
Intranasal Nanotransferosomal Gel for Quercetin Brain Targeting: II. Antidepressant Effect in an Experimental Animal Model

 , , , , , ,
, , , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
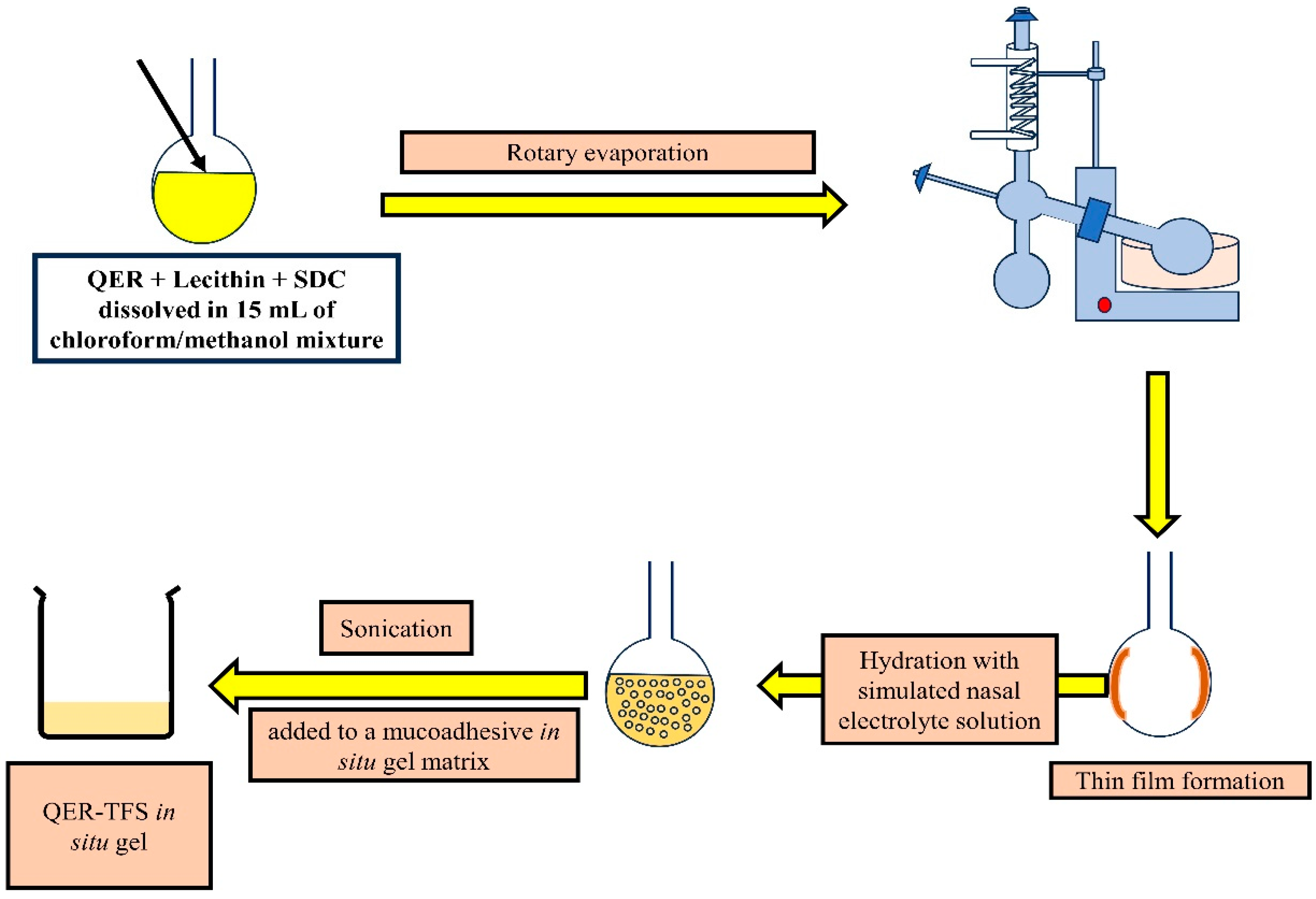
2.2.1. Preparation of Quercetin-Loaded Transferosomes (QER-TFS) and Quercetin Solution (QER-SOLN)
2.2.2. In Vivo Antidepressant Activity of QER-TFS Gel
Animals
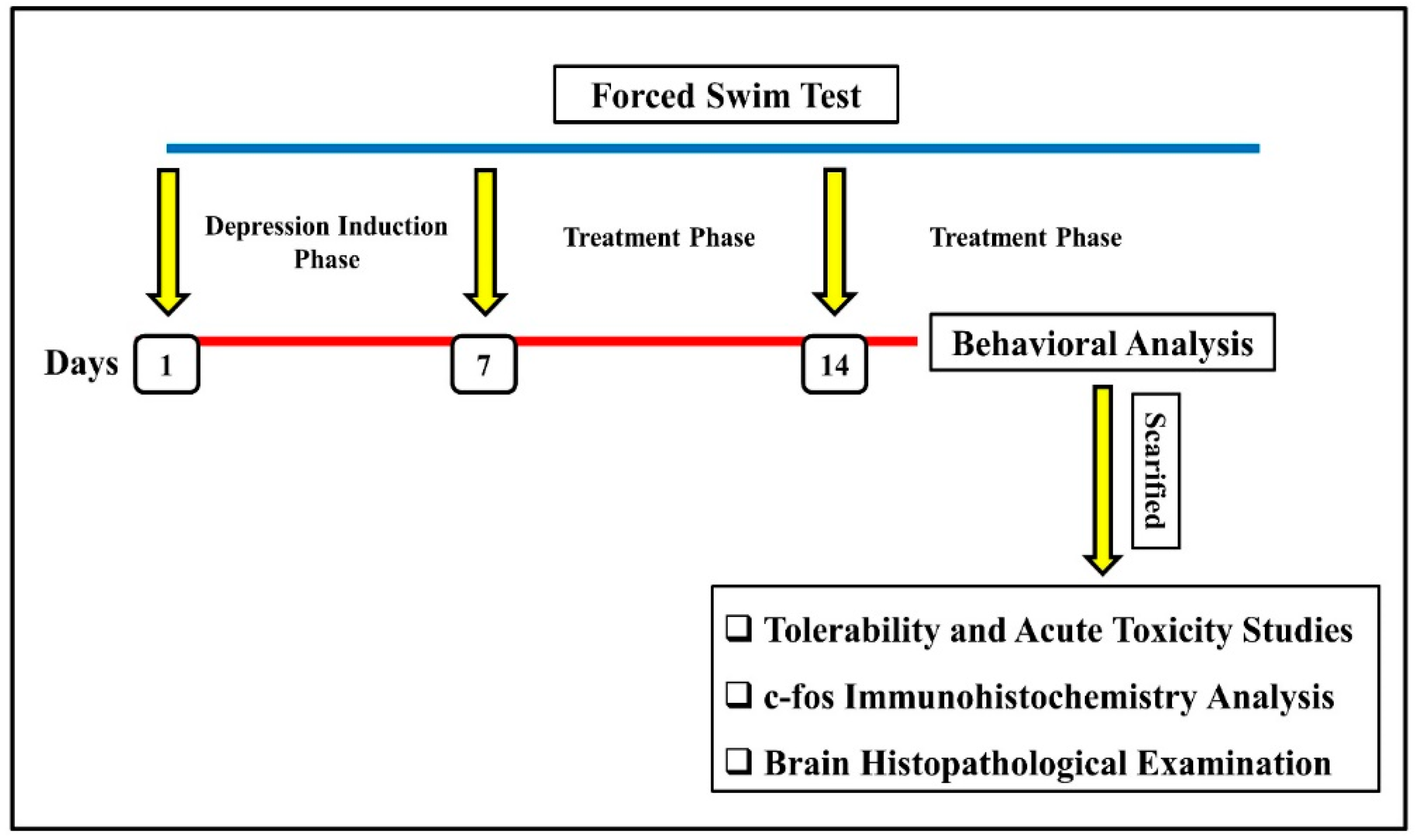
Induction of Depression and Experimental Protocol
Behavioral Analysis
- Immobility, Swimming, and Climbing in the FST
- 2.
- Tail Suspension Test (TST)
- 3.
