
Long-Term Survival and Induction of Operational Tolerance to Murine Islet Allografts by Co-Transplanting Cyclosporine A Microparticles and CTLA4-Ig
 , , , , , and
, , , , , and 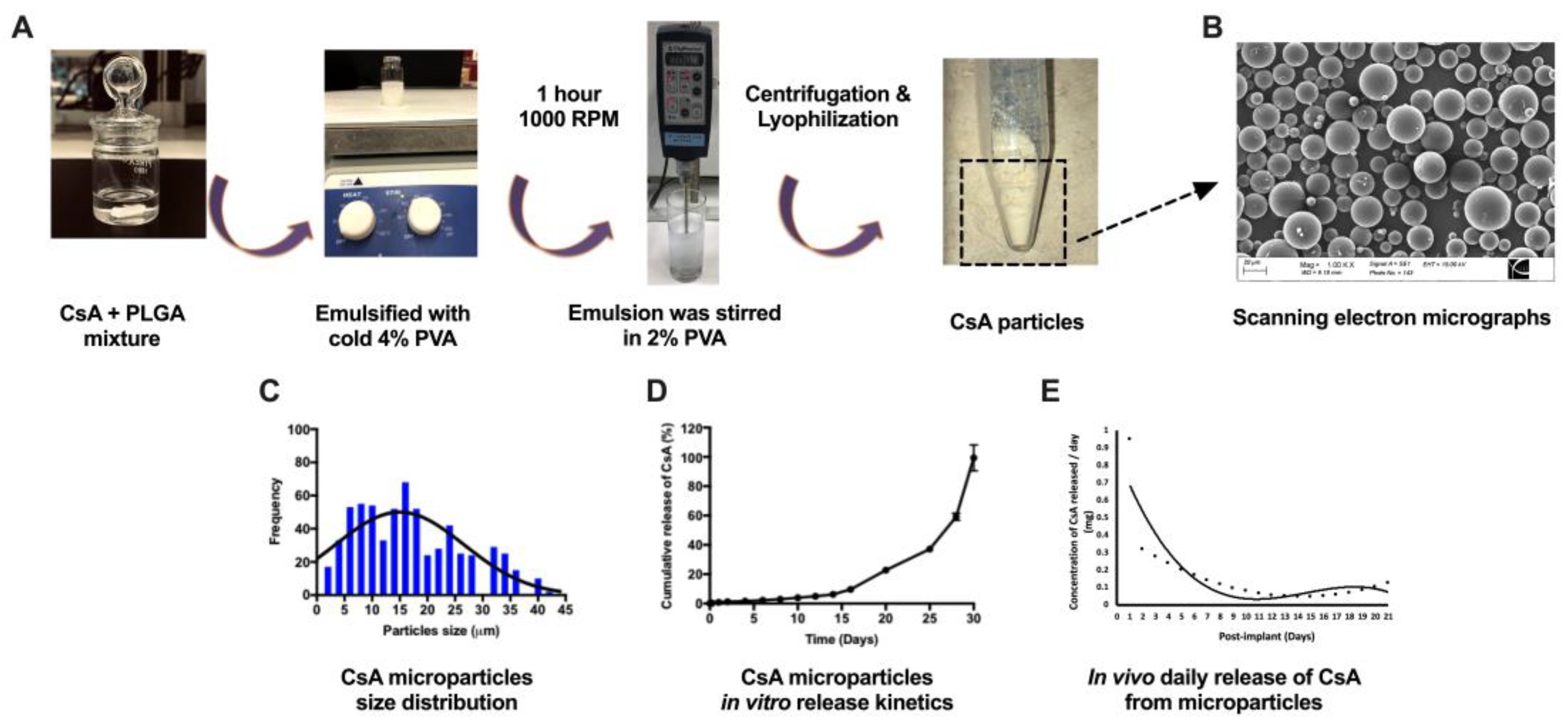{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation and Characterization of CSA-Eluting PLGA Microparticles
2.2. In Vivo Drug Release Characterization
2.3. Islet Isolation, Transplantation, and Metabolic Follow-Up
2.4. Intra-Islet Graft Proinflammatory Cytokine Analysis and Gene Expression Analysis
2.5. Islet Graft Immunohistochemical Analysis
2.6. Assessment of Tolerance Induction by Allogeneic Skin Graft Transplantation
2.7. Statistical Analysis
3. Results
3.1. CsA Microparticle Preparation and Characterization
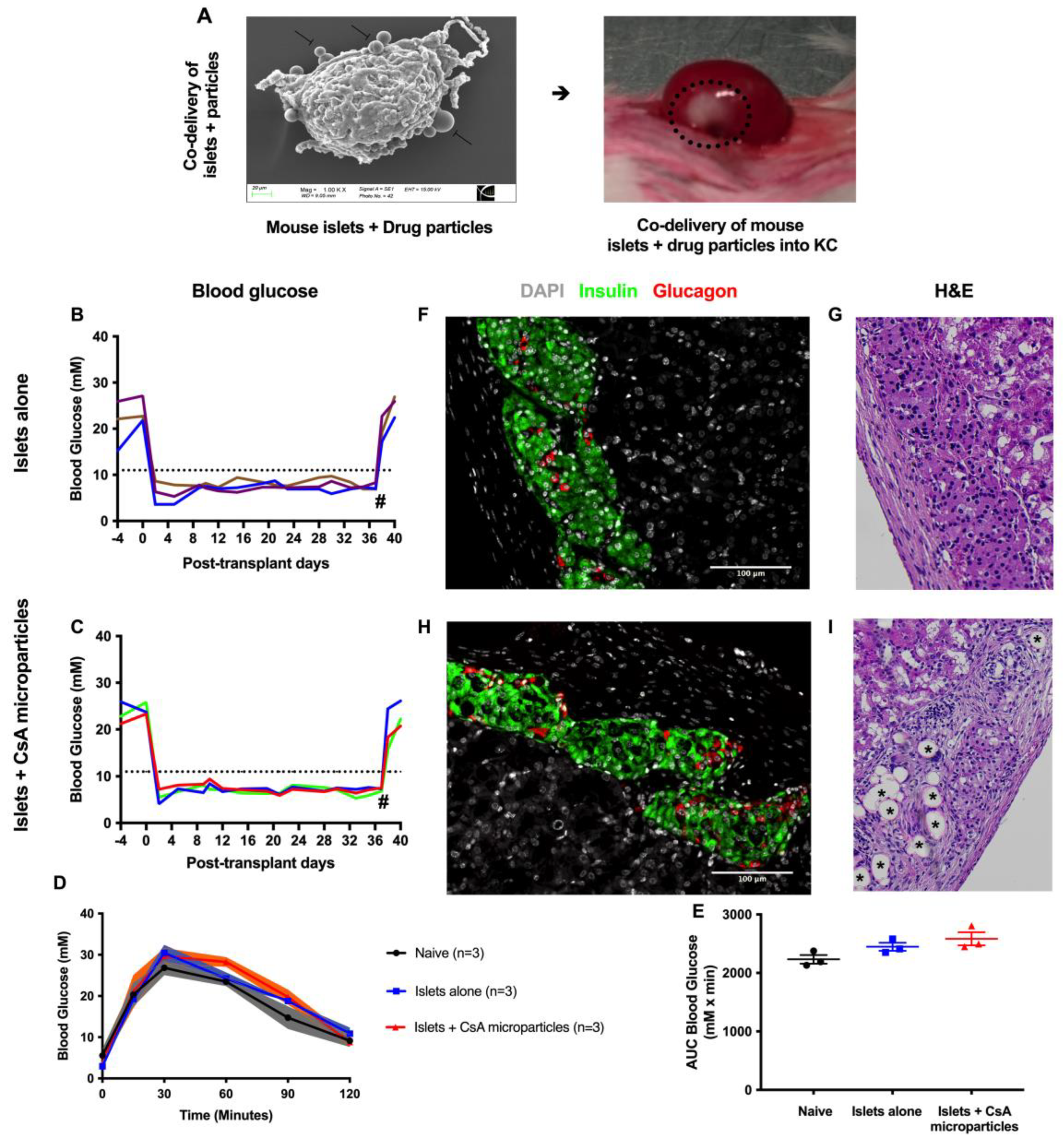
3.2. CsA-Eluting PLGA Microparticles Are Non-Toxic to Murine Syngeneic Islet Grafts
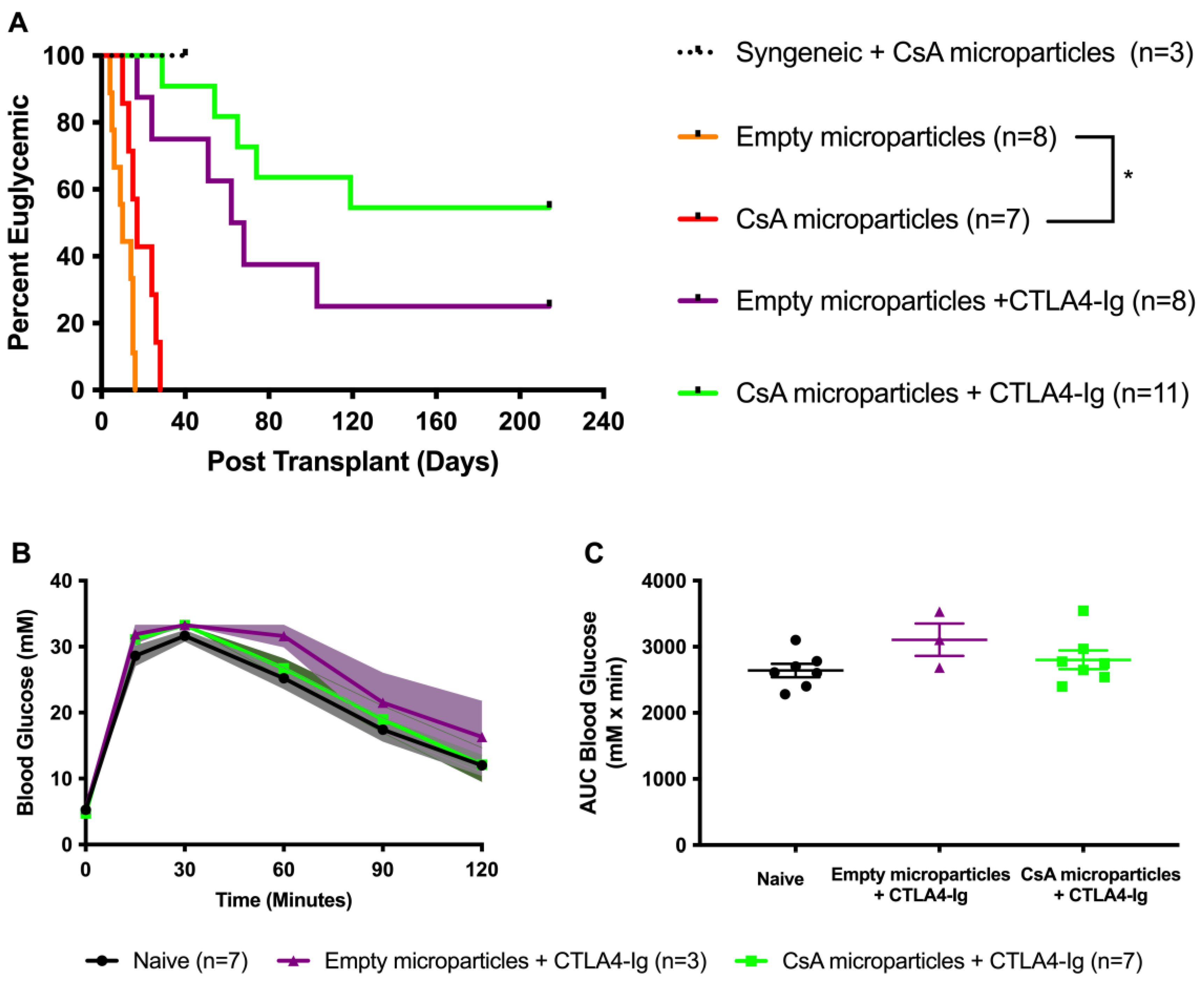
3.3. Co-Localization of CsA-Eluting Microparticles Prolongs Murine Allograft Survival
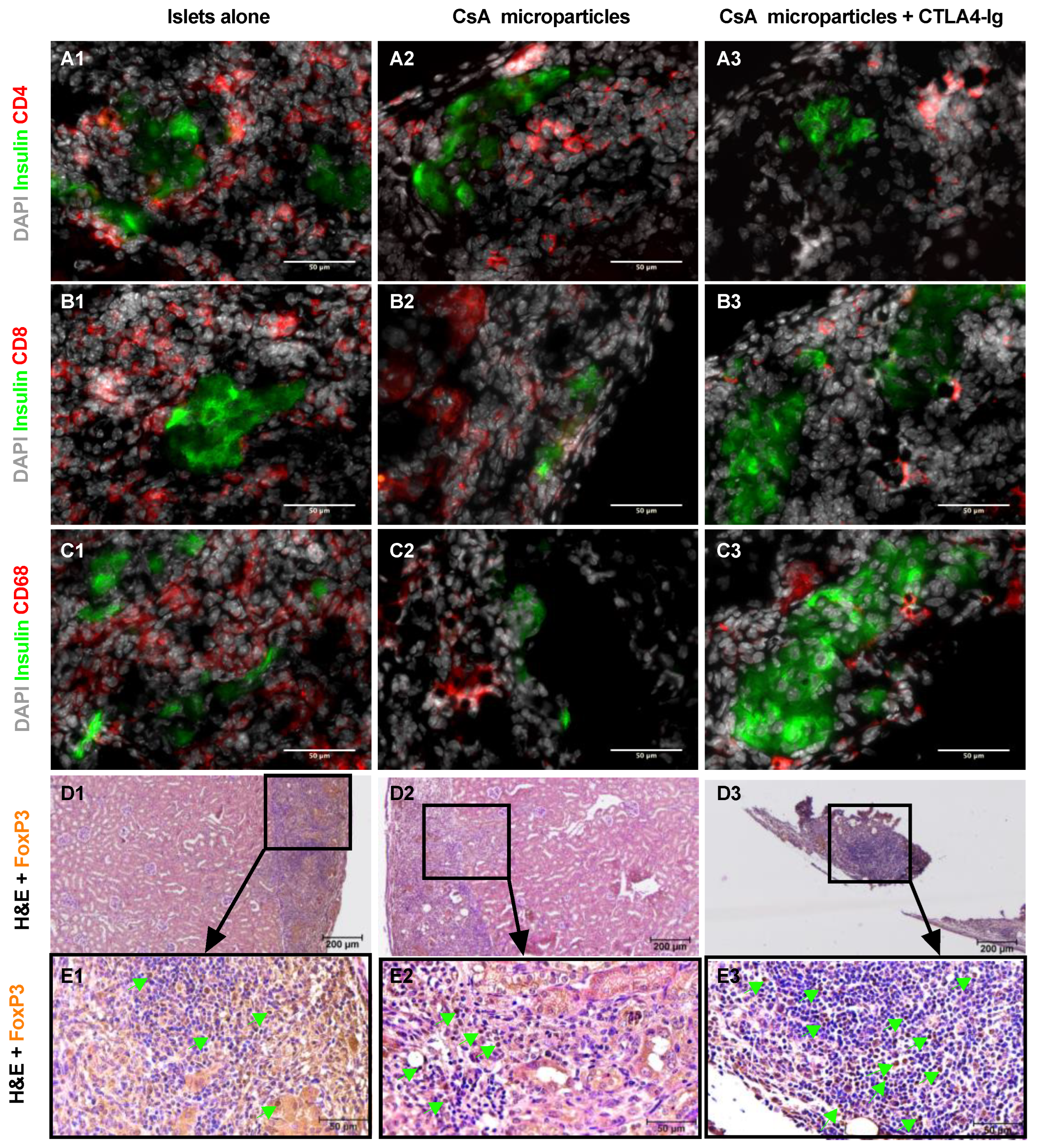
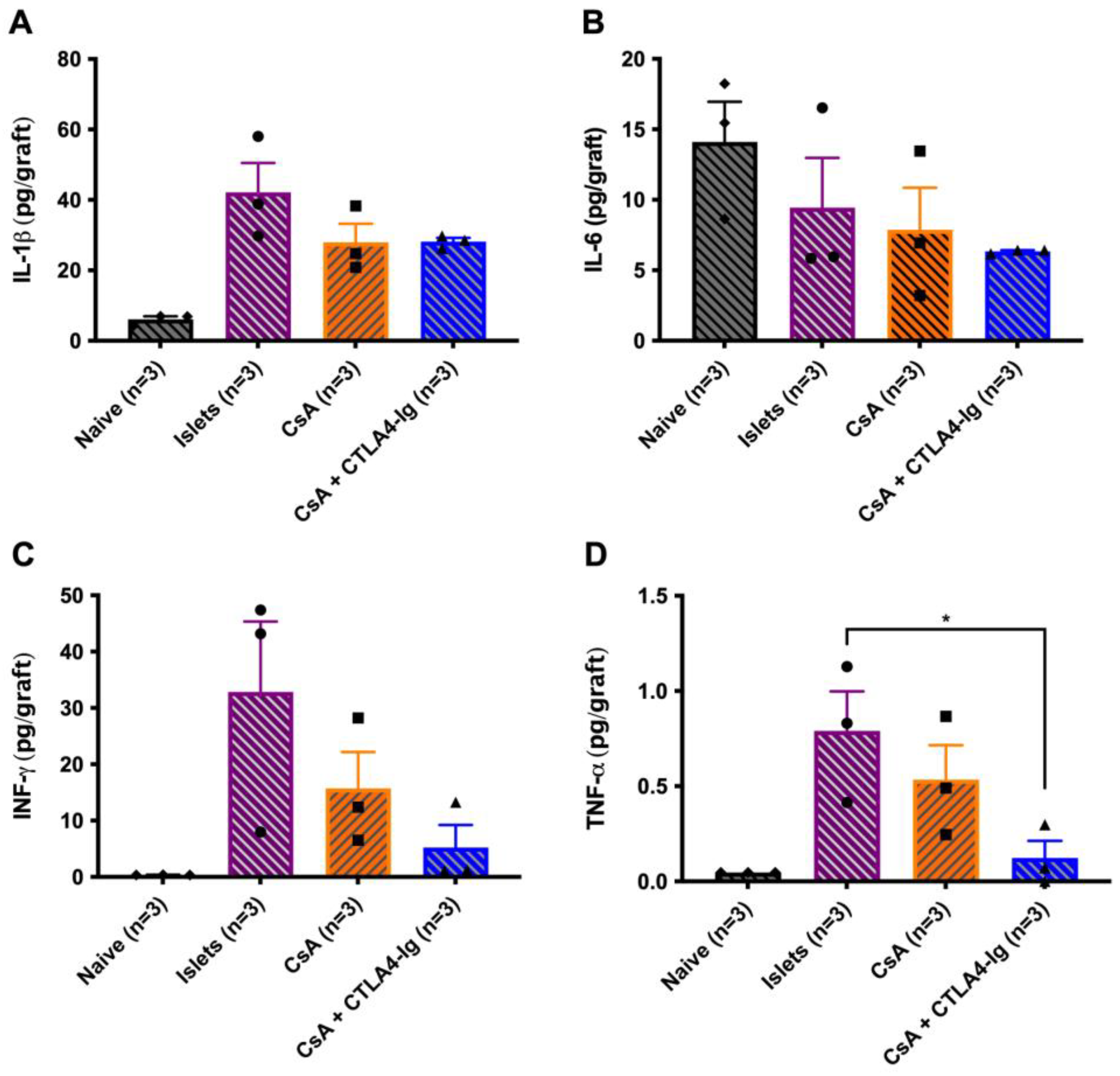
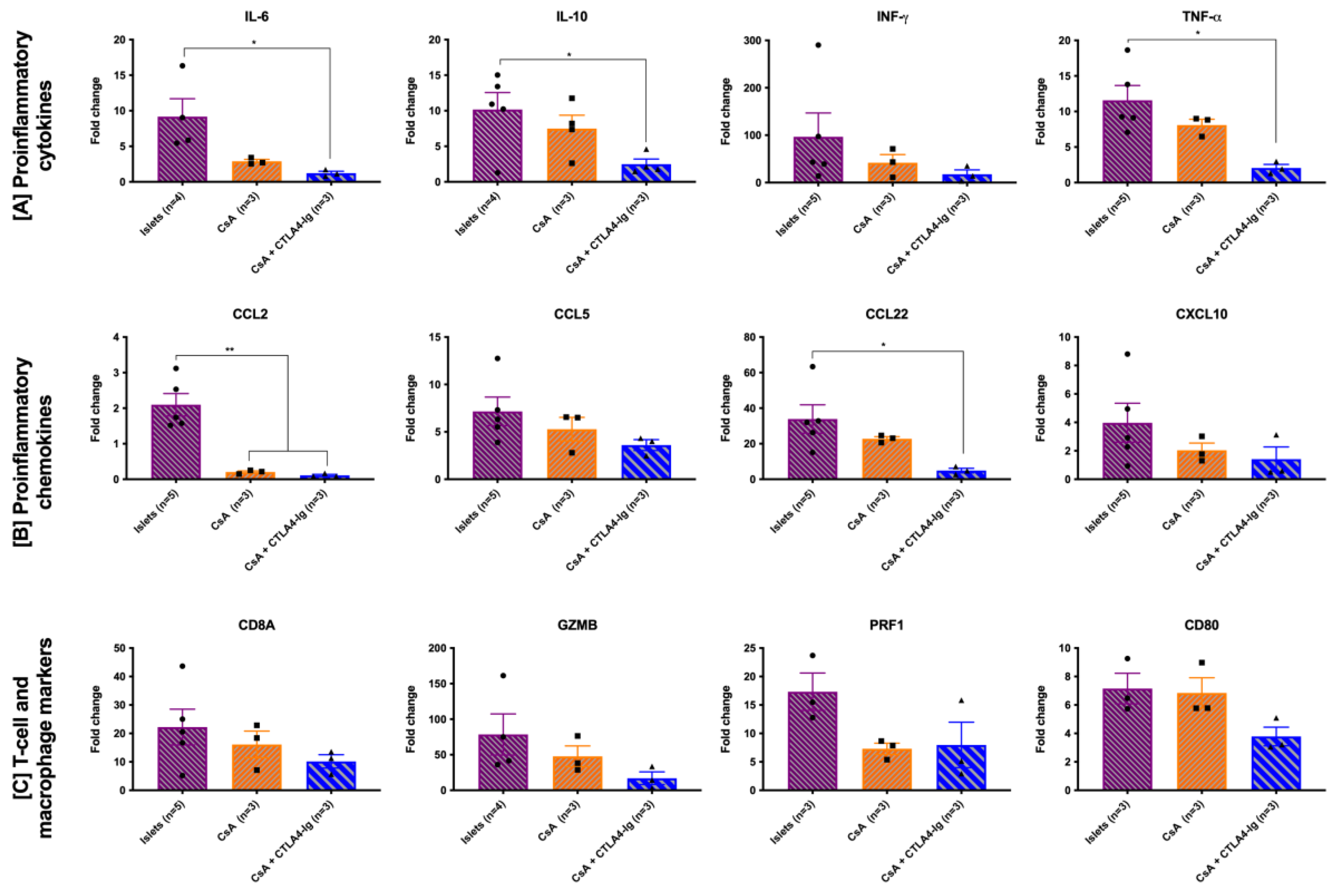
3.4. Co-Localization of CsA Microparticles Modulates Intra-Islet Allograft Proinflammatory Responses and Immune Cell Infiltration
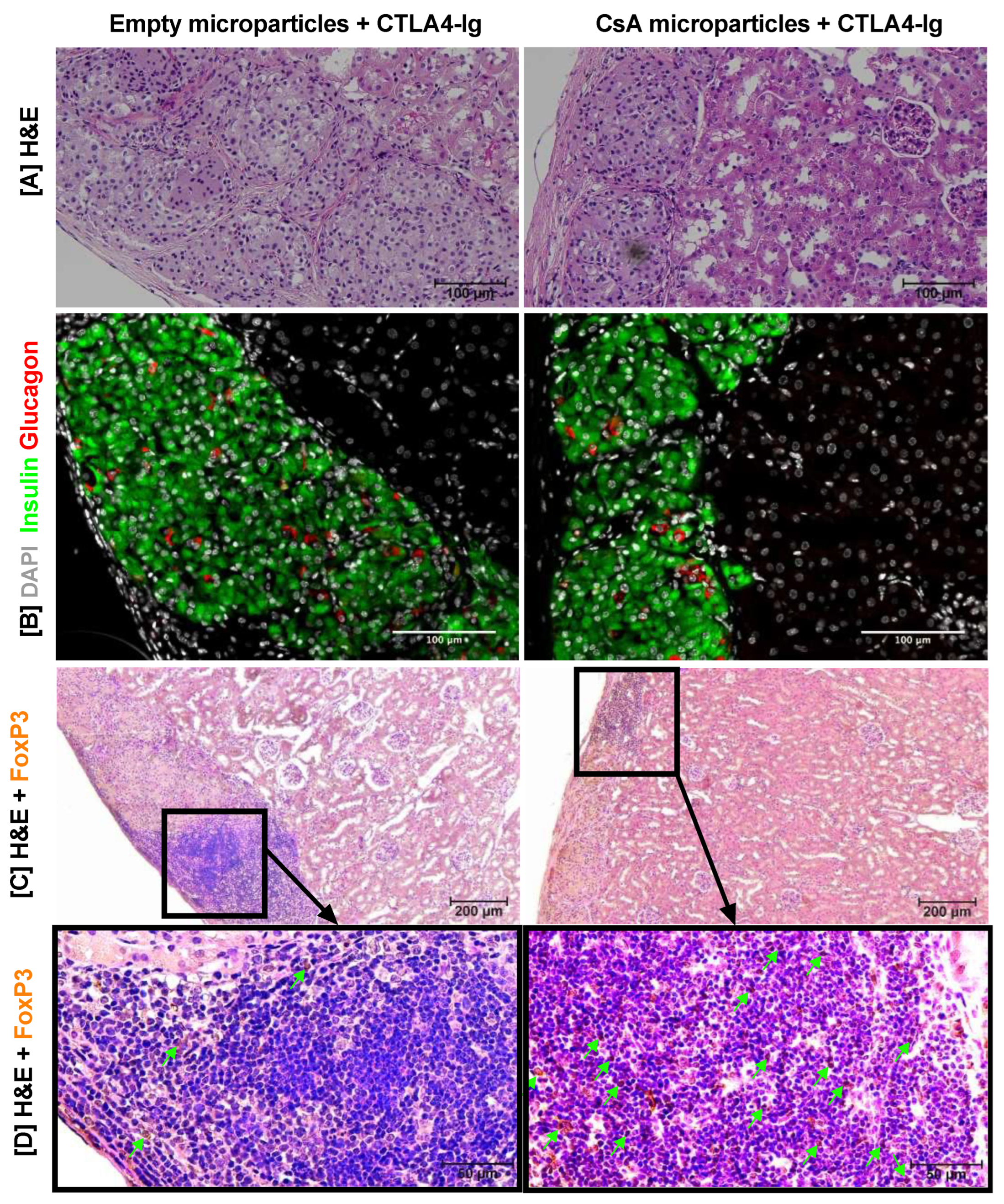
3.5. Histological Characterization of Long-Term Islet Allografts
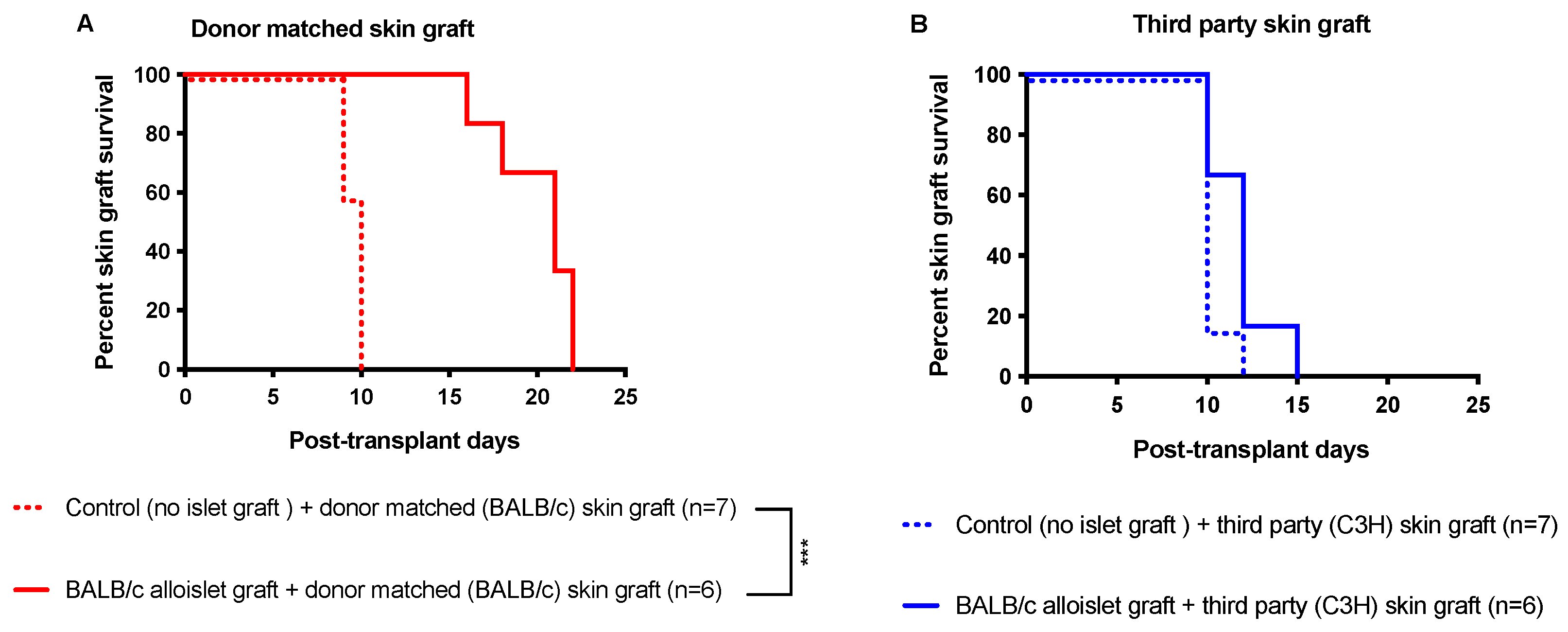
3.6. CsA Microparticles + CTLA4-Ig Treatment Generated Operational Tolerance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rickels, M.R.; Robertson, R.P. Pancreatic Islet Transplantation in Humans: Recent Progress and Future Directions. Endocr. Rev. 2019, 40, 631–668. [Google Scholar] [CrossRef]
- Marfil-Garza, B.A.; Imes, S.; Verhoeff, K.; Hefler, J.; Lam, A.; Dajani, K.; Anderson, B.; O’Gorman, D.; Kin, T.; Bigam, D.; et al. Pancreatic islet transplantation in type 1 diabetes: 20-year experience from a single-centre cohort in Canada. Lancet Diabetes Endocrinol. 2022, 10, 519–532. [Google Scholar] [CrossRef]
