
Immunometabolism Modulation by Extracts from Pistachio Stalks Formulated in Phospholipid Vesicles
 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Extract Preparation
2.2. LC-ESI/Quadrupole-Orbitrap/MS Analysis
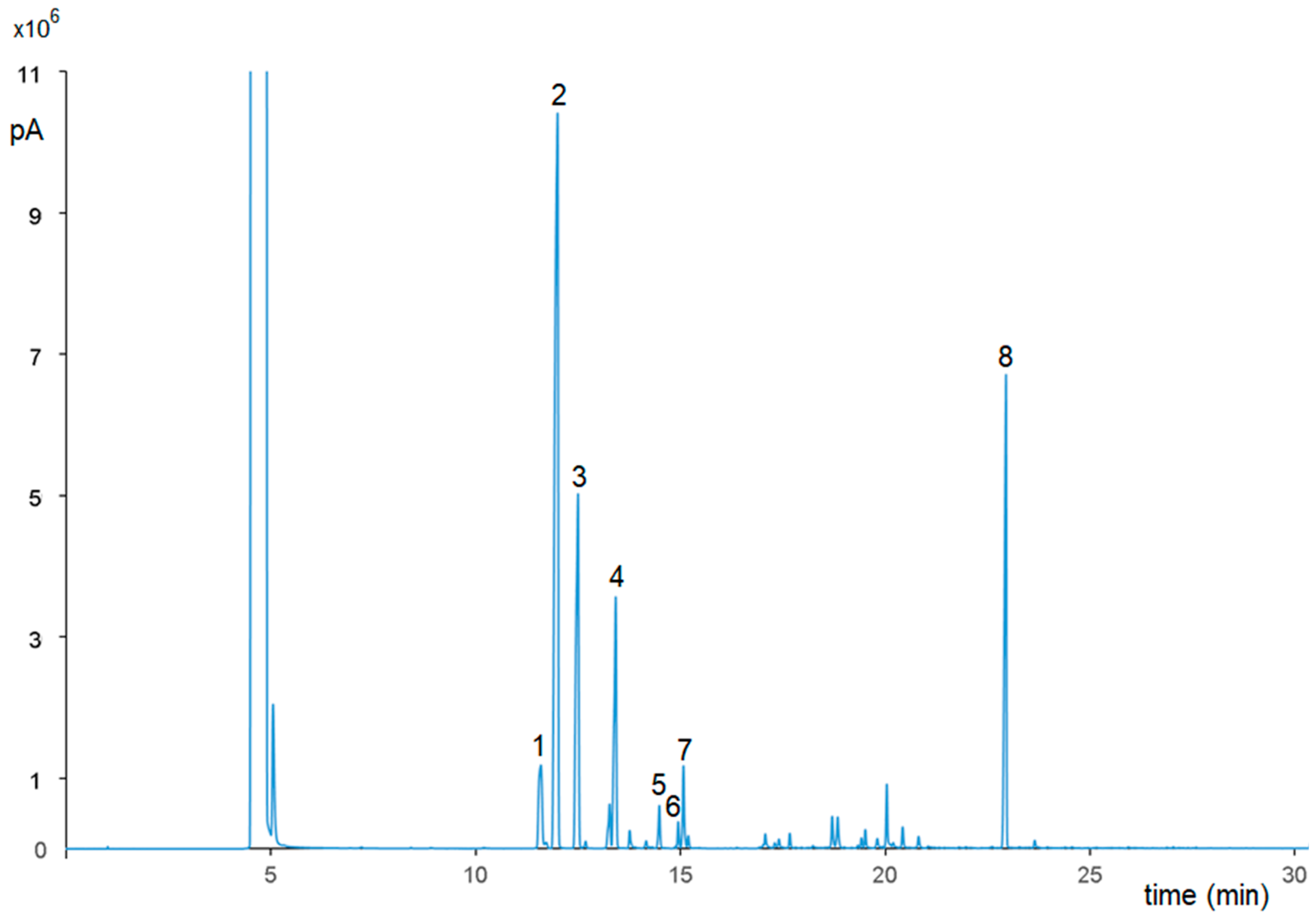
2.3. GC-FID and GC-MS Characterization of P. vera Essential Oil
2.4. Materials
2.5. Vesicle Preparation
2.6. Vesicle Characterization
2.7. Cell Culture and Treatments
2.8. Cell Viability Assay
2.9. Western Blotting
2.10. ACLY Activity
2.11. ROS, NO• and PGE2 Detection
2.12. Cellular Uptake of P. vera Extracts by LC–MS Analysis
2.13. Statistical Analysis
3. Results
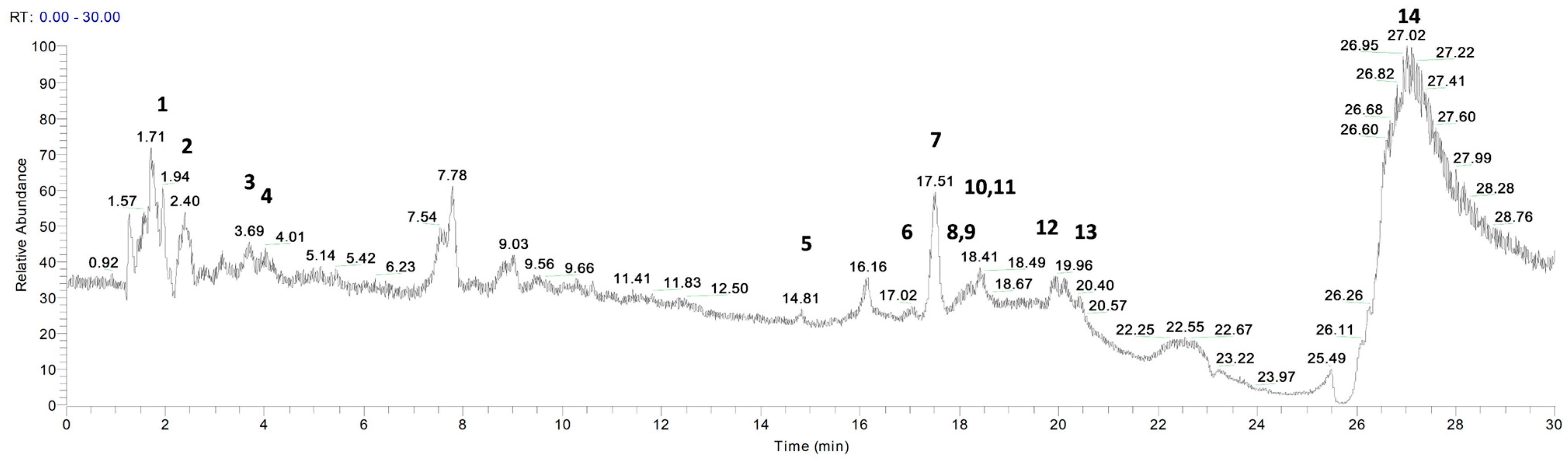
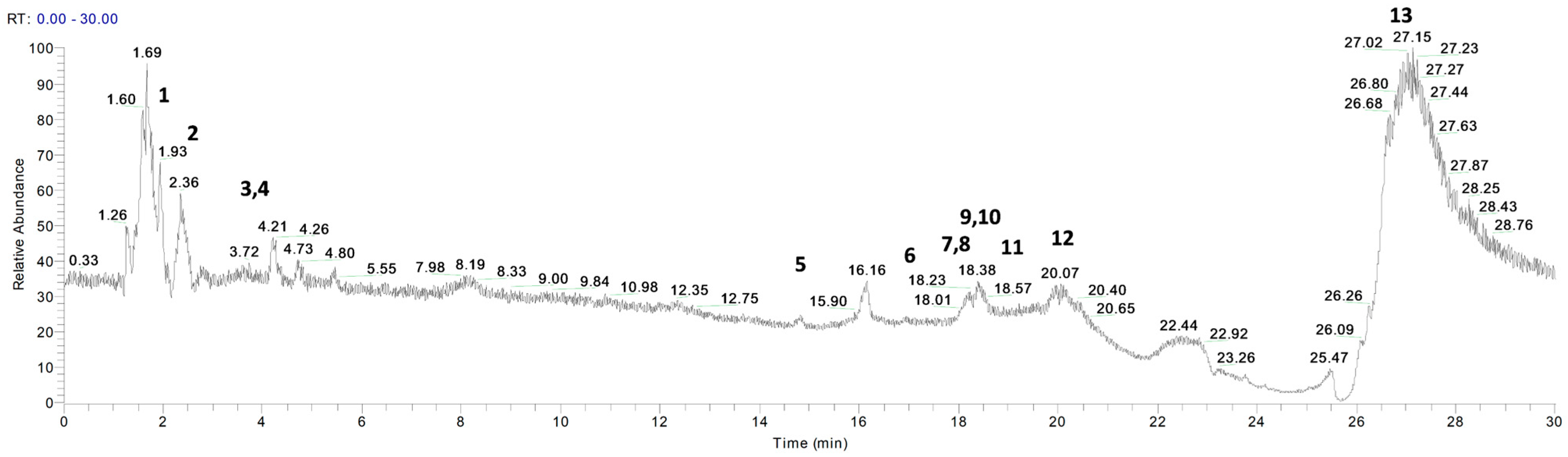
3.1. Phytochemical Profile of P. vera Extracts
3.2. Vesicle Characterization
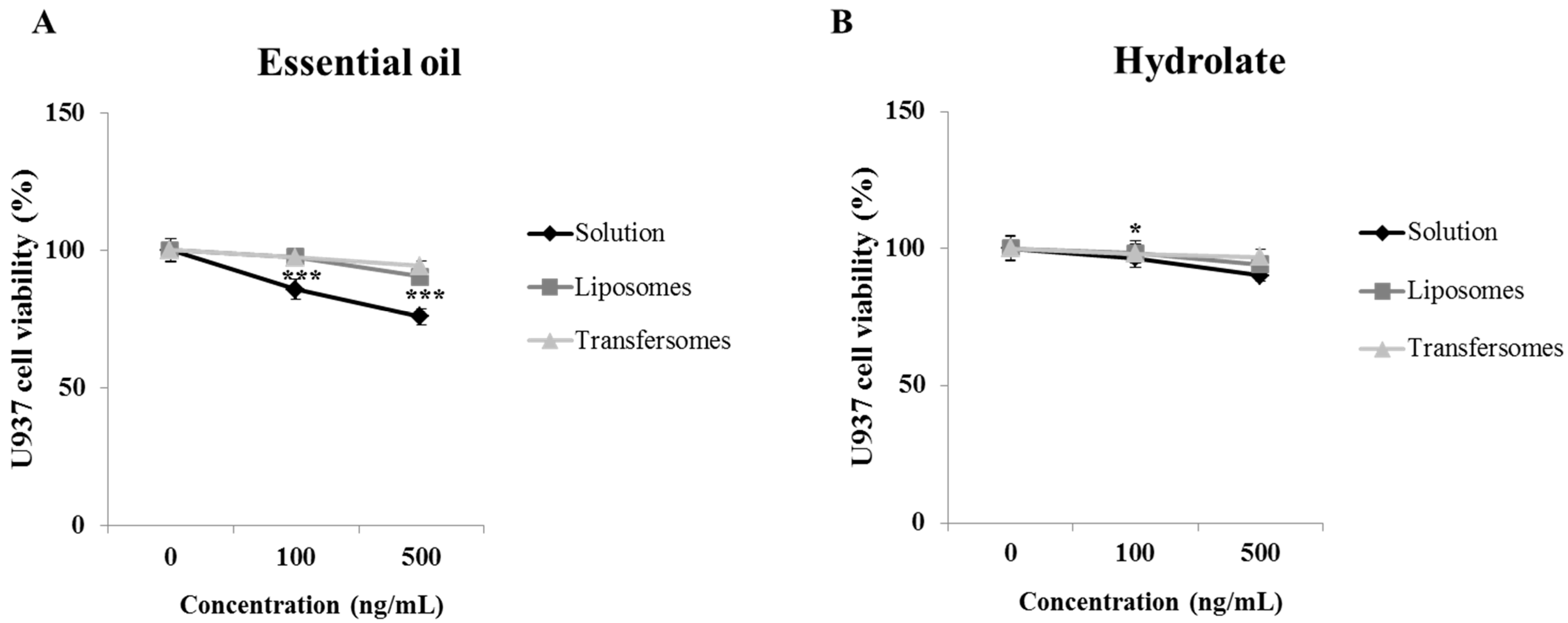
3.3. Cell Viability
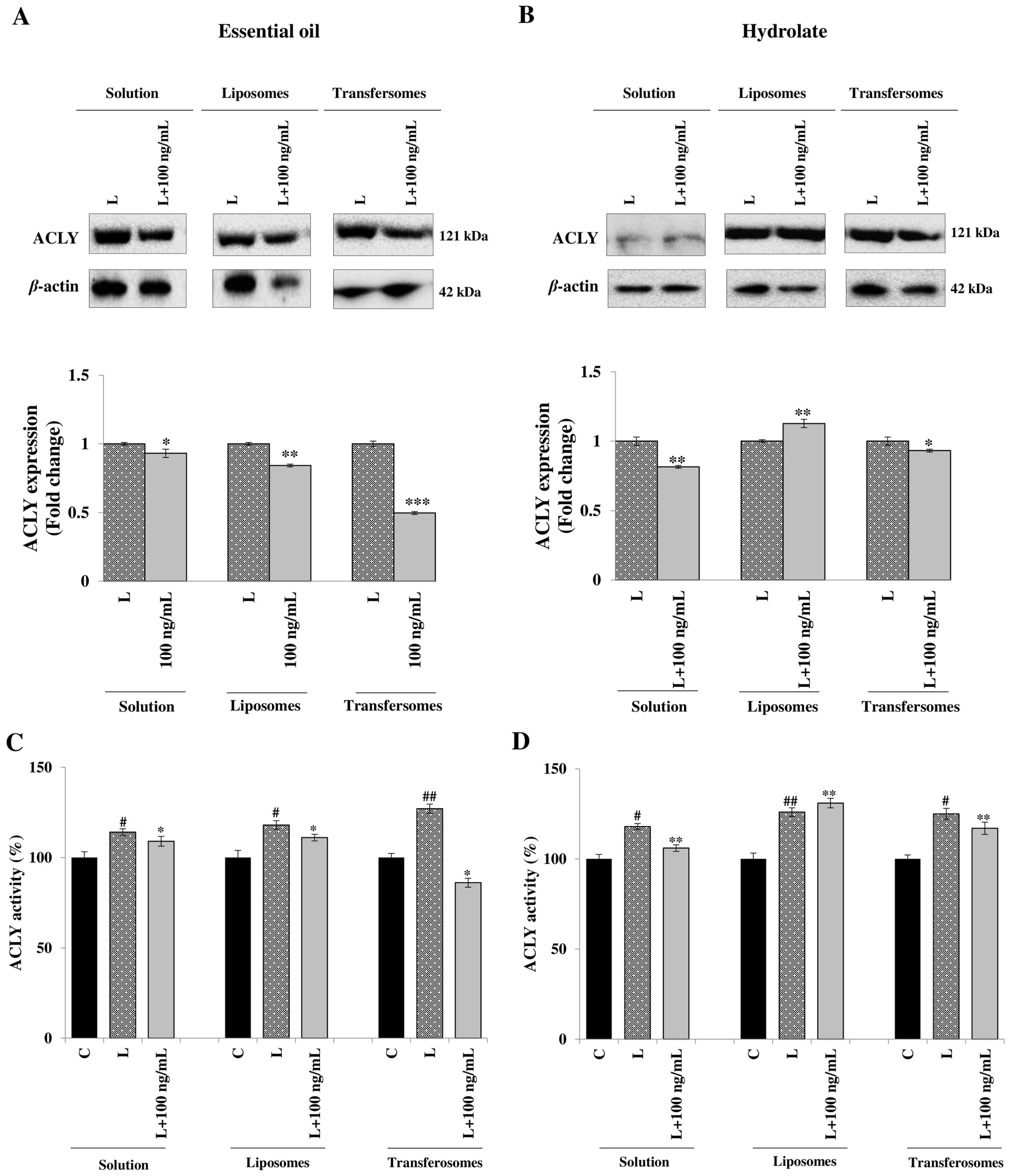
3.4. Effect on ATP Citrate Lyase Expression and Activity
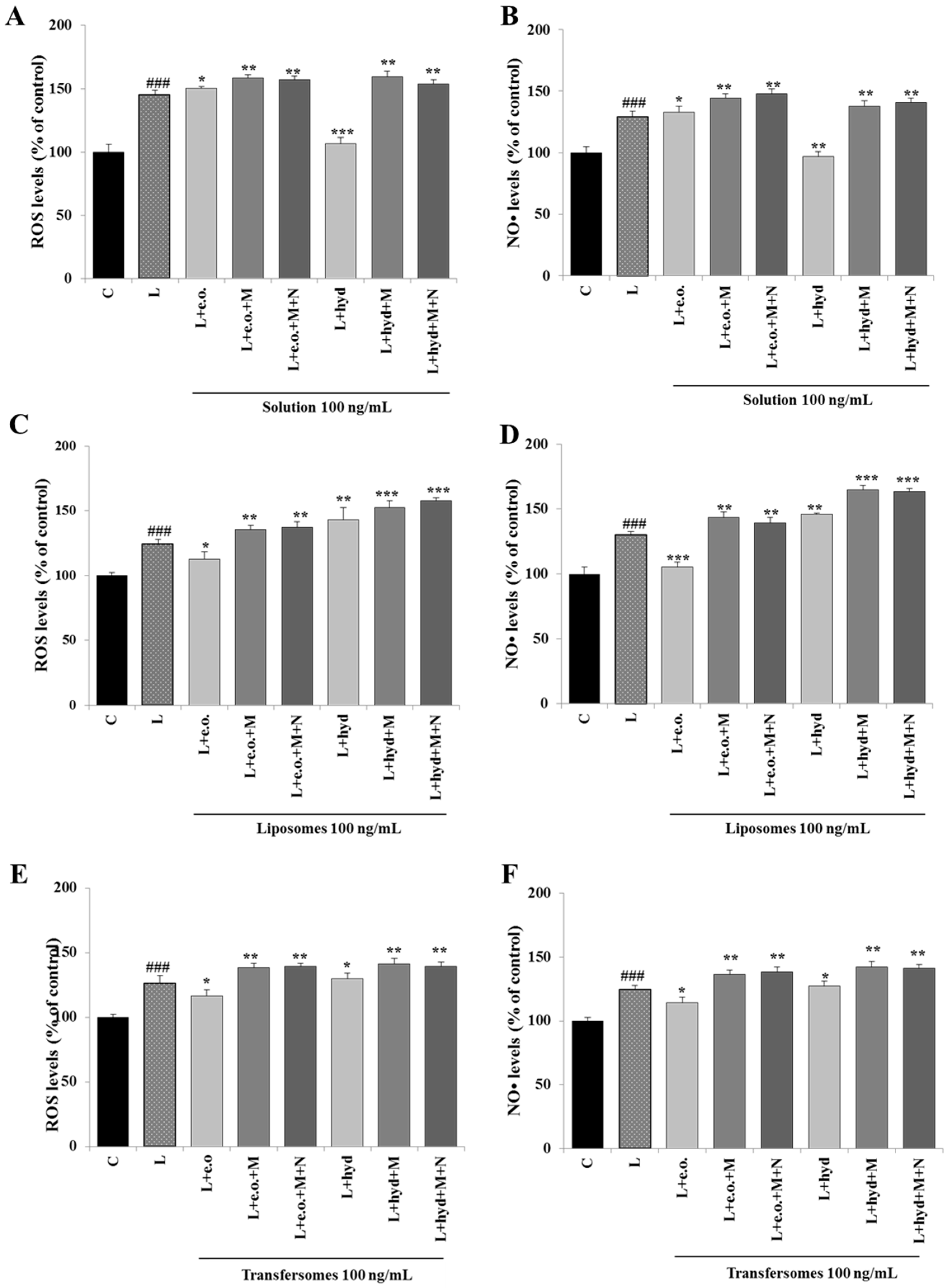
3.5. Effect on ROS and NO• Production
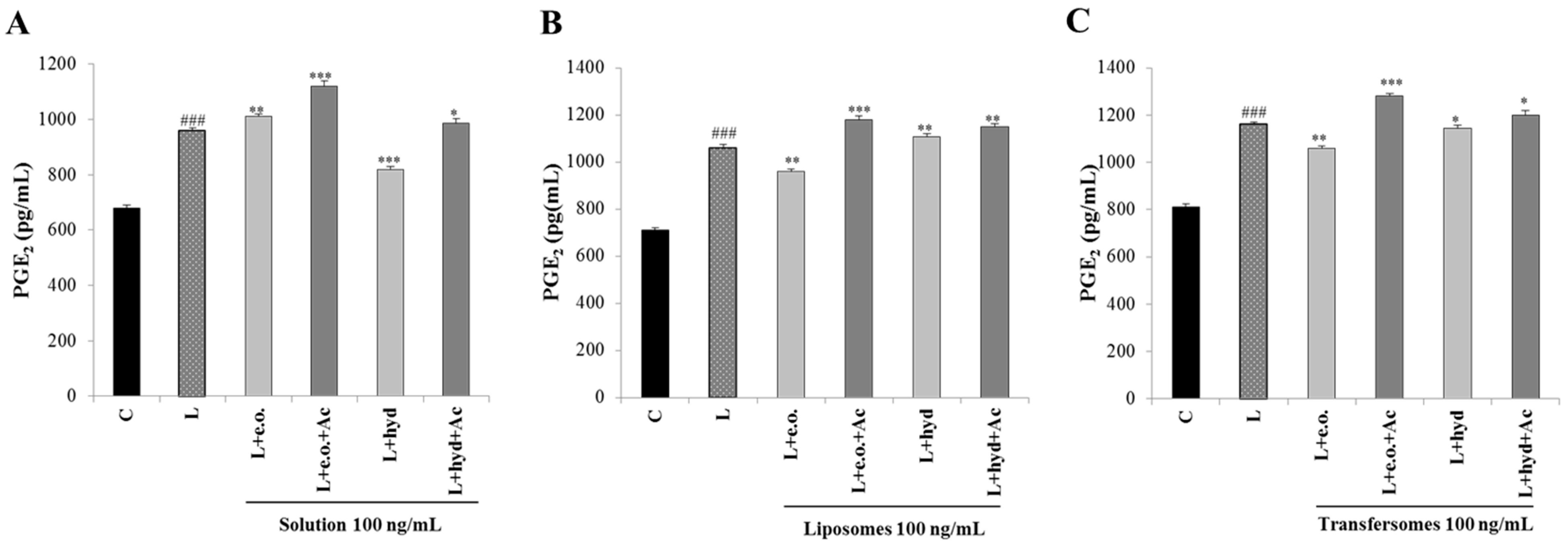
3.6. Effect on PGE2 Secretion
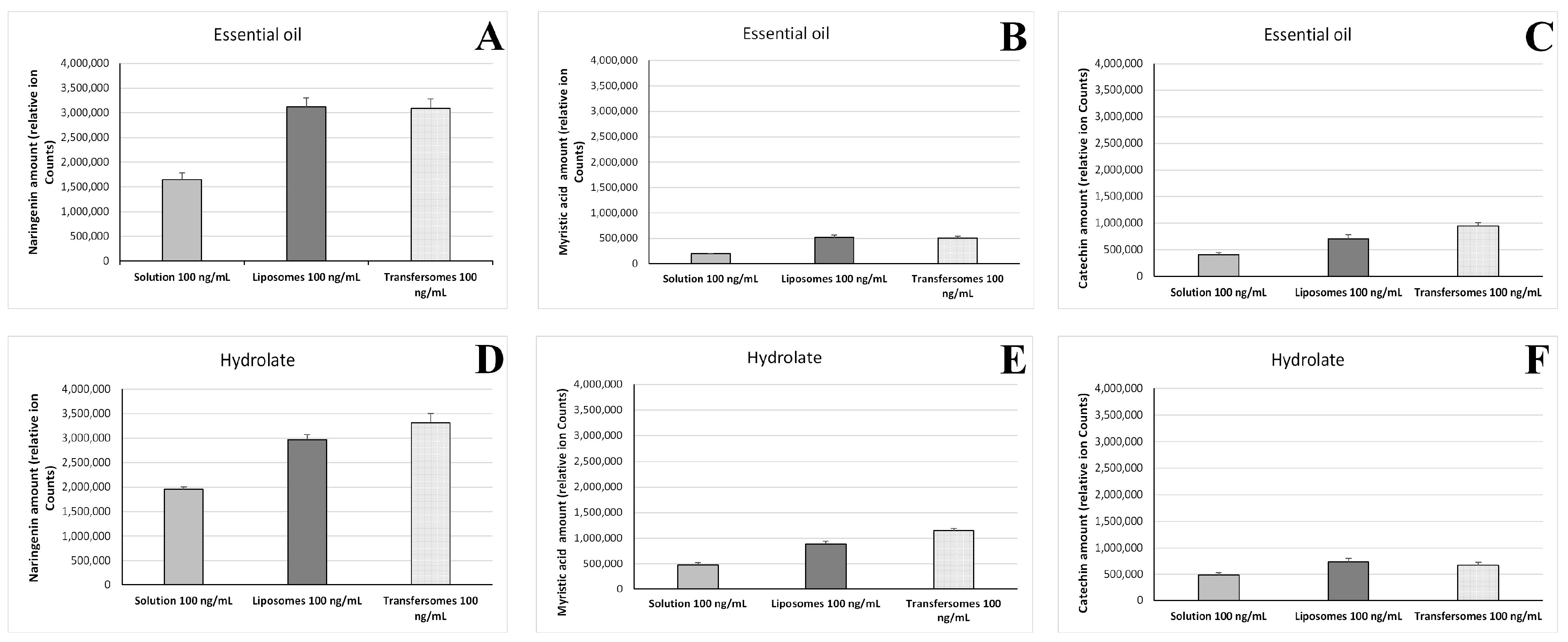
3.7. Intracellular Accumulation of P. vera Extracts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bulló, M.; Juanola-Falgarona, M.; Hernández-Alonso, P.; Salas-Salvadó, J. Nutrition attributes and health effects of pistachio nuts. Br. J. Nutr. 2015, 113 (Suppl. S2), S79–S93. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Barreca, D.; Gervasi, T.; Roussell, M.A.; Klein, B.; Feeney, M.J.; Carughi, A. Pistachio nuts (Pistacia vera L.): Production, nutrients, bioactives and novel health effects. Plants 2021, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Gentile, C.; Tesoriere, L.; Butera, D.; Fazzari, M.; Monastero, M.; Allegra, M.; Livrea, M.A. Antioxidant activity of sicilian pistachio (Pistacia vera L. var. Bronte) nut extract and its bioactive components. J. Agric. Food Chem. 2007, 55, 643–648. [Google Scholar] [CrossRef]