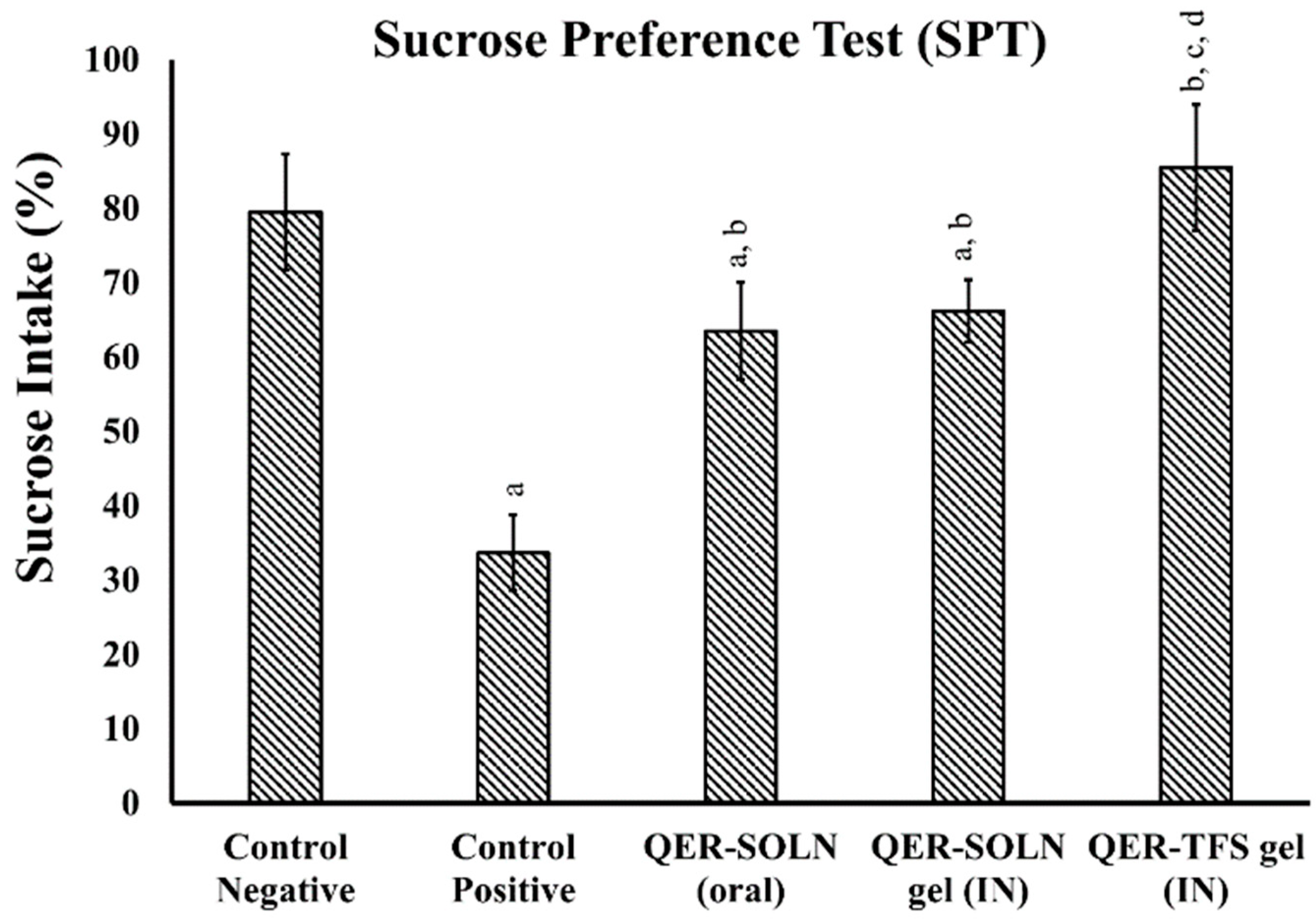
- Sucrose Preference Test (SPT)
- 4.
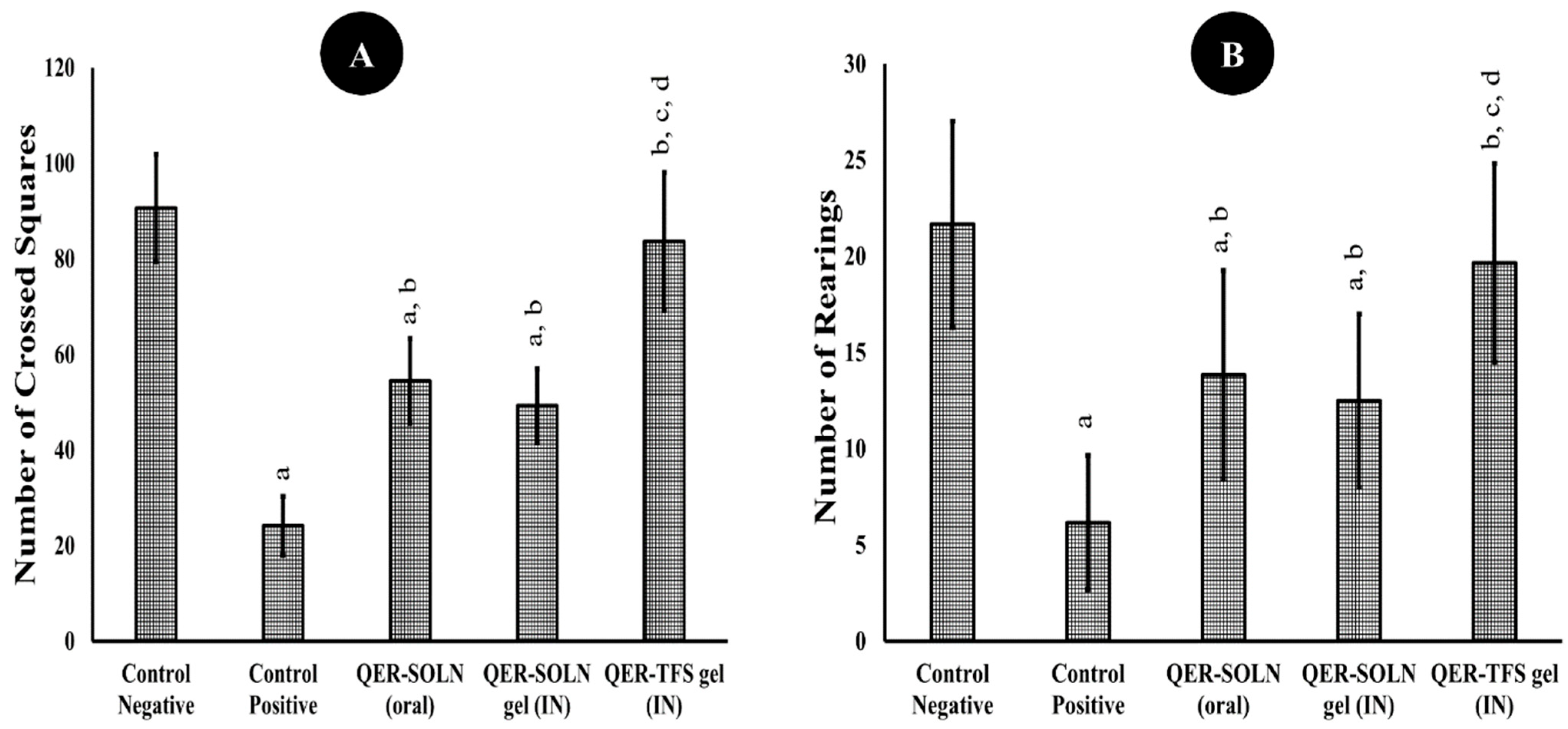
- Open Field Test (OFT)
- 5.
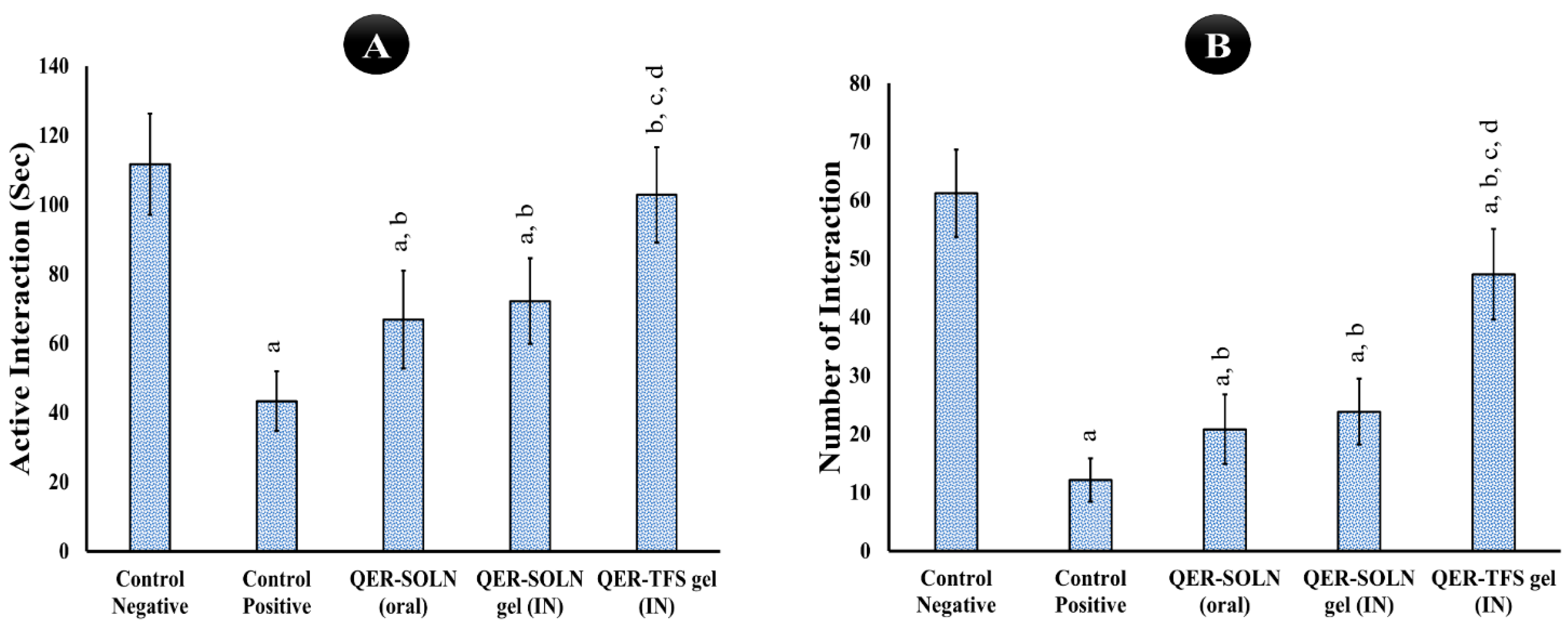
- Social Interaction Test (SIT)
- 6.
