
Lysine-Dendrimer, a New Non-Aggressive Solution to Rebalance the Microbiota of Acne-Prone Skin
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of G2-Dendrimer
2.2. Cell Culture
2.3. IL-8 Production
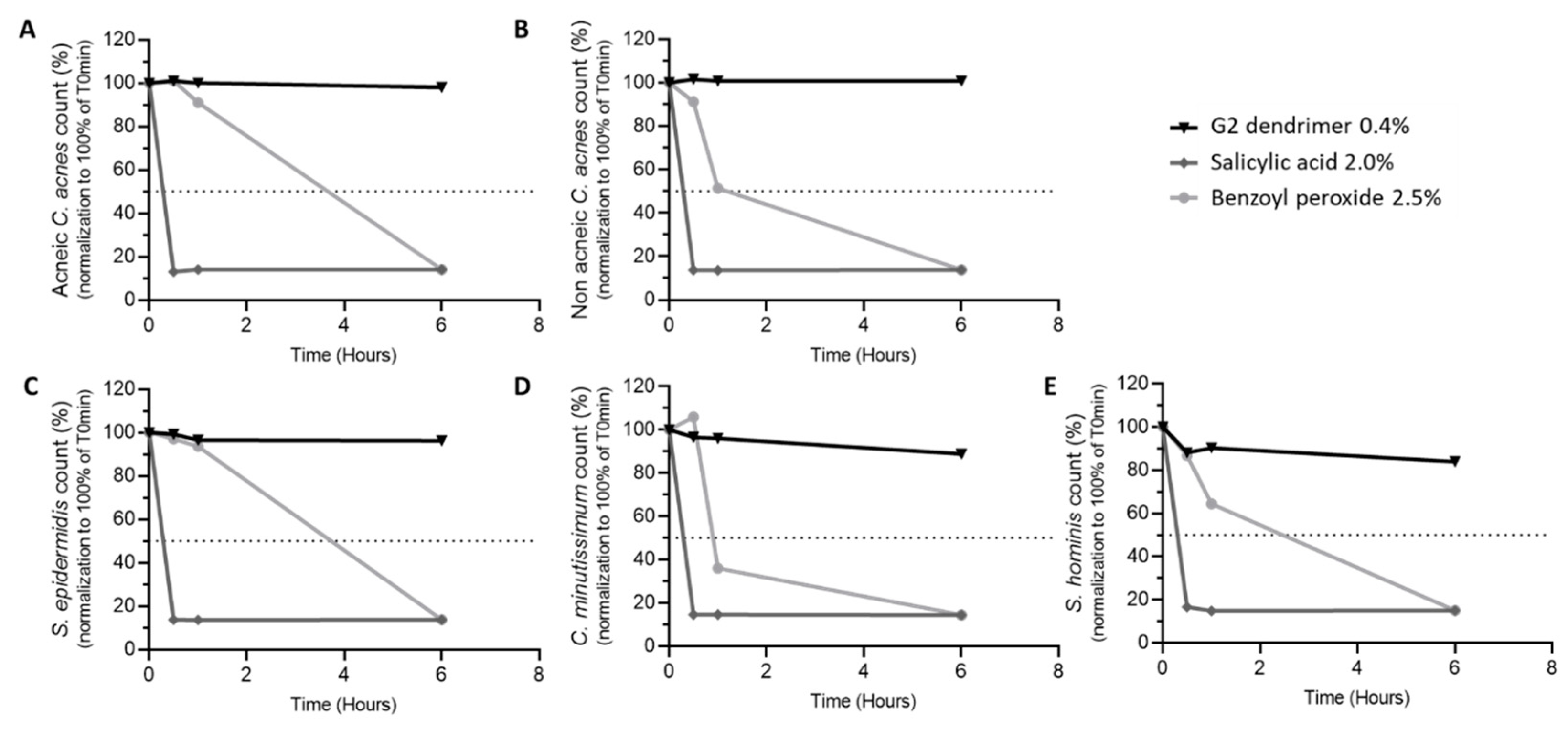
2.4. Bactericidal Activity
2.5. Clinical Study with G2-Dendrimer Versus Benchmark
2.6. Clinical Study with G2-Dendrimer in Full Formulation
2.7. Biofilm Formation on Glass Slides
2.8. Skin Microbiome Sampling
2.9. Statistical Tests
3. Results
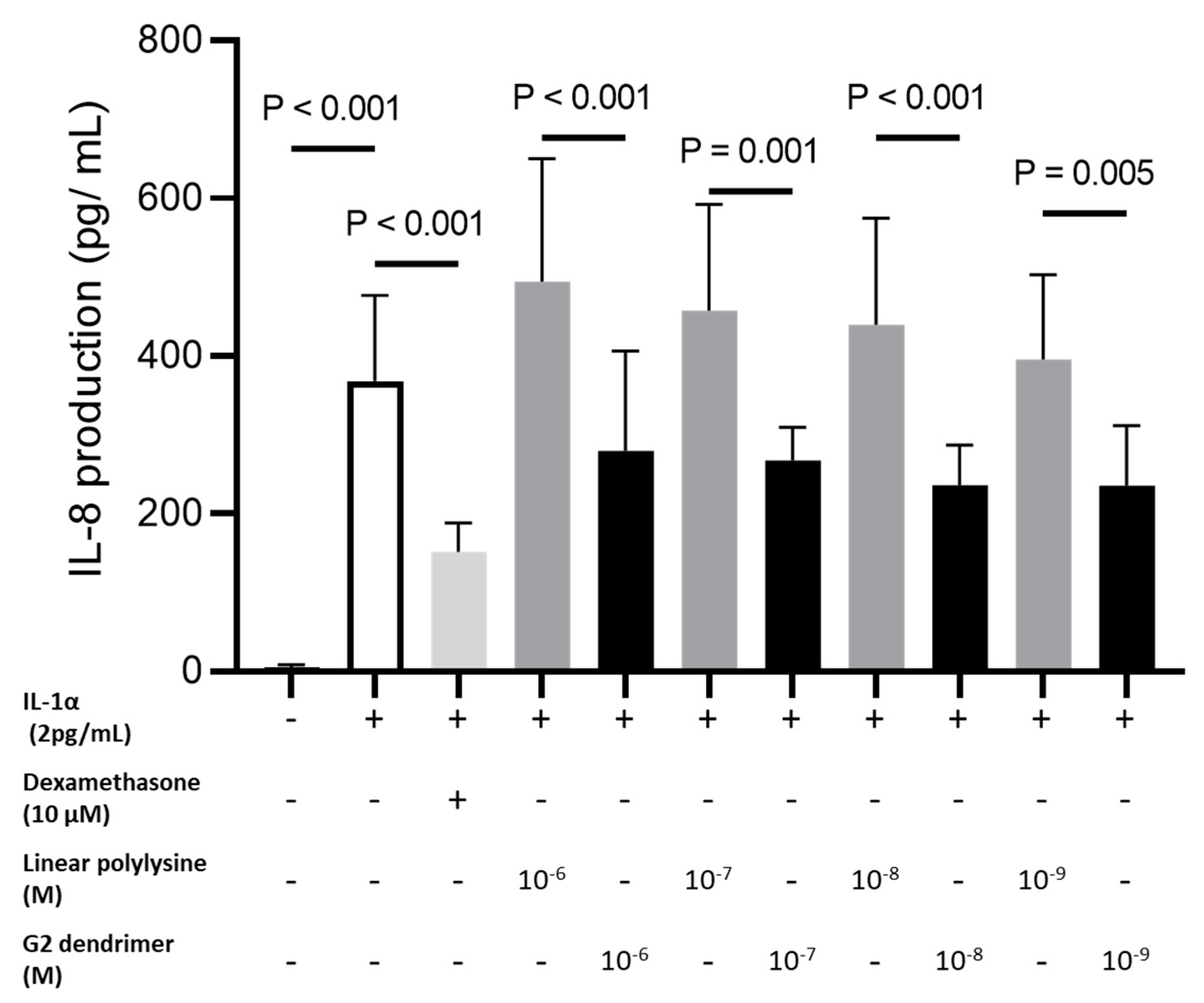
3.1. Anti-Inflammatory Activity of Dendrimer
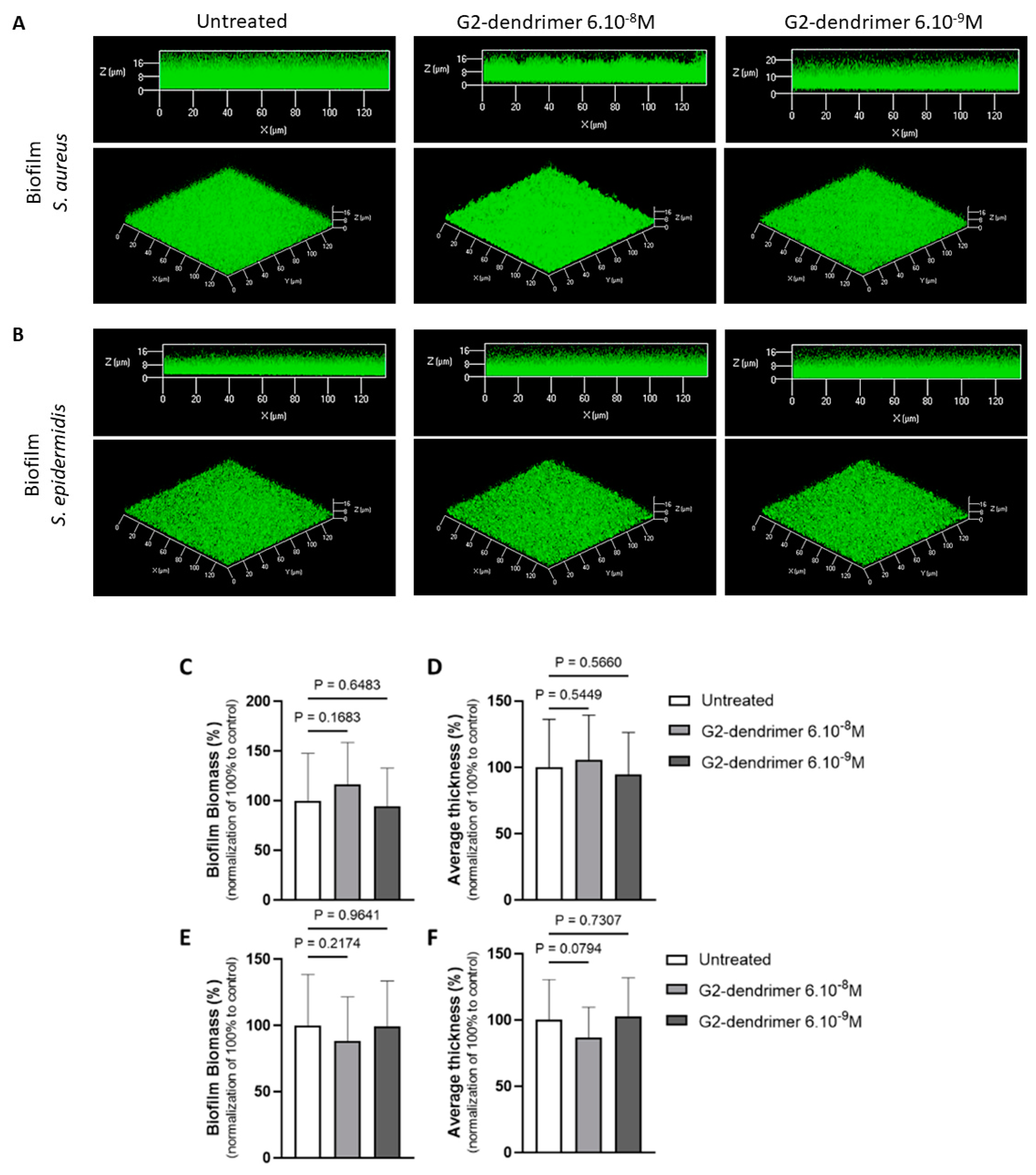
3.2. Effect on Staphyloccocus Aureus and Staphyloccocus Epidermidis Biofilm Formation
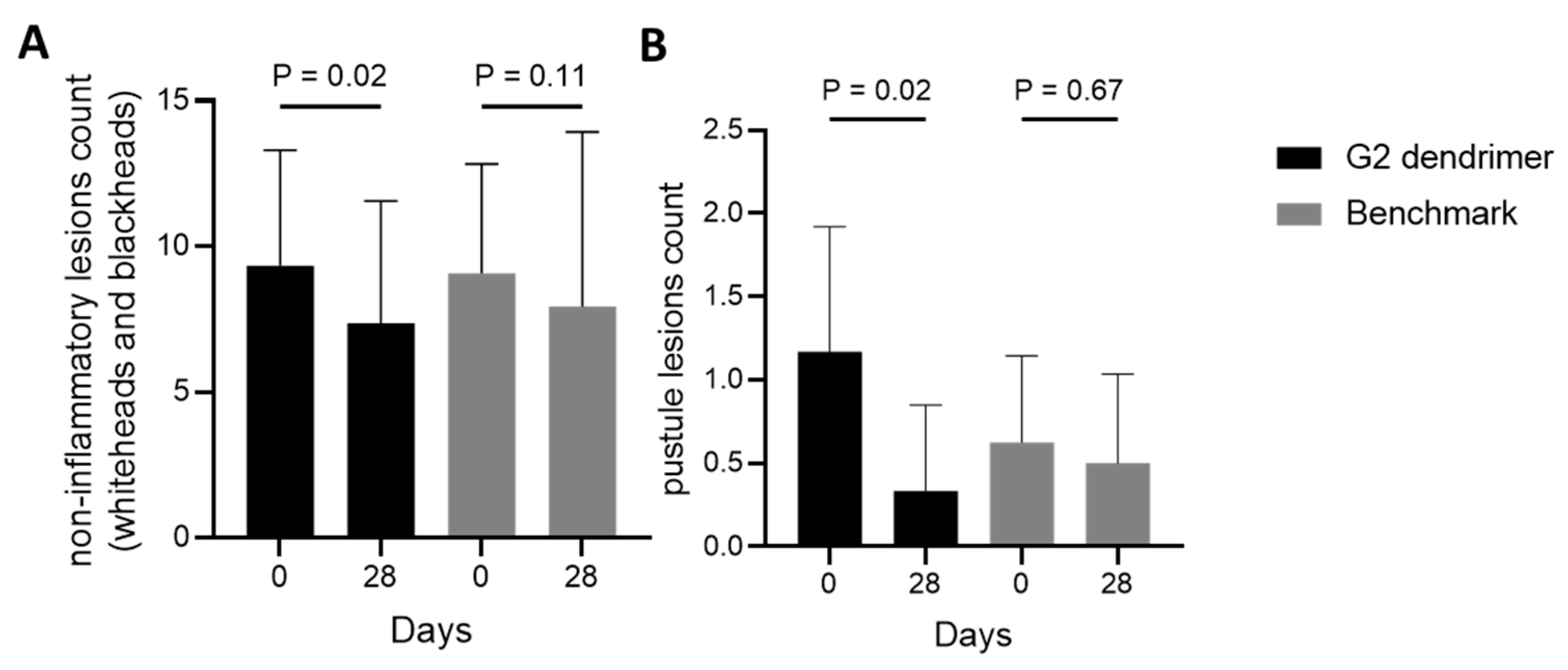
3.3. Efficacy of G2-Dendrimer Versus Benzoyl Peroxide in a Clinical Study
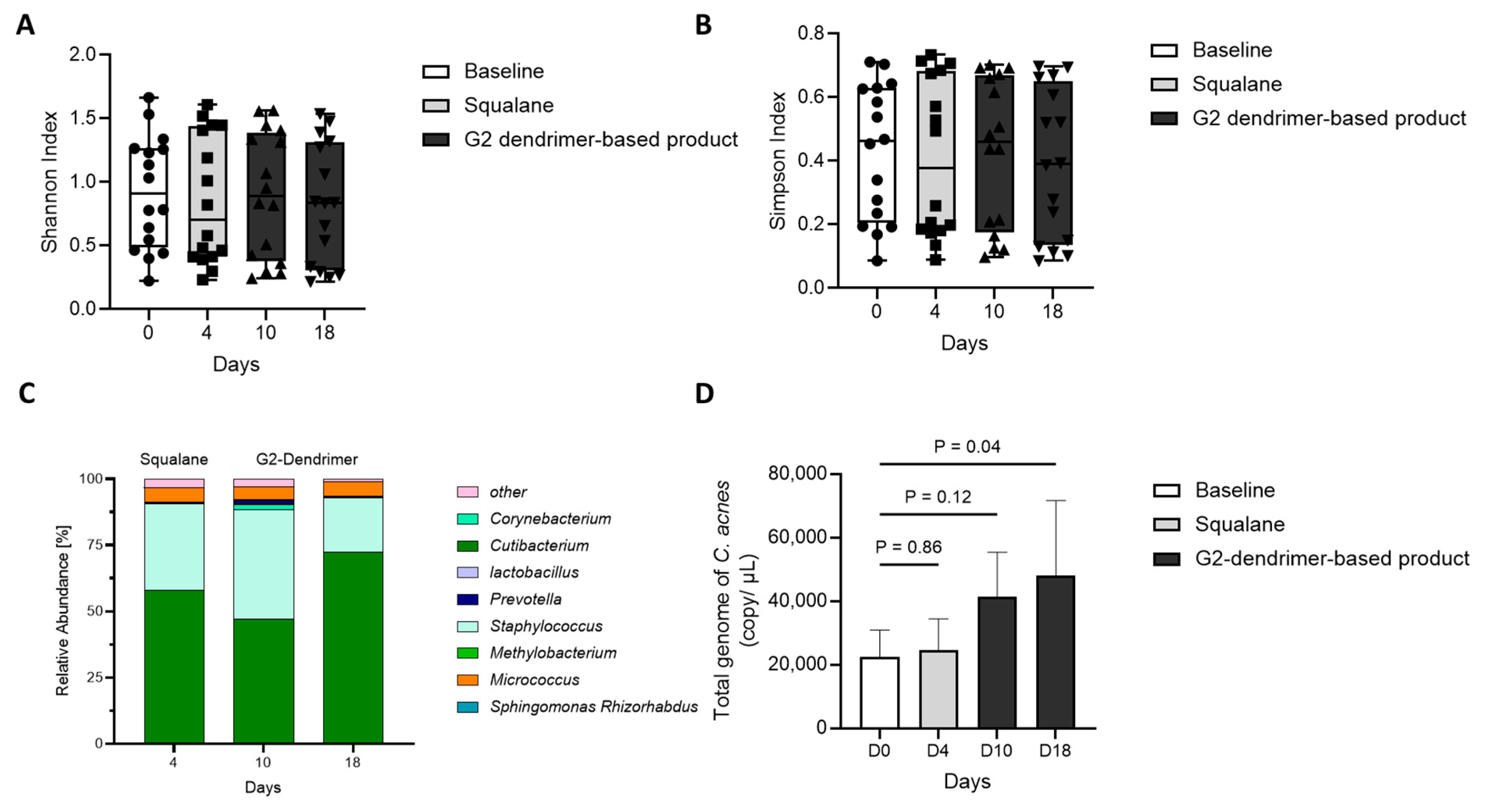
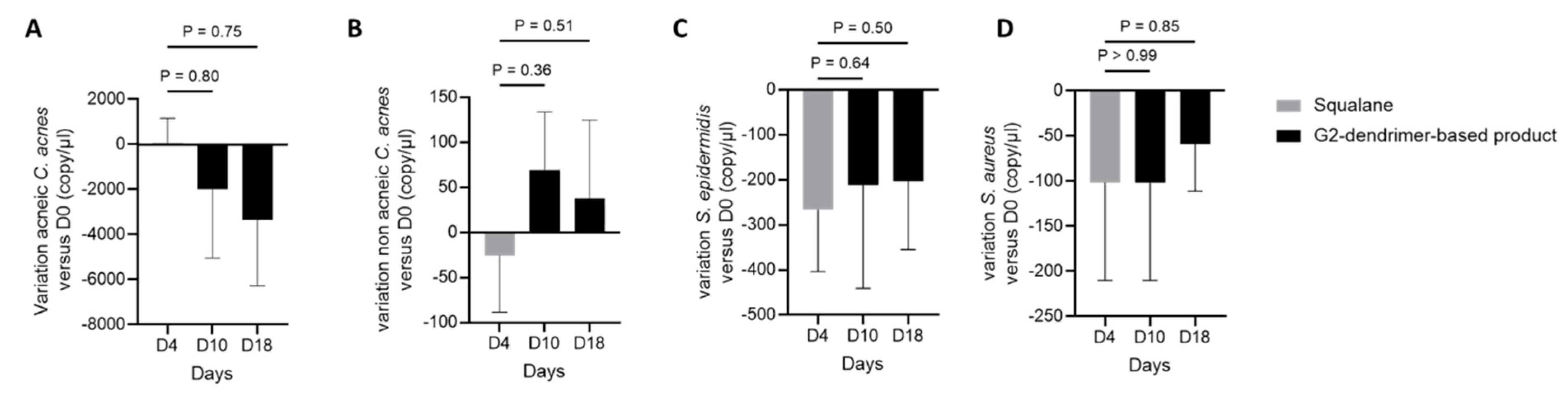
3.4. Impact of G2-Dendrimer on Commensal Bacteria
3.5. Distribution of C. acnes
4. Discussion
5. Conclusions
6. Potential Limitation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dagnelie, M.A.; Poinas, A.; Dréno, B. What is new in adult acne for the last 2 years: Focus on acne pathophysiology and treatments. Int. J. Dermatol. 2022, 61, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Akhavan, A.; Bershad, S. Topical acne drugs: Review of clinical properties, systemic exposure, and safety. Am. J. Clin. Dermatol. 2003, 4, 473–492. [Google Scholar] [CrossRef] [PubMed]
