
Retinoic Acid Receptor Is a Novel Therapeutic Target for Postoperative Cognitive Dysfunction


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
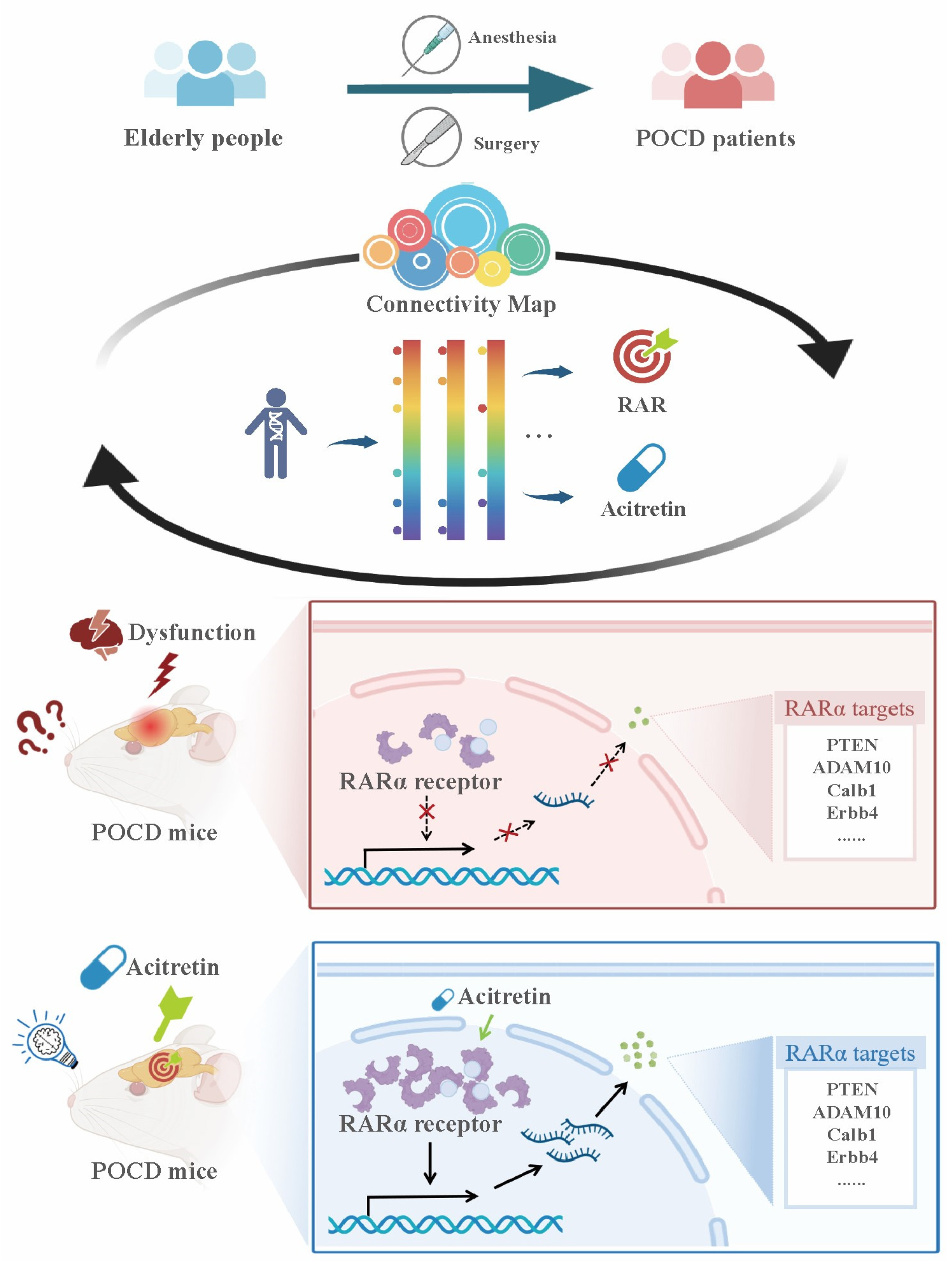
2.2. CMap Analysis of POCD Signature
2.3. Animals
2.4. Anesthesia and Surgical Procedure
2.5. Physiological Parameters Measurements
2.6. Behavioral Tests
2.7. Brain Tissue Harvest
2.8. Western Blotting Analysis
2.9. qRT-PCR
2.10. Data Analysis and Statistics
3. Results
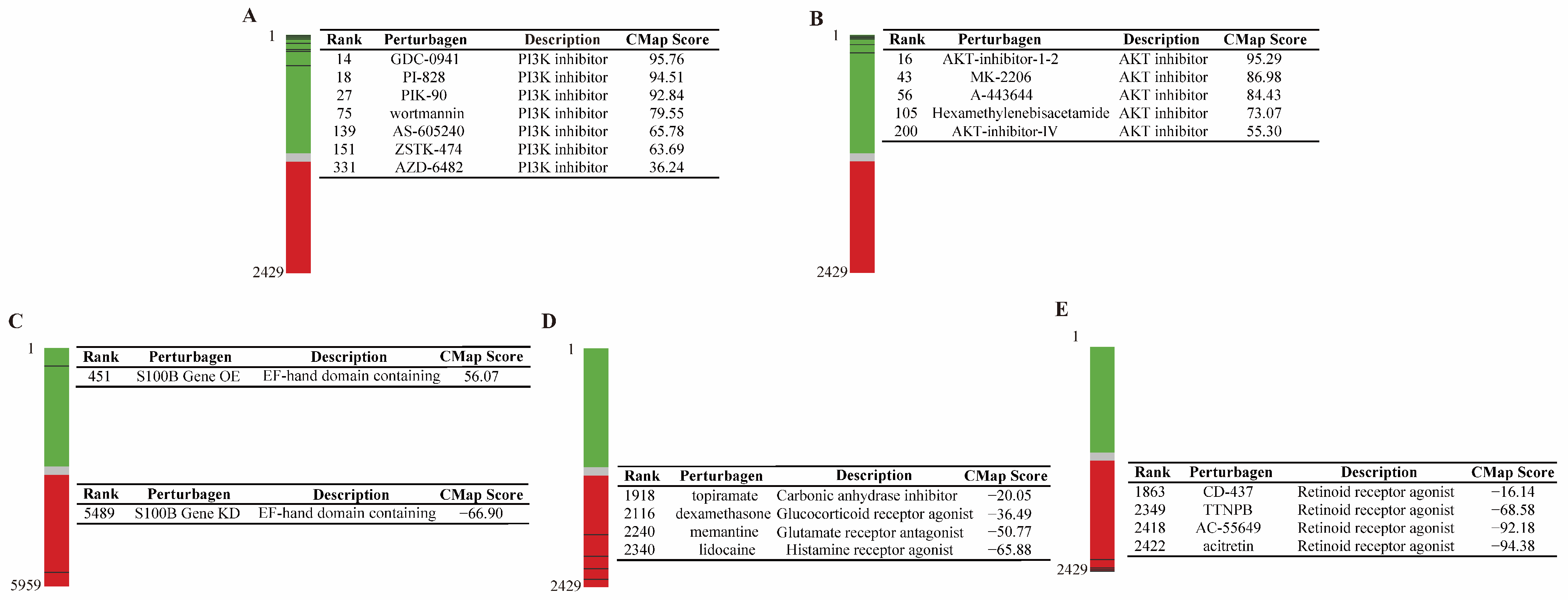
3.1. The Reliability and Predictability for the CMap Model of POCD
3.2. RAR Was Predicated as a Novel Therapeutic Target for POCD
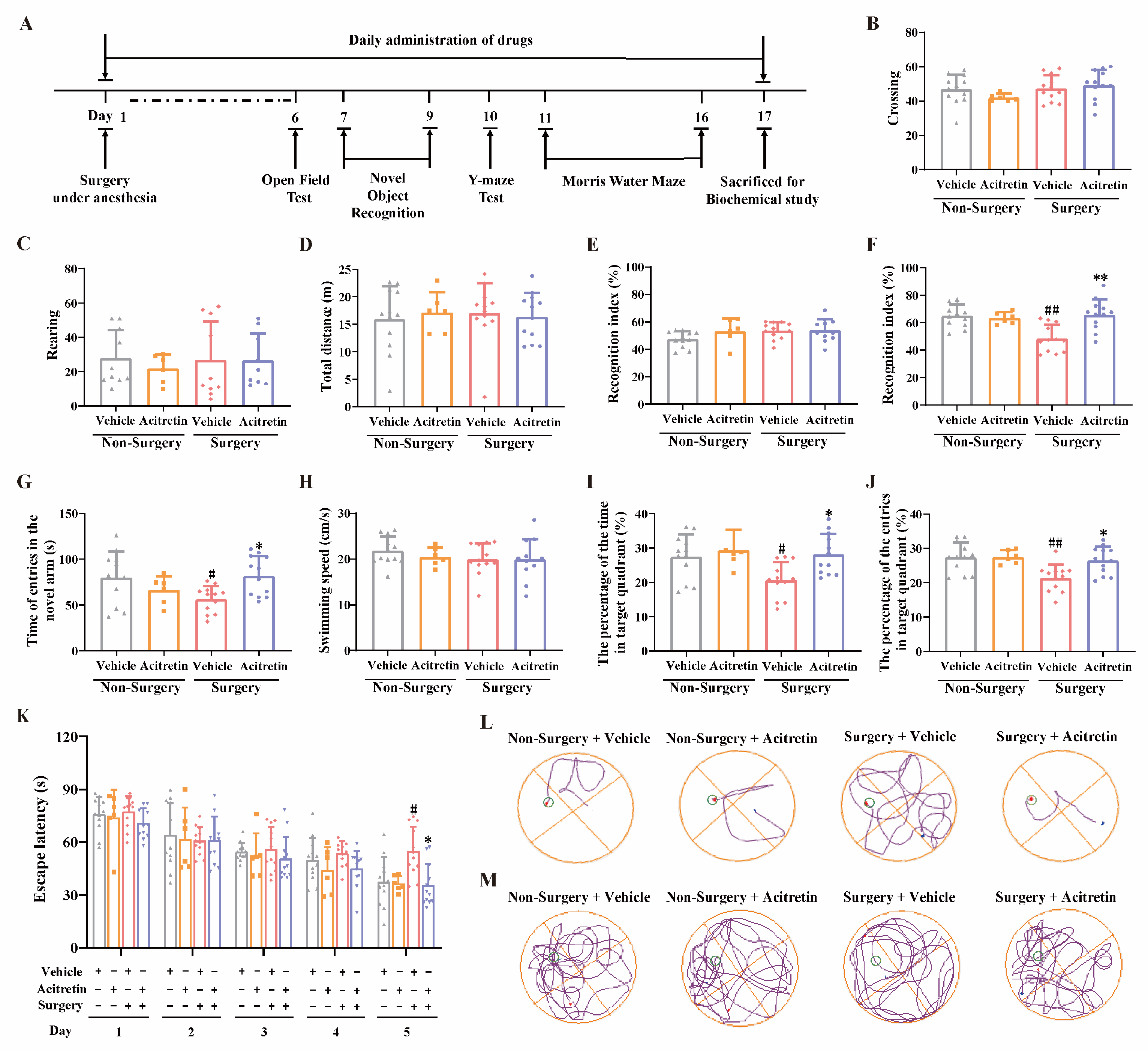
3.3. Acitretin Alleviated Surgery-Induced Cognitive Dysfunction in Aged Mice
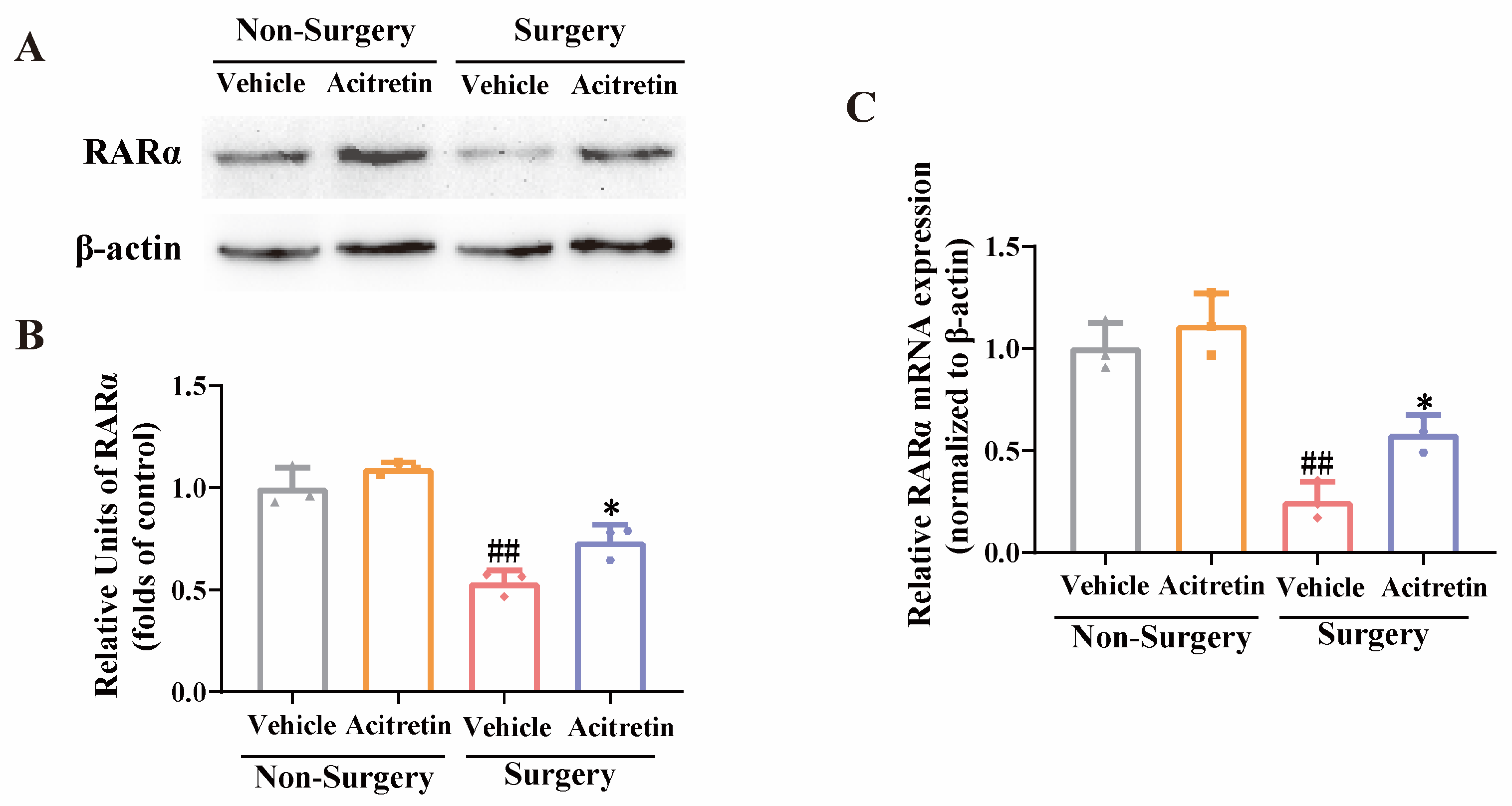
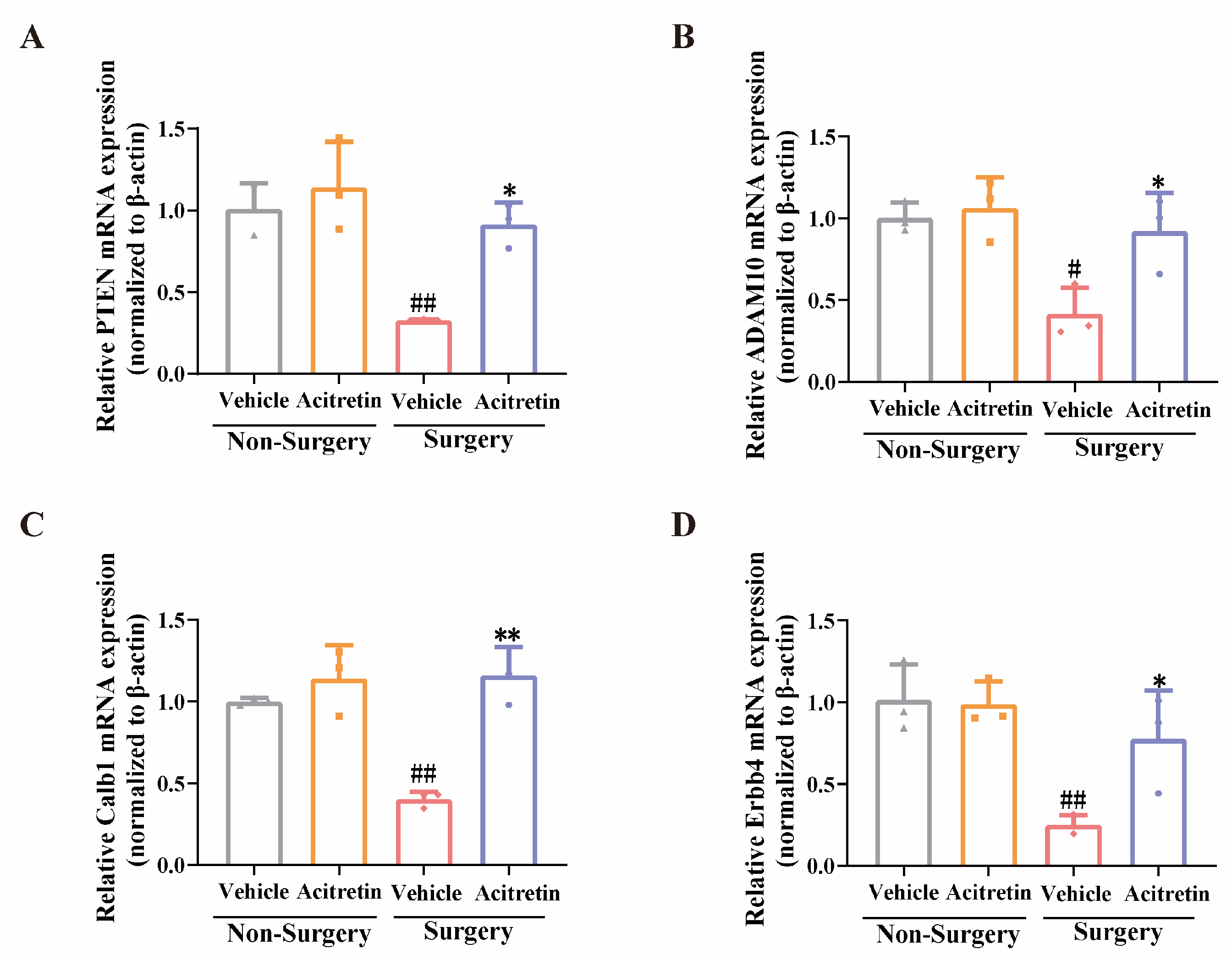
3.4. Acitretin Attenuated the Reduction of RARα Expression and Increased mRNA Expression of RARα-Target Genes in Aged Mice after Surgery
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, B.; Yuan, H.; Zhai, X.; Li, X.; Qin, J.; Chen, J.; Meng, B. High-Pressure Pneumoperitoneum Aggravates Surgery-Induced Neuroinflammation and Cognitive Dysfunction in Aged Mice. Mediat. Inflamm. 2020, 2020, 6983193. [Google Scholar] [CrossRef] [PubMed]
- Boone, M.D.; Sites, B.; von Recklinghausen, F.M.; Mueller, A.; Taenzer, A.H.; Shaefi, S. Economic Burden of Postoperative Neurocognitive Disorders Among US Medicare Patients. JAMA Netw. Open 2020, 3, e208931. [Google Scholar] [CrossRef]