- Li, Y.; Frei, A.W.; Labrada, I.M.; Rong, Y.; Liang, J.P.; Samojlik, M.M.; Sun, C.; Barash, S.; Keselowsky, B.G.; Bayer, A.L.; et al. Immunosuppressive PLGA TGF-beta1 Microparticles Induce Polyclonal and Antigen-Specific Regulatory T Cells for Local Immunomodulation of Allogeneic Islet Transplants. Front. Immunol. 2021, 12, 653088. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Phung, C.D.; Kim, J.O.; Yong, C.S.; Kim, J.R.; Yook, S.; Jeong, J.H. The impact of locally-delivered tacrolimus-releasing microspheres and polyethylene glycol-based islet surface modification on xenogeneic islet survival. J. Control. Release 2021, 336, 274–284. [Google Scholar] [CrossRef]
- Chen, M.C.M.; Wang, J.L.; Tzen, J.T.C. Elevating Bioavailability of cyclosporine A via encapsulation in artificial oil bodies stabilized by caleosin. Biotechnol. Prog. 2005, 21, 1297–1301. [Google Scholar] [CrossRef]
- Wee, Y.M.; Choi, M.Y.; Kang, C.H.; Kim, Y.H.; Kim, J.H.; Lee, S.K.; Yu, S.Y.; Kim, S.C.; Han, D.J. The synergistic effect of Tautomycetin on Cyclosporine A-mediated immunosuppression in a rodent islet allograft model. Mol. Med. 2010, 16, 298–306. [Google Scholar] [CrossRef]
- Mishra, J.; Davani, A.J.; Natarajan, G.K.; Kwok, W.M.; Stowe, D.F.; Camara, A.K.S. Cyclosporin A Increases Mitochondrial Buffering of Calcium: An Additional Mechanism in Delaying Mitochondrial Permeability Transition Pore Opening. Cells 2019, 8, 1052. [Google Scholar] [CrossRef]
- Basadonna, G.; Kakizaki, K.; Merrell, R.C. Effect of cyclosporine on established islet autografts. J. Surg. Res. 1986, 40, 450–454. [Google Scholar] [CrossRef]
- Bugliani, M.; Masini, M.; Liechti, R.; Marselli, L.; Xenarios, I.; Boggi, U.; Filipponi, F.; Masiello, P.; Marchetti, P. The direct effects of tacrolimus and cyclosporin A on isolated human islets: A functional, survival and gene expression study. Islets 2009, 1, 106–110. [Google Scholar] [CrossRef]
- Hahn, H.J.; Laube, F.; Lucke, S.; Kloting, I.; Kohnert, K.D.; Warzock, R. Toxic effects of cyclosporine on the endocrine pancreas of Wistar rats. Transplantation 1986, 41, 44–47. [Google Scholar] [CrossRef]
- Gamble, A.; Pepper, A.R.; Bruni, A.; Shapiro, A.M.J. The journey of islet cell transplantation and future development. Islets 2018, 10, 80–94. [Google Scholar] [CrossRef]