- Feldman, S.; Careccia, R.E.; Barham, K.L.; Hancox, J. Diagnosis and treatment of acne. Am. Fam. Physician 2004, 69, 2123–2130. [Google Scholar] [PubMed]
- Ochsendorf, F.R. Cutibacterium acnes in acne pathophysiology—The chicken or the egg? Br. J. Dermatol. 2019, 181, 657–658. [Google Scholar] [CrossRef]
- Brzuszkiewicz, E.; Weiner, J.; Wollherr, A.; Thürmer, A.; Hüpeden, J.; Lomholt, H.B.; Kilian, M.; Gottschalk, G.; Daniel, R.; Mollenkopf, H.-J.; et al. Comparative genomics and transcriptomics of Propionibacterium acnes. PLoS ONE 2011, 6, e21581. [Google Scholar] [CrossRef]
- Fitz-Gibbon, S.; Tomida, S.; Chiu, B.-H.; Nguyen, L.; Du, C.; Liu, M.; Elashoff, D.; Erfe, M.C.; Loncaric, A.; Kim, J.; et al. Propionibacterium acnes strain populations in the human skin microbiome associated with acne. J. Investig. Dermatol. 2013, 133, 2152–2160. [Google Scholar] [CrossRef] [Green Version]
- Ramasamy, S.; Barnard, E.; Dawson, T.L., Jr.; Li, H. The role of the skin microbiota in acne pathophysiology. Br. J. Dermatol. 2019, 181, 691–699. [Google Scholar] [CrossRef]
- Dréno, B.; Dagnelie, M.A.; Khammari, A.; Corvec, S. The Skin Microbiome: A New Actor in Inflammatory Acne. Am. J. Clin. Dermatol. 2020, 21, 18–24. [Google Scholar] [CrossRef]