- Arora, S.S.; Gooch, J.L.; García, P.S. Postoperative cognitive dysfunction, Alzheimer’s disease, and anesthesia. Int. J. Neurosci. 2014, 124, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Huang, K.; Zhu, B.; Zhou, B.; Ahmad Harb, A.K.; Liu, L.; Wu, X. Neuropsychological Tests in Post-operative Cognitive Dysfunction: Methods and Applications. Front. Psychol. 2021, 12, 684307. [Google Scholar] [CrossRef] [PubMed]
- Okumura, D.; Hagino, M.; Yamagishi, A.; Kaibori, Y.; Munira, S.; Saito, Y.; Nakayama, Y. Inhibitors of the VEGF Receptor Suppress HeLa S3 Cell Proliferation via Misalignment of Chromosomes and Rotation of the Mitotic Spindle, Causing a Delay in M-Phase Progression. Int. J. Mol. Sci. 2018, 19, 4014. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Ma, Q.; Ma, D. Low-dose Sevoflurane Attenuates Cardiopulmonary Bypass (CPB)- induced Postoperative Cognitive Dysfunction (POCD) by Regulating Hippocampus Apoptosis via PI3K/AKT Pathway. Curr. Neurovascular Res. 2020, 17, 232–240. [Google Scholar] [CrossRef]
- Kim, J.H.; Jung, H.; Lee, Y.; Sohn, J.H. Surgery Performed Under Propofol Anesthesia Induces Cognitive Impairment and Amyloid Pathology in ApoE4 Knock-In Mouse Model. Front. Aging Neurosci. 2021, 13, 658860. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.X.; Xiao, Q.X.; Liu, Q.; Deng, R.; Bian, J.; Deng, I.B.; Al-Hawwas, M.; Yu, F.X. Microarray Expression Profiles of lncRNAs and mRNAs in Postoperative Cognitive Dysfunction. Front. Neurosci. 2018, 12, 694. [Google Scholar] [CrossRef]
- Liang, X.; Zhu, W.; Lv, Z.; Zou, Q. Molecular Computing and Bioinformatics. Molecules 2019, 24, 2358. [Google Scholar] [CrossRef]
- Luo, S.; Li, H.; Mo, Z.; Lei, J.; Zhu, L.; Huang, Y.; Fu, R.; Li, C.; Huang, Y.; Liu, K.; et al. Connectivity map identifies luteolin as a treatment option of ischemic stroke by inhibiting MMP9 and activation of the PI3K/Akt signaling pathway. Exp. Mol. Med. 2019, 51, 1–11. [Google Scholar] [CrossRef]