- Yeh, T.S.; Yuan, C.; Ascherio, A.; Rosner, B.A.; Willett, W.C.; Blacker, D. Long-term dietary flavonoid Intake and subjective cognitive decline in US men and women. Neurology 2021, 97, e1041–e1056. [Google Scholar] [CrossRef]
- Özçelik, B.; Aslan, M.; Orhan, I.; Karaoglu, T. Antibacterial, antifungal, and antiviral activities of the lipophylic extracts of Pistacia vera. Microbiol. Res. 2005, 160, 159–164. [Google Scholar] [CrossRef]
- Paterniti, I.; Impellizzeri, D.; Cordaro, M.; Siracusa, R.; Bisignano, C.; Gugliandolo, E.; Carughi, A.; Esposito, E.; Mandalari, G.; Cuzzocrea, S. The anti-inflammatory and antioxidant potential of pistachios (Pistacia vera L.) in vitro and in vivo. Nutrients 2017, 9, 915. [Google Scholar] [CrossRef]
- Sarkhail, P.; Navidpour, L.; Rahimifard, M.; Hosseini, N.M.; Souri, E. Bioassay-guided fractionation and identification of wound healing active compound from Pistacia vera L. hull extract. J. Ethnopharmacol. 2020, 248, 112335. [Google Scholar] [CrossRef]
- Pålsson-McDermott, E.M.; O’Neill, L.A.J. Targeting immunometabolism as an anti-inflammatory strategy. Cell. Res. 2020, 30, 300–314. [Google Scholar] [CrossRef]
- Infantino, V.; Pierri, C.L.; Iacobazzi, V. Metabolic routes in inflammation: The citrate pathway and its potential as therapeutic target. Curr. Med. Chem. 2019, 26, 7104–7116. [Google Scholar] [CrossRef]
- Santarsiero, A.; Convertini, P.; Todisco, S.; Pierri, C.L.; De Grassi, A.; Williams, N.C.; Iacobazzi, D.; De Stefano, G.; O’Neill, L.A.J.; Infantino, V. Acly nuclear translocation in human macrophages drives proinflammatory gene expression by Nf-Kb acetylation. Cells 2021, 10, 2962. [Google Scholar] [CrossRef]
- Santarsiero, A.; Convertini, P.; Vassallo, A.; Santoro, V.; Todisco, S.; Iacobazzi, D.; Fondufe-Mittendorf, Y.; Martelli, G.; de Oliveira, M.R.; Montanaro, R.; et al. Phenolic compounds of red wine aglianico del vulture modulate the functional activity of macrophages via Inhibition of NF-κB and the citrate pathway. Oxid. Med. Cell. Longev. 2021, 2021, 1–15. [Google Scholar] [CrossRef]
