
Light-Based Anti-Biofilm and Antibacterial Strategies

Abstract
:1. Introduction
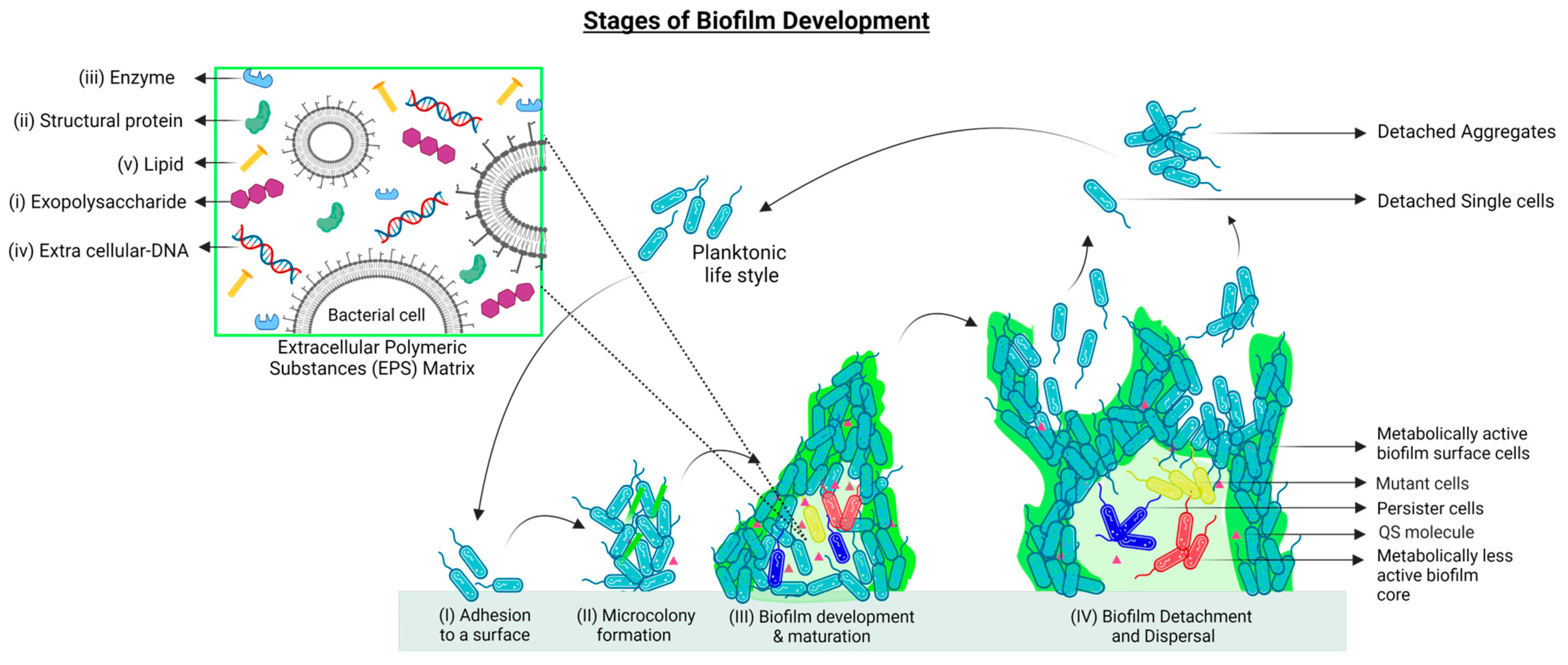
2. Biofilm Formation and Development

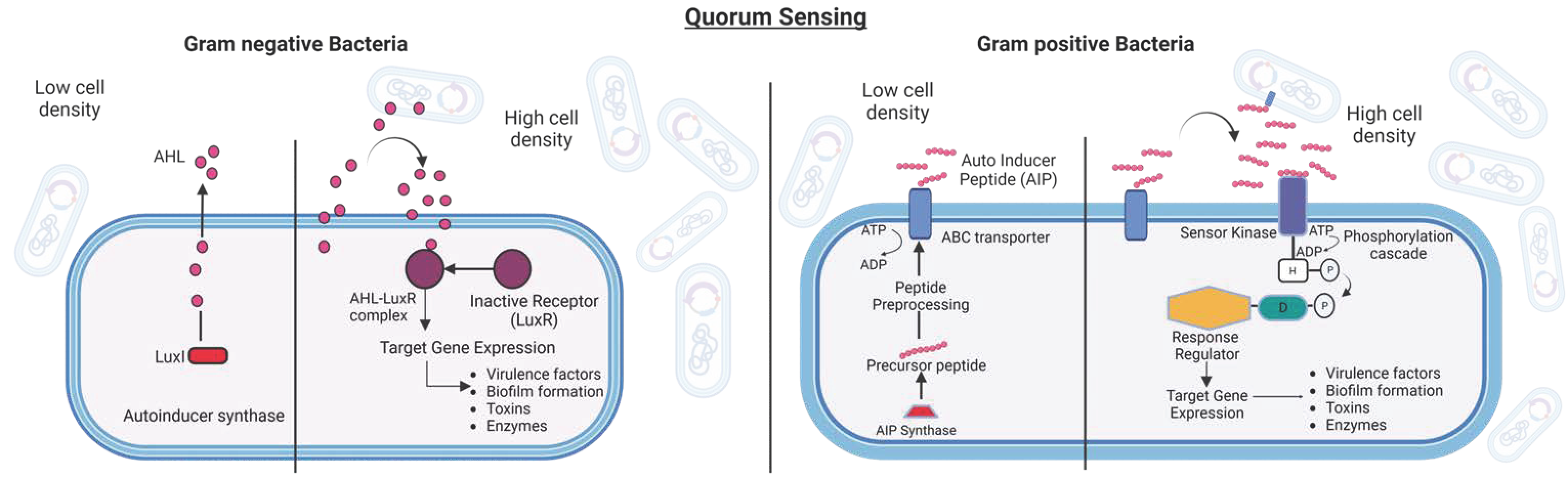
Quorum Sensing

3. Mechanism through Which Biofilms Combat Antibiotics
3.1. Metabolic Activity Heterogeneity and Tolerance Acquisition
3.2. Adaptive Stress Responses
3.3. Antibiotic Resistance
3.4. Horizontal Gene Transfer (HGT)
3.5. Efflux Pumps in Biofilm Resistance
4. Biofilm Disruption Strategies
4.1. Anti-Adhesion Strategies
4.2. Quorum Quenching or Quorum Sensing Inhibition
4.2.1. Targeting QS Signaling Molecules
4.2.2. Targeting Signaling Molecule Receptors
4.2.3. Blocking the Signaling Cascade
4.2.4. Targeting the EPS Chemical Composition and Structure
4.3. Targeting Persister Cells
4.4. Targeting Efflux Pumps
5. Light-Based Antibiofilm Strategies
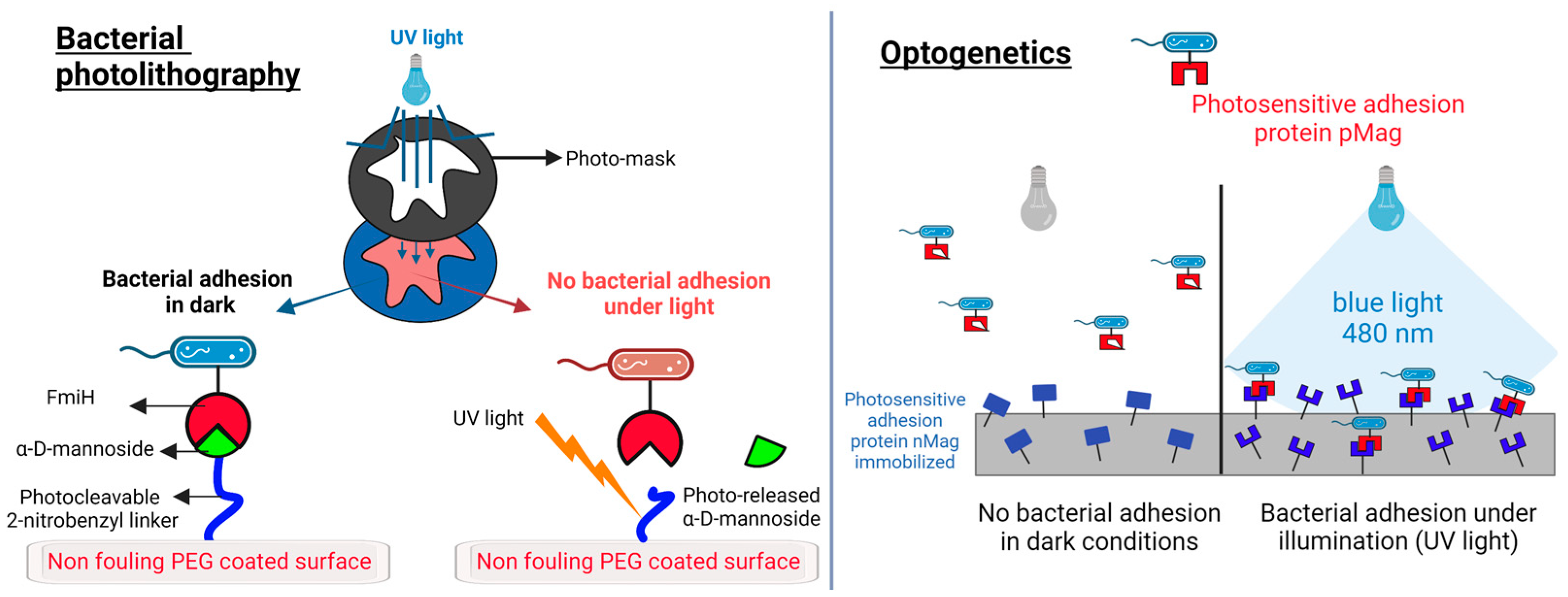
5.1. Bacterial Adhesion: Light-Triggered Control of Bacterial Adhesion

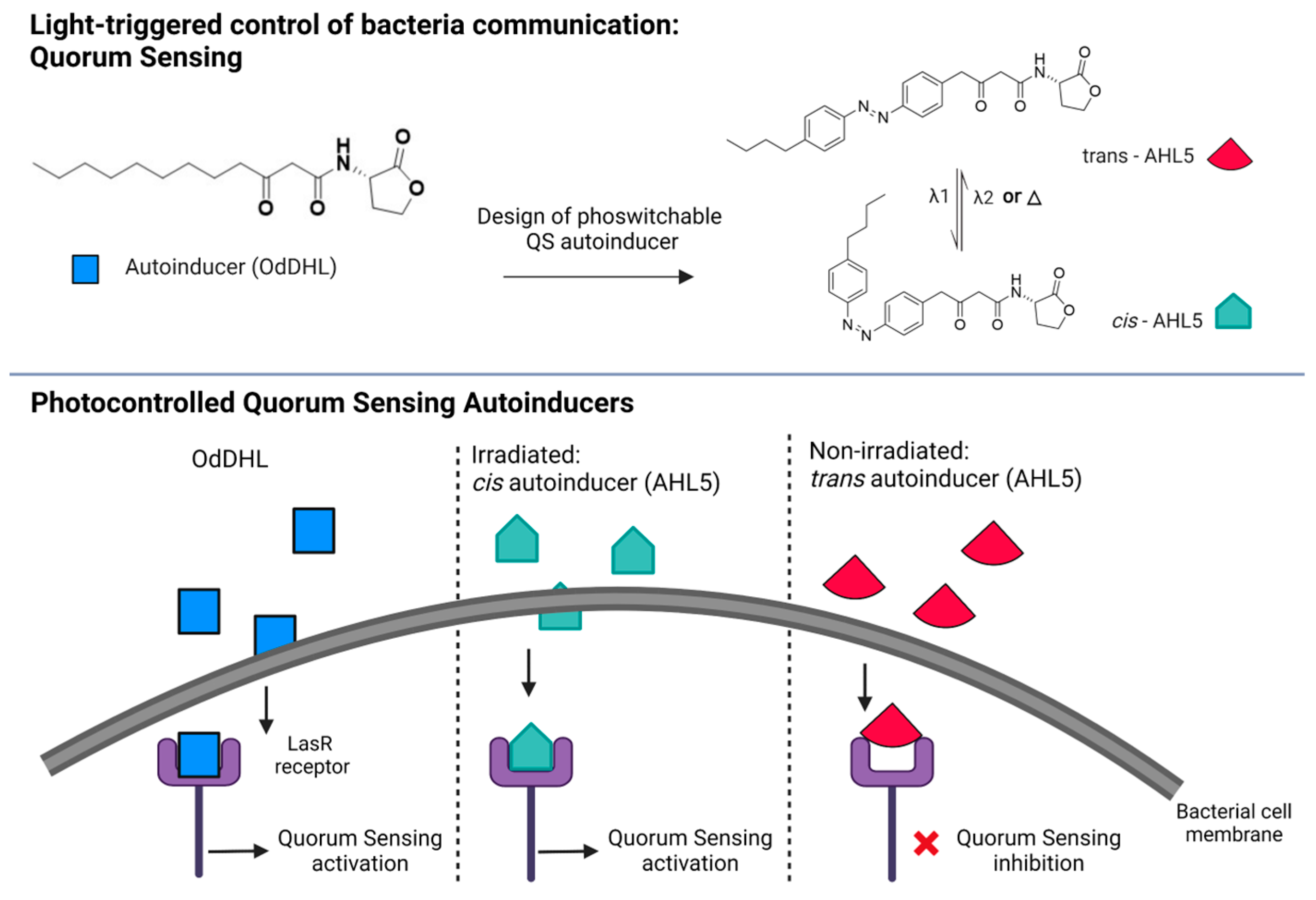
5.2. Bacterial Communication: Photoswitchable Modulators of Quorum Sensing
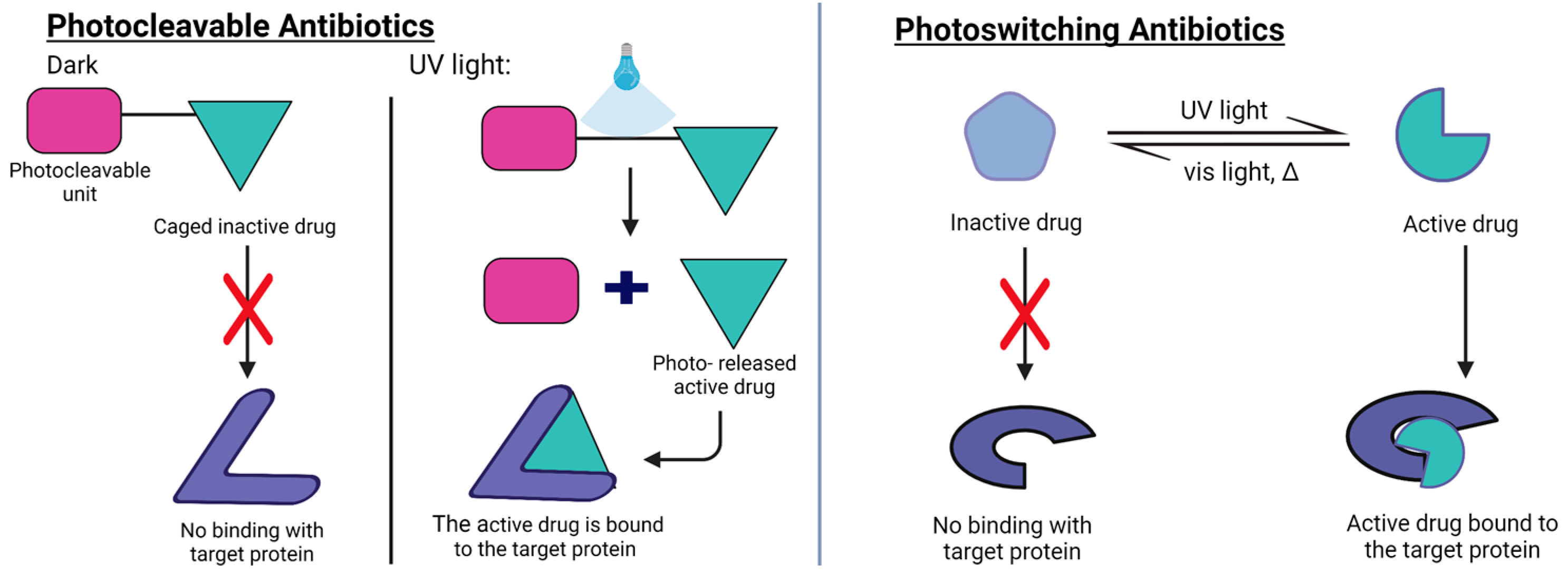
5.3. Biofilm Maturation and Planktonic Phase: Photocleavable and Photoswitchable Antibiotics
6. Light-Based Materials Strategies to Tackle Bacterial Infections
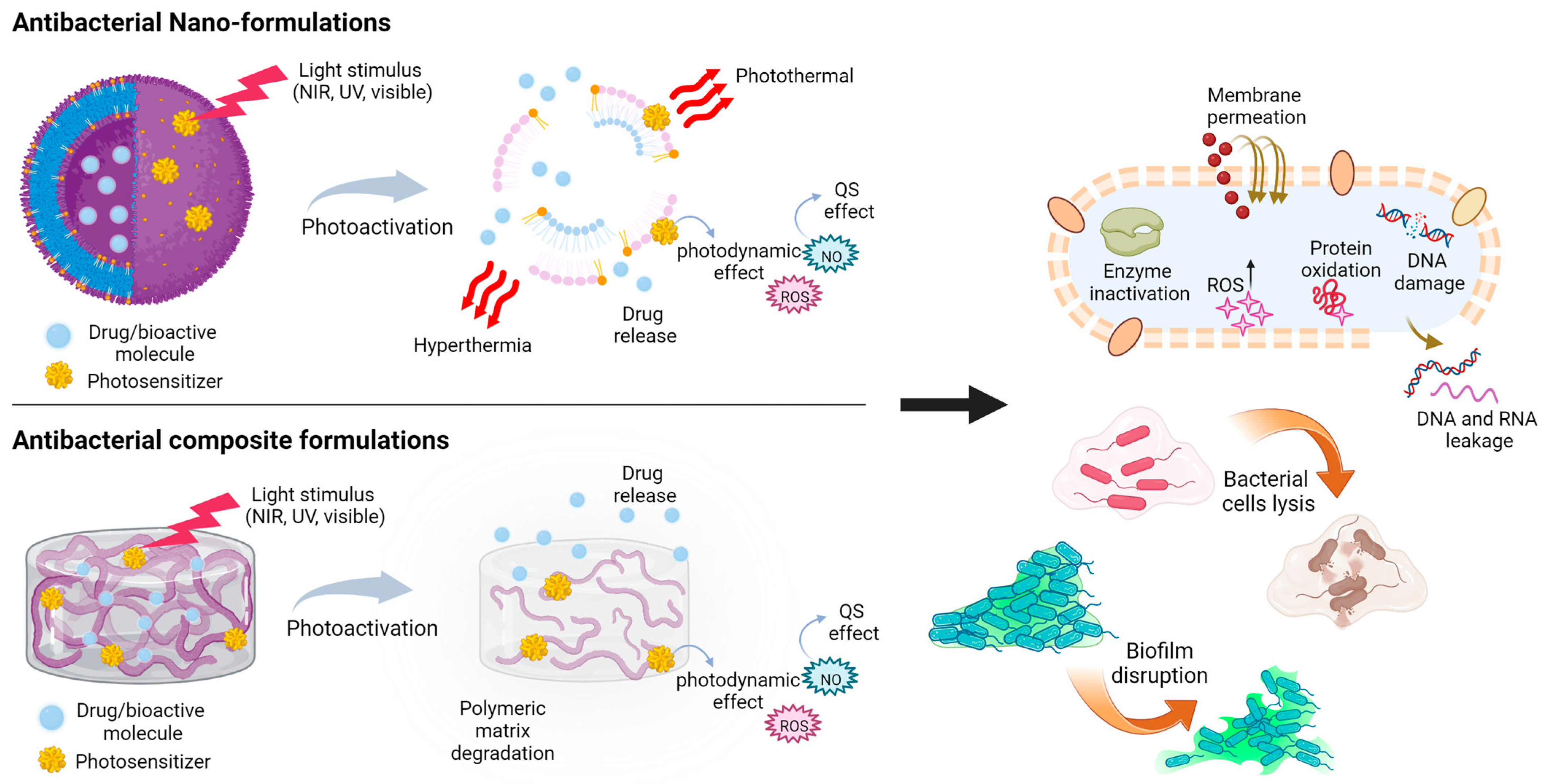
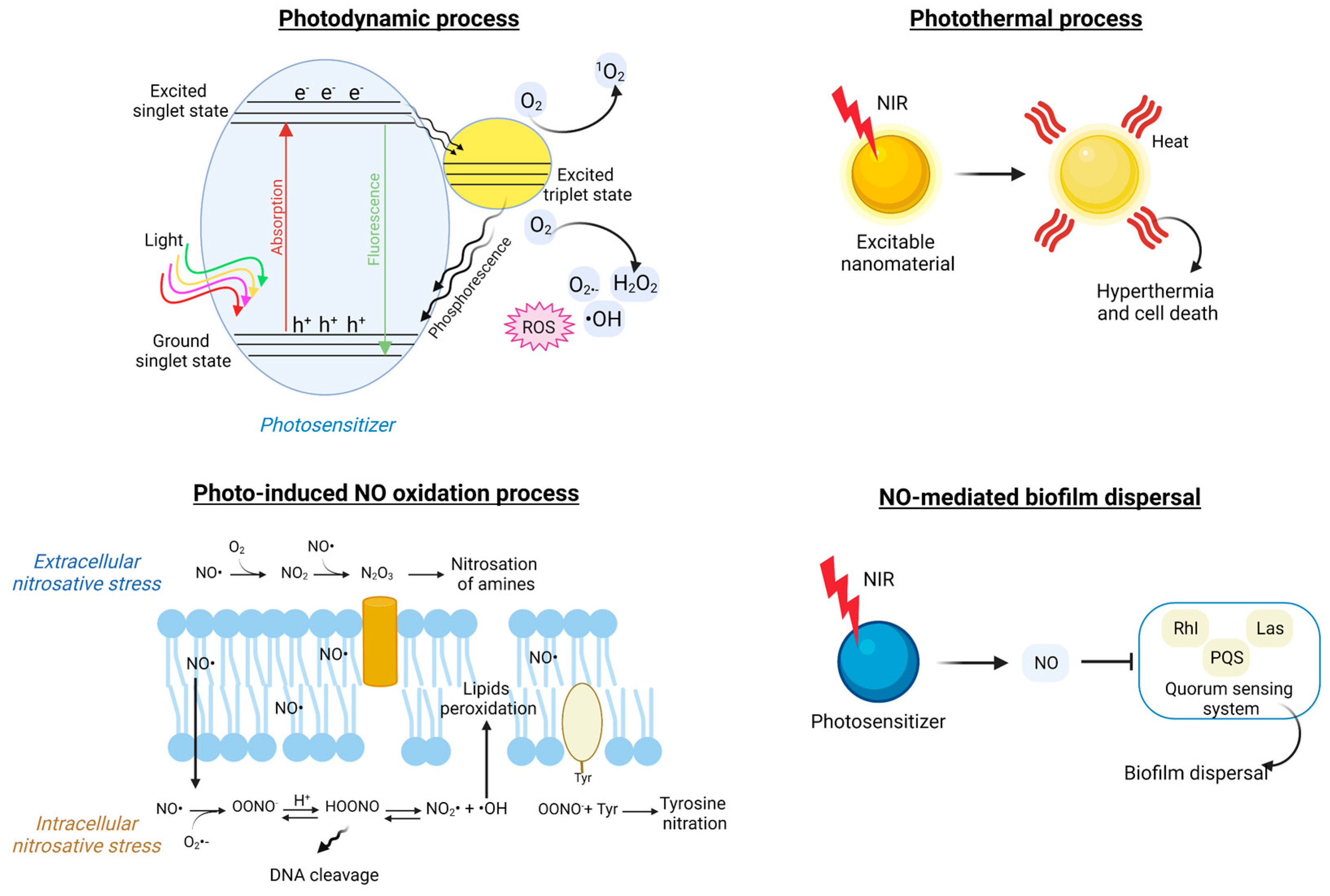
6.1. Light-Triggered Nano-Formulations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Materials | Formulation | Light | Mechanism | Target Microorganism | Application |
|---|---|---|---|---|---|
| PVA-Prussian blue nanoparticle hydrogel films | Nanoparticles in hydrogels | NIR | Localized photothermal therapy | Pseudomonas aeruginosa [258] | |
| Sodium alginate hydrogel loaded with Cu2O and Bi12O17Cl2 NPs | Nanoparticles in hydrogels | NIR | Hydrogel crosslinking, film formation, and ROS generation | Staphylococcus aureus, Escherichia coli, and Streptococcus mutans [259] | tooth whitening and biofilm removal |
| Upconversion nanoparticles (UCNPs) and porphyrinic MOFs (PCN-224) NPs doped with L-arginine and incorporated in PVDF electrospun fibers | Nanoparticles in nanofibers | NIR | ROS generation and nitric oxide-assisted photodynamic therapy | Staphylococcus aureus and Pseudomonas aeruginosa [260] | wound healing |
| Upconversion nanoparticles (UCNPs) incorporated in PVDF electrospun fibers | Nanoparticles in nanofibers | NIR | ROS generation | Staphylococcus aureus and Escherichia coli [261] | wound healing |
| PVA microneedles with a metal-organic framework and multifunctional porphyrin-like metal center NPs | Microneedles | NIR | Photothermal conversion and nanozyme/peroxidase properties of NPs | Staphylococcus aureus [260] | wound healing |
| Iodophilic MOF UiO-66 containing Au nanorods coated with SiO2 and embedded in PVP | Nanoparticles in films | NIR | Photoactive nanoparticles | Staphylococcus aureus and Escherichia coli [262] | nosocomial infections |
| Ag-sodium lignin sulfonate NPs and polypyrrole-polydopamine NPs in poly(ethylene glycol) diacrylate hydrogel | Nanoparticles in hydrogels | NIR | Photothermal activity and antibacterial Ag ion release | Staphylococcus aureus and Escherichia coli [263] | wound dressings |
| PLGA-PCL-methylene blue fibers | Nanofibers | Visible | Controlled matrix degradation and photosensitizer release, photodynamic therapy, and ROS generation | Escherichia coli and Streptococcus mutans [264] | |
| Conjugated polymer NPs + cell-penetrating peptides embedded in polyisocyanides hydrogel | Nanoparticles in hydrogels | White and NIR | Synergistic photodynamic and photothermal therapy | Staphylococcus aureus, Escherichia coli, and Aspergillus niger [248] | clinical infections |
| Light-responsive TiO2 nanotubes and thermo-responsive copolymer | Functionalized composite surface | UV | ROS generation | Staphylococcus aureus and Escherichia coli [265] | anti-adhesion |
| Porphyrin photosensitizer and PLGA-encapsulated bFGF nanospheres embedded in carboxymethyl chitosan-sodium alginate | Nanoparticles in hydrogels | Visible | Photodynamic chemotherapy | Staphylococcus aureus and MDR-Staphylococcus aureus [266] | burn wounds |
| ZnO incorporated with Ag NPs, embedded in carboxymethyl cellulose hydrogel | Nanoparticles in hydrogels | Visible | Ag and Zn ions are released and ROS generation occurs | Staphylococcus aureus and Escherichia coli [267] | |
| Porphyrin-based porous organic polymers | Nanoparticles in films | Visible | Photothermal effect and ROS generation | Methicillin-resistant Staphylococcus aureus [268] | wound healing |
| Riboflavin-modified PVC film | Functionalized composite surface | Blue light | ROS generation | Pseudomonas aeruginosa [269] | |
| Hydrogel of polyvinyl alcohol modified with chitosan, polydopamine, and NO release donor/red phosphorous nanofilm | Functionalized composite surface | NIR | Peroxynitrite (ONOO−) generation, controlled release, and hyperthermia | MDR-Staphylococcus aureus [270] | bone implants |
| PPy-poly dopamine NPs embedded in NIPAm/acrylic acid hydrogel | Nanoparticles in hydrogels | NIR | Light-triggered tunable hydrogel deformation and adhesion and photothermal therapy | Staphylococcus aureus and Escherichia coli [271] | wound healing |
