
Cationic Microbubbles for Non-Selective Binding of Cavitation Nuclei to Bacterial Biofilms


Abstract
:1. Introduction
2. Materials and Methods
2.1. Production of MBs
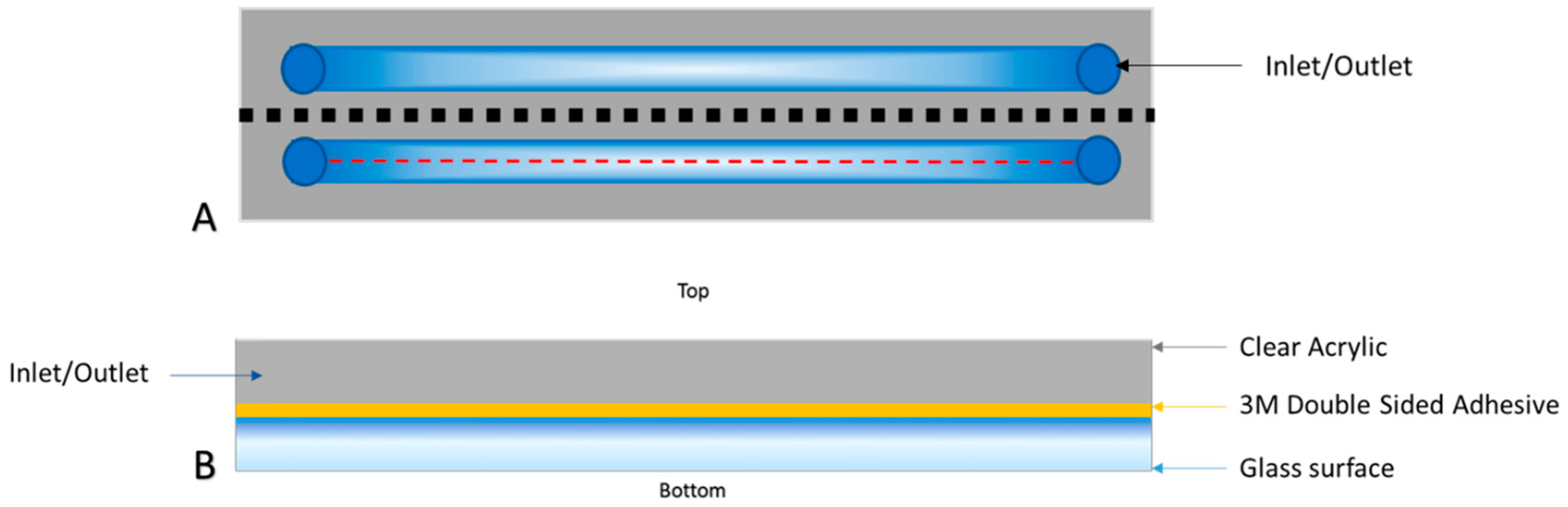
2.2. Development of Microfluidic Flow Cells for Creating Surfaces with Augmented Electrostatic Charge
2.3. Assessment of MB Charge
2.4. Determination of MB Binding Characteristics under Static Conditions
2.5. Assessment of MB Non-Selective Binding Strength under Flow
2.6. Assessment of Non-Selective Interaction of MBs with P. aeruginosa Biofilms