- Williams, H.C.; Dellavalle, R.P.; Garner, S. Acne vulgaris. Lancet 2012, 379, 361–372. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, Y.; Lazic Mosler, E.; Hu, J.; Li, H.; Zhang, Y.; Liu, J.; Zhang, Q. Topical benzoyl peroxide for acne. Cochrane Database Syst. Rev. 2020, 3, CD011154. [Google Scholar] [CrossRef]
- Thiboutot, D.; Zaenglein, A.; Weiss, J.; Webster, G.; Calvarese, B.; Chen, D. An aqueous gel fixed combination of clindamycin phosphate 1.2% and benzoyl peroxide 2.5% for the once-daily treatment of moderate to severe acne vulgaris: Assessment of efficacy and safety in 2813 patients. J. Am. Acad. Dermatol. 2008, 59, 792–800. [Google Scholar] [CrossRef]
- Al-Talib, H.; Al-Khateeb, A.; Hameed, A.; Murugaiah, C. Efficacy and safety of superficial chemical peeling in treatment of active acne vulgaris. An. Bras. Dermatol. 2017, 92, 212–216. [Google Scholar] [CrossRef] [Green Version]
- Lam, M.; Hu, A.; Fleming, P.; Lynde, C.W. The Impact of Acne Treatment on Skin Bacterial Microbiota: A Systematic Review. J. Cutan. Med. Surg. 2022, 26, 93–97. [Google Scholar] [CrossRef]
- O’neill, A.M.; Gallo, R.L. Host-microbiome interactions and recent progress into understanding the biology of acne vulgaris. Microbiome 2018, 6, 177. [Google Scholar] [CrossRef]