- Flores, C.; Fouquet, G.; Moura, I.C.; Maciel, T.T.; Hermine, O. Lessons to Learn From Low-Dose Cyclosporin-A: A New Approach for Unexpected Clinical Applications. Front. Immunol. 2019, 10, 588. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, X.; Nepovimova, E.; Wang, Y.; Yang, H.; Kuca, K. Mechanism of cyclosporine A nephrotoxicity: Oxidative stress, autophagy, and signalings. Food Chem. Toxicol. 2018, 118, 889–907. [Google Scholar] [CrossRef] [PubMed]
- Arita, S.; Kasraie, A.; Une, S.; Ohtsuka, S.; Smith, C.V.; Mullen, Y. Prolongation of islet allograft survival in mice by combined treatment with pravastatin and low-dose cyclosporine. Cell Transplant. 2001, 10, 639–644. [Google Scholar] [CrossRef]
- Lee, D.Y.; Lee, S.; Nam, J.H.; Byun, Y. Minimization of immunosuppressive therapy after islet transplantation: Combined action of heme oxygenase-1 and PEGylation to islet. Am. J. Transplant. 2006, 6, 1820–1828. [Google Scholar] [CrossRef] [PubMed]
- Drachenberg, C.B.; Klassen, D.K.; Weir, M.R.; Wiland, A.; Fink, J.C.; Bartlett, S.T.; Cangro, C.B.; Blahut, S.; Papadimitriou, J.C. Islet cell damage associated with tacrolimus and cyclosporine: Morphological features in pancreas allograft biopsies and clinical correlation. Transplantation 1999, 68, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Goyal, R.; Macri, L.; Kohn, J. Formulation Strategy for the Delivery of Cyclosporine A: Comparison of Two Polymeric Nanospheres. Sci. Rep. 2015, 5, 13065. [Google Scholar] [CrossRef]
- Kuppan, P.; Kelly, S.; Polishevska, K.; Hojanepesov, O.; Seeberger, K.; Korbutt, G.S.; Pepper, A.R. Co-localized immune protection using dexamethasone-eluting micelles in a murine islet allograft model. Am. J. Transplant. 2020, 20, 714–725. [Google Scholar] [CrossRef]
- Azzi, J.; Tang, L.; Moore, R.; Tong, R.; El Haddad, N.; Akiyoshi, T.; Mfarrej, B.; Yang, S.; Jurewicz, M.; Ichimura, T.; et al. Polylactide-cyclosporin A nanoparticles for targeted immunosuppression. FASEB J. 2010, 24, 3927–3938. [Google Scholar] [CrossRef]