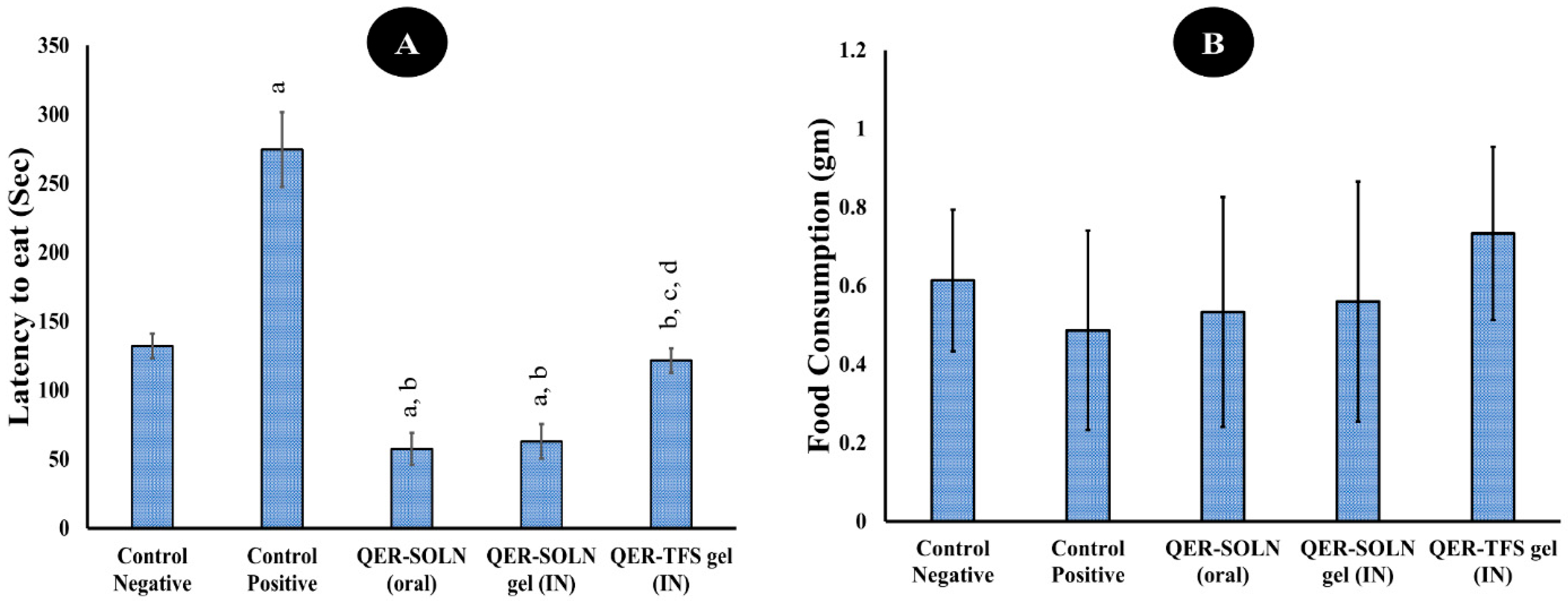
- Novelty-Suppressed Feeding (NSF)
c-fos Immunohistochemistry Analysis
Tolerability and Acute Toxicity Studies
Brain Histopathological Examination
2.3. Statistical Analysis
3. Results and Discussion
3.1. Pharmacodynamic Studies
3.1.1. Behavioral Analysis
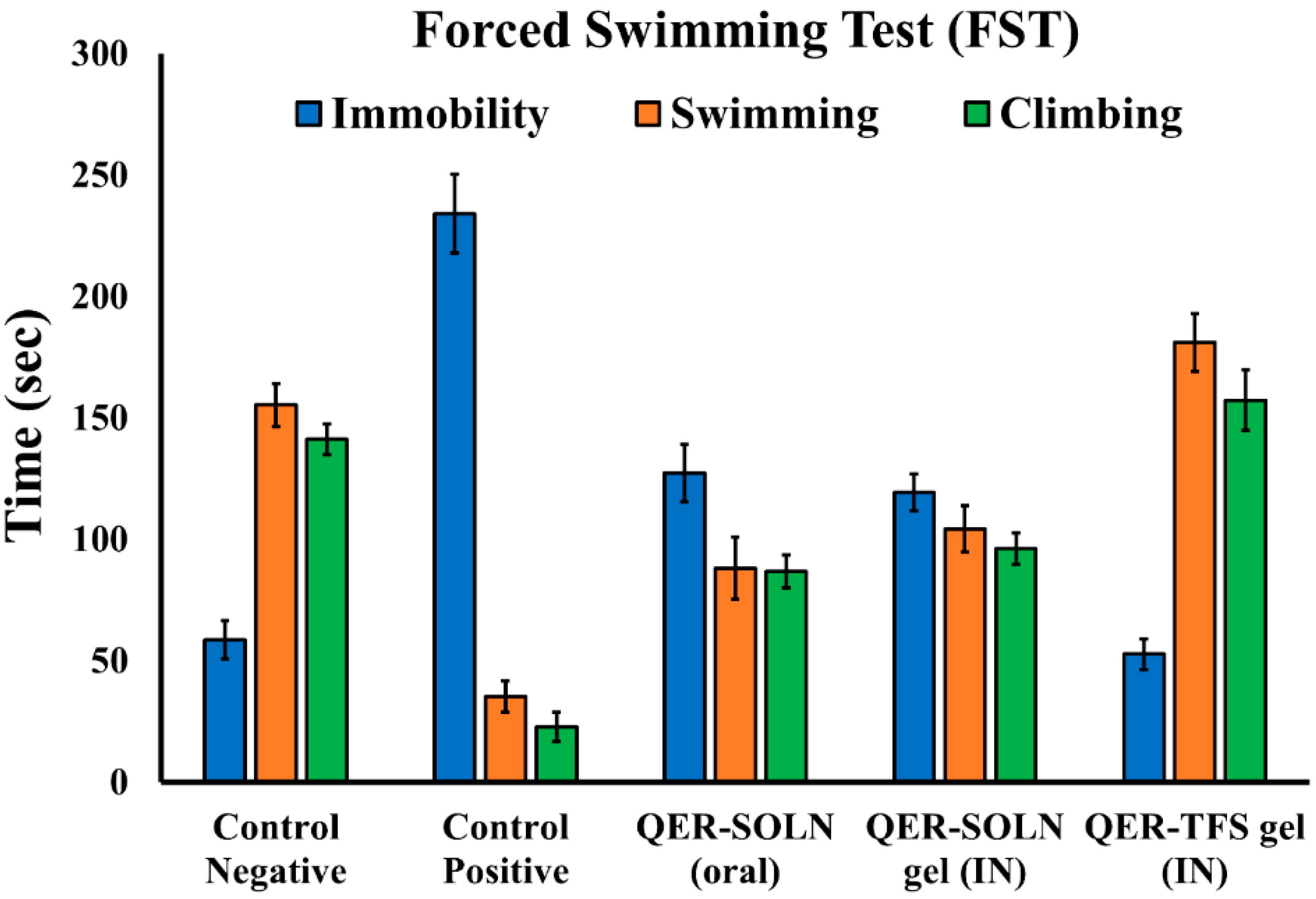
Immobility, Climbing, and Swimming in the FST
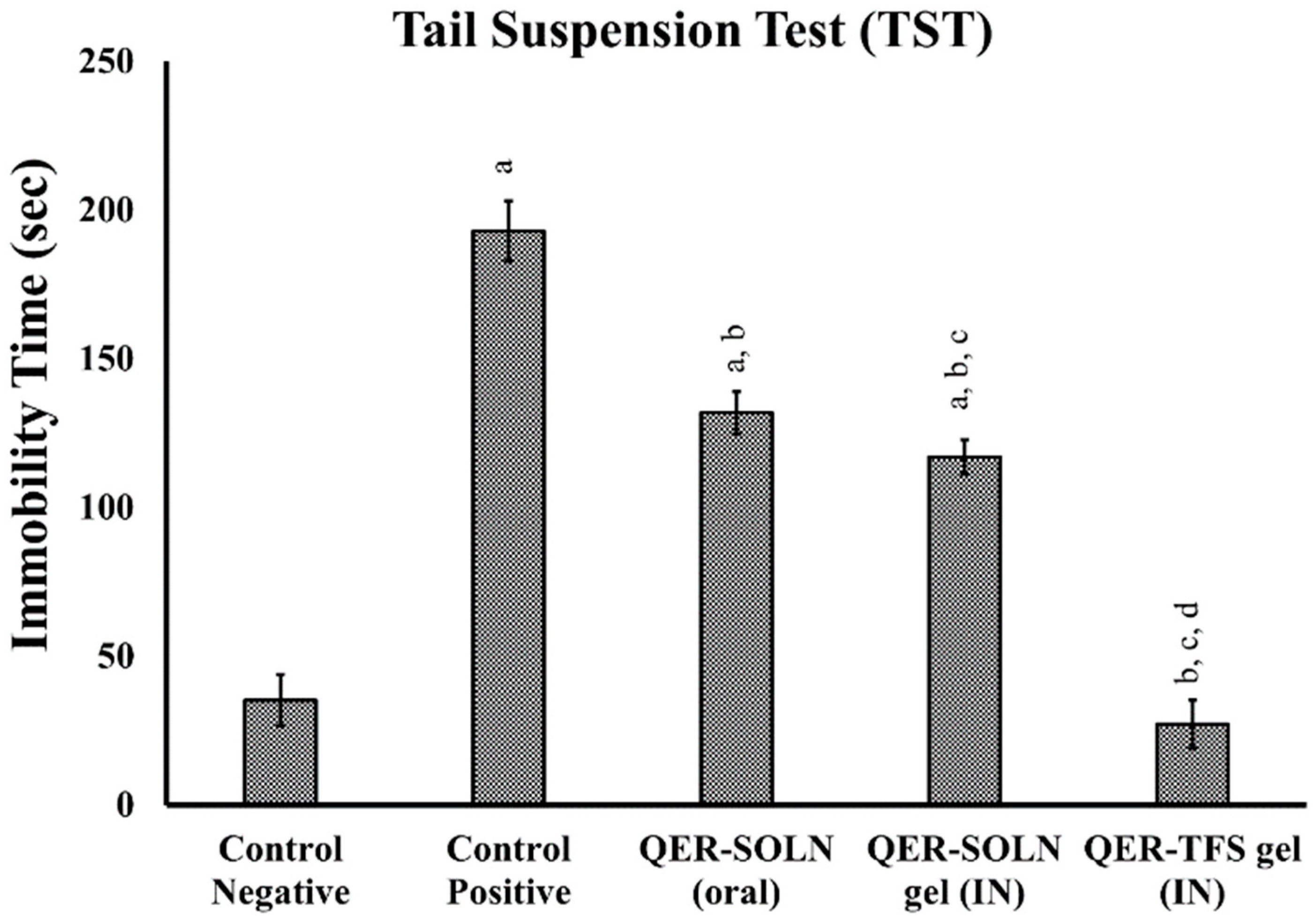
Tail Suspension Test (TST)
Sucrose Preference Test (SPT)
Open Field Test (OFT)
Social Interaction Test (SIT)
Novelty-Suppressed Feeding (NSF)
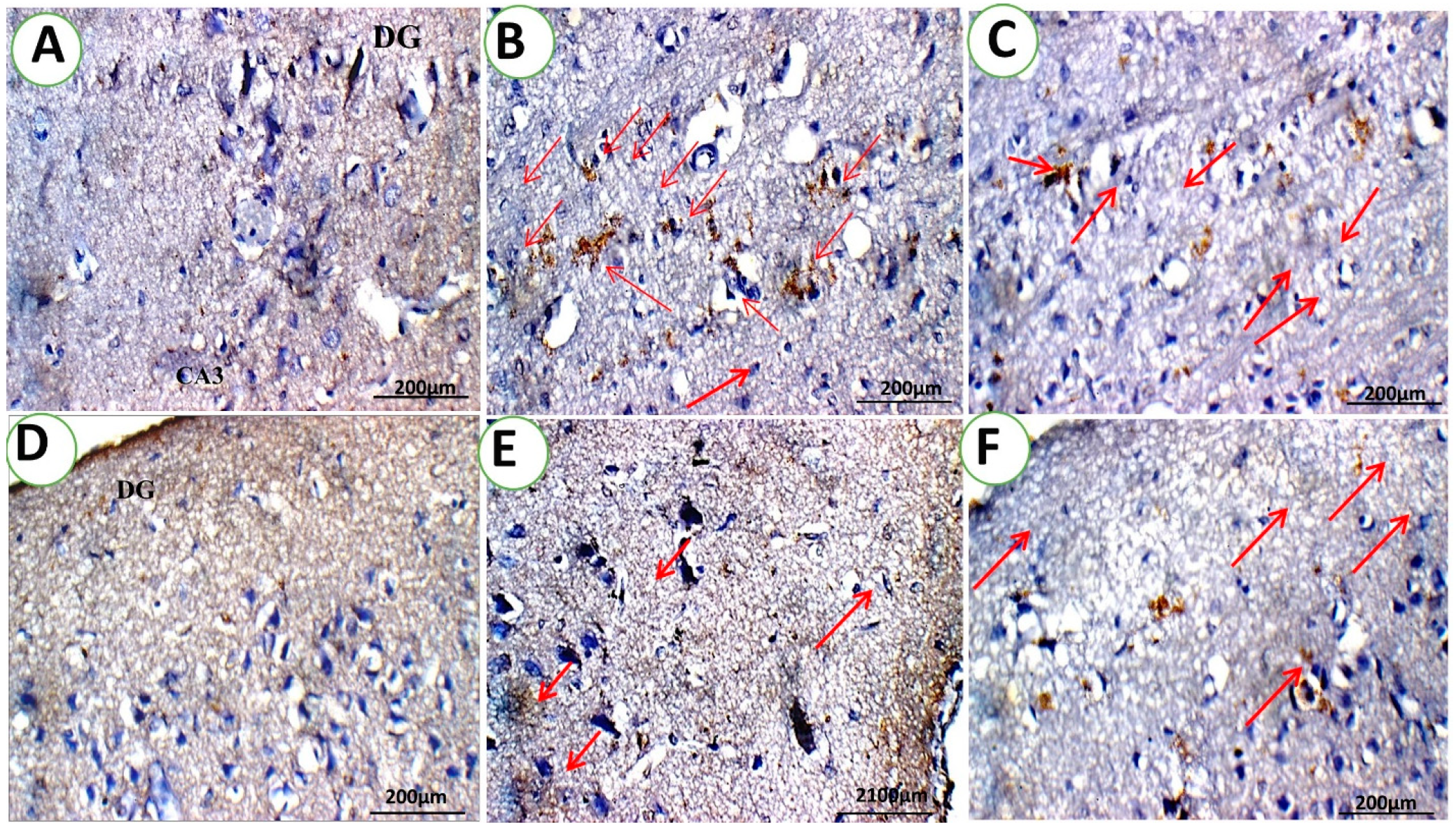
3.1.2. c-fos Immunohistochemistry Analysis
3.1.3. Tolerability and Acute Toxicity Studies
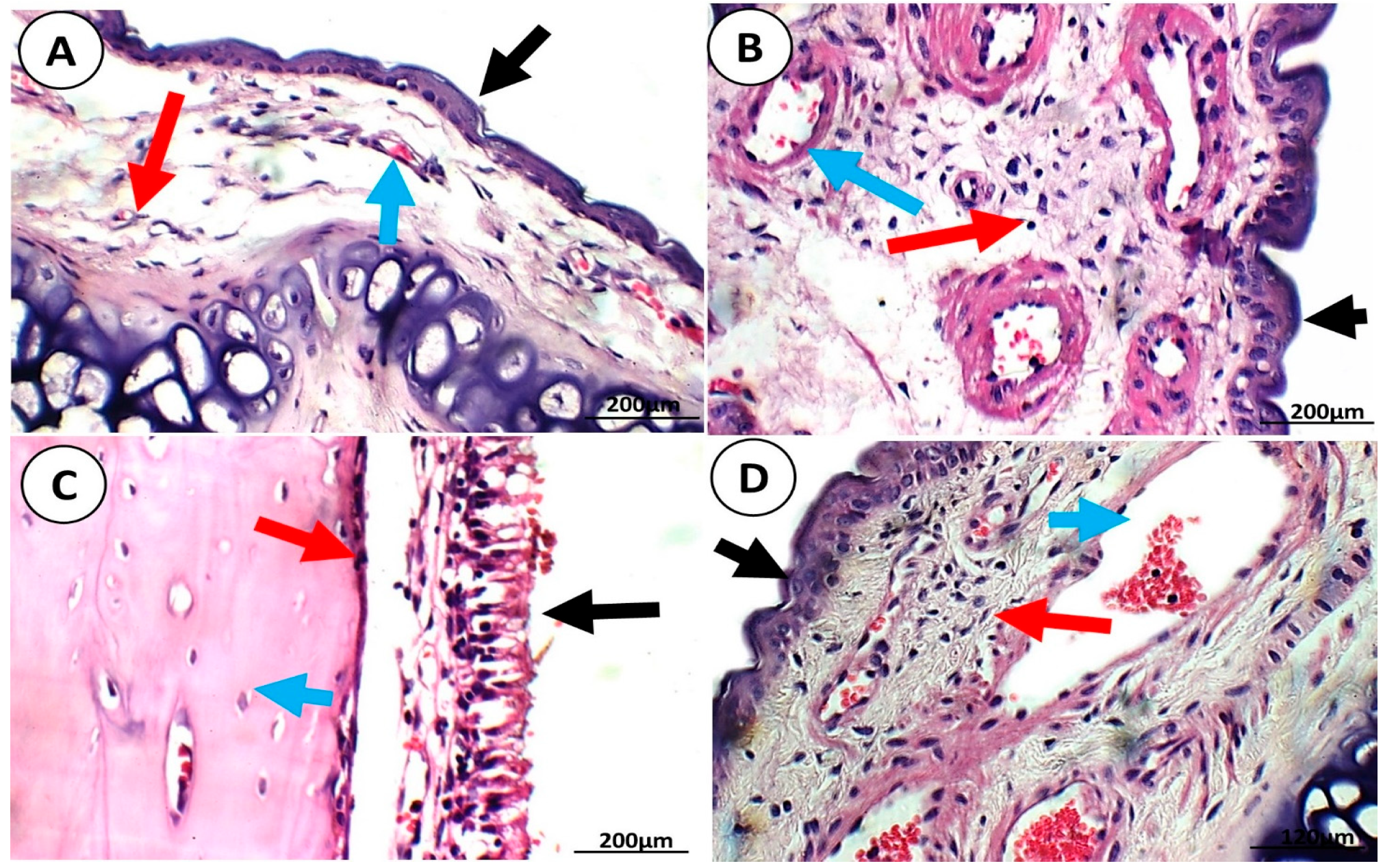
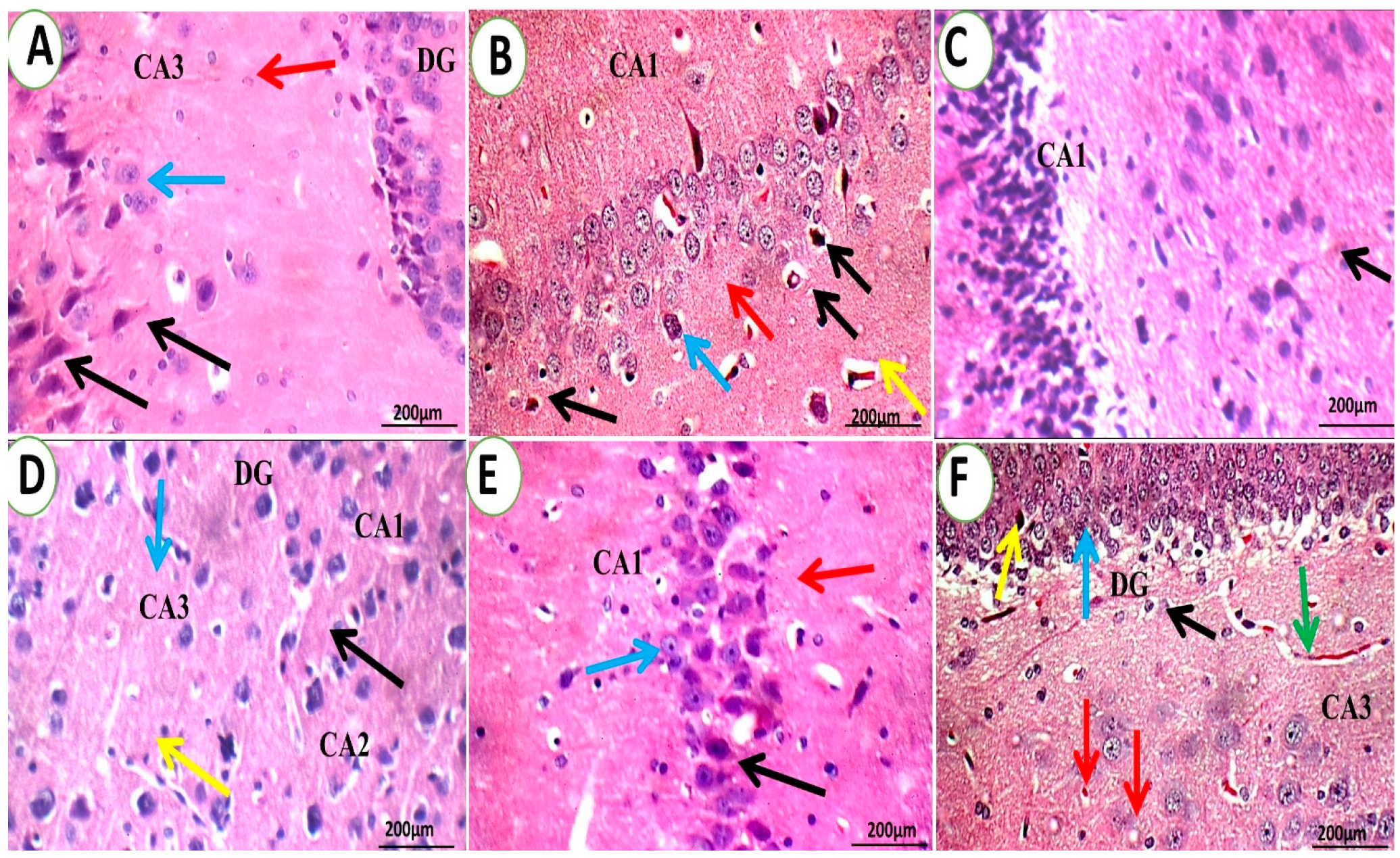
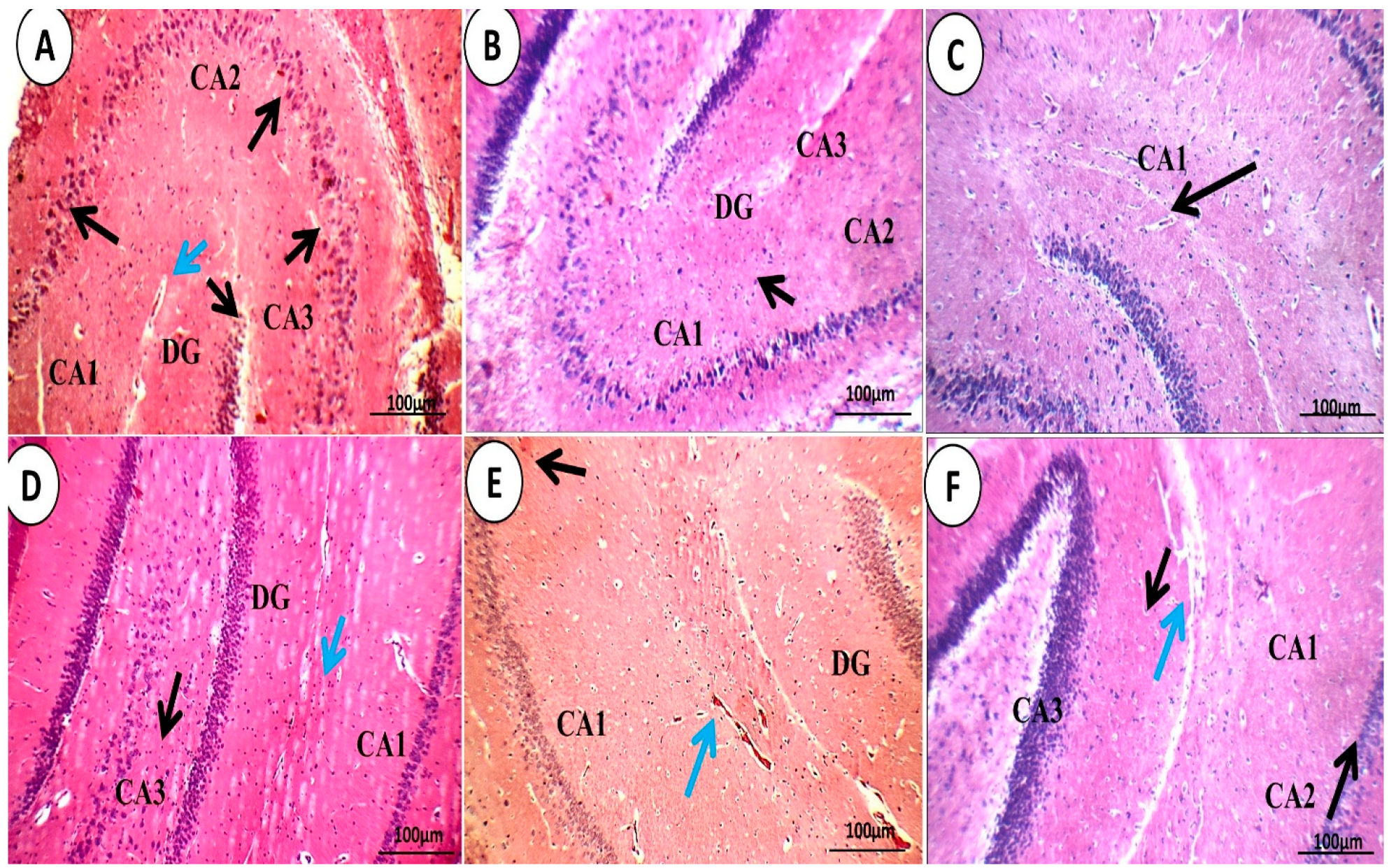
3.1.4. Brain Histopathological Examination
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Silvestro, S.; Bramanti, P.; Mazzon, E. Role of quercetin in depressive-like behaviors: Findings from animal models. Appl. Sci. 2021, 11, 7116. [Google Scholar] [CrossRef]
- Zhang, J.; Ning, L.; Wang, J. Dietary quercetin attenuates depressive-like behaviors by inhibiting astrocyte reactivation in response to stress. Biochem. Biophys. Res. Commun. 2020, 533, 1338–1346. [Google Scholar] [CrossRef]
- Kendler, K.S.; Thornton, L.M.; Gardner, C.O. Stressful life events and previous episodes in the etiology of major depression in women: An evaluation of the “kindling” hypothesis. Am. J. Psychiatry 2000, 157, 1243–1251. [Google Scholar] [CrossRef]
- Monroe, S.M.; Hadjiyannakis, K. The social environment and depression: Focusing on severe life stress. In Handbook of Depression; Guilford Press: New York, NY, USA, 2002. [Google Scholar]
- Frodl, T.; Reinhold, E.; Koutsouleris, N.; Reiser, M.; Meisenzahl, E.M. Interaction of childhood stress with hippocampus and prefrontal cortex volume reduction in major depression. J. Psychiatr. Res. 2010, 44, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Desmyter, S.; Van Heeringen, C.; Audenaert, K. Structural and functional neuroimaging studies of the suicidal brain. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 796–808. [Google Scholar] [CrossRef] [PubMed]
- Kendler, K.S.; Karkowski, L.M.; Prescott, C.A. Causal relationship between stressful life events and the onset of major depression. Am. J. Psychiatry 1999, 156, 837–841. [Google Scholar] [CrossRef]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (neuro) degenerative processes in that illness. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 676–692. [Google Scholar]
- Racagni, G.; Popoli, M. The pharmacological properties of antidepressants. Int. Clin. Psychopharmacol. 2010, 25, 117–131. [Google Scholar] [CrossRef]
- Dantzer, R.; O’connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Raison, C.L.; Capuron, L.; Miller, A.H. Cytokines sing the blues: Inflammation and the pathogenesis of depression. Trends Immunol. 2006, 27, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Moret, C.; Isaac, M.; Briley, M. Problems associated with long-term treatment with selective serotonin reuptake inhibitors. J. Psychopharmacol. 2009, 23, 967–974. [Google Scholar] [CrossRef]
- Kilts, C.D. Potential new drug delivery systems for antidepressants: An overview. J. Clin. Psychiatry 2003, 64, 31–33. [Google Scholar]
- Falzon, C.C.; Balabanova, A. Phytotherapy: An introduction to herbal medicine. Prim. Care Clin. Off. Pract. 2017, 44, 217–227. [Google Scholar] [CrossRef]