- Lee, S.Y.; Song, M.Y.; Kim, D.; Park, C.; Park, D.K.; Kim, D.G.; Yoo, J.S.; Kim, Y.H. A Proteotranscriptomic-Based Computational Drug-Repositioning Method for Alzheimer’s Disease. Front. Pharmacol. 2019, 10, 1653. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Narayan, R.; Corsello, S.M.; Peck, D.D.; Natoli, T.E.; Lu, X.; Gould, J.; Davis, J.F.; Tubelli, A.A.; Asiedu, J.K.; et al. A Next Generation Connectivity Map: L1000 Platform and the First 1,000,000 Profiles. Cell 2017, 171, 1437–1452.e1417. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wu, X.; Ye, L.; Bai, Y.; Zhang, H.; Xuan, Z.; Feng, Y.; Zhang, P.; Chen, Y.; Yan, Y.; et al. Edaravone at high concentrations attenuates cognitive dysfunctions induced by abdominal surgery under general anesthesia in aged mice. Metab. Brain Dis. 2020, 35, 373–383. [Google Scholar] [CrossRef]
- Wang, R.P.; Huang, J.; Chan, K.W.Y.; Leung, W.K.; Goto, T.; Ho, Y.S.; Chang, R.C. IL-1β and TNF-α play an important role in modulating the risk of periodontitis and Alzheimer’s disease. J. Neuroinflamm. 2023, 20, 71. [Google Scholar] [CrossRef] [PubMed]
- Sohn, E.; Lim, H.S.; Kim, Y.J.; Kim, B.Y.; Kim, J.H.; Jeong, S.J. Elaeagnus glabra f. oxyphylla Attenuates Scopolamine-Induced Learning and Memory Impairments in Mice by Improving Cholinergic Transmission via Activation of CREB/NGF Signaling. Nutrients 2019, 11, 1205. [Google Scholar] [CrossRef]
- Mou, C.Y.; Xie, Y.F.; Wei, J.X.; Wang, Q.Y.; Le, J.Y.; Bao, Y.J.; Zhang, P.P.; Mao, Y.C.; Huang, X.H.; Pan, H.B.; et al. Rose Bengal inhibits β-amyloid oligomers-induced tau hyperphosphorylation via acting on Akt and CDK5 kinases. Psychopharmacology 2022, 239, 3579–3593. [Google Scholar] [CrossRef]