- Naeem, M.; Bae, J.; Oshi, M.A.; Kim, M.S.; Moon, H.R.; Lee, B.L.; Im, E.; Jung, Y.; Yoo, J.W. Colon-targeted delivery of cyclosporine A using dual-functional Eudragit((R)) FS30D/PLGA nanoparticles ameliorates murine experimental colitis. Int. J. Nanomed. 2018, 13, 1225–1240. [Google Scholar] [CrossRef]
- Yavuz, B.; Bozdag Pehlivan, S.; Kaffashi, A.; Calamak, S.; Ulubayram, K.; Palaska, E.; Cakmak, H.B.; Unlu, N. In vivo tissue distribution and efficacy studies for cyclosporin A loaded nano-decorated subconjunctival implants. Drug Deliv. 2016, 23, 3279–3284. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Kim, H.J.; Byun, N.R.; Park, C.G. Donor-Specific Regulatory T Cell-Mediated Immune Tolerance in an Intrahepatic Murine Allogeneic Islet Transplantation Model with Short-Term Anti-CD154 mAb Single Treatment. Cell Transplant. 2020, 29, 963689720913876. [Google Scholar] [CrossRef]
- Woodward, K.B.; Zhao, H.; Shrestha, P.; Batra, L.; Tan, M.; Grimany-Nuno, O.; Bandura-Morgan, L.; Askenasy, N.; Shirwan, H.; Yolcu, E.S. Pancreatic islets engineered with a FasL protein induce systemic tolerance at the induction phase that evolves into long-term graft-localized immune privilege. Am. J. Transplant. 2020, 20, 1285–1295. [Google Scholar] [CrossRef] [PubMed]
- Bruni, A.; Gala-Lopez, B.; Pepper, A.R.; Abualhassan, N.S.; Shapiro, A.J. Islet cell transplantation for the treatment of type 1 diabetes: Recent advances and future challenges. Diabetes Metab. Syndr. Obes. 2014, 7, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.; Appari, M.; Forbes, S. Considerations and challenges of islet transplantation and future therapies on the horizon. Am. J. Physiol. Endocrinol. Metab. 2022, 322, E109–E117. [Google Scholar] [CrossRef] [PubMed]
- Bauer, W.; Gyenesei, A.; Kretowski, A. The Multifactorial Progression from the Islet Autoimmunity to Type 1 Diabetes in Children. Int. J. Mol. Sci. 2021, 22, 7493. [Google Scholar] [CrossRef]
- Takaki, T.; Shimoda, M. Pancreatic islet transplantation: Toward definitive treatment for diabetes mellitus. Glob Health Med. 2020, 2, 200–211. [Google Scholar] [CrossRef]