2.7. Statistical Analysis
3. Results
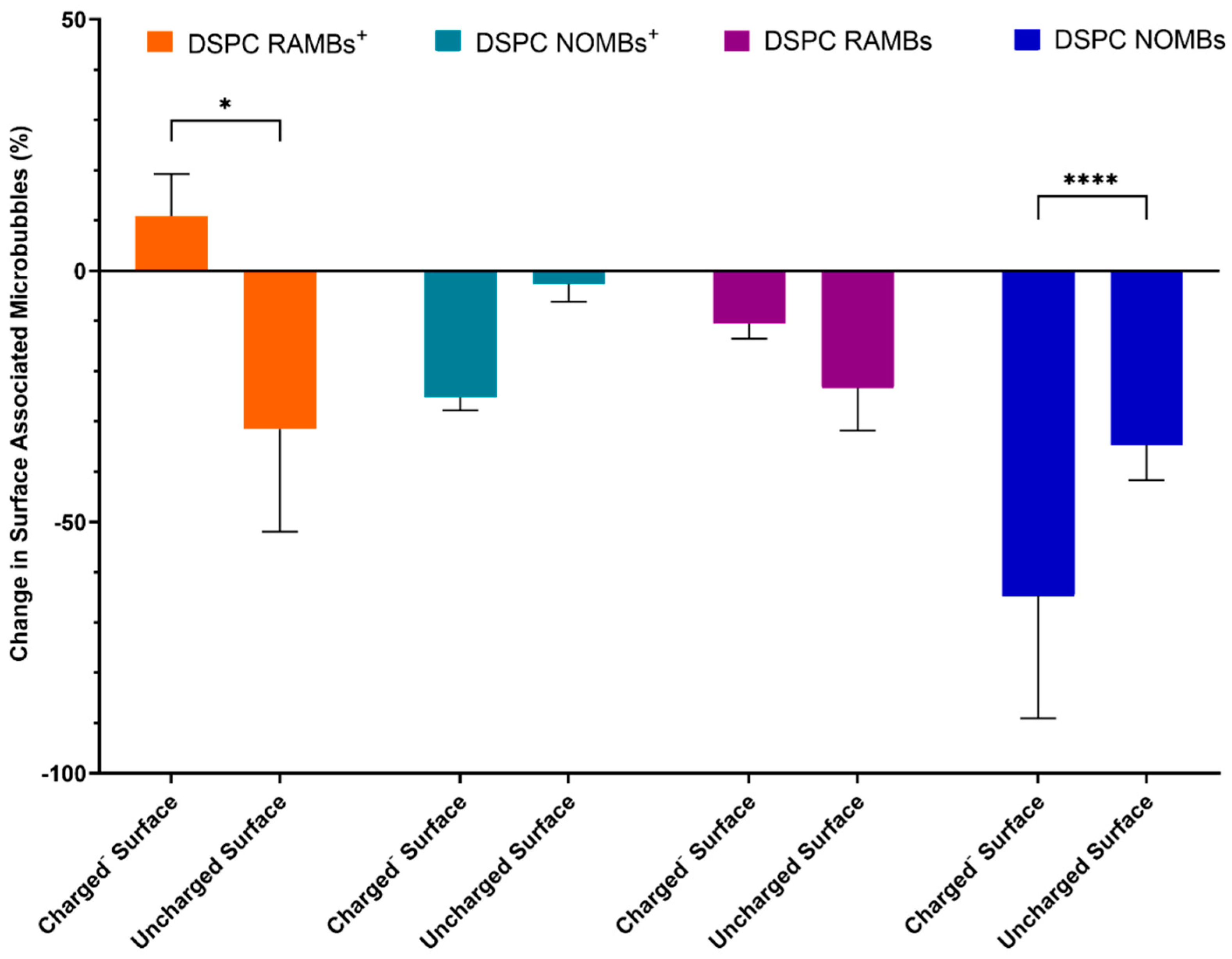
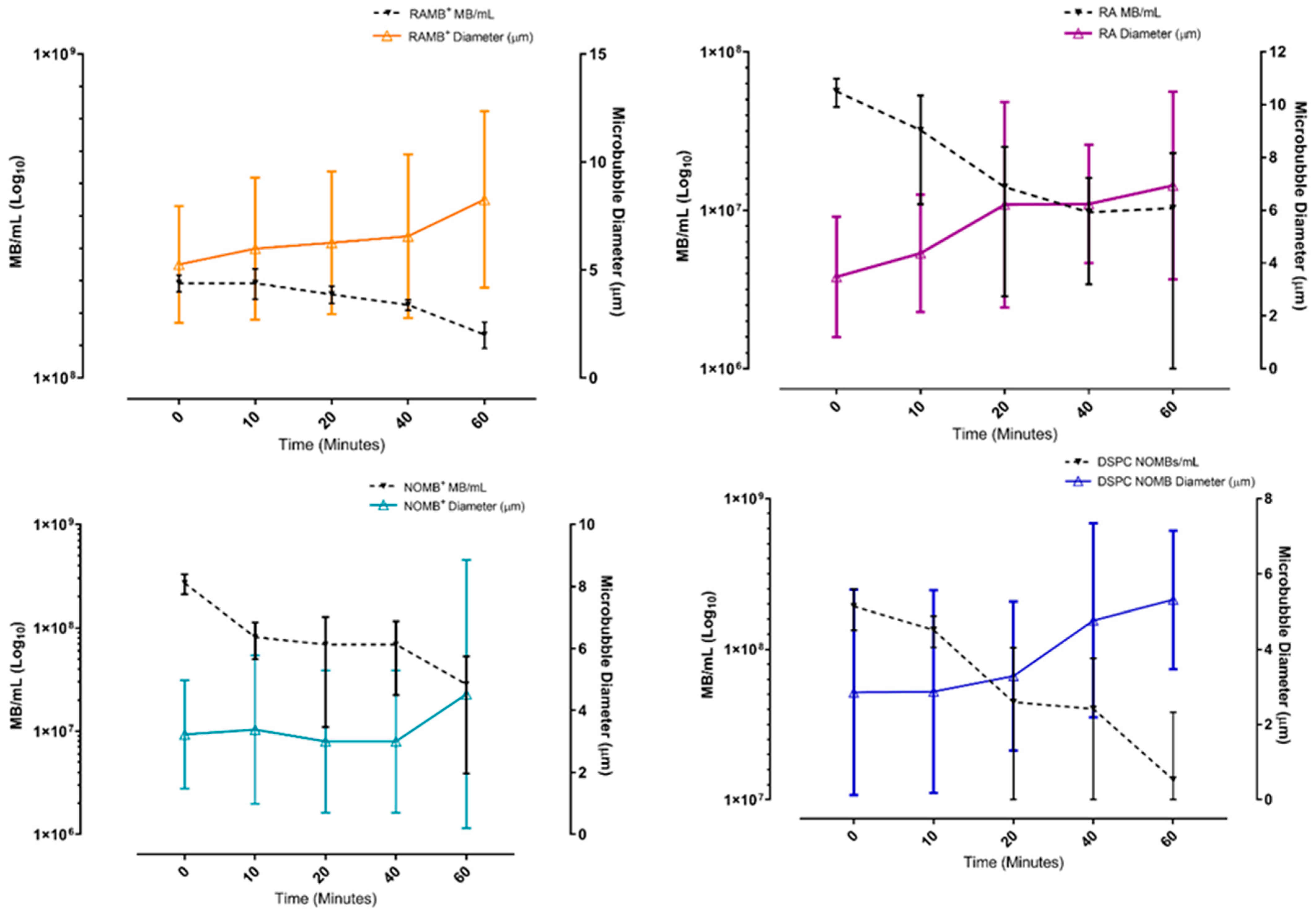
3.1. Microbubble Binding Characteristics in Static Conditions
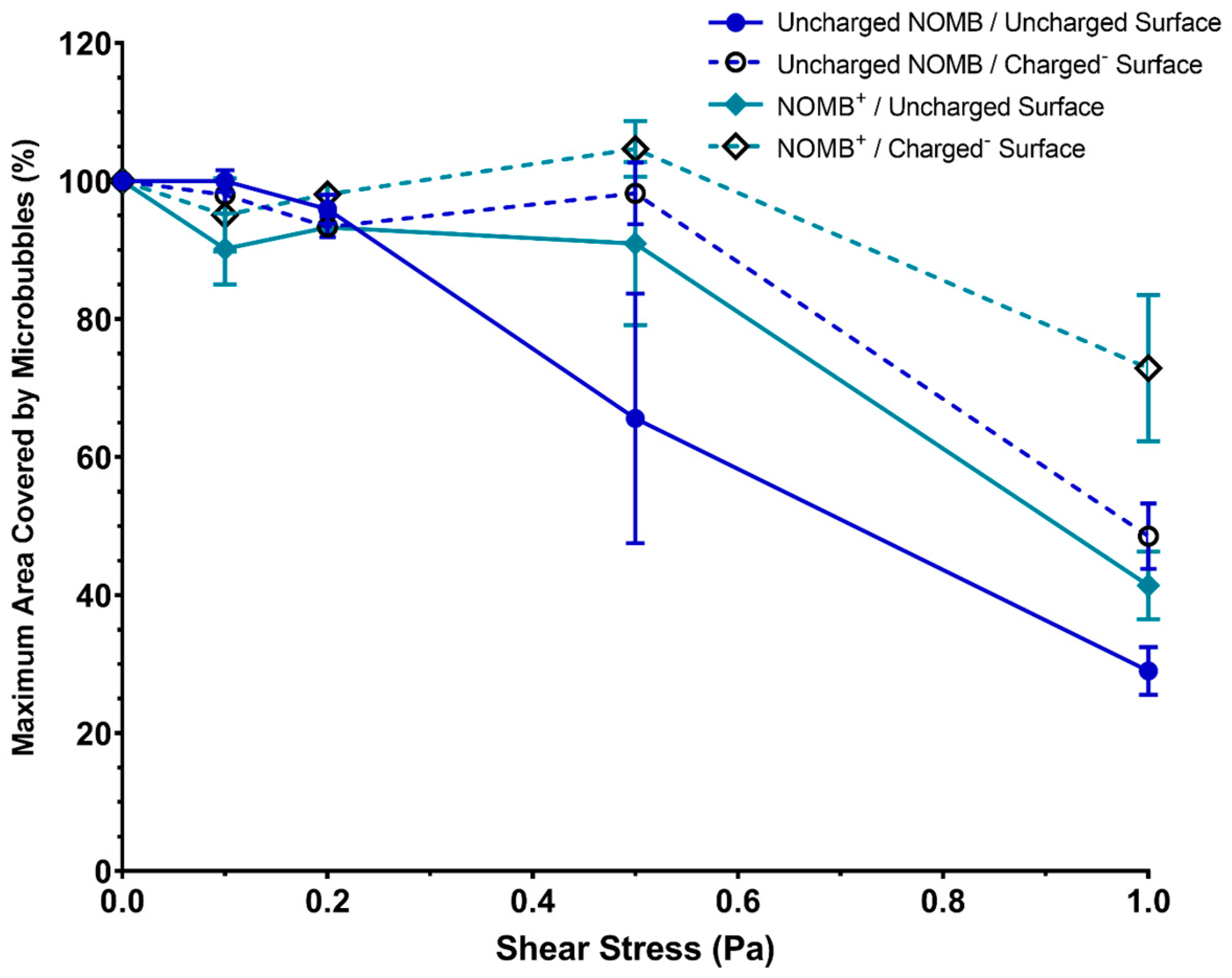
3.2. Assessment of MB Non-Selective Binding Strength in Dynamic Conditions
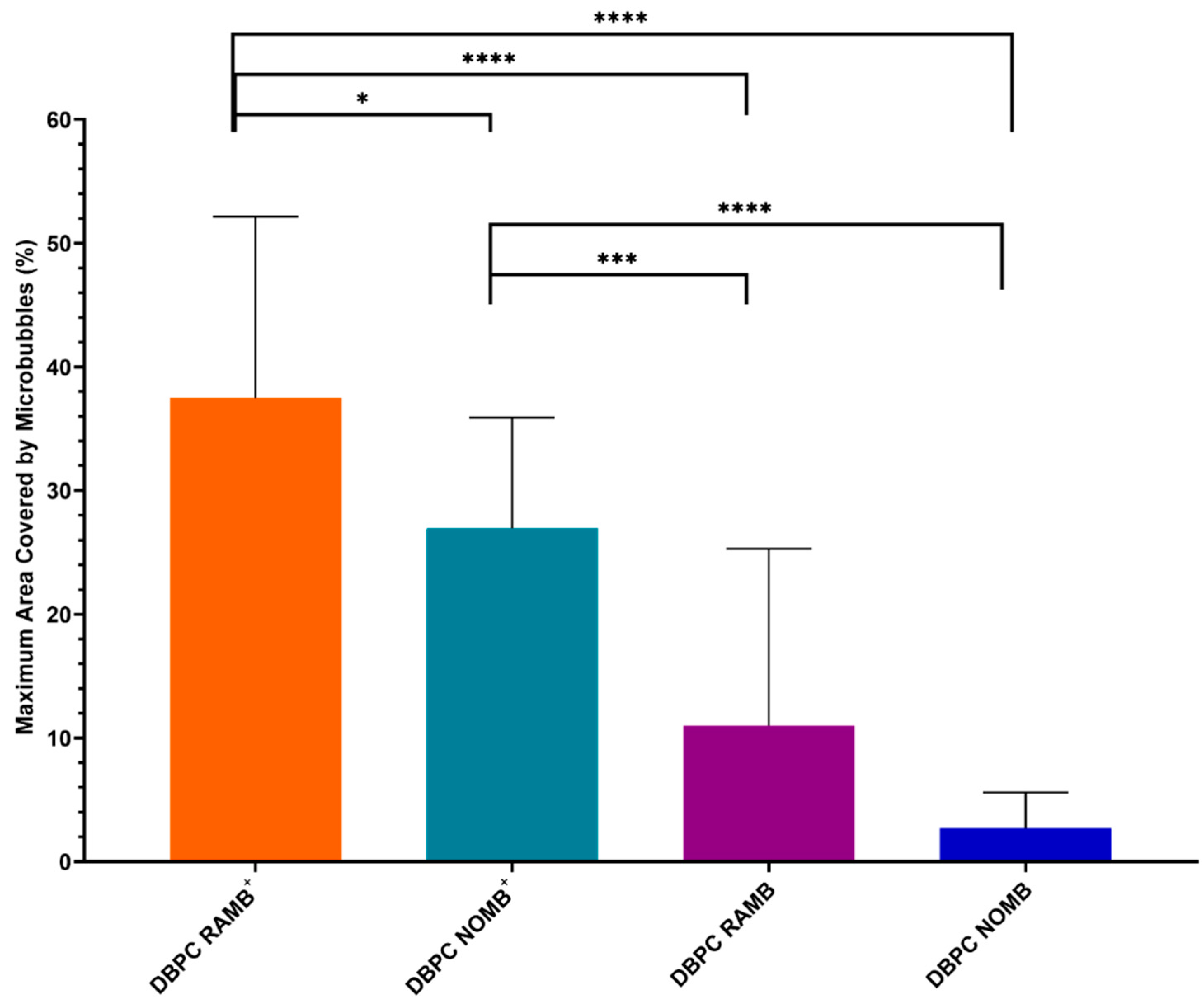
3.3. Assessment of MB Interaction with P. aeruginosa Biofilms
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
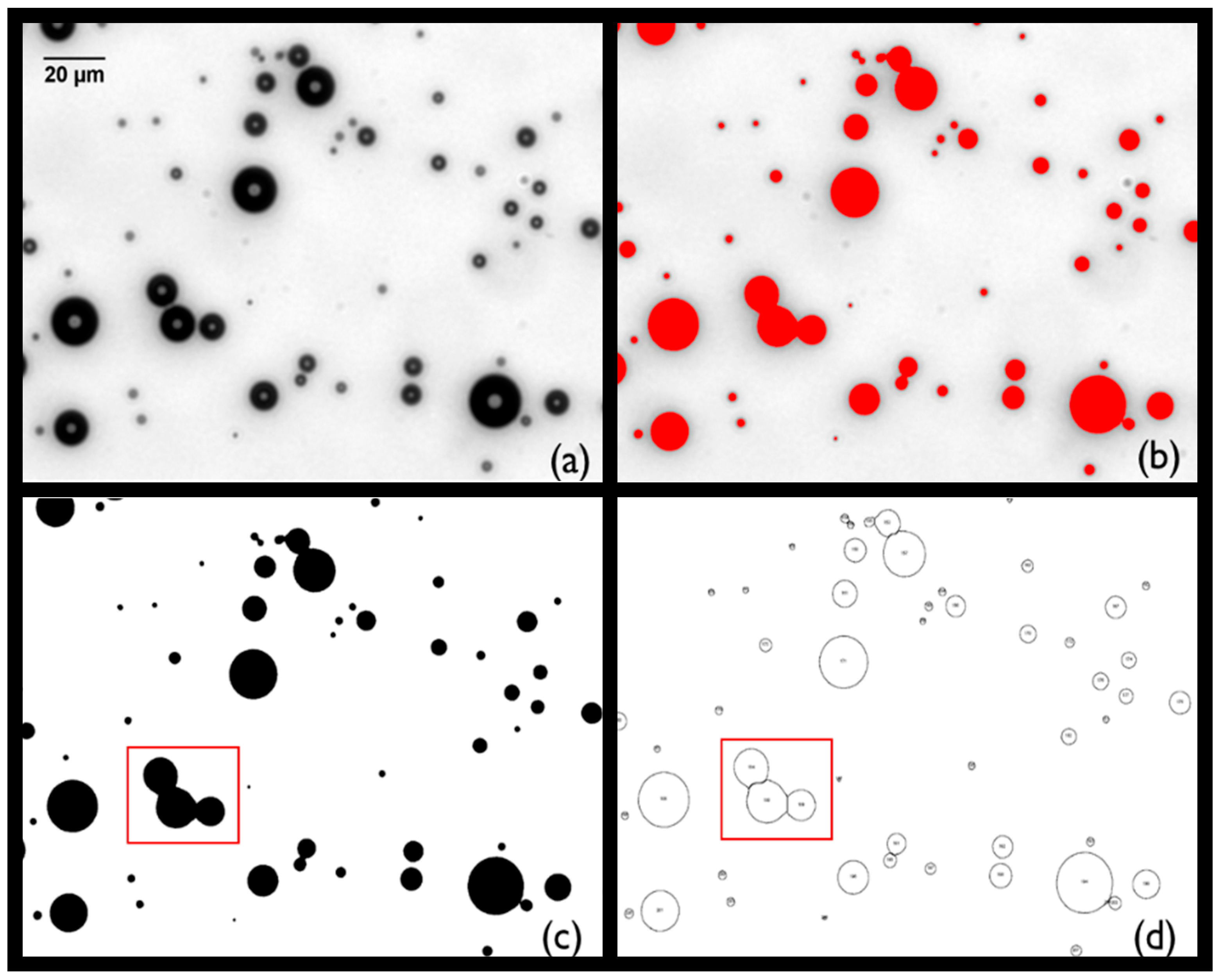