- Bakre, A.; Olayemi, J.; Ojo, O.; Odusanya, S.; Agu, G.; Aderibigbe, A. Antidepressant-like effect of ethanol extract of Blighia unijugata Bak.(Sapindaceae) leaves in acute and chronic models of depression in mice. Niger. J. Physiol. Sci. 2019, 34, 191–199. [Google Scholar]
- Chávez-Morales, Y.; Jiménez-Ferrer, E.; Martínez-Hernández, G.B.; Tortoriello, J.; Román-Ramos, R.; Zamilpa, A.; Herrera-Ruiz, M. Effect of standardized fractions and tiliroside from leaves of Tilia americana on depression tests in mice. Iran. J. Pharm. Res. IJPR 2019, 18, 1931. [Google Scholar] [PubMed]
- Dovichi, S.S.; Lajolo, F.M. Flavonoids and their relationship to diseases of the central nervous system. Nutr. Rev. Soc. Bras. Aliment. Nutr. 2011, 36, 123–135. [Google Scholar]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, P.; Serrano-García, N.; Medina-Campos, O.N.; Pedraza-Chaverri, J.; Ögren, S.O.; Rojas, C. Antidepressant-like effect of a Ginkgo biloba extract (EGb761) in the mouse forced swimming test: Role of oxidative stress. Neurochem. Int. 2011, 59, 628–636. [Google Scholar] [CrossRef]
- Sakakibara, H.; Yoshino, S.; Kawai, Y.; Terao, J. Antidepressant-like effect of onion (Allium cepa L.) powder in a rat behavioral model of depression. Biosci. Biotechnol. Biochem. 2008, 72, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Bhutada, P.; Mundhada, Y.; Bansod, K.; Ubgade, A.; Quazi, M.; Umathe, S.; Mundhada, D. Reversal by quercetin of corticotrophin releasing factor induced anxiety-and depression-like effect in mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2010, 34, 955–960. [Google Scholar] [CrossRef]
- Kawabata, K.; Kawai, Y.; Terao, J. Suppressive effect of quercetin on acute stress-induced hypothalamic-pituitary-adrenal axis response in Wistar rats. J. Nutr. Biochem. 2010, 21, 374–380. [Google Scholar] [CrossRef]
- Dok-Go, H.; Lee, K.H.; Kim, H.J.; Lee, E.H.; Lee, J.; Song, Y.S.; Lee, Y.-H.; Jin, C.; Lee, Y.S.; Cho, J. Neuroprotective effects of antioxidative flavonoids, quercetin,(+)-dihydroquercetin and quercetin 3-methyl ether, isolated from Opuntia ficus-indica var. saboten. Brain Res. 2003, 965, 130–136. [Google Scholar] [CrossRef]
- Haleagrahara, N.; Radhakrishnan, A.; Lee, N.; Kumar, P. Flavonoid quercetin protects against swimming stress-induced changes in oxidative biomarkers in the hypothalamus of rats. Eur. J. Pharmacol. 2009, 621, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Bhalani, D.V.; Nutan, B.; Kumar, A.; Singh Chandel, A.K. Bioavailability enhancement techniques for poorly aqueous soluble drugs and therapeutics. Biomedicines 2022, 10, 2055. [Google Scholar] [CrossRef]
- Eissa, E.M.; Elkomy, M.H.; Eid, H.M.; Ali, A.A.; Abourehab, M.A.; Alsubaiyel, A.M.; Naguib, I.A.; Alsalahat, I.; Hassan, A.H. Intranasal delivery of granisetron to the brain via nanostructured cubosomes-based in situ gel for improved management of chemotherapy-induced emesis. Pharmaceutics 2022, 14, 1374. [Google Scholar] [CrossRef]
- Eid, H.M.; Elkomy, M.H.; El Menshawe, S.F.; Salem, H.F. Transfersomal nanovesicles for nose-to-brain delivery of ofloxacin for better management of bacterial meningitis: Formulation, optimization by Box-Behnken design, characterization and in vivo pharmacokinetic study. J. Drug Deliv. Sci. Technol. 2019, 54, 101304. [Google Scholar] [CrossRef]