- di Masi, A.; Leboffe, L.; De Marinis, E.; Pagano, F.; Cicconi, L.; Rochette-Egly, C.; Lo-Coco, F.; Ascenzi, P.; Nervi, C. Retinoic acid receptors: From molecular mechanisms to cancer therapy. Mol. Aspects Med. 2015, 41, 1–115. [Google Scholar] [CrossRef]
- Uva, P.; Bosco, M.C.; Eva, A.; Conte, M.; Garaventa, A.; Amoroso, L.; Cangelosi, D. Connectivity Map Analysis Indicates PI3K/Akt/mTOR Inhibitors as Potential Anti-Hypoxia Drugs in Neuroblastoma. Cancers 2021, 13, 2809. [Google Scholar] [CrossRef]
- Lee, J.Y.; Gallo, R.A.; Ledon, P.J.; Tao, W.; Tse, D.T.; Pelaez, D.; Wester, S.T. Integrating Differential Gene Expression Analysis with Perturbagen-Response Signatures May Identify Novel Therapies for Thyroid-Associated Orbitopathy. Transl. Vis. Sci. Technol. 2020, 9, 39. [Google Scholar] [CrossRef]
- Han, X.; Cheng, X.; Xu, J.; Liu, Y.; Zhou, J.; Jiang, L.; Gu, X.; Xia, T. Activation of TREM2 attenuates neuroinflammation via PI3K/Akt signaling pathway to improve postoperative cognitive dysfunction in mice. Neuropharmacology 2022, 219, 109231. [Google Scholar] [CrossRef]
- Lv, Z.; Che, L.; Du, Y.; Yu, J.; Su, E.; Liu, H.; Chen, D. Mechanism of Mongolian Medicine Eerdun Wurile in Improving Postoperative Cognitive Dysfunction Through Activation of the PI3K Signaling Pathway. Front. Neurosci. 2021, 15, 769759. [Google Scholar] [CrossRef]
- Wang, W.; Ma, Y.; Liu, Y.; Wang, P.; Liu, Y. Effects of Dexmedetomidine Anesthesia on Early Postoperative Cognitive Dysfunction in Elderly Patients. ACS Chem. Neurosci. 2022, 13, 2309–2314. [Google Scholar] [CrossRef]