- Infantino, V.; Iacobazzi, V.; Palmieri, F.; Menga, A. ATP-citrate lyase is essential for macrophage inflammatory response. Biochem. Biophys. Res. Commun. 2013, 440, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Convertini, P.; Menga, A.; Andria, G.; Scala, I.; Santarsiero, A.; Castiglione Morelli, M.A.; Iacobazzi, V.; Infantino, V. The contribution of the citrate pathway to oxidative stress in Down syndrome. Immunology 2016, 149, 423–431. [Google Scholar] [CrossRef]
- Santarsiero, A.; Leccese, P.; Convertini, P.; Padula, A.; Abriola, P.; D’Angelo, S.; Bisaccia, F.; Infantino, V. New insights into behçet’s syndrome metabolic reprogramming: Citrate pathway dysregulation. Mediat. Inflamm. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Yan, H.; Pang, S.; Ya, M.; Qiu, F.; Qin, P.; Zeng, C.; Lu, Y. Liposomes as multifunctional nano-carriers for medicinal natural products. Front. Chem. 2022, 10, 963004. [Google Scholar] [CrossRef] [PubMed]
- Rahman, H.S.; Othman, H.H.; Hammadi, N.I.; Yeap, S.K.; Amin, K.M.; Abdul Samad, N.; Alitheen, N.B. Novel drug delivery systems for loading of natural plant extracts and their biomedical applications. Int. J. Nanomed. 2020, 15, 2439–2483. [Google Scholar] [CrossRef]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass. Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef]
- Vassallo, A.; Armentano, M.F.; Miglionico, R.; Caddeo, C.; Chirollo, C.; Gualtieri, M.J.; Ostuni, A.; Bisaccia, F.; Faraone, I.; Milella, L. Hura crepitans L. extract: Phytochemical characterization, antioxidant activity, and nanoformulation. Pharmaceutics 2020, 12, 553. [Google Scholar] [CrossRef]
- Aversano, R.; Contaldi, F.; Adelfi, M.G.; D’Amelia, V.; Diretto, G.; De Tommasi, N.; Vaccaro, C.; Vassallo, A.; Carputo, D. Comparative metabolite and genome analysis of tuber-bearing potato species. Phytochemistry 2017, 137, 42–51. [Google Scholar] [CrossRef]
- Mariconda, A.; Vassallo, A.; Bonomo, M.G.; Calabrone, L.; Salzano, G.; Claps, M.; Sinicropi, M.S.; Capasso, A.; Saturnino, C. Herbal formulations of Thymus serpyllum L. and Hypericum perforatum L. from southern Italy: Preparation and chemical characterization. Pharmacologyonline 2020, 1, 1–10. [Google Scholar]