- Chien, A.L.; Tsai, J.; Leung, S.; Mongodin, E.F.; Nelson, A.M.; Kang, S.; Garza, L.A. Association of Systemic Antibiotic Treatment of Acne with Skin Microbiota Characteristics. JAMA Dermatol. 2019, 155, 425–434. [Google Scholar] [CrossRef]
- Grice, E.A.; Segre, J.A. The skin microbiome. Nat. Rev. Microbiol. 2011, 9, 244–253. [Google Scholar] [CrossRef]
- Cavallo, I.; Sivori, F.; Truglio, M.; De Maio, F.; Lucantoni, F.; Cardinali, G.; Pontone, M.; Bernardi, T.; Sanguinetti, M.; Capitanio, B.; et al. Skin dysbiosis and Cutibacterium acnes biofilm in inflammatory acne lesions of adolescents. Sci. Rep. 2022, 12, 21104. [Google Scholar] [CrossRef]
- Attia-Vigneau, J.; Barreau, M.; Le Toquin, E.; Feuilloley, M.G.J.; Loing, E.; Lesouhaitier, O. Polylysine dendrigraft is able to differentially impact Cutibacterium acnes strains preventing acneic skin. Exp. Dermatol. 2022, 31, 1056–1064. [Google Scholar] [CrossRef]
- Radzishevsky, I.S.; Rotem, S.; Bourdetsky, D.; Navon-Venezia, S.; Carmeli, Y.; Mor, A. Improved antimicrobial peptides based on acyl-lysine oligomers. Nat. Biotechnol. 2007, 25, 657–659. [Google Scholar] [CrossRef]
- Borrel, V.; Thomas, P.; Catovic, C.; Racine, P.-J.; Konto-Ghiorghi, Y.; Lefeuvre, L.; Duclairoir-Poc, C.; Zouboulis, C.C.; Feuilloley, M.G.J. Acne and Stress: Impact of Catecholamines on Cutibacterium acnes. Front. Med. 2019, 6, 155. [Google Scholar] [CrossRef]