Appendix A.1. Characterisation of MBs Size and Concentration


Appendix A.2. Assessing Flow Cells for Leakage under Increasing Flow Rates

References
- Ciofu, O.; Rojo-Molinero, E.; Macià, M.D.; Oliver, A. Antibiotic Treatment of Biofilm Infections. APMIS 2017, 125, 304–319. [Google Scholar] [CrossRef]
- Carniello, V.; Peterson, B.W.; van der Mei, H.C.; Busscher, H.J. Physico-Chemistry from Initial Bacterial Adhesion to Surface-Programmed Biofilm Growth. Adv. Colloid. Interface Sci. 2018, 261, 1–14. [Google Scholar] [CrossRef]
- Cámara, M.; Green, W.; MacPhee, C.E.; Rakowska, P.D.; Raval, R.; Richardson, M.C.; Slater-Jefferies, J.; Steventon, K.; Webb, J.S. Economic Significance of Biofilms: A Multidisciplinary and Cross-Sectoral Challenge. NPJ Biofilms Microbiomes 2022, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, J. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations. 2014. Available online: https://amr-review.org/sites/default/files/AMR%20Review%20Paper%20-%20Tackling%20a%20crisis%20for%20the%20health%20and%20wealth%20of%20nations_1.pdf (accessed on 11 May 2023).