- Arena, K.; Cacciola, F.; Mangraviti, D.; Zoccali, M.; Rigano, F.; Marino, N.; Dugo, P.; Mondello, L. Determination of the polyphenolic fraction of Pistacia vera L. kernel extracts by comprehensive two-dimensional liquid chromatography coupled to mass spectrometry detection. Anal. Bioanal. Chem. 2019, 411, 4819–4829. [Google Scholar] [CrossRef] [PubMed]
- Sonmezdag, A.S.; Kelebek, H.; Selli, S. Characterization and comparative evaluation of volatile, phenolic and antioxidant properties of pistachio (Pistacia vera L.) hull. J. Essent. Oil Res. 2017, 29, 262–270. [Google Scholar] [CrossRef]
- Huang, H.S.; Yu, H.S.; Yen, C.H.; Liaw, E.T. Multiple-reaction monitoring tandem mass method for determination of phenolics and water-soluble vitamins in Eccoilopus formosanus. Molecules 2020, 25, 3632. [Google Scholar] [CrossRef]
- Infantino, V.; Dituri, F.; Convertini, P.; Santarsiero, A.; Palmieri, F.; Todisco, S.; Mancarella, S.; Giannelli, G.; Iacobazzi, V. Epigenetic upregulation and functional role of the mitochondrial aspartate/glutamate carrier isoform 1 in hepatocellular carcinoma. Biochim. Biophys. Acta. Mol. Basis. Dis. 2019, 1865, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Santarsiero, A.; Onzo, A.; Pascale, R.; Acquavia, M.A.; Coviello, M.; Convertini, P.; Todisco, S.; Marsico, M.; Pifano, C.; Iannece, P.; et al. Pistacia lentiscus hydrosol: Untargeted metabolomic analysis and anti-inflammatory activity mediated by NF-κB and the citrate pathway. Oxid. Med. Cell. Long. 2020, 2020, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Takeda, Y.; Suzuki, F.; Inoue, H. [27] ATP citrate lyase (citrate-cleavage enzyme). In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1969; Volume 13, pp. 153–160. [Google Scholar]
- Caddeo, C.; Miglionico, R.; Rinaldi, R.; Nigro, I.; Lamorte, D.; Chiummiento, L.; Lupattelli, P.; Funicello, M.; D’Orsi, R.; Valenti, D.; et al. PEGylated liposomes loaded with carbamate inhibitor ANP0903 trigger apoptosis by enhancing ER stress in Hepg2 cancer cells. Int. J. Mol. Sci. 2023, 24, 4552. [Google Scholar] [CrossRef]