| Ciprofloxacin-loaded PEG hydrogel | Hydrogel | UV | Light-triggered drug release and photo-cleavable molecular cage | Staphylococcus aureus [272] | wound healing |
| Dibenzaldehyde-grafted poly (ethylene glycol), lauric acid-terminated chitosan, and curcumin-loaded mesoporous polydopamine NPs | Nanoparticles in hydrogels | NIR | Light-triggered drug release, hyperthermia with cellular component leakage, and disruption of the bacterial membrane | Staphylococcus aureus and Escherichia coli [273] | wound healing |
| Polysaccharide hydrogel encapsulating ferric tannate NPs and vancomycin | Nanoparticles in hydrogels | NIR | Hyperthermia and light-triggered drug release | Staphylococcus aureus [274] | wound healing |
| Prussian blue and tannic acid-loaded polyacrylamide Hydrogel | Hydrogel | NIR | Photothermal therapy | Staphylococcus aureus [275] | wound healing |
| Curcumin-based metal-organic framework + vancomycin, and chitosan | Nanoparticles in hydrogels | NIR | Bacterial capturing, Zn ions, and antibiotic release | Staphylococcus aureus [276] | wound healing |
| TiO2 nanorod array | Functionalized composite surface | NIR | Hyperthermia, ROS generation, and bacterial membrane puncture | Staphylococcus aureus and Escherichia coli [277] | bone implants |
| Chitosan microspheres loaded with rose bengal and polypyrrole in PVA hydrogel | Nanoparticles in hydrogels | Visible and NIR | Photothermal and photodynamic therapy | Staphylococcus aureus and Escherichia coli [278] | wound healing |
| Aloe-Emodin/Carbon Nanoparticle Hybrid PEG hydrogel | Nanoparticles in hydrogels | NIR | ROS generation and drug release | Staphylococcus aureus and Escherichia coli [279] | wound healing |
| PVA-(GS-Linker-MPEG) hydrogel loaded with Cy3/Cy5-silica NPs and UCNPs | Nanoparticles in hydrogels | NIR | NIR-UV conversion and light-triggered antibiotic release | Staphylococcus aureus [280] | infected wounds |
| Rose bengal/graphene oxide/PVA/chitosan hybrid hydrogel | Nanoparticles in hydrogels | Visible and NIR | Photothermal therapy and ROS generation | Staphylococcus aureus and Escherichia coli [281] | wound healing |
| Photochromic low-MW supramolecular hydrogel, drug loaded | Hydrogel | Visible | Light-triggered hydrogel dissolution and drug release | Escherichia coli [282] | |
| Catechol-conjugated poly(vinylpyrrolidone) sulfobetaine/polyaniline | Polymer coating | NIR | Photothermal therapy | Staphylococcus aureus and Escherichia coli [283] | |
| Pectin—Ag/AgCl/ZnO plasmonic hybrid nanocomposites | Nanoparticles in hydrogels | Visible | Photocatalytic nanostructures, ROS generation, and Zn and Ag ion release | Staphylococcus aureus and Escherichia coli [284] | |
| Berberine-microalgae/carboxymethyl chitosan/sodium alginate hydrogel | Hydrogel | Visible | Light-triggered drug release, ROS generation, QS downregulation, and inhibition and destruction of the biofilm | Methicillin-resistant Staphylococcus aureus [285] | infected wounds |
| Chlorinated e6-methacrylated silk fibroin | Film | UV and NIR | Photodynamic therapy | Staphylococcus aureus [286] | surgical wounds |
| Dopamine-folic acid hydrogel loaded with transition metal ions + carbon quantum dot-decorated ZnO NPs | Nanoparticles in hydrogels | Visible and NIR | Photothermal therapy, ROS generation, Zn ion release, and bacteria wall penetration | Staphylococcus aureus and Escherichia coli [287] | wound healing |

6.2. Light-Responsive Hydrogels and Polymeric Composite Structures
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mahamuni-Badiger, P.P.; Patil, P.M.; Badiger, M.V.; Patel, P.R.; Thorat-Gadgil, B.S.; Pandit, A.; Bohara, R.A. Biofilm formation to inhibition: Role of zinc oxide-based nanoparticles. Mater. Sci. Eng. C 2020, 108, 110319. [Google Scholar] [CrossRef] [PubMed]
- Uruén, C.; Chopo-Escuin, G.; Tommassen, J.; Mainar-Jaime, R.; Arenas, J. Biofilms as Promoters of Bacterial Antibiotic Resistance and Tolerance. Antibiotics 2021, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Baylay, A.J.; Piddock, L.J.; Webber, M.A. Molecular mechanisms of antibiotic resistance—Part I. In Bacterial Resistance to Antibiotics—From Molecules to Man; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 1–26. [Google Scholar]
- Shi, J.; Yan, Y.; Links, M.G.; Li, L.; Dillon, J.-A.R.; Horsch, M.; Kusalik, A. Antimicrobial resistance genetic factor identification from whole-genome sequence data using deep feature selection. BMC Bioinform. 2019, 20, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Li, Z.; Li, L.; Sun, Y.; Shi, L.; Li, W.; Zhang, J.; Wu, Y.; Xu, H.; Wang, M. Detection of multidrug resistant pathogenic bacteria and novel complex class 1 integrons in campus atmospheric particulate matters. Sci. Total Environ. 2023, 856, 158976. [Google Scholar] [CrossRef] [PubMed]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Rice, L.B. Federal Funding for the Study of Antimicrobial Resistance in Nosocomial Pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Goverment of the United Kingdom: London, UK, 2016.