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus Biofilm: An Emerging Battleground in Microbial Communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Meiers, J.; Zahorska, E.; Röhrig, T.; Hauck, D.; Wagner, S.; Titz, A. Directing Drugs to Bugs: Antibiotic-Carbohydrate Conjugates Targeting Biofilm-Associated Lectins of Pseudomonas Aeruginosa. J. Med. Chem. 2020, 63, 11707–11724. [Google Scholar] [CrossRef] [PubMed]
- Palmioli, A.; Sperandeo, P.; Polissi, A.; Airoldi, C. Targeting Bacterial Biofilm: A New LecA Multivalent Ligand with Inhibitory Activity. ChemBioChem 2019, 20, 2911–2915. [Google Scholar] [CrossRef]
- Heta, S.; Robo, I. The Side Effects of the Most Commonly Used Group of Antibiotics in Periodontal Treatments. Med. Sci. 2018, 6, 6. [Google Scholar] [CrossRef]
- De Gunzburg, J.; Ghozlane, A.; Ducher, A.; Le Chatelier, E.; Duval, X.; Ruppé, E.; Armand-Lefevre, L.; Sablier-Gallis, F.; Burdet, C.; Alavoine, L.; et al. Protection of the Human Gut Microbiome from Antibiotics. J. Infect. Dis. 2018, 217, 628–636. [Google Scholar] [CrossRef]
- Sirsi, S.; Borden, M. Microbubble Compositions, Properties and Biomedical Applications. Bubble Sci. Eng. Technol. 2009, 1, 3–17. [Google Scholar] [CrossRef]
- Lattwein, K.R.; Shekhar, H.; Kouijzer, J.J.P.; van Wamel, W.J.B.; Holland, C.K.; Kooiman, K. Sonobactericide: An Emerging Treatment Strategy for Bacterial Infections. Ultrasound Med. Biol. 2020, 46, 193–215. [Google Scholar] [CrossRef]
- Unger, E.C.; Porter, T.; Culp, W.; Labell, R.; Matsunaga, T.; Zutshi, R. Therapeutic Applications of Lipid-Coated Microbubbles. Adv. Drug Deliv. Rev. 2004, 56, 1291–1314. [Google Scholar] [CrossRef] [PubMed]
- Kooiman, K.; Vos, H.J.; Versluis, M.; de Jong, N. Acoustic Behavior of Microbubbles and Implications for Drug Delivery. Adv. Drug Deliv. Rev. 2014, 72, 28–48. [Google Scholar] [CrossRef]
- Stride, E.; Saffari, N. Microbubble Ultrasound Contrast Agents: A Review. Proc. Inst. Mech. Eng. H 2003, 217, 429–447. [Google Scholar] [CrossRef]
- Ibsen, S.; Schutt, C.E.; Esener, S. Microbubble-Mediated Ultrasound Therapy: A Review of Its Potential in Cancer Treatment; Dove Press: Macclesfield, UK, 2013; Volume 7, pp. 375–388. [Google Scholar]
- Kooiman, K.; Roovers, S.; Langeveld, S.A.G.; Kleven, R.T.; Dewitte, H.; O’Reilly, M.A.; Escoffre, J.M.; Bouakaz, A.; Verweij, M.D.; Hynynen, K.; et al. Ultrasound-Responsive Cavitation Nuclei for Therapy and Drug Delivery. Ultrasound Med. Biol. 2020, 46, 1296–1325. [Google Scholar] [CrossRef]
- LuTheryn, G.; Glynne-Jones, P.; Webb, J.S.; Carugo, D. Ultrasound-mediated Therapies for the Treatment of Biofilms in Chronic Wounds: A Review of Present Knowledge. Microb. Biotechnol. 2019, 1751–7915, 13471. [Google Scholar] [CrossRef] [PubMed]
- Izadifar, Z.; Babyn, P.; Chapman, D. Ultrasound Cavitation/Microbubble Detection and Medical Applications. J. Med. Biol. Eng. 2018, 39, 259–276. [Google Scholar] [CrossRef]
- Martin, M.J.; Chung, E.M.L.; Goodall, A.H.; Martina, A.D.; Ramnarine, K.V.; Fan, L.; Hainsworth, S.V.; Naylor, A.R.; Evans, D.H. Enhanced Detection of Thromboemboli with the Use of Targeted Microbubbles. Stroke 2007, 38, 2726–2732. [Google Scholar] [CrossRef]
- Zhao, S.; Borden, M.; Bloch, S.H.; Kruse, D.E.; Ferrara, K.W.; Dayton, P.A. Increasing Binding Efficiency of Ultrasound Targeted Agents with Radiation Force. In Proceedings of the Proceedings—IEEE Ultrasonics Symposium, Montreal, QC, Canada, 23–27 August 2004; Volume 2, pp. 1114–1117. [Google Scholar]
- Weller, G.E.R.; Villanueva, F.S.; Klibanov, A.L.; Wagner, W.R. Modulating Targeted Adhesion of an Ultrasound Contrast Agent to Dysfunctional Endothelium. Ann. Biomed. Eng. 2002, 30, 1012–1019. [Google Scholar] [CrossRef]
- Owen, J.; Rademeyer, P.; Chung, D.; Cheng, Q.; Holroyd, D.; Coussios, C.; Friend, P.; Pankhurst, Q.A.; Stride, E. Magnetic Targeting of Microbubbles against Physiologically Relevant Flow Conditions. Interface Focus 2015, 5, 20150001. [Google Scholar] [CrossRef]