- Kanak, M.A.; Takita, M.; Kunnathodi, F.; Lawrence, M.C.; Levy, M.F.; Naziruddin, B. Inflammatory response in islet transplantation. Int. J. Endocrinol. 2014, 2014, 451035. [Google Scholar] [CrossRef]
- Wang, X.; Brown, N.K.; Wang, B.; Shariati, K.; Wang, K.; Fuchs, S.; Melero-Martin, J.M.; Ma, M. Local Immunomodulatory Strategies to Prevent Allo-Rejection in Transplantation of Insulin-Producing Cells. Adv. Sci. 2021, 8, e2003708. [Google Scholar] [CrossRef]
- Barra, J.M.; Kozlovskaya, V.; Kharlampieva, E.; Tse, H.M. Localized Immunosuppression With Tannic Acid Encapsulation Delays Islet Allograft and Autoimmune-Mediated Rejection. Diabetes 2020, 69, 1948–1960. [Google Scholar] [CrossRef]
- Kuppan, P.; Kelly, S.; Polishevska, K.; Seeberger, K.; Rosko, M.; Korbutt, G.S.; Pepper, A.R. 209.1: Co-localized Immune Protection Using Cyclosporine A Eluting Micelles in a Murine Islet Allograft Model. Transplantation 2021, 105, S13. [Google Scholar] [CrossRef]
- Razavi, M.; Wang, J.; Thakor, A.S. Localized drug delivery graphene bioscaffolds for cotransplantation of islets and mesenchymal stem cells. Sci. Adv. 2021, 7, eabf9221. [Google Scholar] [CrossRef] [PubMed]
- Frei, A.W.; Li, Y.; Jiang, K.; Buchwald, P.; Stabler, C.L. Local delivery of fingolimod from three-dimensional scaffolds impacts islet graft efficacy and microenvironment in a murine diabetic model. J. Tissue Eng. Regen. Med. 2018, 12, 393–404. [Google Scholar] [CrossRef]
- Ozbay, L.A.; Smidt, K.; Mortensen, D.M.; Carstens, J.; Jorgensen, K.A.; Rungby, J. Cyclosporin and tacrolimus impair insulin secretion and transcriptional regulation in INS-1E beta-cells. Br. J. Pharmacol. 2011, 162, 136–146. [Google Scholar] [CrossRef]
- Tang, L.; Azzi, J.; Kwon, M.; Mounayar, M.; Tong, R.; Yin, Q.; Moore, R.; Skartsis, N.; Fan, T.M.; Abdi, R.; et al. Immunosuppressive Activity of Size-Controlled PEG-PLGA Nanoparticles Containing Encapsulated Cyclosporine A. J. Transplant. 2012, 2012, 896141. [Google Scholar] [CrossRef]
- Lagreca, E.; Onesto, V.; Di Natale, C.; La Manna, S.; Netti, P.A.; Vecchione, R. Recent advances in the formulation of PLGA microparticles for controlled drug delivery. Prog. Biomater. 2020, 9, 153–174. [Google Scholar] [CrossRef] [PubMed]