- Kumar, M.; Misra, A.; Babbar, A.; Mishra, A.; Mishra, P.; Pathak, K. Intranasal nanoemulsion based brain targeting drug delivery system of risperidone. Int. J. Pharm. 2008, 358, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-Z.; Zhang, Y.-Q.; Wang, Z.-Z.; Wu, K.; Lou, J.-N.; Qi, X.-R. Enhanced brain distribution and pharmacodynamics of rivastigmine by liposomes following intranasal administration. Int. J. Pharm. 2013, 452, 344–354. [Google Scholar] [CrossRef]
- Patel, R.J.; Parikh, R.H. Intranasal delivery of topiramate nanoemulsion: Pharmacodynamic, pharmacokinetic and brain uptake studies. Int. J. Pharm. 2020, 585, 119486. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.I.; Baboota, S.; Ahuja, A.; Ali, M.; Ali, J.; Sahni, J.K.; Bhatnagar, A. Pharmacoscintigraphic evaluation of potential of lipid nanocarriers for nose-to-brain delivery of antidepressant drug. Int. J. Pharm. 2014, 470, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Elkomy, M.H.; Zaki, R.M.; Alsaidan, O.A.; Elmowafy, M.; Zafar, A.; Shalaby, K.; Abdelgawad, M.A.; Abo El-Ela, F.I.; Rateb, M.E.; Naguib, I.A. Intranasal Nanotransferosomal Gel for Quercetin Brain Targeting: I. Optimization, Characterization, Brain Localization, and Cytotoxic Studies. Pharmaceutics 2023, 15, 1805. [Google Scholar] [CrossRef] [PubMed]
- Eid, H.M.; Ali, A.A.; Ali, A.M.A.; Eissa, E.M.; Hassan, R.M.; El-Ela, F.I.A.; Hassan, A.H. Potential Use of Tailored Citicoline Chitosan-Coated Liposomes for Effective Wound Healing in Diabetic Rat Model. Int. J. Nanomed. 2022, 17, 555. [Google Scholar] [CrossRef] [PubMed]
- Eid, H.M.; Naguib, I.A.; Alsantali, R.I.; Alsalahat, I.; Hegazy, A.M. Novel chitosan-coated niosomal formulation for improved management of bacterial conjunctivitis: A highly permeable and efficient ocular nanocarrier for azithromycin. J. Pharm. Sci. 2021, 110, 3027–3036. [Google Scholar] [CrossRef]
- Elkomy, M.H.; Alruwaili, N.K.; Elmowafy, M.; Shalaby, K.; Zafar, A.; Ahmad, N.; Alsalahat, I.; Ghoneim, M.M.; Eissa, E.M.; Eid, H.M. Surface-Modified Bilosomes Nanogel Bearing a Natural Plant Alkaloid for Safe Management of Rheumatoid Arthritis Inflammation. Pharmaceutics 2022, 14, 563. [Google Scholar] [CrossRef] [PubMed]
- Elkomy, M.H.; Eid, H.M.; Elmowafy, M.; Shalaby, K.; Zafar, A.; Abdelgawad, M.A.; Rateb, M.E.; Ali, M.R.; Alsalahat, I.; Abou-Taleb, H.A. Bilosomes as a promising nanoplatform for oral delivery of an alkaloid nutraceutical: Improved pharmacokinetic profile and snowballed hypoglycemic effect in diabetic rats. Drug Deliv. 2022, 29, 2694–2704. [Google Scholar] [CrossRef]
- Kaur, P.; Garg, T.; Vaidya, B.; Prakash, A.; Rath, G.; Goyal, A.K. Brain delivery of intranasal in situ gel of nanoparticulated polymeric carriers containing antidepressant drug: Behavioral and biochemical assessment. J. Drug Target. 2015, 23, 275–286. [Google Scholar] [CrossRef]
- Chenu, F.; Guiard, B.; Bourin, M.; Gardier, A. Antidepressant-like activity of selective serotonin reuptake inhibitors combined with a NK1 receptor antagonist in the mouse forced swimming test. Behav. Brain Res. 2006, 172, 256–263. [Google Scholar] [CrossRef]
- Can, A.; Dao, D.T.; Terrillion, C.E.; Piantadosi, S.C.; Bhat, S.; Gould, T.D. The tail suspension test. JoVE (J. Vis. Exp.) 2012, 59, e3769. [Google Scholar] [CrossRef]
- Dang, R.; Zhou, X.; Tang, M.; Xu, P.; Gong, X.; Liu, Y.; Jiao, H.; Jiang, P. Fish oil supplementation attenuates neuroinflammation and alleviates depressive-like behavior in rats submitted to repeated lipopolysaccharide. Eur. J. Nutr. 2018, 57, 893–906. [Google Scholar] [CrossRef]