- Caudwell, J.A.; Tinkler, J.M.; Johnson, B.R.G.; McDowall, K.J.; Alsulaimani, F.; Tiede, C.; Tomlinson, D.C.; Freear, S.; Turnbull, W.B.; Evans, S.D.; et al. Protein-Conjugated Microbubbles for the Selective Targeting of S. Aureus Biofilms. Biofilm 2022, 4, 100074. [Google Scholar] [CrossRef]
- Sommer, R.; Wagner, S.; Rox, K.; Varrot, A.; Hauck, D.; Wamhoff, E.-C.; Schreiber, J.; Ryckmans, T.; Brunner, T.; Rademacher, C.; et al. Glycomimetic, Orally Bioavailable LecB Inhibitors Block Biofilm Formation of Pseudomonas Aeruginosa. J. Am. Chem. Soc. 2018, 140, 2537–2545. [Google Scholar] [CrossRef] [PubMed]
- Johansson, E.M.V.; Crusz, S.A.; Kolomiets, E.; Buts, L.; Kadam, R.U.; Cacciarini, M.; Bartels, K.M.; Diggle, S.P.; Cámara, M.; Williams, P.; et al. Inhibition and Dispersion of Pseudomonas Aeruginosa Biofilms by Glycopeptide Dendrimers Targeting the Fucose-Specific Lectin LecB. Chem. Biol. 2008, 15, 1249–1257. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.; Hauck, D.; Hoffmann, M.; Sommer, R.; Joachim, I.; Müller, R.; Imberty, A.; Varrot, A.; Titz, A. Covalent Lectin Inhibition and Application in Bacterial Biofilm Imaging. Angew. Chem. Int. Ed. 2017, 56, 16559–16564. [Google Scholar] [CrossRef] [PubMed]
- Takalkar, A.M.; Klibanov, A.L.; Rychak, J.J.; Lindner, J.R.; Ley, K. Binding and Detachment Dynamics of Microbubbles Targeted to P-Selectin under Controlled Shear Flow. J. Control Release 2004, 96, 473–482. [Google Scholar] [CrossRef]
- Renner, L.D.; Weibel, D.B. Physicochemical Regulation of Biofilm Formation. MRS Bull. 2011, 36, 347–355. [Google Scholar] [CrossRef]
- Harper, R.A.; Carpenter, G.H.; Proctor, G.B.; Harvey, R.D.; Gambogi, R.J.; Geonnotti, A.R.; Hider, R.; Jones, S.A. Diminishing Biofilm Resistance to Antimicrobial Nanomaterials through Electrolyte Screening of Electrostatic Interactions. Colloids. Surf. B Biointerfaces 2019, 173, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Khelissa, S.O.; Abdallah, M.; Jama, C.; Barras, A.; Chihib, N.E. Comparative Study on the Impact of Growth Conditions on the Physiology and the Virulence of Pseudomonas Aeruginosa Biofilm and Planktonic Cells. J. Food Prot. 2019, 82, 1357–1363. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Zhang, H.; Bai, R. Advances in Ultrasound-Targeted Microbubble-Mediated Gene Therapy for Liver Fibrosis. Acta Pharm. Sin. B 2017, 7, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Carugo, D.; Owen, J.; Crake, C.; Lee, J.Y.; Stride, E. Biologicallyand Acoustically Compatible Chamber for Studying Ultrasound-Mediated Delivery of Therapeutic Compounds. Ultrasound Med. Biol. 2015, 41, 1927–1937. [Google Scholar] [CrossRef]
- Diakova, G.B.; Du, Z.; Klibanov, A.L. Targeted Ultrasound Contrast Imaging of Tumor Vasculature with Positively Charged Microbubbles. Invest. Radiol. 2020, 55, 736–740. [Google Scholar] [CrossRef]
- Barraud, N.; Storey, M.V.; Moore, Z.P.; Webb, J.S.; Rice, S.A.; Kjelleberg, S. Nitric Oxide-Mediated Dispersal in Single- and Multi-Species Biofilms of Clinically and Industrially Relevant Microorganisms. Microb. Biotechnol. 2009, 2, 370–378. [Google Scholar] [CrossRef]
- Barraud, N.; Kelso, M.J.; Rice, S.A.; Kjelleberg, S. Nitric Oxide: A Key Mediator of Biofilm Dispersal with Applications in Infectious Diseases. Curr. Pharm. Des. 2015, 21, 31–42. [Google Scholar] [CrossRef] [PubMed]
- LuTheryn, G.; Hind, C.; Campbell, C.; Crowther, A.; Wu, Q.; Keller, S.B.; Glynne-Jones, P.; Sutton, J.M.; Webb, J.S.; Gray, M.; et al. Bactericidal and Anti-Biofilm Effects of Uncharged and Cationic Ultrasound-Responsive Nitric Oxide Microbubbles on Pseudomonas Aeruginosa Biofilms. Front. Cell Infect. Microbiol. 2022, 12, 1130. [Google Scholar] [CrossRef]