- Belkaid, Y.; Segre, J.A. Dialogue between skin microbiota and immunity. Science 2014, 346, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Ring, H.C.; Thorsen, J.; Saunte, D.M.; Lilje, B.; Bay, L.; Riis, P.T.; Larsen, N.; Andersen, L.O.; Nielsen, H.V.; Miller, I.M.; et al. The Follicular Skin Microbiome in Patients with Hidradenitis Suppurativa and Healthy Controls. JAMA Dermatol. 2017, 153, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Collet, H.; Souaid, E.; Cottet, H.; Deratani, A.; Boiteau, L.; Dessalces, G.; Rossi, J.-C.; Commeyras, A.; Pascal, R. An expeditious multigram-scale synthesis of lysine dendrigraft (DGL) polymers by aqueous N-carboxyanhydride polycondensation. Chemistry 2010, 16, 2309–2316. [Google Scholar] [CrossRef] [PubMed]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersbøll, B.K.; Molin, S. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology 2000, 146 Pt 10, 2395–2407. [Google Scholar] [CrossRef] [Green Version]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Hillion, M.; Mijouin, L.; Jaouen, T.; Barreau, M.; Meunier, P.; Lefeuvre, L.; Lati, E.; Chevalier, S.; Feuilloley, M.G. Comparative study of normal and sensitive skin aerobic bacterial populations. MicrobiologyOpen 2013, 2, 953–961. [Google Scholar] [CrossRef]