- Ramazani, F.; Chen, W.; van Nostrum, C.F.; Storm, G.; Kiessling, F.; Lammers, T.; Hennink, W.E.; Kok, R.J. Strategies for encapsulation of small hydrophilic and amphiphilic drugs in PLGA microspheres: State-of-the-art and challenges. Int. J. Pharm. 2016, 499, 358–367. [Google Scholar] [CrossRef]
- Liu, P.C.; Ssu, C.T.; Tsao, Y.P.; Liou, T.L.; Tsai, C.Y.; Chou, C.T.; Chen, M.H.; Leu, C.M. Cytotoxic T lymphocyte-associated antigen-4-Ig (CTLA-4-Ig) suppresses Staphylococcus aureus-induced CD80, CD86, and pro-inflammatory cytokine expression in human B cells. Arthritis Res. Ther. 2020, 22, 64. [Google Scholar] [CrossRef]
- Pawlick, R.; Gala-Lopez, B.; Pepper, A.R.; McCall, M.; Ziff, O.; Shapiro, A.M. The combination of anti-NKG2D and CTLA-4 Ig therapy prolongs islet allograft survival in amurine model. Am. J. Transplant. 2014, 14, 2367–2374. [Google Scholar] [CrossRef]
- Steurer, W.; Nickerson, P.W.; Steele, A.W.; Steiger, J.; Zheng, X.X.; Strom, T.B. Ex vivo coating of islet cell allografts with murine CTLA4/Fc promotes graft tolerance. J. Immunol. 1995, 155, 1165–1174. [Google Scholar] [CrossRef]
- Levisetti, M.G.; Padrid, P.P.; Szot, G.L.; Mittal, N.; Meehan, S.M.; Wardrip, C.L.; Gray, G.S.; Bruce, D.S.; Thistlethwaite, J.R., Jr.; Bluestone, J.A. Immunosuppressive effects of human CTLA4Ig in a non-human primate model of allogeneic pancreatic islets transplantation. J. Immunol. 1997, 159, 5187–5191. [Google Scholar] [CrossRef] [PubMed]
- Badell, I.R.; Russell, M.C.; Cardona, K.; Shaffer, V.O.; Turner, A.P.; Avila, J.G.; Cano, J.A.; Leopardi, F.V.; Song, M.; Strobert, E.A.; et al. CTLA4Ig prevents alloantibody formation following nonhuman primate islet transplantation using the CD40-specific antibody 3A8. Am. J. Transplant. 2012, 12, 1918–1923. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.Y.; Pollard, C.A.; Kumar, R.; Drogemuller, C.J.; Naziruddin, B.; Stover, C.; Issa, E.; Isherwood, J.; Cooke, J.; Levy, M.F.; et al. A comparison of the inflammatory response following autologous compared with allogenic islet cell transplantation. Ann. Transl. Med. 2021, 9, 98. [Google Scholar] [CrossRef] [PubMed]