- Lee, P.R.; Brady, D.L.; Shapiro, R.A.; Dorsa, D.M.; Koenig, J.I. Social interaction deficits caused by chronic phencyclidine administration are reversed by oxytocin. Neuropsychopharmacology 2005, 30, 1883–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodnoff, S.R.; Suranyi-Cadotte, B.; Aitken, D.H.; Quirion, R.; Meaney, M.J. The effects of chronic antidepressant treatment in an animal model of anxiety. Psychopharmacology 1988, 95, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Dulawa, S.C.; Hen, R. Recent advances in animal models of chronic antidepressant effects: The novelty-induced hypophagia test. Neurosci. Biobehav. Rev. 2005, 29, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Powell, T.R.; Fernandes, C.; Schalkwyk, L.C. Depression-related behavioral tests. Curr. Protoc. Mouse Biol. 2012, 2, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Blasco-Serra, A.; González-Soler, E.M.; Cervera-Ferri, A.; Teruel-Martí, V.; Valverde-Navarro, A.A. A standardization of the novelty-suppressed feeding test protocol in rats. Neurosci. Lett. 2017, 658, 73–78. [Google Scholar] [CrossRef]
- Fukumoto, K.; Chaki, S. Involvement of serotonergic system in the effect of a metabotropic glutamate 5 receptor antagonist in the novelty-suppressed feeding test. J. Pharmacol. Sci. 2015, 127, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-S.; Park, W.-S.; Kim, Y.H.; Kwon, S.-H.; Jang, Y.-J.; Han, D.; Morita, K.; Her, S. Antidepressant-like effects of Cortex Mori Radicis extract via bidirectional phosphorylation of glucocorticoid receptors in the hippocampus. Behav. Brain Res. 2013, 236, 56–61. [Google Scholar] [CrossRef]
- Sarkisova, K.Y.; Midzianovskaia, I.; Kulikov, M. Depressive-like behavioral alterations and c-fos expression in the dopaminergic brain regions in WAG/Rij rats with genetic absence epilepsy. Behav. Brain Res. 2003, 144, 211–226. [Google Scholar] [CrossRef]
- Abo El-Enin, H.A.; Elkomy, M.H.; Naguib, I.A.; Ahmed, M.F.; Alsaidan, O.A.; Alsalahat, I.; Ghoneim, M.M.; Eid, H.M. Lipid nanocarriers overlaid with chitosan for brain delivery of berberine via the nasal route. Pharmaceuticals 2022, 15, 281. [Google Scholar] [CrossRef]
- Elkomy, M.H.; Elmowafy, M.; Shalaby, K.; Azmy, A.F.; Ahmad, N.; Zafar, A.; Eid, H.M. Development and machine-learning optimization of mucoadhesive nanostructured lipid carriers loaded with fluconazole for treatment of oral candidiasis. Drug Dev. Ind. Pharm. 2021, 47, 246–258. [Google Scholar] [CrossRef]
- Panda, D.S.; Eid, H.M.; Elkomy, M.H.; Khames, A.; Hassan, R.M.; Abo El-Ela, F.I.; Yassin, H.A. Berberine encapsulated lecithin–chitosan nanoparticles as innovative wound healing agent in type II diabetes. Pharmaceutics 2021, 13, 1197. [Google Scholar] [CrossRef]
- Belmaker, R.H.; Agam, G. Major depressive disorder. N. Engl. J. Med. 2008, 358, 55–68. [Google Scholar] [CrossRef] [Green Version]
- Pothion, S.; Bizot, J.-C.; Trovero, F.; Belzung, C. Strain differences in sucrose preference and in the consequences of unpredictable chronic mild stress. Behav. Brain Res. 2004, 155, 135–146. [Google Scholar] [CrossRef]
- Willner, P. Chronic mild stress (CMS) revisited: Consistency and behavioural-neurobiological concordance in the effects of CMS. Neuropsychobiology 2005, 52, 90–110. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J.; Hyman, S.E. Animal models of neuropsychiatric disorders. Nat. Neurosci. 2010, 13, 1161–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willner, P.; Towell, A.; Sampson, D.; Sophokleous, S.; Muscat, R.a. Reduction of sucrose preference by chronic unpredictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharmacology 1987, 93, 358–364. [Google Scholar] [CrossRef]