- Maleeva, G.; Wutz, D.; Rustler, K.; Nin-Hill, A.; Rovira, C.; Petukhova, E.; Bautista-Barrufet, A.; Gomila-Juaneda, A.; Scholze, P.; Peiretti, F.; et al. A photoswitchable GABA receptor channel blocker. Br. J. Pharmacol. 2019, 176, 2661–2677. [Google Scholar] [CrossRef] [Green Version]
- Gomila, A.M.J.; Rustler, K.; Maleeva, G.; Nin-Hill, A.; Wutz, D.; Bautista-Barrufet, A.; Rovira, X.; Bosch, M.; Mukhametova, E.; Petukhova, E.; et al. Photocontrol of Endogenous Glycine Receptors In Vivo. Cell Chem. Biol. 2020, 27, 1425–1433.e1427. [Google Scholar] [CrossRef]
- Castagna, R.; Maleeva, G.; Pirovano, D.; Matera, C.; Gorostiza, P. Donor-Acceptor Stenhouse Adduct Displaying Reversible Photoswitching in Water and Neuronal Activity. J. Am. Chem. Soc. 2022, 144, 15595–15602. [Google Scholar] [CrossRef]
- Reiner, A.; Isacoff, E.Y. Tethered ligands reveal glutamate receptor desensitization depends on subunit occupancy. Nat. Chem. Biol. 2014, 10, 273–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riefolo, F.; Matera, C.; Garrido-Charles, A.; Gomila, A.M.J.; Sortino, R.; Agnetta, L.; Claro, E.; Masgrau, R.; Holzgrabe, U.; Batlle, M.; et al. Optical Control of Cardiac Function with a Photoswitchable Muscarinic Agonist. J. Am. Chem. Soc. 2019, 141, 7628–7636. [Google Scholar] [CrossRef]
- Riefolo, F.; Sortino, R.; Matera, C.; Claro, E.; Preda, B.; Vitiello, S.; Traserra, S.; Jimenez, M.; Gorostiza, P. Rational Design of Photochromic Analogues of Tricyclic Drugs. J. Med. Chem. 2021, 64, 9259–9270. [Google Scholar] [CrossRef] [PubMed]
- Matera, C.; Calvé, P.; Casadó-Anguera, V.; Sortino, R.; Gomila, A.M.; Moreno, E.; Gener, T.; Delgado-Sallent, C.; Nebot, P.; Costazza, D. Reversible photocontrol of dopaminergic transmission in wild-type animals. Int. J. Mol. Sci. 2022, 23, 10114. [Google Scholar] [CrossRef] [PubMed]
- Nevola, L.; Varese, M.; Martin-Quiros, A.; Mari, G.; Eckelt, K.; Gorostiza, P.; Giralt, E. Targeted Nanoswitchable Inhibitors of Protein-Protein Interactions Involved in Apoptosis. ChemMedChem 2019, 14, 100–106. [Google Scholar] [CrossRef]
- Eli, S.; Castagna, R.; Mapelli, M.; Parisini, E. Recent Approaches to the Identification of Novel Microtubule-Targeting Agents. Front. Mol. Biosci. 2022, 9, 841777. [Google Scholar] [CrossRef]
- Matera, C.; Gomila, A.M.J.; Camarero, N.; Libergoli, M.; Soler, C.; Gorostiza, P. Photoswitchable Antimetabolite for Targeted Photoactivated Chemotherapy. J. Am. Chem. Soc. 2018, 140, 15764–15773. [Google Scholar] [CrossRef] [Green Version]
- Lerch, M.M.; Hansen, M.J.; van Dam, G.M.; Szymanski, W.; Feringa, B.L. Emerging Targets in Photopharmacology. Angew. Chem. Int. Ed. 2016, 55, 10978–10999. [Google Scholar] [CrossRef]
- Hull, K.; Morstein, J.; Trauner, D. In Vivo Photopharmacology. Chem. Rev 2018, 118, 10710–10747. [Google Scholar] [CrossRef]
- Fuchter, M.J. On the Promise of Photopharmacology Using Photoswitches: A Medicinal Chemist’s Perspective. J. Med. Chem. 2020, 63, 11436–11447. [Google Scholar] [CrossRef]
- Castagna, R.; Kolarski, D.; Durand-de Cuttoli, R.; Maleeva, G. Orthogonal Control of Neuronal Circuits and Behavior Using Photopharmacology. J. Mol. Neurosci. 2022, 72, 1433–1442. [Google Scholar] [CrossRef] [PubMed]
- Fedatto Abelha, T.; Rodrigues Lima Caires, A. Light-Activated Conjugated Polymers for Antibacterial Photodynamic and Photothermal Therapy. Adv. NanoBiomed Res. 2021, 1, 2100012. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, W.; Xu, Q.; Zheng, Y. Progress in Antibacterial Hydrogel Dressing. Gels 2022, 8, 503. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zou, L.; Wang, J.; Jin, Q.; Ji, J. Stimuli-responsive nanoplatforms for antibacterial applications. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2022, 14, e1775. [Google Scholar] [CrossRef]
- Chakraborty, N.; Jha, D.; Roy, I.; Kumar, P.; Gaurav, S.S.; Marimuthu, K.; Ng, O.T.; Lakshminarayanan, R.; Verma, N.K.; Gautam, H.K. Nanobiotics against antimicrobial resistance: Harnessing the power of nanoscale materials and technologies. J. Nanobiotechnology 2022, 20, 375. [Google Scholar] [CrossRef]
- Li, X.; Wu, B.; Chen, H.; Nan, K.; Jin, Y.; Sun, L.; Wang, B. Recent developments in smart antibacterial surfaces to inhibit biofilm formation and bacterial infections. J. Mater. Chem. B 2018, 6, 4274–4292. [Google Scholar] [CrossRef]
- Pierau, L.; Versace, D.-L. Light and hydrogels: A new generation of antimicrobial materials. Materials 2021, 14, 787. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef]
- Vestby, L.K.; Grønseth, T.; Simm, R.; Nesse, L.L. Bacterial biofilm and its role in the pathogenesis of disease. Antibiotics 2020, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Fulaz, S.; Vitale, S.; Quinn, L.; Casey, E. Nanoparticle–biofilm interactions: The role of the EPS matrix. Trends Microbiol. 2019, 27, 915–926. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.-C.; Wingender, J. Relevance of microbial extracellular polymeric substances (EPSs)—Part I: Structural and ecological aspects. Water Sci. Technol. 2001, 43, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Kovach, K.; Davis-Fields, M.; Irie, Y.; Jain, K.; Doorwar, S.; Vuong, K.; Dhamani, N.; Mohanty, K.; Touhami, A.; Gordon, V.D. Evolutionary adaptations of biofilms infecting cystic fibrosis lungs promote mechanical toughness by adjusting polysaccharide production. NPJ Biofilms Microbiomes 2017, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.; Castillo-Pino, E. An introduction to the epidemiology and burden of urinary tract infections. Ther. Adv. Urol. 2019, 11, 1756287219832172. [Google Scholar] [CrossRef] [Green Version]
- Slavkin, H.C. Biofilms, Microbial Ecology and Antoni van Leeuwenhoek. J. Am. Dent. Assoc. 1997, 128, 492–495. [Google Scholar] [CrossRef]
- Sedarat, Z.; Taylor-Robinson, A.W. A Consideration of antibacterial agent efficacies in the treatment and prevention of formation of Staphylococcus aureus biofilm. J. Microbiol. Infect. Dis. 2019, 9, 167–172. [Google Scholar] [CrossRef] [Green Version]
- Allcock, S.; Young, E.H.; Holmes, M.; Gurdasani, D.; Dougan, G.; Sandhu, M.S.; Solomon, L.; Török, M. Antimicrobial resistance in human populations: Challenges and opportunities. Glob. Health Epidemiol. Genom. 2017, 2, e4. [Google Scholar] [CrossRef] [Green Version]
- López, D.; Vlamakis, H.; Kolter, R. Biofilms. Cold Spring Harb. Perspect. Biol. 2010, 2, a000398. [Google Scholar] [CrossRef]
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial biofilms: Development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era. Cold Spring Harb. Perspect. Med. 2013, 3, a010306. [Google Scholar] [CrossRef] [Green Version]
- Petrova, O.E.; Sauer, K. Sticky situations: Key components that control bacterial surface attachment. J. Bacteriol. 2012, 194, 2413–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galie, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef] [PubMed]
- Solano, C.; Echeverz, M.; Lasa, I. Biofilm dispersion and quorum sensing. Curr. Opin. Microbiol. 2014, 18, 96–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Rivas, F.; Ripolles-Avila, C.; Fontecha-Umaña, F.; Ríos-Castillo, A.G.; Rodríguez-Jerez, J.J. Biofilms in the spotlight: Detection, quantification, and removal methods. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1261–1276. [Google Scholar] [CrossRef] [Green Version]
- Srey, S.; Jahid, I.K.; Ha, S.-D. Biofilm formation in food industries: A food safety concern. Food Control 2013, 31, 572–585. [Google Scholar] [CrossRef]
- Møretrø, T.; Heir, E.; Nesse, L.L.; Vestby, L.K.; Langsrud, S. Control of Salmonella in food related environments by chemical disinfection. Food Res. Int. 2012, 45, 532–544. [Google Scholar] [CrossRef]
- Ciofu, O.; Moser, C.; Jensen, P.Ø.; Høiby, N. Tolerance and resistance of microbial biofilms. Nat. Rev. Microbiol. 2022, 20, 621–635. [Google Scholar] [CrossRef]
- Mukherjee, S.; Bassler, B.L. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Lal, A. Quorum sensing: How bacteria talk to each other. Resonance 2009, 14, 866–871. [Google Scholar] [CrossRef]
- Atkinson, S.; Williams, P. Quorum sensing and social networking in the microbial world. J. R. Soc. Interface 2009, 6, 959–978. [Google Scholar] [CrossRef] [Green Version]
- Haque, S.; Yadav, D.K.; Bisht, S.C.; Yadav, N.; Singh, V.; Dubey, K.K.; Jawed, A.; Wahid, M.; Dar, S.A. Quorum sensing pathways in Gram-positive and-negative bacteria: Potential of their interruption in abating drug resistance. J. Chemother. 2019, 31, 161–187. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, F.; Ding, G.; Li, J.; Guo, X.; Zhu, J.; Zhou, L.; Cai, S.; Liu, X.; Luo, Y.; et al. Acyl homoserine lactone-based quorum sensing in a methanogenic archaeon. ISME J. 2012, 6, 1336–1344. [Google Scholar] [CrossRef] [Green Version]
- Withers, H.; Swift, S.; Williams, P. Quorum sensing as an integral component of gene regulatory networks in Gram-negative bacteria. Curr. Opin. Microbiol. 2001, 4, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Ziemichód, A.; Skotarczak, B. QS–systems communication of gram-positive bacterial cells. Acta Biol. 2017, 24, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhao, X. Effects of quorum sensing on the biofilm formation and viable but non-culturable state. Food Res. Int. 2020, 137, 109742. [Google Scholar] [CrossRef] [PubMed]
- Nealson, K.H.; Hastings, J. Bacterial bioluminescence: Its control and ecological significance. Microbiol. Rev. 1979, 43, 496–518. [Google Scholar] [CrossRef]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, I.; Silva, L.R.; Giaouris, E.D.; Melo, L.F.; Simões, M. Quorum sensing in food spoilage and natural-based strategies for its inhibition. Food Res. Int. 2020, 127, 108754. [Google Scholar] [CrossRef] [PubMed]
- Arciola, C.R.; Campoccia, D.; Speziale, P.; Montanaro, L.; Costerton, J.W. Biofilm formation in Staphylococcus implant infections. A review of molecular mechanisms and implications for biofilm-resistant materials. Biomaterials 2012, 33, 5967–5982. [Google Scholar] [CrossRef] [PubMed]
- Spangler, J.R.; Dean, S.N.; Leary, D.H.; Walper, S.A. Response of lactobacillus plantarum WCFS1 to the gram-negative pathogen-associated quorum sensing molecule N-3-oxododecanoyl homoserine lactone. Front. Microbiol. 2019, 10, 715. [Google Scholar] [CrossRef] [Green Version]
- Ji, G.; Beavis, R.C.; Novick, R.P. Cell density control of staphylococcal virulence mediated by an octapeptide pheromone. Proc. Natl. Acad. Sci. USA 1995, 92, 12055–12059. [Google Scholar] [CrossRef]
- Eberhard, A.; Burlingame, A.L.; Eberhard, C.; Kenyon, G.L.; Nealson, K.H.; Oppenheimer, N. Structural identification of autoinducer of Photobacterium fischeri luciferase. Biochemistry 1981, 20, 2444–2449. [Google Scholar] [CrossRef]
- Chen, X.; Schauder, S.; Potier, N.; Van Dorsselaer, A.; Pelczer, I.; Bassler, B.L.; Hughson, F.M. Structural identification of a bacterial quorum-sensing signal containing boron. Nature 2002, 415, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Dizin, E.; Hu, X.; Wavreille, A.-S.; Park, J.; Pei, D. S-Ribosylhomocysteinase (LuxS) is a mononuclear iron protein. Biochemistry 2003, 42, 4717–4726. [Google Scholar] [CrossRef] [PubMed]
- Flavier, A.B.; Ganova-Raeva, L.M.; Schell, M.A.; Denny, T.P. Hierarchical autoinduction in Ralstonia solanacearum: Control of acyl-homoserine lactone production by a novel autoregulatory system responsive to 3-hydroxypalmitic acid methyl ester. J. Bacteriol. 1997, 179, 7089–7097. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.H.; He, Y.; Gao, Y.; Wu, J.E.; Dong, Y.H.; He, C.; Wang, S.X.; Weng, L.X.; Xu, J.L.; Tay, L. A bacterial cell–cell communication signal with cross-kingdom structural analogues. Mol. Microbiol. 2004, 51, 903–912. [Google Scholar] [CrossRef]
- Ohnishi, Y.; Kameyama, S.; Onaka, H.; Horinouchi, S. The A-factor regulatory cascade leading to streptomycin biosynthesis in Streptomyces griseus: Identification of a target gene of the A-factor receptor. Mol. Microbiol. 1999, 34, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Holden, M.T.; Ram Chhabra, S.; De Nys, R.; Stead, P.; Bainton, N.J.; Hill, P.J.; Manefield, M.; Kumar, N.; Labatte, M.; England, D. Quorum-sensing cross talk: Isolation and chemical characterization of cyclic dipeptides from Pseudomonas aeruginosa and other gram-negative bacteria. Mol. Microbiol. 1999, 33, 1254–1266. [Google Scholar] [CrossRef]
- Diggle, S.P.; Winzer, K.; Chhabra, S.R.; Worrall, K.E.; Cámara, M.; Williams, P. The Pseudomonas aeruginosa quinolone signal molecule overcomes the cell density-dependency of the quorum sensing hierarchy, regulates rhl-dependent genes at the onset of stationary phase and can be produced in the absence of LasR. Mol. Microbiol. 2003, 50, 29–43. [Google Scholar] [CrossRef]
- Déziel, E.; Lépine, F.; Milot, S.; He, J.; Mindrinos, M.N.; Tompkins, R.G.; Rahme, L.G. Analysis of Pseudomonas aeruginosa 4-hydroxy-2-alkylquinolines (HAQs) reveals a role for 4-hydroxy-2-heptylquinoline in cell-to-cell communication. Proc. Natl. Acad. Sci. USA 2004, 101, 1339–1344. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, Y.; Ge, Y.; Zhu, X.; Pan, J. Regulatory mechanisms and promising applications of quorum sensing-inhibiting agents in control of bacterial biofilm formation. Front. Microbiol. 2020, 11, 589640. [Google Scholar] [CrossRef] [PubMed]