- Plazonic, F.; LuTheryn, G.; Hind, C.; Clifford, M.; Gray, M.; Stride, E.; Glynne-Jones, P.; Hill, M.; Sutton, J.M.; Carugo, D. Bactericidal Effect of Ultrasound-Responsive Microbubbles and Sub-Inhibitory Gentamicin against Pseudomonas Aeruginosa Biofilms on Substrates With Differing Acoustic Impedance. Ultrasound Med. Biol. 2022, 48, 1888–1898. [Google Scholar] [CrossRef]
- Gu, Y.; Chen, C.C. Eliminating the Interference of Oxygen for Sensing Hydrogen Peroxide with the Polyaniline Modified Electrode. Sensors 2008, 8, 8237–8247. [Google Scholar] [CrossRef]
- Soren, O. Investigation into Novel Nitric Oxide Based Anti-Biofilm Strategies to Target Pseudomonas Aeruginosa Infection in Cystic Fibrosis; University of Southampton: Southampton, UK, 2019. [Google Scholar]
- Owen, J.; Crake, C.; Lee, J.Y.; Carugo, D.; Beguin, E.; Khrapitchev, A.A.; Browning, R.J.; Sibson, N.; Stride, E. A Versatile Method for the Preparation of Particle-Loaded Microbubbles for Multimodality Imaging and Targeted Drug Delivery. Drug Deliv. Transl. Res. 2018, 8, 342–356. [Google Scholar] [CrossRef]
- Delalande, A.; Bastié, C.; Pigeon, L.; Manta, S.; Lebertre, M.; Mignet, N.; Midoux, P.; Pichon, C. Cationic Gas-Filled Microbubbles for Ultrasound-Based Nucleic Acids Delivery. Biosci. Rep. 2017, 37, 20160619. [Google Scholar] [CrossRef] [PubMed]
- Behrens, S.H.; Grier, D.G. The Charge of Glass and Silica Surfaces. J. Chem. Phys. 2001, 115, 6716. [Google Scholar] [CrossRef]
- Westerhof, N.; Stergiopulos, N.; Noble, M.I.M.; Westerhof, N.; Stergiopulos, N.; Noble, M.I.M. Law of Poiseuille. In Snapshots of Hemodynamics; Springer: New York, NY, USA, 2010; pp. 9–14. [Google Scholar]
- Lewis, A.M.; Boose, E.R. Estimating Volume Flow Rates Through Xylem Conduits. Am. J. Bot. 1995, 82, 1112–1116. [Google Scholar] [CrossRef]
- Wang, D.S.; Panje, C.; Pysz, M.A.; Paulmurugan, R.; Rosenberg, J.; Gambhir, S.S.; Schneider, M.; Willmann, J.K. Cationic versus Neutral Microbubbles for Ultrasound-Mediated Gene Delivery in Cancer. Radiology 2012, 264, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Ginés, L.; Mandal, S.; Ashek-I-Ahmed; Cheng, C.L.; Sow, M.; Williams, O.A. Positive Zeta Potential of Nanodiamonds. Nanoscale 2017, 9, 12549–12555. [Google Scholar] [CrossRef]
- Abaka-Wood, G.B.; Addai-Mensah, J.; Skinner, W. A Study of Flotation Characteristics of Monazite, Hematite, and Quartz Using Anionic Collectors. Int. J. Min. Process. 2017, 158, 55–62. [Google Scholar] [CrossRef]
- Chacko, S.A.; Wenthold, P.G. The Negative Ion Chemistry of Nitric Oxide in the Gas Phase. Mass. Spectrom. Rev. 2006, 25, 112–126. [Google Scholar] [CrossRef]
- Weitzberg, E.; Hezel, M.; Lundberg, J.O. Nitrate-Nitrite-Nitric Oxide Pathway: Implications for Anesthesiology and Intensive Care. Anesthesiology 2010, 113, 1460–1475. [Google Scholar] [CrossRef] [PubMed]
- John, Y.; David, V.E.; Mmereki, D. A Comparative Study on Removal of Hazardous Anions from Water by Adsorption: A Review. Int. J. Chem. Eng. 2018, 2018, 1–21. [Google Scholar] [CrossRef]
- Mautner, A.; Kobkeatthawin, T.; Bismarck, A. Efficient Continuous Removal of Nitrates from Water with Cationic Cellulose Nanopaper Membranes. Resour. -Effic. Technol. 2017, 3, 22–28. [Google Scholar] [CrossRef]
- Choi, S.; Yun, Z.; Hong, S.; Ahn, K. The Effect of Co-Existing Ions and Surface Characteristics of Nanomembranes of the Removal of Nitrate and Fluoride. Desalination 2001, 133, 53–64. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Weitzberg, E.; Gladwin, M.T. The Nitrate-Nitrite-Nitric Oxide Pathway in Physiology and Therapeutics. Nat. Rev. Drug Discov. 2008, 7, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.Y.; Coakley, R.; Lau, G.W.; Lymar, S.V.; Gaston, B.; Karabulut, A.C.; Hennigan, R.F.; Hwang, S.H.; Buettner, G.; Schurr, M.J.; et al. Anaerobic Killing of Mucoid Pseudomonas Aeruginosa by Acidified Nitrite Derivatives under Cystic Fibrosis Airway Conditions. J. Clin. Investig. 2006, 116, 436–446. [Google Scholar] [CrossRef]