- Peng, L.; Xu, L.; Ouyang, W. Role of peripheral inflammatory markers in postoperative cognitive dysfunction (POCD): A meta-analysis. PLoS ONE 2013, 8, e79624. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.N.; Whiting, A.; McCaffery, P. Retinoic acid receptor-targeted drugs in neurodegenerative disease. Expert. Opin. Drug Metab. Toxicol. 2020, 16, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Wołoszynowska-Fraser, M.U.; Kouchmeshky, A.; McCaffery, P. Vitamin A and Retinoic Acid in Cognition and Cognitive Disease. Annu. Rev. Nutr. 2020, 40, 247–272. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; van Dam, A.M.; Wang, Y.; Lucassen, P.J.; Zhou, J.N. Retinoic acid and depressive disorders: Evidence and possible neurobiological mechanisms. Neurosci. Biobehav. Rev. 2020, 112, 376–391. [Google Scholar] [CrossRef]
- Jarvis, C.I.; Goncalves, M.B.; Clarke, E.; Dogruel, M.; Kalindjian, S.B.; Thomas, S.A.; Maden, M.; Corcoran, J.P. Retinoic acid receptor-α signalling antagonizes both intracellular and extracellular amyloid-β production and prevents neuronal cell death caused by amyloid-β. Eur. J. Neurosci. 2010, 32, 1246–1255. [Google Scholar] [CrossRef]
- Holthoewer, D.; Endres, K.; Schuck, F.; Hiemke, C.; Schmitt, U.; Fahrenholz, F. Acitretin, an enhancer of alpha-secretase expression, crosses the blood-brain barrier and is not eliminated by P-glycoprotein. Neurodegener. Dis. 2012, 10, 224–228. [Google Scholar] [CrossRef]
- Endres, K.; Fahrenholz, F.; Lotz, J.; Hiemke, C.; Teipel, S.; Lieb, K.; Tüscher, O.; Fellgiebel, A. Increased CSF APPs-α levels in patients with Alzheimer disease treated with acitretin. Neurology 2014, 83, 1930–1935. [Google Scholar] [CrossRef]