- Van Gennip, M.; Christensen, L.D.; Alhede, M.; Phipps, R.; Jensen, P.Ø.; Christophersen, L.; Pamp, S.J.; Moser, C.; Mikkelsen, P.J.; Koh, A.Y. Inactivation of the rhlA gene in Pseudomonas aeruginosa prevents rhamnolipid production, disabling the protection against polymorphonuclear leukocytes. Apmis 2009, 117, 537–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Beer, D.; Stoodley, P.; Roe, F.; Lewandowski, Z. Effects of biofilm structures on oxygen distribution and mass transport. Biotechnol. Bioeng. 1994, 43, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.; Alexandre, K.; Etienne, M. Tolerance and persistence of Pseudomonas aeruginosa in biofilms exposed to antibiotics: Molecular mechanisms, antibiotic strategies and therapeutic perspectives. Front. Microbiol. 2020, 11, 2057. [Google Scholar] [CrossRef]
- Keren, I.; Kaldalu, N.; Spoering, A.; Wang, Y.; Lewis, K. Persister cells and tolerance to antimicrobials. FEMS Microbiol. Lett. 2004, 230, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201. [Google Scholar] [CrossRef] [Green Version]
- Poole, K. Stress responses as determinants of antimicrobial resistance in Pseudomonas aeruginosa: Multidrug efflux and more. Can. J. Microbiol. 2014, 60, 783–791. [Google Scholar] [CrossRef]
- Macia, M.; Rojo-Molinero, E.; Oliver, A. Antimicrobial susceptibility testing in biofilm-growing bacteria. Clin. Microbiol. Infect. 2014, 20, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Laslop, N.; Mankin, A.S. Context-Specific Action of Ribosomal Antibiotics. Annu. Rev. Microbiol. 2018, 72, 185–207. [Google Scholar] [CrossRef]
- Greulich, P.; Scott, M.; Evans, M.R.; Allen, R.J. Growth-dependent bacterial susceptibility to ribosome-targeting antibiotics. Mol. Syst. Biol. 2015, 11, 796. [Google Scholar] [CrossRef]
- Levin, B.R.; Rozen, D.E. Non-inherited antibiotic resistance. Nat. Rev. Microbiol. 2006, 4, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Dalebroux, Z.D.; Swanson, M.S. ppGpp: Magic beyond RNA polymerase. Nat. Rev. Microbiol. 2012, 10, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Salzer, A.; Wolz, C. Role of (p)ppGpp in antibiotic resistance, tolerance, persistence and survival in Firmicutes. Microlife 2023, 4, uqad009. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.N.; Porse, A.; Abdelsamad, A.; Sommer, M.; Høiby, N.; Ciofu, O. Lack of the Major Multifunctional Catalase KatA in Pseudomonas aeruginosa Accelerates Evolution of Antibiotic Resistance in Ciprofloxacin-Treated Biofilms. Antimicrob. Agents Chemother. 2019, 63, 10–1128. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.; Joshi-Datar, A.; Lepine, F.; Bauerle, E.; Olakanmi, O.; Beer, K.; McKay, G.; Siehnel, R.; Schafhauser, J.; Wang, Y.; et al. Active Starvation Responses Mediate Antibiotic Tolerance in Biofilms and Nutrient-Limited Bacteria. Science 2011, 334, 982–986. [Google Scholar] [CrossRef] [Green Version]
- Pamp, S.J.; Gjermansen, M.; Johansen, H.K.; Tolker-Nielsen, T. Tolerance to the antimicrobial peptide colistin in Pseudomonas aeruginosa biofilms is linked to metabolically active cells, and depends on the pmr and mexAB-oprM genes. Mol. Microbiol. 2008, 68, 223–240. [Google Scholar] [CrossRef]
- Herrmann, G.; Yang, L.; Wu, H.; Song, Z.; Wang, H.; Høiby, N.; Ulrich, M.; Molin, S.; Riethmüller, J.; Döring, G. Colistin-tobramycin combinations are superior to monotherapy concerning the killing of biofilm Pseudomonas aeruginosa. J. Infect. Dis. 2010, 202, 1585–1592. [Google Scholar] [CrossRef] [Green Version]
- Fraud, S.; Poole, K. Oxidative stress induction of the MexXY multidrug efflux genes and promotion of aminoglycoside resistance development in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2011, 55, 1068–1074. [Google Scholar] [CrossRef] [Green Version]
- Fetar, H.; Gilmour, C.; Klinoski, R.; Daigle, D.M.; Dean, C.R.; Poole, K. mexEF-oprN multidrug efflux operon of Pseudomonas aeruginosa: Regulation by the MexT activator in response to nitrosative stress and chloramphenicol. Antimicrob. Agents Chemother. 2011, 55, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Fraud, S.; Campigotto, A.J.; Chen, Z.; Poole, K. MexCD-OprJ multidrug efflux system of Pseudomonas aeruginosa: Involvement in chlorhexidine resistance and induction by membrane-damaging agents dependent upon the AlgU stress response sigma factor. Antimicrob. Agents Chemother. 2008, 52, 4478–4482. [Google Scholar] [CrossRef] [Green Version]
- Askoura, M.; Mottawea, W.; Abujamel, T.; Taher, I. Efflux pump inhibitors (EPIs) as new antimicrobial agents against Pseudomonas aeruginosa. Libyan J. Med. 2011, 6, 5870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How antibiotics kill bacteria: From targets to networks. Nat. Rev. Microbiol. 2010, 8, 423–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, P.S.; Franklin, M.J.; Williamson, K.S.; Folsom, J.P.; Boegli, L.; James, G.A. Contribution of stress responses to antibiotic tolerance in Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 2015, 59, 3838–3847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, J.; Schurr, M.J.; Sauer, K. The MerR-like regulator BrlR confers biofilm tolerance by activating multidrug efflux pumps in Pseudomonas aeruginosa biofilms. J. Bacteriol. 2013, 195, 3352–3363. [Google Scholar] [CrossRef] [Green Version]
- Poudyal, B.; Sauer, K. The PA3177 gene encodes an active diguanylate cyclase that contributes to biofilm antimicrobial tolerance but not biofilm formation by Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2018, 62, 10-1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Król, J.E.; Wojtowicz, A.J.; Rogers, L.M.; Heuer, H.; Smalla, K.; Krone, S.M.; Top, E.M. Invasion of E. coli biofilms by antibiotic resistance plasmids. Plasmid 2013, 70, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Usui, M.; Yoshii, Y.; Thiriet-Rupert, S.; Ghigo, J.-M.; Beloin, C. Intermittent antibiotic treatment of bacterial biofilms favors the rapid evolution of resistance. Commun. Biol. 2023, 6, 275. [Google Scholar] [CrossRef]
- Colizzi, S.E.; van Dijk, B.; Merks, R.M.H.; Rozen, D.E.; Vroomans, R.M.A. Evolution of genome fragility enables microbial division of labor. Mol. Syst. Biol. 2023, 19, e11353. [Google Scholar] [CrossRef]
- Soto, S.M. Role of efflux pumps in the antibiotic resistance of bacteria embedded in a biofilm. Virulence 2013, 4, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Buroni, S.; Matthijs, N.; Spadaro, F.; Van Acker, H.; Scoffone, V.C.; Pasca, M.R.; Riccardi, G.; Coenye, T. Differential roles of RND efflux pumps in antimicrobial drug resistance of sessile and planktonic Burkholderia cenocepacia cells. Antimicrob. Agents Chemother. 2014, 58, 7424–7429. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, K.; Furukawa, S.; Ogihara, H.; Morinaga, Y. Roles of multidrug efflux pumps on the biofilm formation of Escherichia coli K-12. Biocontrol Sci. 2011, 16, 69–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooun, A.; Liu, S.; Lewis, K. A dose-response study of antibiotic resistance in Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 2000, 44, 640–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenover, F.C. Mechanisms of antimicrobial resistance in bacteria. Am. J. Med. 2006, 119, S3–S10. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, D.; Das Talukdar, A.; Dutta Choudhury, M.; Maurya, A.P.; Paul, D.; Dhar Chanda, D.; Chakravorty, A.; Bhattacharjee, A. Transcriptional analysis of MexAB-OprM efflux pumps system of Pseudomonas aeruginosa and its role in carbapenem resistance in a tertiary referral hospital in India. PLoS ONE 2015, 10, e0133842. [Google Scholar] [CrossRef]
- Suresh, M.; Nithya, N.; Jayasree, P.; Vimal, K.; Manish Kumar, P. Mutational analyses of regulatory genes, mexR, nalC, nalD and mexZ of mexAB-oprM and mexXY operons, in efflux pump hyperexpressing multidrug-resistant clinical isolates of Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2018, 34, 83. [Google Scholar] [CrossRef]
- Paxman, J.J.; Lo, A.W.; Sullivan, M.J.; Panjikar, S.; Kuiper, M.; Whitten, A.E.; Wang, G.; Luan, C.-H.; Moriel, D.G.; Tan, L. Unique structural features of a bacterial autotransporter adhesin suggest mechanisms for interaction with host macromolecules. Nat. Commun. 2019, 10, 1967. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Zhang, S.; Zhou, X.; Tian, S. Anti-biofilm and bacteriostatic effects of three flavonoid compounds on Streptococcus mutans. Biofouling 2023, 39, 245–256. [Google Scholar] [CrossRef]
- An, S.-J.; Namkung, J.-U.; Ha, K.-W.; Jun, H.-K.; Kim, H.Y.; Choi, B.-K. Inhibitory effect of d-arabinose on oral bacteria biofilm formation on titanium discs. Anaerobe 2022, 75, 102533. [Google Scholar] [CrossRef]
- Gong, H.; Hu, X.; Liao, M.; Fa, K.; Ciumac, D.; Clifton, L.A.; Sani, M.-A.; King, S.M.; Maestro, A.; Separovic, F. Structural disruptions of the outer membranes of gram-negative bacteria by rationally designed amphiphilic antimicrobial peptides. ACS Appl. Mater. Interfaces 2021, 13, 16062–16074. [Google Scholar] [CrossRef]
- Angsantikul, P.; Thamphiwatana, S.; Zhang, Q.; Spiekermann, K.; Zhuang, J.; Fang, R.H.; Gao, W.; Obonyo, M.; Zhang, L. Coating nanoparticles with gastric epithelial cell membrane for targeted antibiotic delivery against Helicobacter pylori infection. Adv. Ther. 2018, 1, 1800016. [Google Scholar] [CrossRef]
- Huebinger, R.M.; Stones, D.H.; de Souza Santos, M.; Carlson, D.L.; Song, J.; Vaz, D.P.; Keen, E.; Wolf, S.E.; Orth, K.; Krachler, A.M. Targeting bacterial adherence inhibits multidrug-resistant Pseudomonas aeruginosa infection following burn injury. Sci. Rep. 2016, 6, 39341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, W.; Jin, J.; Park, S.; Kim, J.-Y.; Lee, M.-J.; Sun, H.; Kwon, J.-S.; Lee, H.; Choi, S.-H.; Hong, J. Quantitative Interpretation of Hydration Dynamics Enabled the Fabrication of a Zwitterionic Antifouling Surface. ACS Appl. Mater. Interfaces 2020, 12, 7951–7965. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Deng, S.; Lu, Z.; She, Y.; Xie, J.; Cong, Z.; Zhang, W.; Liu, R. Using In Vivo Assessment on Host Defense Peptide Mimicking Polymer-Modified Surfaces for Combating Implant Infections. ACS Appl. Bio Mater. 2021, 4, 3811–3829. [Google Scholar] [CrossRef]
- Zhu, J.; Uliana, A.; Wang, J.; Yuan, S.; Li, J.; Tian, M.; Simoens, K.; Volodin, A.; Lin, J.; Bernaerts, K.; et al. Elevated salt transport of antimicrobial loose nanofiltration membranes enabled by copper nanoparticles via fast bioinspired deposition. J. Mater. Chem. A 2016, 4, 13211–13222. [Google Scholar] [CrossRef]
- Ďureje, J.; Prošek, Z.; Trejbal, J.; Potocký, Š.; Tesárek, P. Influence of oxygen and argon plasma treatment on wettability and surface morphology of polypropylene microfibers. Acta Polytech. CTU Proc. 2022, 34, 11–14. [Google Scholar] [CrossRef]
- Eckhart, L.; Fischer, H.; Barken, K.B.; Tolker-Nielsen, T.; Tschachler, E. DNase1L2 suppresses biofilm formation by Pseudomonas aeruginosa and Staphylococcus aureus. Br. J. Dermatol. 2007, 156, 1342–1345. [Google Scholar] [CrossRef] [PubMed]
- Alkawash, M.A.; Soothill, J.S.; Schiller, N.L. Alginate lyase enhances antibiotic killing of mucoid Pseudomonas aeruginosa in biofilms. Apmis 2006, 114, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Weldrick, P.J.; Hardman, M.J.; Paunov, V.N. Enhanced Clearing of Wound-Related Pathogenic Bacterial Biofilms Using Protease-Functionalized Antibiotic Nanocarriers. ACS Appl. Mater. Interfaces 2019, 11, 43902–43919. [Google Scholar] [CrossRef]
- Rajasekharan, S.K.; Ramesh, S. Cellulase inhibits Burkholderia cepacia biofilms on diverse prosthetic materials. Pol. J. Microbiol. 2013, 62, 327–330. [Google Scholar] [CrossRef]
- Kalpana, B.J.; Aarthy, S.; Pandian, S.K. Antibiofilm activity of α-amylase from Bacillus subtilis S8-18 against biofilm forming human bacterial pathogens. Appl. Biochem. Biotechnol. 2012, 167, 1778–1794. [Google Scholar] [CrossRef]
- Kalia, M.; Yadav, V.K.; Singh, P.K.; Dohare, S.; Sharma, D.; Narvi, S.S.; Agarwal, V. Designing quorum sensing inhibitors of Pseudomonas aeruginosa utilizing FabI: An enzymic drug target from fatty acid synthesis pathway. 3 Biotech 2019, 9, 40. [Google Scholar] [CrossRef]
- Storz, M.P.; Allegretta, G.; Kirsch, B.; Empting, M.; Hartmann, R.W. From in vitro to in cellulo: Structure–activity relationship of (2-nitrophenyl) methanol derivatives as inhibitors of PqsD in Pseudomonas aeruginosa. Org. Biomol. Chem. 2014, 12, 6094–6104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Lu, C. Biological activity and identification of a peptide inhibitor of LuxS from Streptococcus suis serotype 2. FEMS Microbiol. Lett. 2009, 294, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.-H.; Gusti, A.R.; Zhang, Q.; Xu, J.-L.; Zhang, L.-H. Identification of quorum-quenching N-acyl homoserine lactonases from Bacillus species. Appl. Environ. Microbiol. 2002, 68, 1754–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peppoloni, S.; Pericolini, E.; Colombari, B.; Pinetti, D.; Cermelli, C.; Fini, F.; Prati, F.; Caselli, E.; Blasi, E. The β-lactamase inhibitor boronic acid derivative SM23 as a new anti-Pseudomonas aeruginosa biofilm. Front. Microbiol. 2020, 11, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.S.; Ryu, E.-J.; Li, L.; Choi, B.-K.; Kim, B.M. New bicyclic brominated furanones as potent autoinducer-2 quorum-sensing inhibitors against bacterial biofilm formation. Eur. J. Med. Chem. 2017, 137, 76–87. [Google Scholar] [CrossRef]