- Hua-Cheng, Y.; Xiong, C.; Manas, D.; Xin-Hong, Z.; Tian-Ming, G. Behavioral animal models of depression. Neurosci. Bull. 2010, 26, 327–337. [Google Scholar]
- Zhu, X.-H.; Yan, H.-C.; Zhang, J.; Qu, H.-D.; Qiu, X.-S.; Chen, L.; Li, S.-J.; Cao, X.; Bean, J.C.; Chen, L.-H. Intermittent hypoxia promotes hippocampal neurogenesis and produces antidepressant-like effects in adult rats. J. Neurosci. 2010, 30, 12653–12663. [Google Scholar] [CrossRef] [Green Version]
- Tong, G.-F.; Qin, N.; Sun, L.-W. Development and evaluation of Desvenlafaxine loaded PLGA-chitosan nanoparticles for brain delivery. Saudi Pharm. J. 2017, 25, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Rénéric, J.-P.; Lucki, I. Antidepressant behavioral effects by dual inhibition of monoamine reuptake in the rat forced swimming test. Psychopharmacology 1998, 136, 190–197. [Google Scholar] [CrossRef]
- Santarelli, L.; Saxe, M.; Gross, C.; Surget, A.; Battaglia, F.; Dulawa, S.; Weisstaub, N.; Lee, J.; Duman, R.; Arancio, O. Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science 2003, 301, 805–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessa, J.M.; Mesquita, A.R.; Oliveira, M.; Pêgo, J.M.; Cerqueira, J.J.; Palha, J.A.; Almeida, O.F.; Sousa, N. A trans-dimensional approach to the behavioral aspects of depression. Front. Behav. Neurosci. 2009, 3, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragunow, M. A role for immediate-early transcription factors in learning and memory. Behav. Genet. 1996, 26, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Warburton, E.; Kuśmierek, P.; Aggleton, J.P.; Kowalska, D.; Brown, M. Fos imaging reveals differential neuronal activation of areas of rat temporal cortex by novel and familiar sounds. Eur. J. Neurosci. 2001, 14, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Sarkisova, K.I.; Midzianovskaia, I.; Kulikov, M. Depression-like changes in behavior and c-fos gene expression in dopaminergic brain structures in WAG/Rij rats. Zhurnal Vyss. Nervn. Deiatelnosti Im. IP Pavlov 2002, 52, 733–742. [Google Scholar]
- Silveira, M.C.L.; Zangrossi, H., Jr.; de Barros Viana, M.; Silveira, R.; Graeff, F.G. Differential expression of Fos protein in the rat brain induced by performance of avoidance or escape in the elevated T-maze. Behav. Brain Res. 2001, 126, 13–21. [Google Scholar] [CrossRef]
- Elkomy, M.H.; Ali, A.A.; Eid, H.M. Chitosan on the surface of nanoparticles for enhanced drug delivery: A comprehensive review. J. Control. Release 2022, 351, 923–940. [Google Scholar] [CrossRef]
- Alam, M.I.; Baboota, S.; Ahuja, A.; Ali, M.; Ali, J.; Sahni, J.K. Intranasal administration of nanostructured lipid carriers containing CNS acting drug: Pharmacodynamic studies and estimation in blood and brain. J. Psychiatr. Res. 2012, 46, 1133–1138. [Google Scholar] [CrossRef]
- Haque, S.; Md, S.; Sahni, J.K.; Ali, J.; Baboota, S. Development and evaluation of brain targeted intranasal alginate nanoparticles for treatment of depression. J. Psychiatr. Res. 2014, 48, 1–12. [Google Scholar] [CrossRef]
- Hermann, D.M.; Bassetti, C.L. Implications of ATP-binding cassette transporters for brain pharmacotherapies. Trends Pharmacol. Sci. 2007, 28, 128–134. [Google Scholar] [CrossRef]