- Krummel, M.F.; Bartumeus, F.; Gerard, A. T cell migration, search strategies and mechanisms. Nat. Rev. Immunol. 2016, 16, 193–201. [Google Scholar] [CrossRef]
- Eizirik, D.L.; Mandrup-Poulsen, T. A choice of death--the signal-transduction of immune-mediated beta-cell apoptosis. Diabetologia 2001, 44, 2115–2133. [Google Scholar] [CrossRef]
- Grey, S.T.; Longo, C.; Shukri, T.; Patel, V.I.; Csizmadia, E.; Daniel, S.; Arvelo, M.B.; Tchipashvili, V.; Ferran, C. Genetic engineering of a suboptimal islet graft with A20 preserves beta cell mass and function. J. Immunol. 2003, 170, 6250–6256. [Google Scholar] [CrossRef]
- Yoshimatsu, G.; Kunnathodi, F.; Saravanan, P.B.; Shahbazov, R.; Chang, C.; Darden, C.M.; Zurawski, S.; Boyuk, G.; Kanak, M.A.; Levy, M.F.; et al. Pancreatic beta-Cell-Derived IP-10/CXCL10 Isletokine Mediates Early Loss of Graft Function in Islet Cell Transplantation. Diabetes 2017, 66, 2857–2867. [Google Scholar] [CrossRef]
- Zammit, N.W.; Walters, S.N.; Seeberger, K.L.; O’Connell, P.J.; Korbutt, G.S.; Grey, S.T. A20 as an immune tolerance factor can determine islet transplant outcomes. JCI Insight 2019, 4, e131028. [Google Scholar] [CrossRef]
- Krzystyniak, A.; Golab, K.; Witkowski, P.; Trzonkowski, P. Islet cell transplant and the incorporation of Tregs. Curr. Opin. Organ. Transplant. 2014, 19, 610–615. [Google Scholar] [CrossRef]
- Marfil-Garza, B.A.; Hefler, J.; Bermudez De Leon, M.; Pawlick, R.; Dadheech, N.; Shapiro, A.M.J. Progress in Translational Regulatory T Cell Therapies for Type 1 Diabetes and Islet Transplantation. Endocr. Rev. 2021, 42, 198–218. [Google Scholar] [CrossRef]
- Peng, Y.; Laouar, Y.; Li, M.O.; Green, E.A.; Flavell, R.A. TGF-beta regulates in vivo expansion of Foxp3-expressing CD4+CD25+ regulatory T cells responsible for protection against diabetes. Proc. Natl. Acad. Sci. USA 2004, 101, 4572–4577. [Google Scholar] [CrossRef] [PubMed]
- Barahona de Brito, C.; Patra, A.K. NFAT Factors Are Dispensable for the Development but Are Critical for the Maintenance of Foxp3(+) Regulatory T Cells. Cells 2022, 11, 1397. [Google Scholar] [CrossRef] [PubMed]