- Scoffone, V.C.; Chiarelli, L.R.; Makarov, V.; Brackman, G.; Israyilova, A.; Azzalin, A.; Forneris, F.; Riabova, O.; Savina, S.; Coenye, T. Discovery of new diketopiperazines inhibiting Burkholderia cenocepacia quorum sensing in vitro and in vivo. Sci. Rep. 2016, 6, 32487. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Mei, Y.; He, B.; Sun, X.; Li, J. Reducing quorum sensing-mediated virulence factor expression and biofilm formation in hafnia alvei by using the potential quorum sensing inhibitor L-carvone. Front. Microbiol. 2019, 9, 3324. [Google Scholar] [CrossRef]
- Czajkowski, R.; Krzyżanowska, D.; Karczewska, J.; Atkinson, S.; Przysowa, J.; Lojkowska, E.; Williams, P.; Jafra, S. Inactivation of AHLs by Ochrobactrum sp. A44 depends on the activity of a novel class of AHL acylase. Environ. Microbiol. Rep. 2011, 3, 59–68. [Google Scholar] [CrossRef]
- Yu, L.; Li, W.; Zhang, M.; Cui, Y.; Chen, X.; Ni, J.; Yu, L.; Shang, F.; Xue, T. Imidazole decreases the ampicillin resistance of an Escherichia coli strain isolated from a cow with mastitis by inhibiting the function of autoinducer 2. J. Dairy Sci. 2018, 101, 3356–3362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, B.; Eberl, L.; Feucht, W.; Polster, J. Influence of polyphenols on bacterial biofilm formation and quorum-sensing. Z. Für Naturforschung C 2003, 58, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Fujii, K.; Yamasaki, O.; Oono, T.; Iwatsuki, K. Antibacterial action of several tannins against Staphylococcus aureus. J. Antimicrob. Chemother. 2001, 48, 487–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.-H.; Hu, Z.-Q.; Shimamura, T.; Hara, Y. Inhibition by epigallocatechin gallate (EGCg) of conjugative R plasmid transfer in Escherichia coli. J. Infect. Chemother. 2001, 7, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Goo, E.; Yu, S.; Choi, O.; Lee, J.; Kim, J.; Kim, H.; Igarashi, J.; Suga, H.; Moon, J.S. Small-molecule inhibitor binding to an N-acyl-homoserine lactone synthase. Proc. Natl. Acad. Sci. USA 2011, 108, 12089–12094. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M.; D’morris, S.; Paul, V.; Warrier, S.; Vasudevan, A.K.; Vanuopadath, M.; Nair, S.S.; Paul-Prasanth, B.; Mohan, C.G.; Biswas, R. Mechanistic understanding of Phenyllactic acid mediated inhibition of quorum sensing and biofilm development in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2017, 101, 8223–8236. [Google Scholar] [CrossRef] [PubMed]
- Kalia, V.C. Quorum sensing inhibitors: An overview. Biotechnol. Adv. 2013, 31, 224–245. [Google Scholar] [CrossRef]
- Hentzer, M.; Wu, H.; Andersen, J.B.; Riedel, K.; Rasmussen, T.B.; Bagge, N.; Kumar, N.; Schembri, M.A.; Song, Z.; Kristoffersen, P. Attenuation of Pseudomonas aeruginosa virulence by quorum sensing inhibitors. EMBO J. 2003, 22, 3803–3815. [Google Scholar] [CrossRef]
- Aliyu, A.B.; Koorbanally, N.A.; Moodley, B.; Singh, P.; Chenia, H.Y. Quorum sensing inhibitory potential and molecular docking studies of sesquiterpene lactones from Vernonia blumeoides. Phytochemistry 2016, 126, 23–33. [Google Scholar] [CrossRef]
- Vandeputte, O.M.; Kiendrebeogo, M.; Rasamiravaka, T.; Stevigny, C.; Duez, P.; Rajaonson, S.; Diallo, B.; Mol, A.; Baucher, M.; El Jaziri, M. The flavanone naringenin reduces the production of quorum sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Microbiology 2011, 157, 2120–2132. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Li, T.; Gu, Y.; Zhang, Q.; Wang, E.; Li, W.; Wang, X.; Li, Y.; Li, H. Design, Synthesis, and Biological Evaluation of N-Acyl-Homoserine Lactone Analogs of Quorum Sensing in Pseudomonas aeruginosa. Front. Chem. 2022, 10, 948687. [Google Scholar] [CrossRef]
- Truchado, P.; Gimenez-Bastida, J.-A.; Larrosa, M.; Castro-Ibáñez, I.; Espín, J.C.; Tomas-Barberan, F.A.; García-Conesa, M.T.; Allende, A. Inhibition of quorum sensing (QS) in Yersinia enterocolitica by an orange extract rich in glycosylated flavanones. J. Agric. Food Chem. 2012, 60, 8885–8894. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Zhang, M. Analysis of the antibacterial effect of an Edwardsiella tarda LuxS inhibitor. Springerplus 2016, 5, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, E.-J.; Sim, J.; Sim, J.; Lee, J.; Choi, B.-K. D-Galactose as an autoinducer 2 inhibitor to control the biofilm formation of periodontopathogens. J. Microbiol. 2016, 54, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Curtis, M.M.; Russell, R.; Moreira, C.G.; Adebesin, A.M.; Wang, C.; Williams, N.S.; Taussig, R.; Stewart, D.; Zimmern, P.; Lu, B. QseC inhibitors as an antivirulence approach for Gram-negative pathogens. MBio 2014, 5, e02165-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Packiavathy, I.A.S.V.; Agilandeswari, P.; Musthafa, K.S.; Pandian, S.K.; Ravi, A.V. Antibiofilm and quorum sensing inhibitory potential of Cuminum cyminum and its secondary metabolite methyl eugenol against Gram negative bacterial pathogens. Food Res. Int. 2012, 45, 85–92. [Google Scholar] [CrossRef]
- El-Shaer, S.; Shaaban, M.; Barwa, R.; Hassan, R. Control of quorum sensing and virulence factors of Pseudomonas aeruginosa using phenylalanine arginyl β-naphthylamide. J. Med. Microbiol. 2016, 65, 1194–1204. [Google Scholar] [CrossRef] [PubMed]
- Kitzenberg, D.A.; Lee, J.S.; Mills, K.B.; Kim, J.-S.; Liu, L.; Vázquez-Torres, A.; Colgan, S.P.; Kao, D.J. Adenosine Awakens Metabolism to Enhance Growth-Independent Killing of Tolerant and Persister Bacteria across Multiple Classes of Antibiotics. Mbio 2022, 13, e00480-22. [Google Scholar] [CrossRef]
- Koul, A.; Dendouga, N.; Vergauwen, K.; Molenberghs, B.; Vranckx, L.; Willebrords, R.; Ristic, Z.; Lill, H.; Dorange, I.; Guillemont, J.; et al. Diarylquinolines target subunit c of mycobacterial ATP synthase. Nat. Chem. Biol. 2007, 3, 323–324. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Wood, T.K. Combatting persister cells with substituted indoles. Front. Microbiol. 2020, 11, 1565. [Google Scholar] [CrossRef]
- Palmqvist, N.; Foster, T.; Fitzgerald, J.R.; Josefsson, E.; Tarkowski, A. Fibronectin-binding proteins and fibrinogen-binding clumping factors play distinct roles in staphylococcal arthritis and systemic inflammation. J. Infect. Dis. 2005, 191, 791–798. [Google Scholar] [CrossRef]
- Yang, X.; Hou, J.; Tian, Y.; Zhao, J.; Sun, Q.; Zhou, S. Antibacterial surfaces: Strategies and applications. Sci. China Technol. Sci. 2022, 65, 1000–1010. [Google Scholar] [CrossRef]
- Paluch, E.; Piecuch, A.; Obłąk, E.; Lamch, Ł.; Wilk, K.A. Antifungal activity of newly synthesized chemodegradable dicephalic-type cationic surfactants. Colloids Surf. B Biointerfaces 2018, 164, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Piewngam, P.; Chiou, J.; Chatterjee, P.; Otto, M. Alternative approaches to treat bacterial infections: Targeting quorum-sensing. Expert Rev. Anti Infect. Ther. 2020, 18, 499–510. [Google Scholar] [CrossRef]
- Kalia, V.C.; Patel, S.K.S.; Kang, Y.C.; Lee, J.-K. Quorum sensing inhibitors as antipathogens: Biotechnological applications. Biotechnol. Adv. 2019, 37, 68–90. [Google Scholar] [CrossRef]
- Vashistha, A.; Sharma, N.; Nanaji, Y.; Kumar, D.; Singh, G.; Barnwal, R.P.; Yadav, A.K. Quorum sensing inhibitors as Therapeutics: Bacterial biofilm inhibition. Bioorganic Chem. 2023, 136, 106551. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.-H.; Xu, J.-L.; Li, X.-Z.; Zhang, L.-H. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc. Natl. Acad. Sci. USA 2000, 97, 3526–3531. [Google Scholar] [CrossRef] [PubMed]
- Marin, S.D.L.; Xu, Y.; Meijler, M.M.; Janda, K.D. Antibody catalyzed hydrolysis of a quorum sensing signal found in Gram-negative bacteria. Bioorganic Med. Chem. Lett. 2007, 17, 1549–1552. [Google Scholar] [CrossRef]
- Park, J.; Jagasia, R.; Kaufmann, G.F.; Mathison, J.C.; Ruiz, D.I.; Moss, J.A.; Meijler, M.M.; Ulevitch, R.J.; Janda, K.D. Infection control by antibody disruption of bacterial quorum sensing signaling. Chem. Biol. 2007, 14, 1119–1127. [Google Scholar] [CrossRef] [Green Version]
- Zang, T.; Lee, B.W.; Cannon, L.M.; Ritter, K.A.; Dai, S.; Ren, D.; Wood, T.K.; Zhou, Z.S. A naturally occurring brominated furanone covalently modifies and inactivates LuxS. Bioorg. Med. Chem. Lett. 2009, 19, 6200–6204. [Google Scholar] [CrossRef] [Green Version]
- Vani, S.; Vadakkan, K.; Mani, B. A narrative review on bacterial biofilm: Its formation, clinical aspects and inhibition strategies. Future J. Pharm. Sci. 2023, 9, 50. [Google Scholar] [CrossRef]
- Milly, T.A.; Tal-Gan, Y. Targeting peptide-based quorum sensing systems for the treatment of gram-positive bacterial infections. Pept. Sci. 2023, 115, e24298. [Google Scholar] [CrossRef] [PubMed]
- Geske, G.D.; O’Neill, J.C.; Miller, D.M.; Wezeman, R.J.; Mattmann, M.E.; Lin, Q.; Blackwell, H.E. Comparative analyses of N-acylated homoserine lactones reveal unique structural features that dictate their ability to activate or inhibit quorum sensing. Chembiochem 2008, 9, 389–400. [Google Scholar] [CrossRef] [Green Version]
- Chambial, S.; Dwivedi, S.; Shukla, K.K.; John, P.J.; Sharma, P. Vitamin C in disease prevention and cure: An overview. Indian J. Clin. Biochem. 2013, 28, 314–328. [Google Scholar] [CrossRef] [Green Version]
- Ren, D.; Bedzyk, L.A.; Ye, R.W.; Thomas, S.M.; Wood, T.K. Differential gene expression shows natural brominated furanones interfere with the autoinducer-2 bacterial signaling system of Escherichia coli. Biotechnol. Bioeng. 2004, 88, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Song, Z.; Hentzer, M.; Andersen, J.B.; Molin, S.; Givskov, M.; Høiby, N. Synthetic furanones inhibit quorum-sensing and enhance bacterial clearance in Pseudomonas aeruginosa lung infection in mice. J. Antimicrob. Chemother. 2004, 53, 1054–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, M. Overview of antibacterial, antitoxin, antiviral, and antifungal activities of tea flavonoids and teas. Mol. Nutr. Food Res. 2007, 51, 116–134. [Google Scholar] [CrossRef]
- Gopu, V.; Meena, C.K.; Shetty, P.H. Quercetin influences quorum sensing in food borne bacteria: In-vitro and in-silico evidence. PLoS ONE 2015, 10, e0134684. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Lau, G.W. Inhibition of competence development, horizontal gene transfer and virulence in Streptococcus pneumoniae by a modified competence stimulating peptide. PLoS Pathog. 2011, 7, e1002241. [Google Scholar] [CrossRef] [Green Version]
- Koirala, B.; Lin, J.; Lau, G.W.; Tal-Gan, Y. Development of a Dominant Negative Competence-Stimulating Peptide (dnCSP) that Attenuates Streptococcus pneumoniae Infectivity in a Mouse Model of Acute Pneumonia. ChemBioChem 2018, 19, 2380–2386. [Google Scholar] [CrossRef]
- Peng, H.; Cheng, Y.; Ni, N.; Li, M.; Choudhary, G.; Chou, H.T.; Lu, C.D.; Tai, P.C.; Wang, B. Synthesis and evaluation of new antagonists of bacterial quorum sensing in Vibrio harveyi. ChemMedChem Chem. Enabling Drug Discov. 2009, 4, 1457–1468. [Google Scholar]
- Luo, Y.; Yang, Q.; Zhang, D.; Yan, W. Mechanisms and control strategies of antibiotic resistance in pathological biofilms. J. Microbiol. Biotechnol. 2021, 31, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Muras, A.; Mallo, N.; Otero-Casal, P.; Pose-Rodríguez, J.M.; Otero, A. Quorum sensing systems as a new target to prevent biofilm-related oral diseases. Oral Dis. 2022, 28, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.H.; Choi, C.H. Role of LuxIR homologue AnoIR in Acinetobacter nosocomialis and the effect of virstatin on the expression of anoR gene. J. Microbiol. Biotechnol. 2015, 25, 1390–1400. [Google Scholar] [CrossRef] [PubMed]
- Munir, S.; Shah, A.A.; Shahid, M.; Manzoor, I.; Aslam, B.; Rasool, M.H.; Saeed, M.; Ayaz, S.; Khurshid, M. Quorum sensing interfering strategies and their implications in the management of biofilm-associated bacterial infections. Braz. Arch. Biol. Technol. 2020, 63, e20190555. [Google Scholar] [CrossRef]
- Jiang, Y.; Geng, M.; Bai, L. Targeting biofilms therapy: Current research strategies and development hurdles. Microorganisms 2020, 8, 1222. [Google Scholar] [CrossRef]
- Baelo, A.; Levato, R.; Julián, E.; Crespo, A.; Astola, J.; Gavaldà, J.; Engel, E.; Mateos-Timoneda, M.A.; Torrents, E. Disassembling bacterial extracellular matrix with DNase-coated nanoparticles to enhance antibiotic delivery in biofilm infections. J. Control. Release 2015, 209, 150–158. [Google Scholar] [CrossRef]
- Whitchurch, C.B.; Tolker-Nielsen, T.; Ragas, P.C.; Mattick, J.S. Extracellular DNA required for bacterial biofilm formation. Science 2002, 295, 1487. [Google Scholar] [CrossRef]
- Banerjee, A.; Chowdhury, P.; Bauri, K.; Saha, B.; De, P. Inhibition and eradication of bacterial biofilm using polymeric materials. Biomater. Sci. 2023, 11, 11–36. [Google Scholar] [CrossRef]
- Cornelissen, A.; Ceyssens, P.-J.; T’syen, J.; Van Praet, H.; Noben, J.-P.; Shaburova, O.V.; Krylov, V.N.; Volckaert, G.; Lavigne, R. The T7-related Pseudomonas putida phage φ15 displays virion-associated biofilm degradation properties. PLoS ONE 2011, 6, e18597. [Google Scholar] [CrossRef] [Green Version]