- Salama, H.A.; Mahmoud, A.A.; Kamel, A.O.; Hady, M.A.; Awad, G.A. Phospholipid based colloidal poloxamer–nanocubic vesicles for brain targeting via the nasal route. Colloids Surf. B Biointerfaces 2012, 100, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Ceña, V.; Játiva, P. Nanoparticle crossing of blood–brain barrier: A road to new therapeutic approaches to central nervous system diseases. Future Med. 2018, 13, 1513–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betzer, O.; Shilo, M.; Opochinsky, R.; Barnoy, E.; Motiei, M.; Okun, E.; Yadid, G.; Popovtzer, R. The effect of nanoparticle size on the ability to cross the blood–brain barrier: An in vivo study. Nanomedicine 2017, 12, 1533–1546. [Google Scholar] [CrossRef] [PubMed]
- Dong, X. Current strategies for brain drug delivery. Theranostics 2018, 8, 1481. [Google Scholar] [CrossRef]
- Lim, W.M.; Rajinikanth, P.S.; Mallikarjun, C.; Kang, Y.B. Formulation and delivery of itraconazole to the brain using a nanolipid carrier system. Int. J. Nanomed. 2014, 9, 2117–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rana, I.; Khan, N.; Ansari, M.M.; Shah, F.A.; ud Din, F.; Sarwar, S.; Imran, M.; Qureshi, O.S.; Choi, H.-I.; Lee, C.-H. Solid lipid nanoparticles-mediated enhanced antidepressant activity of duloxetine in lipopolysaccharide-induced depressive model. Colloids Surf. B Biointerfaces 2020, 194, 111209. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Treatment | Dose | Parameters | ||
|---|---|---|---|---|---|
| Immobility (s) | Swimming (s) | Climbing (s) | |||
| 1 | Control Negative | 2 mL/kg (normal saline, not depressed) | 58.7 ± 7.8 | 155.3 ± 8.7 | 141.2 ± 6.3 |
| 2 | Control Positive | No treatment, depressed | 234.2 ± 16.1 (a) | 35.6 ± 6.5 (a) | 22.8 ± 6.0 (a) |
| 3 | QER-SOLN (oral) | 300 mg/kg (quercetin, depressed) | 127.3 ± 11.8 (a,b) | 88.2 ± 12.8 (a,b) | 86.8 ± 6.7 (a,b) |
| 4 | QER-SOLN gel (IN) | 10 mg/kg (quercetin, depressed) | 119.4 ± 7.5 (a,b) | 104.3 ± 9.5 (a,b,c) | 96.2 ± 6.4 (a,b) |
| 5 | QER-TFS gel (IN) | 10 mg/kg (quercetin, depressed) | 52.8 ± 6.4 (b,c,d) | 181.2 ± 11.9 (a,b,c,d) | 157.3 ± 12.5 (a,b,c,d) |
| Group | Treatment | Positive Immune Cells (%) * |
|---|---|---|
| 1 | Control Negative (normal) | 15.00 ± 3.00 |
| 2 | Control Positive (depressed) | 85.00 ± 1.00 |
| 3 | QER-SOLN (oral) | 75.00 ± 3.00 |
| 4 | QER-SOLN gel (IN) | 67.00 ± 3.00 |
| 5 | QER-TFS gel (IN) | 5.00 ± 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elkomy, M.H.; Abo El-Ela, F.I.; Zaki, R.M.; Alsaidan, O.A.; Elmowafy, M.; Zafar, A.; Shalaby, K.; Abdelgawad, M.A.; Omar, H.A.; Salama, R.; et al. Intranasal Nanotransferosomal Gel for Quercetin Brain Targeting: II. Antidepressant Effect in an Experimental Animal Model. Pharmaceutics 2023, 15, 2095. https://doi.org/10.3390/pharmaceutics15082095
Elkomy MH, Abo El-Ela FI, Zaki RM, Alsaidan OA, Elmowafy M, Zafar A, Shalaby K, Abdelgawad MA, Omar HA, Salama R, et al. Intranasal Nanotransferosomal Gel for Quercetin Brain Targeting: II. Antidepressant Effect in an Experimental Animal Model. Pharmaceutics. 2023; 15(8):2095. https://doi.org/10.3390/pharmaceutics15082095
Chicago/Turabian StyleElkomy, Mohammed H., Fatma I. Abo El-Ela, Randa Mohammed Zaki, Omar A. Alsaidan, Mohammed Elmowafy, Ameeduzzafar Zafar, Khaled Shalaby, Mohamed A. Abdelgawad, Hany A. Omar, Rania Salama, and et al. 2023. "Intranasal Nanotransferosomal Gel for Quercetin Brain Targeting: II. Antidepressant Effect in an Experimental Animal Model" Pharmaceutics 15, no. 8: 2095. https://doi.org/10.3390/pharmaceutics15082095