- Gültekin-Özgüven, M.; Davarcı, F.; Paslı, A.A.; Demir, N.; Özçelik, B. Determination of phenolic compounds by ultra high liquid chromatography-tandem mass spectrometry: Applications in nuts. LWT 2015, 64, 42–49. [Google Scholar] [CrossRef]
- Cardullo, N.; Leanza, M.; Muccilli, V.; Tringali, C. Valorization of agri-food waste from pistachio hard shells: Extraction of polyphenols as natural antioxidants. Resources 2021, 10, 45. [Google Scholar] [CrossRef]
- Fabani, M.P.; Luna, L.; Baroni, M.V.; Monferran, M.V.; Ighani, M.; Tapia, A.; Wunderlin, D.A.; Feresin, G.E. Pistachio (Pistacia vera var Kerman) from argentinean cultivars. A natural product with potential to improve human health. J. Funct. Foods 2013, 5, 1347–1356. [Google Scholar] [CrossRef]
- Abdolshahi, A.; Majd, M.H.; Rad, J.S.; Taheri, M.; Shabani, A.; Teixeira da Silva, J.A. Choice of solvent extraction technique affects fatty acid composition of pistachio (Pistacia vera L.) oil. J. Food Sci. Technol. 2015, 52, 2422–2427. [Google Scholar] [CrossRef]
- Garavand, F.; Madadlou, A.; Moini, S. Determination of phenolic profile and antioxidant activity of pistachio hull using high-performance liquid chromatography–diode array detector–electro-spray ionization–mass spectrometry as affected by ultrasound and microwave. Int. J. Food Prop. 2017, 20, 19–29. [Google Scholar] [CrossRef]
- Harandi, H.; Falahati-Pour, S.K.; Mahmoodi, M.; Faramarz, S.; Maleki, H.; Nasab, F.B.; Shiri, H.; Fooladi, S.; Nematollahi, M.H. Nanoliposomal formulation of pistachio hull extract: Preparation, characterization and anti-cancer evaluation through Bax/Bcl2 modulation. Mol. Biol. Rep. 2022, 49, 2735–2743. [Google Scholar] [CrossRef] [PubMed]
- Todisco, S.; Convertini, P.; Iacobazzi, V.; Infantino, V. TCA cycle rewiring as emerging metabolic signature of hepatocellular carcinoma. Cancers 2020, 12, 68. [Google Scholar] [CrossRef] [PubMed]