- Fleming, D.; Rumbaugh, K.P. Approaches to dispersing medical biofilms. Microorganisms 2017, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Iwase, T.; Uehara, Y.; Shinji, H.; Tajima, A.; Seo, H.; Takada, K.; Agata, T.; Mizunoe, Y. Staphylococcus epidermidis Esp inhibits Staphylococcus aureus biofilm formation and nasal colonization. Nature 2010, 465, 346–349. [Google Scholar] [CrossRef]
- Shukla, S.K.; Rao, T.S. Staphylococcus aureus biofilm removal by targeting biofilm-associated extracellular proteins. Indian J. Med. Res. 2017, 146, S1. [Google Scholar] [PubMed]
- Marx, C.; Gardner, S.; Harman, R.M.; Van de Walle, G.R. The mesenchymal stromal cell secretome impairs methicillin-resistant Staphylococcus aureus biofilms via cysteine protease activity in the equine model. Stem Cells Transl. Med. 2020, 9, 746–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, D.; Huang, C.; Huang, H.; Zhao, Y.; Khan, M.R.U.; Zhao, H.; Huang, L. Antibacterial Mechanism of Curcumin: A Review. Chem. Biodivers. 2020, 17, e2000171. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting microbial biofilms: Current and prospective therapeutic strategies. Nat. Rev. Microbiol. 2017, 15, 740–755. [Google Scholar] [CrossRef]
- Palomares-Navarro, J.J.; Bernal-Mercado, A.T.; González-Aguilar, G.A.; Ortega-Ramirez, L.A.; Martínez-Téllez, M.A.; Ayala-Zavala, J.F. Antibiofilm Action of Plant Terpenes in Salmonella Strains: Potential Inhibitors of the Synthesis of Extracellular Polymeric Substances. Pathogens 2023, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Eisha, S.; Park, S.; Morris, A.; Martin, I. Targeting Matrix Exopolysaccharides to Disrupt Pseudomonas aeruginosa Biofilms in Cystic Fibrosis. Med. Pharmacol. 2023. [Google Scholar] [CrossRef]
- Wood, T.K. Combatting bacterial persister cells. Biotechnol. Bioeng. 2016, 113, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.S.; Bekale, L.A.; Molchanova, N.; Nielsen, J.E.; Wright, M.; Bacacao, B.; Diamond, G.; Jenssen, H.v.; Santa Maria, P.L.; Barron, A.E. Anti-persister and Anti-biofilm Activity of Self-Assembled Antimicrobial Peptoid Ellipsoidal Micelles. ACS Infect. Dis. 2022, 8, 1823–1830. [Google Scholar] [CrossRef]
- Necula, G.; Bacalum, M.; Radu, M. Interaction of Tryptophan-and Arginine-Rich Antimicrobial Peptide with E. coli Outer Membrane—A Molecular Simulation Approach. Int. J. Mol. Sci. 2023, 24, 2005. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.W.; Mah, T.-F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Cao, J.; Zhou, C.; Liu, H.; Zhang, X.; Zhou, T. Biofilm formation restrained by subinhibitory concentrations of tigecyclin in Acinetobacter baumannii is associated with downregulation of efflux pumps. Chemotherapy 2017, 62, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Singh, A.; Khan, A.U. Nanoparticles as efflux pump and biofilm inhibitor to rejuvenate bactericidal effect of conventional antibiotics. Nanoscale Res. Lett. 2017, 12, 454. [Google Scholar] [CrossRef] [PubMed]
- Richmond, G.E.; Evans, L.P.; Anderson, M.J.; Wand, M.E.; Bonney, L.C.; Ivens, A.; Chua, K.L.; Webber, M.A.; Sutton, J.M.; Peterson, M.L. The Acinetobacter baumannii two-component system AdeRS regulates genes required for multidrug efflux, biofilm formation, and virulence in a strain-specific manner. MBio 2016, 7, e00430-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chetri, S. The culmination of multidrug-resistant efflux pumps vs. meager antibiotic arsenal era: Urgent need for an improved new generation of EPIs. Front. Microbiol. 2023, 14, 1149418. [Google Scholar] [CrossRef]
- Kaur, R.; Liu, S. Antibacterial surface design—Contact kill. Prog. Surf. Sci. 2016, 91, 136–153. [Google Scholar] [CrossRef]
- Schneider-Chaabane, A.; Bleicher, V.; Rau, S.; Al-Ahmad, A.; Lienkamp, K. Stimulus-Responsive Polyzwitterionic Surfaces Made from Itaconic Acid: Self-Triggered Antimicrobial Activity, Protein Repellency, and Cell Compatibility. ACS Appl. Mater. Interfaces 2020, 12, 21242–21253. [Google Scholar] [CrossRef]
- Liu, Z.; Yi, Y.; Song, L.; Chen, Y.; Tian, L.; Zhao, J.; Ren, L. Biocompatible mechano-bactericidal nanopatterned surfaces with salt-responsive bacterial release. Acta Biomater. 2022, 141, 198–208. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, W.; Qin, X.; Cui, M.; Guo, Y.; Wang, T.; Wang, K.; Shi, Z.; Zhang, C.; Li, W.; et al. Recent Progress on Bioinspired Antibacterial Surfaces for Biomedical Application. Biomimetics 2022, 7, 88. [Google Scholar] [CrossRef]
- Di Martino, M.; Sessa, L.; Di Matteo, M.; Panunzi, B.; Piotto, S.; Concilio, S. Azobenzene as Antimicrobial Molecules. Molecules 2022, 27, 5643. [Google Scholar] [CrossRef]
- Chen, F.; Ricken, J.; Xu, D.; Wegner, S.V. Bacterial Photolithography: Patterning Escherichia coli Biofilms with High Spatial Control Using Photocleavable Adhesion Molecules. Adv. Biosyst. 2019, 3, e1800269. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Riedel-Kruse, I.H. Biofilm Lithography enables high-resolution cell patterning via optogenetic adhesin expression. Proc. Natl. Acad. Sci. USA 2018, 115, 3698–3703. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Chavez, M.S.; Naughton, K.L.; Niman, C.M.; Atkinson, J.T.; Gralnick, J.A.; El-Naggar, M.Y.; Boedicker, J.Q. Light-Induced Patterning of Electroactive Bacterial Biofilms. ACS Synth. Biol. 2022, 11, 2327–2338. [Google Scholar] [CrossRef]
- Bruchmann, J.; Pini, I.; Gill, T.S.; Schwartz, T.; Levkin, P.A. Patterned SLIPS for the Formation of Arrays of Biofilm Microclusters with Defined Geometries. Adv. Healthc. Mater. 2017, 6, 1601082. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Li, J.; Bai, Y.; Zhang, Y.; Sun, H.; Zhang, X. A bacterial infection-microenvironment activated nanoplatform based on spiropyran-conjugated glycoclusters for imaging and eliminating of the biofilm. Chem. Eng. J. 2020, 399, 125787. [Google Scholar] [CrossRef]
- Chen, F.; Wegner, S.V. Blue Light Switchable Bacterial Adhesion as a Key Step toward the Design of Biofilms. ACS Synth. Biol. 2017, 6, 2170–2174. [Google Scholar] [CrossRef]
- Wei, J.; Jin, F. Illuminating bacterial behaviors with optogenetics. Curr. Opin. Solid State Mater. Sci. 2022, 26, 101023. [Google Scholar] [CrossRef]
- Hoffman, S.M.; Tang, A.Y.; Avalos, J.L. Optogenetics Illuminates Applications in Microbial Engineering. Annu. Rev. Chem. Biomol. Eng. 2022, 13, 373–403. [Google Scholar] [CrossRef]
- Lindner, F.; Diepold, A. Optogenetics in bacteria—Applications and opportunities. FEMS Microbiol. Rev. 2022, 46, fuab055. [Google Scholar] [CrossRef]
- Chen, F.; Wegner, S.V. Blue-Light-Switchable Bacterial Cell-Cell Adhesions Enable the Control of Multicellular Bacterial Communities. ACS Synth. Biol. 2020, 9, 1169–1180. [Google Scholar] [CrossRef]
- Fang, K.; Park, O.-J.; Hong, S.H. Controlling biofilms using synthetic biology approaches. Biotechnol. Adv. 2020, 40, 107518. [Google Scholar] [CrossRef]
- Hong, S.H.; Hegde, M.; Kim, J.; Wang, X.; Jayaraman, A.; Wood, T.K. Synthetic quorum-sensing circuit to control consortial biofilm formation and dispersal in a microfluidic device. Nat. Commun. 2012, 3, 613. [Google Scholar] [CrossRef] [Green Version]
- Van der Berg, J.; Velema, W.; Szymanski, W.; Driessen, A.; Feringa, B. Controlling the activity of quorum sensing autoinducers with light. Chem. Sci. 2015, 6, 3593–3598. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.J.; Hille, J.I.; Szymanski, W.; Driessen, A.J.; Feringa, B.L. Easily accessible, highly potent, photocontrolled modulators of bacterial communication. Chem 2019, 5, 1293–1301. [Google Scholar] [CrossRef]
- Lauxen, A.I.; Kobauri, P.; Wegener, M.; Hansen, M.J.; Galenkamp, N.S.; Maglia, G.; Szymanski, W.; Feringa, B.L.; Kuipers, O.P. Mechanism of resistance development in E. coli against TCAT, a trimethoprim-based photoswitchable antibiotic. Pharmaceuticals 2021, 14, 392. [Google Scholar] [CrossRef]
- Hou, D.; Wang, R.; Wang, Z.; Yang, G.; Xu, Z.; Zeng, Q.; Chen, Y. A light-activatable antibiotic with high activation efficiency and uncompromised bactericidal potency in the activated state. J. Leather Sci. Eng. 2021, 3, 8. [Google Scholar] [CrossRef]
- Shchelik, I.S.; Tomio, A.; Gademann, K. Design, Synthesis, and Biological Evaluation of Light-Activated Antibiotics. ACS Infect. Dis. 2021, 7, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Garcia, E.; Lozano, C.; Garcia-Iriepa, C.; Marazzi, M.; Winter, A.H.; Torres, C.; Sampedro, D. Controlling Antimicrobial Activity of Quinolones Using Visible/NIR Light-Activated BODIPY Photocages. Pharmaceutics 2022, 14, 1070. [Google Scholar] [CrossRef]
- Testolin, G.; Richter, J.; Ritter, A.; Prochnow, H.; Kohnke, J.; Bronstrup, M. Optical Modulation of Antibiotic Resistance by Photoswitchable Cystobactamids. Chemistry 2022, 28, e202201297. [Google Scholar] [CrossRef] [PubMed]
- Wegener, M.; Hansen, M.J.; Driessen, A.J.; Szymanski, W.; Feringa, B.L. Photocontrol of antibacterial activity: Shifting from UV to red light activation. J. Am. Chem. Soc. 2017, 139, 17979–17986. [Google Scholar] [CrossRef] [PubMed]
- Diguet, A.; Yanagisawa, M.; Liu, Y.J.; Brun, E.; Abadie, S.; Rudiuk, S.; Baigl, D. UV-induced bursting of cell-sized multicomponent lipid vesicles in a photosensitive surfactant solution. J. Am. Chem. Soc. 2012, 134, 4898–4904. [Google Scholar] [CrossRef] [PubMed]
- Szymanski, W.; Beierle, J.M.; Kistemaker, H.A.; Velema, W.A.; Feringa, B.L. Reversible photocontrol of biological systems by the incorporation of molecular photoswitches. Chem. Rev. 2013, 113, 6114–6178. [Google Scholar] [CrossRef] [Green Version]
- Mutter, N.L.; Volaric, J.; Szymanski, W.; Feringa, B.L.; Maglia, G. Reversible Photocontrolled Nanopore Assembly. J. Am. Chem. Soc. 2019, 141, 14356–14363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duchesne, I.; Galstian, T.; Sorelli, L.; Rodrigue, D.; Fabbri, F.; Fafard, M. Photodynamic control of bacterial motility by means of azobenzene molecules. J. Photochem. Photobiol. 2021, 8, 100074. [Google Scholar] [CrossRef]
- de Souza-Guerreiro, T.C.; Bondelli, G.; Grobas, I.; Donini, S.; Sesti, V.; Bertarelli, C.; Lanzani, G.; Asally, M.; Paterno, G.M. Membrane Targeted Azobenzene Drives Optical Modulation of Bacterial Membrane Potential. Adv. Sci. 2023, 10, e2205007. [Google Scholar] [CrossRef]
- Canaparo, R.; Foglietta, F.; Giuntini, F.; Della Pepa, C.; Dosio, F.; Serpe, L. Recent Developments in Antibacterial Therapy: Focus on Stimuli-Responsive Drug-Delivery Systems and Therapeutic Nanoparticles. Molecules 2019, 24, 1991. [Google Scholar] [CrossRef] [Green Version]
- Ferreres, G.; Ivanova, K.; Ivanov, I.; Tzanov, T. Nanomaterials and Coatings for Managing Antibiotic-Resistant Biofilms. Antibiotics 2023, 12, 310. [Google Scholar] [CrossRef]
- Tallet, L.; Gribova, V.; Ploux, L.; Vrana, N.E.; Lavalle, P. New Smart Antimicrobial Hydrogels, Nanomaterials, and Coatings: Earlier Action, More Specific, Better Dosing? Adv. Healthc. Mater. 2021, 10, e2001199. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shan, M.; Zhang, S.; Chen, X.; Liu, W.; Chen, J.; Liu, X. Stimuli-Responsive Antibacterial Materials: Molecular Structures, Design Principles, and Biomedical Applications. Adv. Sci. 2022, 9, e2104843. [Google Scholar] [CrossRef]
- Borzenkov, M.; Pallavicini, P.; Taglietti, A.; D’Alfonso, L.; Collini, M.; Chirico, G. Photothermally active nanoparticles as a promising tool for eliminating bacteria and biofilms. Beilstein J. Nanotechnol. 2020, 11, 1134–1146. [Google Scholar] [CrossRef]
- Zhao, Y.; Dai, X.; Wei, X.; Yu, Y.; Chen, X.; Zhang, X.; Li, C. Near-Infrared Light-Activated Thermosensitive Liposomes as Efficient Agents for Photothermal and Antibiotic Synergistic Therapy of Bacterial Biofilm. ACS Appl. Mater. Interfaces 2018, 10, 14426–14437. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Wang, J.; Wang, Y.; Chen, A.; Wang, C.; Mo, W.; Li, Y.; Yuan, Q.; Zhang, Y. Photon-Responsive Antibacterial Nanoplatform for Synergistic Photothermal-/Pharmaco-Therapy of Skin Infection. ACS Appl. Mater. Interfaces 2019, 11, 300–310. [Google Scholar] [CrossRef]
- Dong, X.; Overton, C.M.; Tang, Y.; Darby, J.P.; Sun, Y.P.; Yang, L. Visible Light-Activated Carbon Dots for Inhibiting Biofilm Formation and Inactivating Biofilm-Associated Bacterial Cells. Front. Bioeng. Biotechnol. 2021, 9, 786077. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhang, B.; Zhang, Y.; Su, R.; Li, P.; Su, W. Fluorescent Carbon Dot-Curcumin Nanocomposites for Remarkable Antibacterial Activity with Synergistic Photodynamic and Photothermal Abilities. ACS Appl. Bio. Mater. 2021, 4, 6703–6718. [Google Scholar] [CrossRef] [PubMed]
- Candreva, A.; De Rose, R.; Perrotta, I.D.; Guglielmelli, A.; La Deda, M. Light-induced clusterization of gold nanoparticles: A new photo-triggered antibacterial against E. coli proliferation. Nanomaterials 2023, 13, 746. [Google Scholar] [CrossRef]
- Wang, N.; Xu, H.; Sun, S.; Guo, P.; Wang, Y.; Qian, C.; Zhong, Y.; Yang, D. Wound therapy via a photo-responsively antibacterial nano-graphene quantum dots conjugate. J. Photochem. Photobiol. B 2020, 210, 111978. [Google Scholar] [CrossRef] [PubMed]
- García, A.; González, B.; Harvey, C.; Izquierdo-Barba, I.; Vallet-Regí, M. Effective reduction of biofilm through photothermal therapy by gold core@shell based mesoporous silica nanoparticles. Microporous Mesoporous Mater. 2021, 328, 111489. [Google Scholar] [CrossRef]
- Bagchi, D.; Rathnam, V.S.; Lemmens, P.; Banerjee, I.; Pal, S.K. NIR-light-active ZnO-based nanohybrids for bacterial biofilm treatment. ACS Omega 2018, 3, 10877–10885. [Google Scholar] [CrossRef]
