial aggregation, thus affecting the process of biofilm formation. It also inhibited the acid production and acid tolerance of
S. mutans. In the presence of cinnamaldehyde, the gene expression in the biofilms was reduced [
14].
Hence, TC, derived from a natural substance, has shown significant potential to serve as an antimicrobial modality, particularly in inhibiting caries-promoting bacterial growth.
Dentifrices comprise several antimicrobial agents, such as fluoride and zinc salts. Given the antibacterial properties we have shown, it may be beneficial to add TC to dentifrice, lozenges, mouthwashes, etc., as a natural antimicrobial agent.
An important issue that must be considered is the large amount of evidence in the literature regarding allergy to cinnamon [
28,
29,
30]. Cinnamaldehyde is a known sensitizer in humans and it has been reported that cinnamaldehyde is a frequent cause of allergic reactions. In human dermatological studies, the level for cinnamaldehyde sensitization has been set at 0.5% [
21]. Thus, this highlights the need for research into cinnamaldehyde derivates using the lowest concentration possible to achieve good antibacterial effects. Therefore, care must be taken when using TC as a caries-preventive modality.
Further studies are needed to prove the anti-cariogenic potential of TC, with or without different antibacterial therapeutics.

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}