- Janvier, X.; Boukerb, A.M.; Feuilloley, M.G.J.; Groboillot, A. Draft Genome Sequences of Four Commensal Strains of Staphylococcus and Pseudomonas Isolated from Healthy Human Skin. Microbiol. Resour. Announc. 2021, 10, e01032-20. [Google Scholar] [CrossRef]
- Bent, S.J.; Forney, L.J. The tragedy of the uncommon: Understanding limitations in the analysis of microbial diversity. ISME J. 2008, 2, 689–695. [Google Scholar] [CrossRef] [Green Version]
- DeJong, T.M. A Comparison of Three Diversity Indices Based on Their Components of Richness and Evenness. Oikos 1975, 26, 222. [Google Scholar] [CrossRef]
- Scharschmidt, T.C.; Fischbach, M.A. What lives on our skin: Ecology, genomics and therapeutic opportunities of the skin microbiome. Drug Discov. Today Dis. Mech. 2013, 10, e83–e89. [Google Scholar] [CrossRef] [Green Version]
- Grice, E.A. The intersection of microbiome and host at the skin interface: Genomic- and metagenomic-based insights. Genome Res. 2015, 25, 1514–1520. [Google Scholar] [CrossRef]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; NISC Comparative Sequencing Program; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; et al. Topographical and Temporal Diversity of the Human Skin Microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef] [Green Version]
- Barnard, E.; Shi, B.; Kang, D.; Craft, N.; Li, H. The balance of metagenomic elements shapes the skin microbiome in acne and health. Sci. Rep. 2016, 6, 39491. [Google Scholar] [CrossRef] [Green Version]