- Magg, T.; Mannert, J.; Ellwart, J.W.; Schmid, I.; Albert, M.H. Subcellular localization of FOXP3 in human regulatory and nonregulatory T cells. Eur. J. Immunol. 2012, 42, 1627–1638. [Google Scholar] [CrossRef] [PubMed]
- Shahaf, G.; Moser, H.; Ozeri, E.; Mizrahi, M.; Abecassis, A.; Lewis, E.C. alpha-1-antitrypsin gene delivery reduces inflammation, increases T-regulatory cell population size and prevents islet allograft rejection. Mol. Med. 2011, 17, 1000–1011. [Google Scholar] [CrossRef]
- Marfil-Garza, B.A.; Pawlick, R.L.; Szeto, J.; Kroger, C.; Tahiliani, V.; Hefler, J.; Dadheech, N.; Seavey, M.M.; Wolf, J.; Jasuja, R.R.; et al. Tumor necrosis factor receptor superfamily member 25 (TNFRSF25) agonists in islet transplantation: Endogenous in vivo regulatory T cell expansion promotes prolonged allograft survival. Am. J. Transplant. 2022, 22, 1101–1114. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuppan, P.; Wong, J.; Kelly, S.; Lin, J.; Worton, J.; Castro, C.; Paramor, J.; Seeberger, K.; Cuesta-Gomez, N.; Anderson, C.C.; et al. Long-Term Survival and Induction of Operational Tolerance to Murine Islet Allografts by Co-Transplanting Cyclosporine A Microparticles and CTLA4-Ig. Pharmaceutics 2023, 15, 2201. https://doi.org/10.3390/pharmaceutics15092201
Kuppan P, Wong J, Kelly S, Lin J, Worton J, Castro C, Paramor J, Seeberger K, Cuesta-Gomez N, Anderson CC, et al. Long-Term Survival and Induction of Operational Tolerance to Murine Islet Allografts by Co-Transplanting Cyclosporine A Microparticles and CTLA4-Ig. Pharmaceutics. 2023; 15(9):2201. https://doi.org/10.3390/pharmaceutics15092201
Chicago/Turabian StyleKuppan, Purushothaman, Jordan Wong, Sandra Kelly, Jiaxin Lin, Jessica Worton, Chelsea Castro, Joy Paramor, Karen Seeberger, Nerea Cuesta-Gomez, Colin C. Anderson, and et al. 2023. "Long-Term Survival and Induction of Operational Tolerance to Murine Islet Allografts by Co-Transplanting Cyclosporine A Microparticles and CTLA4-Ig" Pharmaceutics 15, no. 9: 2201. https://doi.org/10.3390/pharmaceutics15092201