- Infantino, V.; Convertini, P.; Cucci, L.; Panaro, M.A.; Di Noia, M.A.; Calvello, R.; Palmieri, F.; Iacobazzi, V. The mitochondrial citrate carrier: A new player in inflammation. Biochem. J. 2011, 438, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Infantino, V.; Iacobazzi, V.; Menga, A.; Avantaggiati, M.L.; Palmieri, F. A key role of the mitochondrial citrate carrier (SLC25A1) in TNFα- and IFNγ-triggered inflammation. Biochim. Biophys. Acta 2014, 1839, 1217–1225. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P90G | P. vera Extract | H2O | |
|---|---|---|---|
| Empty liposomes | 90 mg/mL | - | 2 mL |
| P. vera liposomes | 90 mg/mL | 10 mg/mL | 2 mL |
| P90G | P. vera Extract | Tween 80 | H2O | |
|---|---|---|---|---|
| Empty transfersomes | 90 mg/mL | - | 5 mg/mL | 2 mL |
| P. vera transfersomes | 90 mg/mL | 10 mg/mL | 5 mg/mL | 2 mL |
| Peak No. | Compound | Rt (min) | Molecular Formula | Molecular Weight | [M+H]+ | Δ Ppm | MS/MS |
|---|---|---|---|---|---|---|---|
| 1 | Vanillic acid | 2.06 | C8H8O4 | 168.18 | 169 | 0.18 | 142, 97 |
| 2 | p-coumaric acid | 2.52 | C9H8O3 | 164.05 | 165 | −0.31 | 147, 82 |
| 3 | Sinapinic acid | 3.60 | C11H12O5 | 224.07 | 225 | 0.71 | 207 |
| 4 | Myristic acid | 3.73 | C14H28O2 | 228.21 | 229 | −0.21 | 225, 191 |
| 5 | Caffeic acid | 14.57 | C9H8O4 | 180.04 | 181 | 0.41 | 181, 163 |
| 6 | Catechin | 16.95 | C15H14O6 | 290.08 | 291 | 0.38 | 245, 151, 109 |
| 7 | α-pinene | 17.49 | C10H16 | 136.24 | 137 | 0.24 | 109, 95, 81 |
| 8 | Ascorbic acid | 17.96 | C6H8O6 | 176.04 | 177 | −0.43 | 177, 159 |
| 9 | Tryptophan | 17.99 | C11H12N2O2 | 204.09 | 205 | 0.39 | 205, 144 |
| 10 | Naringenin | 18.36 | C15H12O5 | 272.07 | 273 | 0.57 | 255, 244,95 |
| 11 | Cinnamic acid | 18.41 | C9H8O2 | 148.05 | 149 | −0.34 | 131, 121, 65 |
| 12 | Luteolin | 18.83 | C15H10O6 | 286.24 | 287 | 0.24 | 269, 243, 135 |
| 13 | Ferulic acid | 20.33 | C10H10O4 | 194.06 | 195 | −0.26 | 177 |
| 14 | Tyrosine | 26.88 | C9H11NO3 | 181.07 | 182 | 0.36 | 182, 165, 136 |
| Peak No. | Compound | Rt (min) | Molecular Formula | Molecular Weight | [M+H]+ | Δ Ppm | MS/MS |
|---|---|---|---|---|---|---|---|