- Gribbon, E.M.; Cunliffe, W.J.; Holland, K.T. Interaction of Propionibacterium acnes with skin lipids in vitro. J. Gen. Microbiol. 1993, 139, 1745–1751. [Google Scholar] [CrossRef] [Green Version]
- Dreno, B.; Martin, R.; Moyal, D.; Henley, J.B.; Khammari, A.; Seité, S. Skin microbiome and acne vulgaris: Staphylococcus, a new actor in acne. Exp. Dermatol. 2017, 26, 798–803. [Google Scholar] [CrossRef] [Green Version]
- Bek-Thomsen, M.; Lomholt, H.B.; Kilian, M. Acne is Not Associated with Yet-Uncultured Bacteria. J. Clin. Microbiol. 2008, 46, 3355–3360. [Google Scholar] [CrossRef] [Green Version]
- Otto, M. Staphylococcus colonization of the skin and antimicrobial peptides. Expert Rev. Dermatol. 2010, 5, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Shima, S.; Matsuoka, H.; Iwamoto, T.; Sakai, H. Antimicrobial action of epsilon-poly-L-lysine. J. Antibiot. 1984, 37, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Kircik, L.H. The role of benzoyl peroxide in the new treatment paradigm for acne. J. Drugs Dermatol. 2013, 12, s73–s76. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (Trade Name) | INCI Name | Active Lotion (%) |
|---|---|---|
| Dermofeel PA-3 | Sodium phytate (and) aqua (and) alcohol | 0.10 |
| Ecogel | Lysolecithin (and) sclerotium gum (and) xanthan gum (and) pullulan | 2.00 |