- Sodhi, R.K.; Singh, N. Retinoids as potential targets for Alzheimer’s disease. Pharmacol. Biochem. Behav. 2014, 120, 117–123. [Google Scholar] [CrossRef]
- Yuan, X.Z.; Sun, S.; Tan, C.C.; Yu, J.T.; Tan, L. The Role of ADAM10 in Alzheimer’s Disease. J. Alzheimers Dis. 2017, 58, 303–322. [Google Scholar] [CrossRef] [PubMed]
- Spina Nagy, G.; Kawamoto, E.M.; Bridi, J.C. The role of PTEN signaling in synaptic function: Implications in autism spectrum disorder. Neurosci. Lett. 2021, 759, 136015. [Google Scholar] [CrossRef] [PubMed]
- Qi, D.; Hou, X.; Jin, C.; Chen, X.; Pan, C.; Fu, H.; Song, L.; Xue, J. HNSC exosome-derived MIAT improves cognitive disorders in rats with vascular dementia via the miR-34b-5p/CALB1 axis. Am. J. Transl. Res. 2021, 13, 10075–10093. [Google Scholar] [PubMed]
- Audisio, C.; Nicolino, S.; Scevola, A.; Tos, P.; Geuna, S.; Battiston, B.; Perroteau, I. ErbB receptors modulation in different types of peripheral nerve regeneration. Neuroreport 2008, 19, 1605–1609. [Google Scholar] [CrossRef] [PubMed]
- Sierra-Mondragon, E.; Rodríguez-Muñoz, R.; Namorado-Tonix, C.; Molina-Jijon, E.; Romero-Trejo, D.; Pedraza-Chaverri, J.; Reyes, J.L. All-Trans Retinoic Acid Attenuates Fibrotic Processes by Downregulating TGF-β1/Smad3 in Early Diabetic Nephropathy. Biomolecules 2019, 9, 525. [Google Scholar] [CrossRef]
- Singh, A.B.; Guleria, R.S.; Nizamutdinova, I.T.; Baker, K.M.; Pan, J. High glucose-induced repression of RAR/RXR in cardiomyocytes is mediated through oxidative stress/JNK signaling. J. Cell. Physiol. 2012, 227, 2632–2644. [Google Scholar] [CrossRef]