| 1 | Vanillic acid | 1.99 | C8H8O4 | 168.18 | 169 | 0.17 | 142, 97 |
| 2 | p-coumaric acid | 2.50 | C9H8O3 | 164.05 | 165 | 0.52 | 147, 82 |
| 3 | Sinapinic acid | 3.61 | C11H12O5 | 224.07 | 225 | 0.73 | 207 |
| 4 | Myristic acid | 3.72 | C14H28O2 | 228.21 | 229 | −0.22 | 225, 191 |
| 5 | Caffeic acid | 15.56 | C9H8O4 | 180.04 | 181 | 0.46 | 181, 163 |
| 6 | Catechin | 16.96 | C15H14O6 | 290.08 | 291 | 0.22 | 245, 151, 109 |
| 7 | Ascorbic acid | 17.80 | C6H8O6 | 176.04 | 177 | −0.46 | 177, 159 |
| 8 | Tryptophan | 17.99 | C11H12N2O2 | 204.09 | 205 | 0.29 | 205, 144 |
| 9 | Naringenin | 18.39 | C15H12O5 | 272.07 | 273 | 0.61 | 255, 244 |
| 10 | Cinnamic acid | 18.47 | C9H8O2 | 148.05 | 149 | −0.57 | 131, 121, 65 |
| 11 | Luteolin | 18.22 | C15H10O6 | 286.24 | 287 | 0.26 | 269, 135 |
| 12 | Ferulic acid | 20.33 | C10H10O4 | 194.06 | 195 | −0.62 | 177 |
| 13 | Tyrosine | 26.63 | C9H11NO3 | 181.07 | 182 | 0.23 | 182, 165, 136 |
| Peak No. | Compound | % Area |
|---|---|---|
| 1 | tricyclene | 4.68 |
| 2 | α-pinene | 40.44 |
| 3 | camphene | 15.89 |
| 4 | β-pinene | 9.52 |
| 5 | 3-carene | 1.23 |
| 6 | p-cymene | 0.25 |
| 7 | limonene | 2.24 |
| 8 | bornyl acetate | 15.53 |
| MD (nm ± SD) | PI ± SD | ZP (mV ± SD) | |
|---|---|---|---|
| Empty liposomes | 85 ± 4.3 | 0.30 ± 0.03 | −12 ± 2.8 |
| P. vera essential oil liposomes | ** 76 ± 6.5 | ** 0.19 ± 0.03 | −11 ± 2.6 |
| P. vera hydrolate liposomes | 82 ± 6.9 | 0.28 ± 0.01 | −11 ± 1.8 |
| Empty transfersomes | ** 95 ± 6.1 | ** 0.24 ± 0.03 | * −15 ± 2.1 |
| P. vera essential oil transfersomes | ## 90 ± 9.6 | ° 0.20 ± 0.04 | −13 ± 3.4 |
| P. vera hydrolate transfersomes | °° 85 ± 5.1 | •• 0.23 ± 0.01 | •• −15 ± 2.8 |
| Compound. | Formulation | EE % ± SD |
|---|---|---|
| Naringenin | P. vera essential oil liposomes | 93 + 0.9 |
| P. vera hydrolate liposomes | 94 + 1.5 | |
| P. vera essential oil transfersomes | 89 + 3.1 | |
| P. vera hydrolate transfersomes | 92 + 2.7 | |
| Myristic acid | P. vera essential oil liposomes | 85 + 4.3 |
| P. vera hydrolate liposomes | 84 + 1.9 | |
| P. vera essential oil transfersomes | 85 + 4.8 | |