- Qi, M.; Li, X.; Sun, X.; Li, C.; Tay, F.R.; Weir, M.D.; Dong, B.; Zhou, Y.; Wang, L.; Xu, H.H.K. Novel nanotechnology and near-infrared photodynamic therapy to kill periodontitis-related biofilm pathogens and protect the periodontium. Dent. Mater. 2019, 35, 1665–1681. [Google Scholar] [CrossRef]
- Zhou, K.; Qiu, X.; Xu, L.; Li, G.; Rao, B.; Guo, B.; Pei, D.; Li, A.; He, G. Poly(selenoviologen)-Assembled Upconversion Nanoparticles for Low-Power Single-NIR Light-Triggered Synergistic Photodynamic and Photothermal Antibacterial Therapy. ACS Appl. Mater. Interfaces 2020, 12, 26432–26443. [Google Scholar] [CrossRef]
- Wei, X.; Li, J.; Zhang, Y.; Zheng, Y.; Zhang, Y.; Meng, H.; Wu, G.; Hu, Y.; Gao, Y.; Huang, S.; et al. Synergy between Clinical Microenvironment Targeted Nanoplatform and Near-Infrared Light Irradiation for Managing Pseudomonas aeruginosa Infections. ACS Appl. Mater. Interfaces 2021, 13, 38979–38989. [Google Scholar] [CrossRef]
- Ran, X.; Du, Y.; Wang, Z.; Wang, H.; Pu, F.; Ren, J.; Qu, X. Hyaluronic Acid-Templated Ag Nanoparticles/Graphene Oxide Composites for Synergistic Therapy of Bacteria Infection. ACS Appl. Mater. Interfaces 2017, 9, 19717–19724. [Google Scholar] [CrossRef]
- Song, Z.; Wu, Y.; Cao, Q.; Wang, H.; Wang, X.; Han, H. pH-Responsive, Light-Triggered on-Demand Antibiotic Release from Functional Metal-Organic Framework for Bacterial Infection Combination Therapy. Adv. Funct. Mater. 2018, 28, 1800011. [Google Scholar] [CrossRef]
- Shen, Z.; He, K.; Ding, Z.; Zhang, M.; Yu, Y.; Hu, J. Visible-Light-Triggered Self-Reporting Release of Nitric Oxide (NO) for Bacterial Biofilm Dispersal. Macromolecules 2019, 52, 7668–7677. [Google Scholar] [CrossRef]
- Hong, L.; Liu, X.; Tan, L.; Cui, Z.; Yang, X.; Liang, Y.; Li, Z.; Zhu, S.; Zheng, Y.; Yeung, K.W.K. Rapid Biofilm Elimination on Bone Implants Using Near-Infrared-Activated Inorganic Semiconductor Heterostructures. Adv. Healthc. Mater. 2019, 8, 1900835. [Google Scholar] [CrossRef]
- Cui, Q.; Yuan, H.; Bao, X.; Ma, G.; Wu, M.; Xing, C. Synergistic Photodynamic and Photothermal Antibacterial Therapy Based on a Conjugated Polymer Nanoparticle-Doped Hydrogel. ACS Appl. Bio. Mater. 2020, 3, 4436–4443. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Tao, B.; He, Y.; Mu, C.; Liu, G.; Zhang, J.; Liao, Q.; Liu, P.; Cai, K. Remote eradication of biofilm on titanium implant via near-infrared light triggered photothermal/photodynamic therapy strategy. Biomaterials 2019, 223, 119479. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, X.; Yang, Y.; Zhang, H.; Shi, J.; Yao, X.; Zhang, X. Near-Infrared Light-Triggered Therapy to Combat Bacterial Biofilm Infections by MoSe2/TiO2 Nanorod Arrays on Bone Implants. Adv. Mater. Interfaces 2020, 7, 1901706. [Google Scholar] [CrossRef]
- Huang, H.-H.; Anand, A.; Lin, C.-J.; Lin, H.-J.; Lin, Y.-W.; Harroun, S.G.; Huang, C.-C. LED irradiation of halogen/nitrogen-doped polymeric graphene quantum dots triggers the photodynamic inactivation of bacteria in infected wounds. Carbon 2021, 174, 710–722. [Google Scholar] [CrossRef]
- Xi, J.; Zhang, J.; Qian, X.; An, L.; Fan, L. Using a visible light-triggered pH switch to activate nanozymes for antibacterial treatment. RSC Adv. 2020, 10, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Gan, H.Q.; Chen, G.R.; James, T.D.; Zhang, B.; Hu, Q.; Xu, F.; Hu, X.L.; He, X.P.; Mai, Y. Near-Infrared Light-Triggered Bacterial Eradication Using a Nanowire Nanocomposite of Graphene Nanoribbons and Chitosan-Coated Silver Nanoparticles. Front. Chem. 2021, 9, 767847. [Google Scholar] [CrossRef]
- Wang, K.; Zhan, S.; Sun, H.; Zhang, D.; Wang, J. Hollow porous core–shell ZnFe2O4/AgCl nanocubes coated with EDTA and Ag nanoparticles for enhanced photocatalytic performances of visible–light–driven. Chem. Eng. J. 2020, 400, 125908. [Google Scholar] [CrossRef]
- Yu, S.; Li, G.; Liu, R.; Ma, D.; Xue, W. Dendritic Fe3O4@Poly(dopamine)@PAMAM Nanocomposite as Controllable NO-Releasing Material: A Synergistic Photothermal and NO Antibacterial Study. Adv. Funct. Mater. 2018, 28, 1707440. [Google Scholar] [CrossRef]
- Choi, S.K. Mechanistic basis of light induced cytotoxicity of photoactive nanomaterials. NanoImpact 2016, 3–4, 81–89. [Google Scholar] [CrossRef]
- Sivakumar, P.; Lee, M.; Kim, Y.S.; Shim, M.S. Photo-triggered antibacterial and anticancer activities of zinc oxide nanoparticles. J. Mater. Chem. B 2018, 6, 4852–4871. [Google Scholar] [CrossRef] [PubMed]
- Borzenkov, M.; D’Alfonso, L.; Polissi, A.; Sperandeo, P.; Collini, M.; Dacarro, G.; Taglietti, A.; Chirico, G.; Pallavicini, P. Novel photo-thermally active polyvinyl alcohol-Prussian blue nanoparticles hydrogel films capable of eradicating bacteria and mitigating biofilms. Nanotechnology 2019, 30, 295702. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liu, J.; Xu, Y.; Liu, H.; Zhang, J.; Wang, Y.; Sun, Y.; Zhao, M.; Liao, L.; Wang, X. Fast Cross-Linked Hydrogel as a Green Light-Activated Photocatalyst for Localized Biofilm Disruption and Brush-Free Tooth Whitening. ACS Appl. Mater. Interfaces 2022, 14, 28427–28438. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, J.; Wang, H.; Li, S.; Pan, X.; Xu, B.; Yang, H.; Wu, Q.; Li, W.; Su, X.; et al. NIR Laser-Triggered Microneedle-Based Liquid Band-Aid for Wound Care. Adv. Funct. Mater. 2021, 31, 2100218. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, P.; Fan, Y.; Zhao, J.; Niu, S.; Song, L.; Ma, L.; Ren, L.; Ming, W. Near-infrared triggered antibacterial nanocomposite membrane containing upconversion nanoparticles. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 103, 109797. [Google Scholar] [CrossRef]
- Han, X.; Boix, G.; Balcerzak, M.; Moriones, O.H.; Cano-Sarabia, M.; Cortés, P.; Bastús, N.; Puntes, V.; Llagostera, M.; Imaz, I.; et al. Antibacterial Films Based on MOF Composites that Release Iodine Passively or Upon Triggering by Near-Infrared Light. Adv. Funct. Mater. 2022, 32, 2112902. [Google Scholar] [CrossRef]
- Chen, Q.; Li, S.; Zhao, W.; Zhao, C. A rapid-triggered approach towards antibacterial hydrogel wound dressing with synergic photothermal and sterilization profiles. Biomater. Adv. 2022, 138, 212873. [Google Scholar] [CrossRef]
- Contreras, A.; Raxworthy, M.J.; Wood, S.; Tronci, G. Hydrolytic Degradability, Cell Tolerance and On-Demand Antibacterial Effect of Electrospun Photodynamically Active Fibres. Pharmaceutics 2020, 12, 711. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Hu, J.; Wang, W.; Liu, K.; Zhou, C.; Liu, Z.; Kong, S.; Lin, S.; Deng, Y.; Guo, Z. Thermo and light-responsive strategies of smart titanium-containing composite material surface for enhancing bacterially anti-adhesive property. Chem. Eng. J. 2021, 407, 125783. [Google Scholar] [CrossRef]
- Mai, B.; Jia, M.; Liu, S.; Sheng, Z.; Li, M.; Gao, Y.; Wang, X.; Liu, Q.; Wang, P. Smart Hydrogel-Based DVDMS/bFGF Nanohybrids for Antibacterial Phototherapy with Multiple Damaging Sites and Accelerated Wound Healing. ACS Appl. Mater. Interfaces 2020, 12, 10156–10169. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Xiang, Y.; Liu, X.; Cui, Z.; Yang, X.; Yeung, K.W.K.; Pan, H.; Wang, X.; Chu, P.K.; Wu, S. Photo-Inspired Antibacterial Activity and Wound Healing Acceleration by Hydrogel Embedded with Ag/Ag@AgCl/ZnO Nanostructures. ACS Nano 2017, 11, 9010–9021. [Google Scholar] [CrossRef]
- Li, Z.; Feng, X.; Gao, S.; Jin, Y.; Zhao, W.; Liu, H.; Yang, X.; Hu, S.; Cheng, K.; Zhang, J. Porous Organic Polymer-Coated Band-Aids for Phototherapy of Bacteria-Induced Wound Infection. ACS Appl. Bio. Mater. 2019, 2, 613–618. [Google Scholar] [CrossRef]
- Munoz, M.; El-Khoury, A.; Eren Cimenci, C.; Gonzalez-Gomez, M.; Hunter, R.A.; Lomboni, D.; Variola, F.; Rotstein, B.H.; Vono, L.L.R.; Rossi, L.M.; et al. Riboflavin Surface Modification of Poly(vinyl chloride) for Light-Triggered Control of Bacterial Biofilm and Virus Inactivation. ACS Appl. Mater. Interfaces 2021, 13, 32251–32262. [Google Scholar] [CrossRef]
- Li, Y.; Liu, X.; Li, B.; Zheng, Y.; Han, Y.; Chen, D.F.; Yeung, K.W.K.; Cui, Z.; Liang, Y.; Li, Z.; et al. Near-Infrared Light Triggered Phototherapy and Immunotherapy for Elimination of Methicillin-Resistant Staphylococcus aureus Biofilm Infection on Bone Implant. ACS Nano 2020, 14, 8157–8170. [Google Scholar] [CrossRef]
- Feng, L.; Shi, W.; Chen, Q.; Cheng, H.; Bao, J.; Jiang, C.; Zhao, W.; Zhao, C. Smart Asymmetric Hydrogel with Integrated Multi-Functions of NIR-Triggered Tunable Adhesion, Self-Deformation, and Bacterial Eradication. Adv. Healthc. Mater. 2021, 10, e2100784. [Google Scholar] [CrossRef]
- Shi, Y.; Truong, V.X.; Kulkarni, K.; Qu, Y.; Simon, G.P.; Boyd, R.L.; Perlmutter, P.; Lithgow, T.; Forsythe, J.S. Light-triggered release of ciprofloxacin from an in situ forming click hydrogel for antibacterial wound dressings. J. Mater. Chem. B 2015, 3, 8771–8774. [Google Scholar] [CrossRef]
- Tao, B.; Lin, C.; Yuan, Z.; He, Y.; Chen, M.; Li, K.; Hu, J.; Yang, Y.; Xia, Z.; Cai, K. Near infrared light-triggered on-demand Cur release from Gel-PDA@ Cur composite hydrogel for antibacterial wound healing. Chem. Eng. J. 2021, 403, 126182. [Google Scholar] [CrossRef]
- Deng, H.; Sun, J.; Yu, Z.; Guo, Z.; Xu, C. Low-intensity near-infrared light-triggered spatiotemporal antibiotics release and hyperthermia by natural polysaccharide-based hybrid hydrogel for synergistic wound disinfection. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 118, 111530. [Google Scholar] [CrossRef] [PubMed]
- Hua, L.; Qian, H.; Lei, T.; Liu, W.; He, X.; Hu, Y.; Lei, P. Triggering Drug Release and Thermal-Disrupting Interface Induced Mitigation of Composite Photothermal Hydrogel Treating Infectious Wounds. Front. Bioeng. Biotechnol. 2021, 9, 796602. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Liu, W.; Wei, W.; Zhao, Y.; Zhuang, P.; Wang, X.; Wang, Y.; Hu, Y.; Dai, H. Photothermal Hydrogel Encapsulating Intelligently Bacteria-Capturing Bio-MOF for Infectious Wound Healing. ACS Nano 2022, 16, 19491–19508. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, G.; Chai, M.; Yao, X.; Chen, W.; Chu, P.K. Synergistic antibacterial activity of physical-chemical multi-mechanism by TiO(2) nanorod arrays for safe biofilm eradication on implant. Bioact. Mater. 2021, 6, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, Y.; Han, X.; Zhang, H.; Fan, A.; Yao, X.; Tang, B.; Zhang, X. Light-Triggered Antibacterial Hydrogels Containing Recombinant Growth Factor for Treatment of Bacterial Infections and Improved Wound Healing. ACS Biomater. Sci. Eng. 2021, 7, 1438–1449. [Google Scholar] [CrossRef]
- Xi, J.; Wu, Q.; Xu, Z.; Wang, Y.; Zhu, B.; Fan, L.; Gao, L. Aloe-Emodin/Carbon Nanoparticle Hybrid Gels with Light-Induced and Long-Term Antibacterial Activity. ACS Biomater. Sci. Eng. 2018, 4, 4391–4400. [Google Scholar] [CrossRef]
- Qiao, B.; Pang, Q.; Yuan, P.; Luo, Y.; Ma, L. Smart wound dressing for infection monitoring and NIR-triggered antibacterial treatment. Biomater. Sci. 2020, 8, 1649–1657. [Google Scholar] [CrossRef]
- Li, Y.; Wang, J.; Yang, Y.; Shi, J.; Zhang, H.; Yao, X.; Chen, W.; Zhang, X. A rose bengal/graphene oxide/PVA hybrid hydrogel with enhanced mechanical properties and light-triggered antibacterial activity for wound treatment. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 118, 111447. [Google Scholar] [CrossRef]
- Karcher, J.; Pianowski, Z.L. Photocontrol of Drug Release from Supramolecular Hydrogels with Green Light. Chemistry 2018, 24, 11605–11610. [Google Scholar] [CrossRef]
- Kim, S.H.; Kang, E.B.; Jeong, C.J.; Sharker, S.M.; In, I.; Park, S.Y. Light controllable surface coating for effective photothermal killing of bacteria. ACS Appl. Mater. Interfaces 2015, 7, 15600–15606. [Google Scholar] [CrossRef]
- Yu, N.; Peng, H.; Qiu, L.; Wang, R.; Jiang, C.; Cai, T.; Sun, Y.; Li, Y.; Xiong, H. New pectin-induced green fabrication of Ag@AgCl/ZnO nanocomposites for visible-light triggered antibacterial activity. Int. J. Biol. Macromol. 2019, 141, 207–217. [Google Scholar] [CrossRef]
- Hu, H.; Zhong, D.; Li, W.; Lin, X.; He, J.; Sun, Y.; Wu, Y.; Shi, M.; Chen, X.; Xu, F.; et al. Microalgae-based bioactive hydrogel loaded with quorum sensing inhibitor promotes infected wound healing. Nano Today 2022, 42, 101368. [Google Scholar] [CrossRef]
- Huang, T.; Zhou, Z.; Li, Q.; Tang, X.; Chen, X.; Ge, Y.; Ling, J. Light-triggered adhesive silk-based film for effective photodynamic antibacterial therapy and rapid hemostasis. Front. Bioeng. Biotechnol. 2022, 9, 1346. [Google Scholar] [CrossRef]
- Xiang, Y.; Mao, C.; Liu, X.; Cui, Z.; Jing, D.; Yang, X.; Liang, Y.; Li, Z.; Zhu, S.; Zheng, Y.; et al. Rapid and Superior Bacteria Killing of Carbon Quantum Dots/ZnO Decorated Injectable Folic Acid-Conjugated PDA Hydrogel through Dual-Light Triggered ROS and Membrane Permeability. Small 2019, 15, e1900322. [Google Scholar] [CrossRef]
- Carpenter, A.W.; Schoenfisch, M.H. Nitric oxide release: Part II. Therapeutic applications. Chem. Soc. Rev. 2012, 41, 3742–3752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Li, H.; Tao, X.; Xie, Y.; Yang, L.; Mao, Z.W.; Xia, W. Light-Triggered Nitric Oxide Release by a Photosensitizer to Combat Bacterial Biofilm Infections. Chemistry 2021, 27, 5453–5460. [Google Scholar] [CrossRef] [PubMed]