- Lin, X.; Chen, Y.; Zhang, P.; Chen, G.; Zhou, Y.; Yu, X. The potential mechanism of postoperative cognitive dysfunction in older people. Exp. Gerontol. 2020, 130, 110791. [Google Scholar] [CrossRef]
- Wang, P.; Li, R.; Yuan, Y.; Zhu, M.; Liu, Y.; Jin, Y.; Yin, Y. PTENα is responsible for protection of brain against oxidative stress during aging. FASEB J. 2021, 35, e21943. [Google Scholar] [CrossRef]
- Xu, J.; Hu, C.; Chen, S.; Shen, H.; Jiang, Q.; Huang, P.; Zhao, W. Neuregulin-1 protects mouse cerebellum against oxidative stress and neuroinflammation. Brain Res. 2017, 1670, 32–43. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer ID | Sequences |
|---|---|---|
| RARα | 220406_001A01 | Forward: 5′-TTCTTTCCCCCTATGCTGGGT-3′ |
| 220406_001B01 | Reverse: 5′-GGGAGGGCTGGGTACTATCTC-3′ | |
| PTEN | 211018_005C02 | Forward: 5′-TGGATTCGACTTAGACTTGACCT-3′ |
| 211018_005D02 | Reverse: 5′-GCGGTGTCATAATGTCTCTCAG-3′ | |
| ADAM10 | 211018_005E01 | Forward: 5′-GGGAAGAAATGCAAGCTGAA-3′ |
| 211018_005F01 | Reverse: 5′-CTGTACAGCAGGGTCCTTGAC-3′ | |
| Calb1 | 211018_005A02 | Forward: 5′-TCTGGCTTCATTTCGACGCTG-3′ |
| 211018_005B02 | Reverse: 5′-ACAAAGGATTTCATTTCCGGTGA-3′ | |
| Erbb4 | 211018_005G01 | Forward: 5′-CCTTCCTGCGGTCTATCCGA-3′ |
| 211018_005H01 | Reverse: 5′-CCAAAGTTGCCATCTTTCCTGTA-3′ | |
| β-actin | 220725_002B07 | Forward: 5′-CCTAAGAGGAGGATGGTCGCR-3′ |
| 220725_002C07 | Reverse: 5′-CCTAAGAGGAGGATGGTCGC-3′ |
| Parameters | Group | Anesthesia Duration (min) | Post-Anesthesia | ||
|---|---|---|---|---|---|
| 0 | 10 | 20 | |||
| Arterial O2 saturation (%) | Anesthesia | 94.70 ± 1.15 | 94.64 ± 3.80 | 94.94 ± 3.50 | 96.17 ± 2.44 |
| Anesthesia + Surgery | 92.90 ± 0.40 | 93.20 ± 1.02 | 95.85 ± 2.23 | 94.46 ± 1.19 | |