| P. vera hydrolate transfersomes | 83 + 2.0 | |
| Catechin | P. vera essential oil liposomes | 86 + 3.9 |
| P. vera hydrolate liposomes | 83 + 1.4 | |
| P. vera essential oil transfersomes | 82 + 2.9 | |
| P. vera hydrolate transfersomes | 84 + 3.0 |
| Time | MD (nm ± SD) | PI ± SD | ZP (mV ± SD) | |
|---|---|---|---|---|
| Empty transfersomes | 30 days | 102 ± 3.0 | 0.26 ± 0.03 | −14 ± 2.1 |
| P. vera essential oil transfersomes | 101 ± 7.9 | 0.18 ± 0.01 | −13 ± 1.8 | |
| P. vera hydrolate transfersomes | 89 ± 8.1 | 0.25 ± 0.01 | −14 ± 1.1 | |
| Empty transfersomes | 60 days | 101 ± 0.5 | 0.21 ± 0.03 | −11 ± 1.3 |
| P. vera essential oil transfersomes | 101 ± 5.0 | 0.17 ± 0.03 | −14 ± 1.8 | |
| P. vera hydrolate transfersomes | 91 ± 4.4 | 0.25 ± 0.05 | −10 ± 5.0 | |
| Empty transfersomes | 90 days | 101 ± 4.9 | 0.22 ± 0.02 | −9 ± 2.6 |
| P. vera essential oil transfersomes | 100 ± 7.9 | 0.18 ± 0.01 | −13 ± 1.6 | |
| P. vera hydrolate transfersomes | 96 ± 8.1 | 0.26 ± 0.02 | −11 ± 4.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pani, S.; Pappalardo, I.; Santarsiero, A.; Vassallo, A.; Radice, R.P.; Martelli, G.; Siano, F.; Todisco, S.; Convertini, P.; Caddeo, C.; et al. Immunometabolism Modulation by Extracts from Pistachio Stalks Formulated in Phospholipid Vesicles. Pharmaceutics 2023, 15, 1540. https://doi.org/10.3390/pharmaceutics15051540
Pani S, Pappalardo I, Santarsiero A, Vassallo A, Radice RP, Martelli G, Siano F, Todisco S, Convertini P, Caddeo C, et al. Immunometabolism Modulation by Extracts from Pistachio Stalks Formulated in Phospholipid Vesicles. Pharmaceutics. 2023; 15(5):1540. https://doi.org/10.3390/pharmaceutics15051540
Chicago/Turabian StylePani, Simone, Ilaria Pappalardo, Anna Santarsiero, Antonio Vassallo, Rosa Paola Radice, Giuseppe Martelli, Francesco Siano, Simona Todisco, Paolo Convertini, Carla Caddeo, and et al. 2023. "Immunometabolism Modulation by Extracts from Pistachio Stalks Formulated in Phospholipid Vesicles" Pharmaceutics 15, no. 5: 1540. https://doi.org/10.3390/pharmaceutics15051540