- Carlsson, S.; Govoni, M.; Wiklund, N.P.; Weitzberg, E.; Lundberg, J.O. In Vitro Evaluation of a New Treatment for Urinary Tract Infections Caused by Nitrate-Reducing Bacteria. Antimicrob. Agents Chemother. 2003, 47, 3713–3718. [Google Scholar] [CrossRef]
- Bremus, C.; Herrmann, U.; Bringer-Meyer, S.; Sahm, H. The Use of Microorganisms in L-Ascorbic Acid Production. J. Biotechnol. 2006, 124, 196–205. [Google Scholar] [CrossRef]
- Barraud, N.; Hassett, D.J.; Hwang, S.-H.; Rice, S.A.; Kjelleberg, S.; Webb, J.S. Involvement of Nitric Oxide in Biofilm Dispersal of Pseudomonas Aeruginosa. J. Bacteriol. 2006, 188, 7344–7353. [Google Scholar] [CrossRef]
- Ballermann, B.J.; Dardik, A.; Eng, E.; Liu, A. Shear Stress and the Endothelium. Kidney Int. 1998, 54, S100–S108. [Google Scholar] [CrossRef]
- Langeveld, S.A.G.; Meijlink, B.; Beekers, I.; Olthof, M.; Van Der Steen, A.F.W.; de Jong, N.; Kooiman, K. Theranostic Microbubbles with Homogeneous Ligand Distribution for Higher Binding Efficacy. Pharmaceutics 2022, 14, 311. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, S.D.; Camphausen, R.T.; Hammer, D.A. Sialyl Lewis(x)-Mediated, PSGL-1-Independent Rolling Adhesion on P-Selectin. Biophys. J. 2000, 79, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Edgeworth, A.; Ross, J.A.; Anderson, T.; Butler, M.; McDicken, W.N.; Moran, C. Novel Flow Chamber to Investigate Binding Strength of a Lipid-Based, High-Frequency, Ultrasonic Contrast Agent. Ultrasound 2010, 18, 130–139. [Google Scholar] [CrossRef]
- Stride, E.P.; Coussios, C.C. Cavitation and Contrast: The Use of Bubbles in Ultrasound Imaging and Therapy. Proc. Inst. Mech. Eng. H 2009, 224, 171–191. [Google Scholar] [CrossRef]
- Ferrara, K.; Pollard, R.; Borden, M. Ultrasound Microbubble Contrast Agents: Fundamentals and Application to Gene and Drug Delivery. Annu. Rev. Biomed. Eng. 2007, 9, 415–447. [Google Scholar] [CrossRef]
- Leighton, T.G. What is ultrasound? Prog. Biophys. Mol. Biol. 2007, 93, 3–83. [Google Scholar] [CrossRef]
- Pereno, V.; Lei, J.; Carugo, D.; Stride, E. Microstreaming inside Model Cells Induced by Ultrasound and Microbubbles. Langmuir 2020, 36, 6388–6398. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MB Composition | DSPC | PEG40s | DSEPC |
|---|---|---|---|
| Stock concentrations (mg/mL) | 25 | 10 | 10 |
| Molar ratio | |||
| Uncharged | 9 | 0.5 | 0 |
| Cationic (+) | 9 | 0.5 | 1 |
| Preparation volumes (µL) to produce 2 mL of MB suspension with a final lipid concentration of 4 mg/mL | |||
| Uncharged | 210 | 75 | 0 |
| Cationic (+) | 190 | 68 | 57 |
| Mean Wall Shear Stress (Pa) | 0.1 | 0.2 | 0.5 | 1 |
|---|---|---|---|---|
| Pressure drop along channel (Pa) | 97.31 | 194.63 | 486.57 | 973.14 |
| Volumetric flow rate (mL/min) | 0.06 | 0.13 | 0.32 | 0.65 |
| Microbubble Formulation | Mean MB Diameter (µm) | Mean MB Concentration (MB/mL) | Zeta Potential (mV) |
|---|---|---|---|
| Uncharged RAMB | 3.48 ± 2.28 | 5.63 × 107 | −2.44 ± 2.60 |
| Uncharged NOMB | 2.85 ± 2.72 | 1.71 × 108 | |
| Cationic RAMB | 5.25 ± 2.70 | 1.96 × 108 | 19.74 ± 0.95 |
| Cationic NOMB | 3.22 ± 1.74 | 2.69 × 108 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
LuTheryn, G.; Ho, E.M.L.; Choi, V.; Carugo, D. Cationic Microbubbles for Non-Selective Binding of Cavitation Nuclei to Bacterial Biofilms. Pharmaceutics 2023, 15, 1495. https://doi.org/10.3390/pharmaceutics15051495
LuTheryn G, Ho EML, Choi V, Carugo D. Cationic Microbubbles for Non-Selective Binding of Cavitation Nuclei to Bacterial Biofilms. Pharmaceutics. 2023; 15(5):1495. https://doi.org/10.3390/pharmaceutics15051495
Chicago/Turabian StyleLuTheryn, Gareth, Elaine M. L. Ho, Victor Choi, and Dario Carugo. 2023. "Cationic Microbubbles for Non-Selective Binding of Cavitation Nuclei to Bacterial Biofilms" Pharmaceutics 15, no. 5: 1495. https://doi.org/10.3390/pharmaceutics15051495