| SCB jojoba oil | Simmondsia chinensis seed oil | 3.00 |
| Dermofeel toco 70 non gmo | Tocopherol (and) helianthus annuus (sunflower) seed oil | 0.10 |
| Lipex shea W | Shea butter cetyl esters | 3.00 |
| Saboderm TCC | Caprylic/capric triglyceride | 3.00 |
| Dekaben C4 | Phenoxyethanol (and) methylparaben (and) ethylparaben (and) butylparaben (and) propylparaben | 0.80 |
| G2-dendrimer | Glycerin (and) water (and) polylysine | 0.0005 |
| TOTAL | 100.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leignadier, J.; Drago, M.; Lesouhaitier, O.; Barreau, M.; Dashi, A.; Worsley, O.; Attia-Vigneau, J. Lysine-Dendrimer, a New Non-Aggressive Solution to Rebalance the Microbiota of Acne-Prone Skin. Pharmaceutics 2023, 15, 2083. https://doi.org/10.3390/pharmaceutics15082083
Leignadier J, Drago M, Lesouhaitier O, Barreau M, Dashi A, Worsley O, Attia-Vigneau J. Lysine-Dendrimer, a New Non-Aggressive Solution to Rebalance the Microbiota of Acne-Prone Skin. Pharmaceutics. 2023; 15(8):2083. https://doi.org/10.3390/pharmaceutics15082083
Chicago/Turabian StyleLeignadier, Julie, Marie Drago, Olivier Lesouhaitier, Magalie Barreau, Albert Dashi, Oliver Worsley, and Joan Attia-Vigneau. 2023. "Lysine-Dendrimer, a New Non-Aggressive Solution to Rebalance the Microbiota of Acne-Prone Skin" Pharmaceutics 15, no. 8: 2083. https://doi.org/10.3390/pharmaceutics15082083