- Teirlinck, E.; Barras, A.; Liu, J.; Fraire, J.C.; Lajunen, T.; Xiong, R.; Forier, K.; Li, C.; Urtti, A.; Boukherroub, R. Exploring light-sensitive nanocarriers for simultaneous triggered antibiotic release and disruption of biofilms upon generation of laser-induced vapor nanobubbles. Pharmaceutics 2019, 11, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]



| Mechanism of Action and Target Mechanism | Approach/Molecule with Antibiofilm Activity | Resulting Phenotype | Target Bacteria | |
|---|---|---|---|---|
| Targeting adhesion strategy of bacteria | Genetic engineering | Deletion/mutation of the UpaB gene | Blocks the super adhesion protein (UpaB) to Inhibit biofilm formation | Escherichia coli (UTI infections) [108] |
| Enzyme activity inhibition | Myricetin (a flavonoid) | Inhibits Sortase A, an enzyme that catalyzes initial adhesion between Streptococcus mutans surface protein Pac and lectin, thus prohibiting surface adhesion | Streptococcus mutans [109] | |
| Deactivation of adhesive protein | D-arabinose | Prevents the adhesion of oral bacteria to dental implant surfaces through lectin | Streptococcus oralis, Fusobacterium nucleatum, and Porphyromonas gingivalis [110] | |
| Peptide mimic of bacterial α-helical peptide | Designed helical peptide G(IIKK)3I-NH2 (G3) | Prevents biofilm formation by acting on bacterial surface adhesion parameters; degrades eDNA to destabilize the mature biofilm architecture | Streptococcus mutans in tooth infections [111] | |
| Competition for surface binding sites | Nanoparticles coated with the outer membrane of Helicobacter pylori | Compete with bacteria for binding sites on host gastric epithelial cells and inhibit bacterial adhesion | Helicobacter pylori [112] | |
| Inhibiting Multivalent Adhesion Molecules (MAMs) | MAM7-mimicking inhibitor | Competes with the pathogen for sites on the host to initiate an infection | Mutidrug resistant and Pseudomonas aeruginosa in burn wounds [113] | |
| Surface antiadhesion | Medical devices with antiadhesive and antimicrobial properties | Grafting 2-methacryloyloxyethyl phosphate choline (MPC) onto medical devices | Prevents nonspecific adsorption of proteins and bacteria, resulting in biofilm inhibition | Gram-negative and Gram-positive bacteria [114] |
| Application of a surface coating called mPep (mixed-charge polypeptide) in medical catheters in vivo | Reduces bacterial adhesion | Methicillin-resistant Staphylococcus aureus and Pseudomonas aeruginosa [115] | ||
| Polydopamine-coated membranes with integerated copper nanoparticles (CuNPs) | Decrease Escherichia coli viability by 76% | Escherichia coli [116] | ||
| Modifying the polypropylene surface by reactive ion-etching technology | Reduce Escherichia coli adhesion on pp surfaces | Escherichia coli [117] | ||
| Targeting Biofilm formation strategy | Destruction of EPS | Deoxyribonuclease I (DNase I) | Degradation of eDNA, resulting in the prevention of adhesion | Staphylococcus aureus and Pseudomonas aeruginosa [118] |
| Alginate lyase | Alginate degradation results in the prevention of biofilm formation | Pseudomonas aeruginosa [119] | ||
| Proteases | Hydrolysis of matrix proteins and adhesins in EPS | Staphylococcus aureus [120] | ||
| Cellulase | Degrades EPS | Burkholderia cepacia [121] | ||
| α-Amylase | Mature biofilm degradation after EPS disruption | Staphylococcus aureus [122] | ||
| Inhibition of QS signal molecule production | FabI derivatives | Inhibition of enoyl-ACP reductase, which contributes acyl chain lengths of N-acyl homoserine lactones | Pseudomonas aeruginosa [123] | |
| (2-nitrophenyl) methanol derivatives | Inhibitors of PqsD, a key enzyme of signal molecule biosynthesis | Pseudomonas aeruginosa [124] | ||
| TNRHNPHHLHHV (peptide) | Inhibits LuxS enzyme activity to Inhibit AI-2 production | Streptococcus suis serotype 2 (SS2) [125] | ||
| Degradation of the QS Signaling molecule | AHL-lactonase AiiA | Degrades AHLs to prevent biofilm formation | Pseudomonas aeruginosa, Vibrio cholerae, and Enterobacter cloacae [126] | |
| Boronic acid derivate SM23 | Decreases the signaling molecules 3-oxo-C12-HSL and C4-HSL to reduce biofilm formation | Pseudomonas aeruginosa [127] | ||
| 3-(dibromomethylene) isobenzofuran-1(3H)-one derivatives | Inhibits biofilm formation through the inhibition of AI-2 activity | Fusobacterium nucleatum, Porphyromonas gingivalis, and Tannerella forsythia [128] | ||
| Diketopiperazines | Interferes with the activity of signal molecule synthase CepI, resulting in the prevention of biofilm formation | Burkholderia cenocepacia [129] | ||
| L-carvone | Inhibits QS activity by reducing AHL production (0.5 µL/mL) | Hafnia alvei [130] | ||
| Acylase | Cleaves the amide bond of AHLs | Gram-negative bacteria [131] | ||
| Imidazole | Degrades AI-2 | Escherichia coli [132] | ||
| Epigallocatechin gallate | Interferes with AHL production and AI-2-mediated QS | Staphylococcus aureus, Burkholderia cepacia, and Eikenella corrodens [133,134,135] | ||
| Acyl-HSL analog J8-C8 | Disturbs QS molecule C8-HSL synthesis and affects biofilm formation | Burkholderia glumae [136] | ||
| Targeting QS Signaling Receptors | 3-Phenyllactic acid (PLA) | Binds with QS receptors RhlR and PqsR involved in biofilm formation | Pseudomonas aeruginosa [137] | |
| Furanones and synthetic furanones | Competes with the native autoinducers to bind to the AHL receptors, decreasing virulence factor production and biofilm formation | Pseudomonas aeruginosa [138,139] | ||
| Sesquiterpene lactone | Decreases the affinity of the CviR protein to its receptor, LuxR | Chromobacterium violaceum [140] | ||
| Naringenin, Taxifolin, and Quercetin 4′-O-β-Dglucopyranoside | Inhibit QS-regulated gene expression. Reduce QS via the vfr-mediated lasIR system | Pseudomonas aeruginosa [141] | ||
| N-(3-oxododecanoyl) homoserine lactone derivatives | Block the binding site of the QS molecule, inhibiting biofilm formation and increasing antibiotic sensitivity | Pseudomonas aeruginosa clinical strains [142] | ||
| Flavonoids compounds | Reduce QS signal concentration | Yersinia enterocolitica [143] | ||
| A small peptide 5906 | Prevents homodimer formation, inhibiting LuxS activity | Edwardsiella tarda [144] | ||
| D-galactose | Inhibit AI-2 activity | Periodontopathogens [145] | ||
| N-phenyl-4-(3-phenylthioureido) benzenesulfonamide | Allosterically modifies the AI-3 receptor QseC, impedes virulence expression, and promotes biofilm formation | Escherichia coli (EAEC) O104:H4 [146] | ||
| Blocking the QS Signaling Cascade | Savirin | Targets AgrA to disrupt Agr operon-mediated QS | Staphylococcus aureus [73] | |
| Curcumin | Inhibits QS-controlled protease and biofilm formation | Pseudomonas aeruginosa PAO1 [147] | ||
| Efflux pump inhibitor PAβN | Reduces the extracellular accumulation of QS signals and diminishes the relative expression of the QS cascade (pqsA, pqsR, lasI, lasR, rhlI, and rhlR) | Pseudomonas aeruginosa clinical isolate [148] | ||
| Targeting mature biofilm | Killing persister and dormant cells | Adenosine (ADO) | Activates ATP and GTP synthesis and promotes cell respiration, thereby enhancing the killing of persistent cells by antibiotics | Gram-negative and Gram-positive bacteria [149] |
| Dialylquinoline TMC207 | Targets ATP synthase, thereby damaging the lipopeptide of the bacterial membrane, including persistent cells, and effectively improving the antibiotic treatment success rate | Mycobacterium tuberculosis [150] | ||
| Lead compound X9 | Inhibition of the RelMtb enzyme that is used to enter the persister cell stage. Inhibition of this enzyme kills the persister cells directly | Mycobacterium tuberculosis [151] | ||
| Materials | Formulation | Light | Mechanism | Target Microorganism | Application |
|---|---|---|---|---|---|
| ZnO-based and squaraine nanohybrids | Nanoparticles | NIR | NP internalization, photoinduced interfacial electron transfer, and ROS generation to disrupt bacterial biofilm | Staphylococcus aureus [240] | artificial implants |
| Au core in shell-based mesoporous silica nanoparticles | Nanoparticles | NIR | Incorporation of nitrosothiol groups (-SNO) with a heat-liable linker, NO release upon photothermal stimulation, and antimicrobial Levofloxacin to disrupt bacterial biofilm | Staphylococcus aureus [239] | |
| PEG-b-pLAMA/pAAPBA-b-pDPA NPs loaded with indocyanine green | Nanoparticles | NIR | Acidic infection environment, borate ester linkage cleavage, NP shelling, hyperpyrexia, and ROS generation | Pseudomonas aeruginosa [243] | clinical chronic infections in pulmonary alveoli |
| Distearoyl phosphatidylcholine + betainylate cholesterol micelles loaded with Cypate and Tobramycin | Liposomes | NIR | Photothermal and antibiotic synergy, penetration into biofilm channels, and thermal-triggered drug release | Pseudomonas aeruginosa [233] | |
| Core-shell upconversion nanoparticles and TiO2(UCNPs@TiO2) | Nanoparticles | NIR and UV | Under NIR light, the core UCNPs emit UV light, which triggers the photodynamic function of the shell via energy transfer | Streptococcus sanguinis, Porphyromonas gingivalis, and Fusobacterium nucleatum [241] | periodontitis |
| Graphene oxide—Hyaluronic acid—Ag NPs | Nanoparticles | NIR | GO absorbs light and generates heat; hyaluronidase-triggered photothermal platform and ROS generation | Staphylococcus aureus [244] | |
| 2-nitrobenzaldehyde-modified zeolitic imidazolate mesoporous NPs + rifampicin | Nanoparticles | UV | UV-light (365 nm)-responsive in situ production of acid, pH-dependent degradation of the zeolite framework, antibiotic release | Staphylococcus aureus and Methicillin-resistant Staphylococcus aureus [245] | wound healing |
| Nitric oxide-releasing micelles, PEO-b-polyCouNO + Ciprofloxacin | Nanomicelles | Visible | visible light irradiation, fluorescence turn-on, enabling in situ self-reporting NO release fluorescence turn-on (>185-fold) | Pseudomonas aeruginosa [246] | |
| Ag3PO4 NPs loaded on Bi2S3 nanorods | Nanorods | NIR | ROS generation after enhanced photocatalytic effect via photoactive nanoparticles and semiconductor heterojunction coating | Staphylococcus aureus and Escherichia coli [247] | orthopedic implants |
| Ag-Cu2O/PANI | Nanoparticles | Visible | Intracellular ROS generation is enhanced by photocatalytic particles and conductive polymer | Staphylococcus aureus and Pseudomonas aeruginosa [248] | |
| Black phosphorus quantum dots + vancomycin in a liposome | Liposomes | NIR | Hyperthermia, light-trigger liposome disruption, and release of its antibiotic cargo | Methicillin-resistant Staphylococcus aureus [234] | skin infections |
| Mesoporous polydopamine NPs with the photosensitizer Indocyanine Green | Nanoparticles | NIR | Photothermal/photodynamic therapy | Staphylococcus aureus [249] | bone implants |
| MoSe2/TiO2 Nanorod | Nanorods | NIR | Photothermal/photodynamic therapy | Staphylococcus aureus and Escherichia coli [250] | bone implants |
| 2,2′-(ethylenedioxy)bis (ethylamine) functionalized Carbon Nanodots | Nanodots | Visible | Breakage of the EPS matrix | Bacillus subtilis [235] | |
| Halogen/nitrogen-doped polymeric graphene quantum dots | Quantum dots | LED | ROS generation | Staphylococcus epidermidis, Staphylococcus aureus, and Pseudomonas aeruginosa [251] | wound healing |
| CuS nanoparticles + photoacid generator | Nanoparticles | Visible | Intramolecular photoreaction, pH decrease, and peroxidase activity | Staphylococcus aureus and Escherichia coli [252] | |
| Chitosan-coated silver NPs and graphene nanoribbon nanowires | Nanoparticles | NIR | Photothermal therapy | Pseudomonas aeruginosa [253] | medical patches |
| Graphene quantum dots + erytromycin + mesoporous silica NPs | Nanoparticles | LED | Singlet oxygen production, photodynamic therapy, and drug release | Staphylococcus aureus and Escherichia coli [238] | wound healing |
| Core-shell ZnFe2O4/AgCl@EDTA-Ag composites | Nanoparticles | Visible | Photocatalytic nanostructures and ROS generation | Escherichia coli [254] | medical disinfection |
| Poly(selenoviologen)-Assembled Upconversion Nanoparticles | Nanoparticles | NIR | Photothermal and photodynamic therapy and ROS generation | Methicillin-resistant Staphylococcus aureus [242] | infected wounds |
| Carbon nanodots + curcumin | Nanodots | Visible and NIR | Photothermal and photodynamic therapy and ROS generation | Staphylococcus aureus and Escherichia coli [236] | |
| Au plasmonic NPs | Nanoparticles | Visible | Nanostructure clusterization and photothermal effect | Escherichia coli [237] | |
| Polydopamine-Fe2O3 NPs with NONOates and dendritic poly(amidoamine) | Nanoparticles | NIR | Photothermal therapy and controlled NO release | Staphylococcus aureus and Escherichia coli [255] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kauser, A.; Parisini, E.; Suarato, G.; Castagna, R. Light-Based Anti-Biofilm and Antibacterial Strategies. Pharmaceutics 2023, 15, 2106. https://doi.org/10.3390/pharmaceutics15082106
Kauser A, Parisini E, Suarato G, Castagna R. Light-Based Anti-Biofilm and Antibacterial Strategies. Pharmaceutics. 2023; 15(8):2106. https://doi.org/10.3390/pharmaceutics15082106
Chicago/Turabian StyleKauser, Ambreen, Emilio Parisini, Giulia Suarato, and Rossella Castagna. 2023. "Light-Based Anti-Biofilm and Antibacterial Strategies" Pharmaceutics 15, no. 8: 2106. https://doi.org/10.3390/pharmaceutics15082106