| Heart rate (bpm) | Anesthesia | 413.6 ± 52.8 | 443.5 ± 82.8 | 539.1 ± 117.3 | 549.1 ± 136.9 |
| Anesthesia + Surgery | 541.3 ± 103.1 | 532.8 ± 80.5 | 527.5 ± 60.1 | 511.3 ±70.2 | |
| Breath rate (brpm) | Anesthesia | 133.4 ± 35.1 | 147.5 ± 45.2 | 112.7 ± 7.74 | 124.0 ± 9.29 |
| Anesthesia + Surgery | 112.2 ± 11.3 | 112.7 ± 18.3 | 107.2 ± 17.2 | 139.8 ± 67.9 | |
| Pulse distention (μm) | Anesthesia | 517.3 ± 150.3 | 463.2 ± 152.2 | 487.3 ± 192.5 | 396.9 ± 157.7 |
| Anesthesia + Surgery | 431.5 ± 193.1 | 425.6 ± 182.7 | 435.2 ± 237.8 | 397.3 ± 243.5 | |
| Rectal temperature (°C) | Anesthesia | 38.45 ± 0.86 | 38.18 ± 0.77 | 38.37 ± 0.20 | 38.99 ± 0.20 |
| Anesthesia + Surgery | 38.17 ± 0.33 | 38.56 ± 0.39 | 38.60 ± 0.44 | 38.72 ± 0.48 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, Y.; Rong, W.; Zhu, A.; Chen, Y.; Chen, H.; Hong, Y.; Le, J.; Wang, Q.; Naman, C.B.; Xu, Z.; et al. Retinoic Acid Receptor Is a Novel Therapeutic Target for Postoperative Cognitive Dysfunction. Pharmaceutics 2023, 15, 2311. https://doi.org/10.3390/pharmaceutics15092311
Bao Y, Rong W, Zhu A, Chen Y, Chen H, Hong Y, Le J, Wang Q, Naman CB, Xu Z, et al. Retinoic Acid Receptor Is a Novel Therapeutic Target for Postoperative Cognitive Dysfunction. Pharmaceutics. 2023; 15(9):2311. https://doi.org/10.3390/pharmaceutics15092311
Chicago/Turabian StyleBao, Yongjie, Wenni Rong, An Zhu, Yuan Chen, Huiyue Chen, Yirui Hong, Jingyang Le, Qiyao Wang, C. Benjamin Naman, Zhipeng Xu, and et al. 2023. "Retinoic Acid Receptor Is a Novel Therapeutic Target for Postoperative Cognitive Dysfunction" Pharmaceutics 15, no. 9: 2311. https://doi.org/10.3390/pharmaceutics15092311