
Germanium Dioxide Nanoparticles Mitigate Biochemical and Molecular Changes Characterizing Alzheimer’s Disease in Rats

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Synthesis of Germanium Dioxide Nanoparticles (GeO2NPs)
2.3. Synthesis of Cerium Dioxide Nanoparticles (CeO2NPs)
2.4. Characterization of GeO2NPs and CeO2NPs
2.5. Assessment of GeO2NPs and CeO2NPs Ex-Vivo Antioxidant Activity
2.6. Biological Experiment
2.6.1. Animals
2.6.2. Assessment of NPs Toxicity and In Vivo Antioxidant Activity
2.6.3. Induction of AD
2.6.4. Experimental Protocol
2.6.5. Biochemical Analyses (Serum and Brain)
2.6.6. Quantitative Analysis of microRNAs Expression
2.6.7. Histopathological Procedure
2.7. Statistical Analysis
3. Results
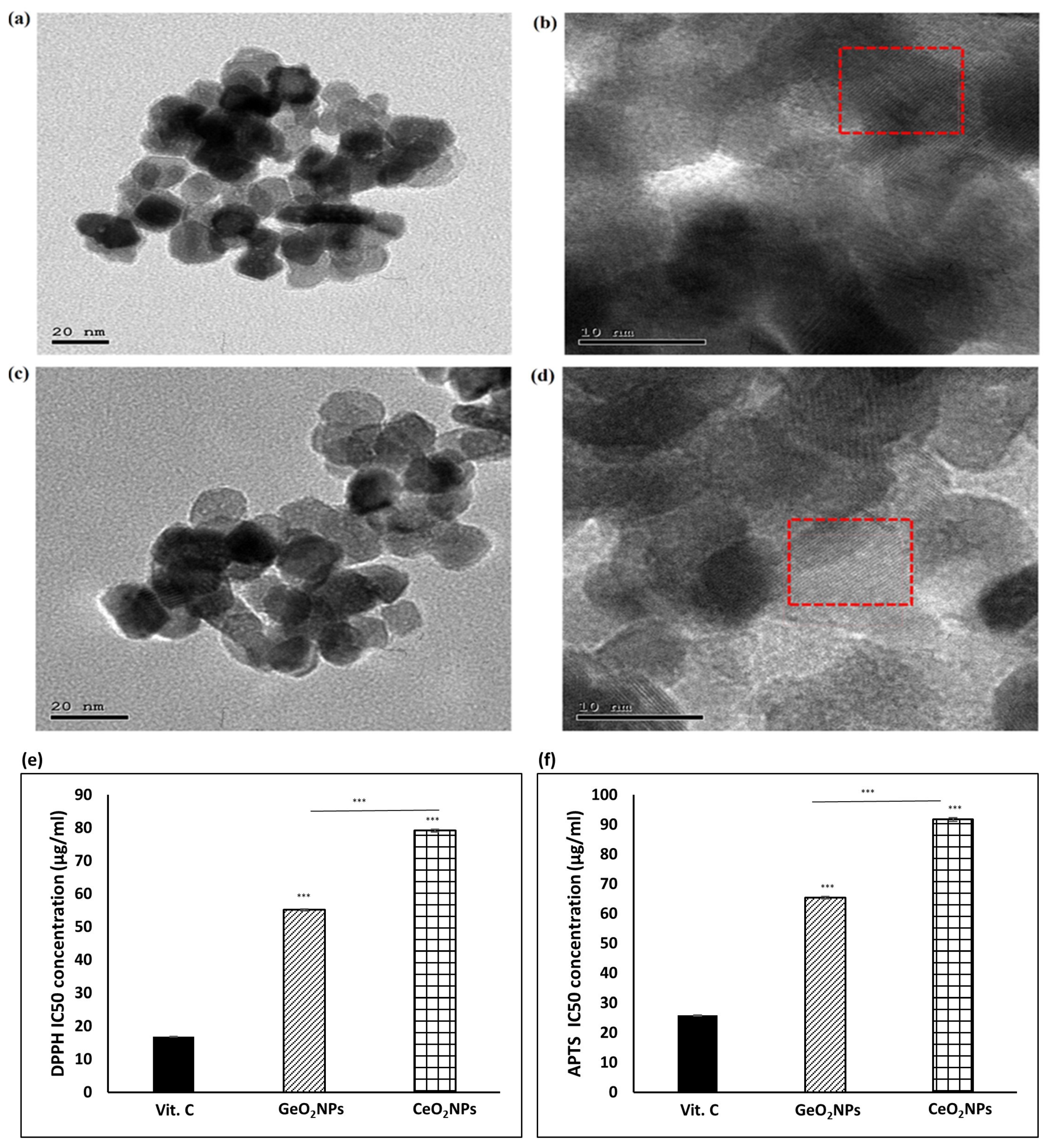
3.1. Morphological Features of the Synthesized GeO2NPs and CeO2NPs and Their Ex-Vivo Antioxidant Activity
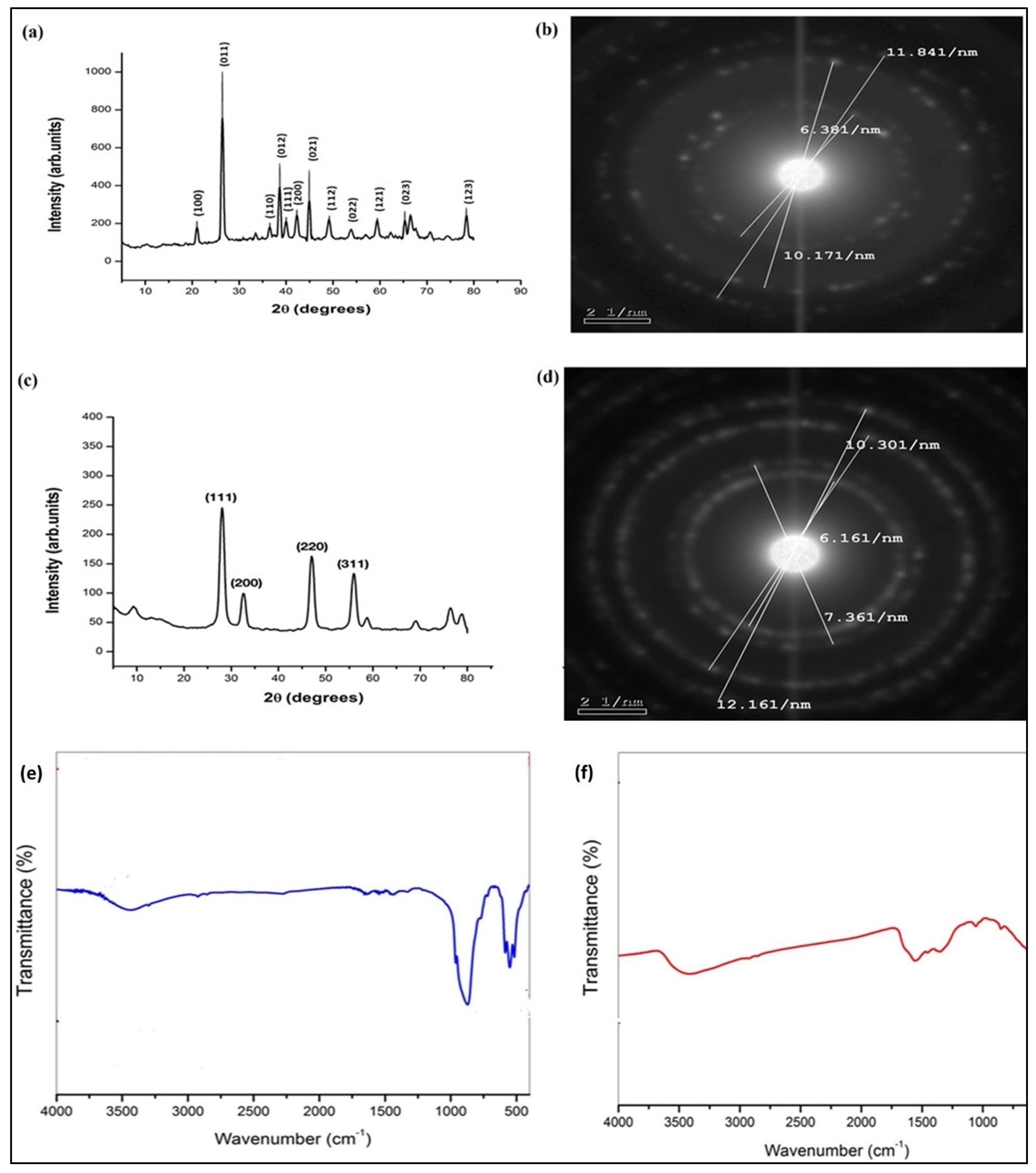
3.2. XRD Pattern, FTIR, and Zeta Potential of the Synthesized GeO2NPs and CeO2NPs
3.3. Safety of GeO2NPs and CeO2NPs and Their In Vivo Antioxidant Activity
3.4. Serum Biochemical AD-Relevant Markers in Response to Treatments
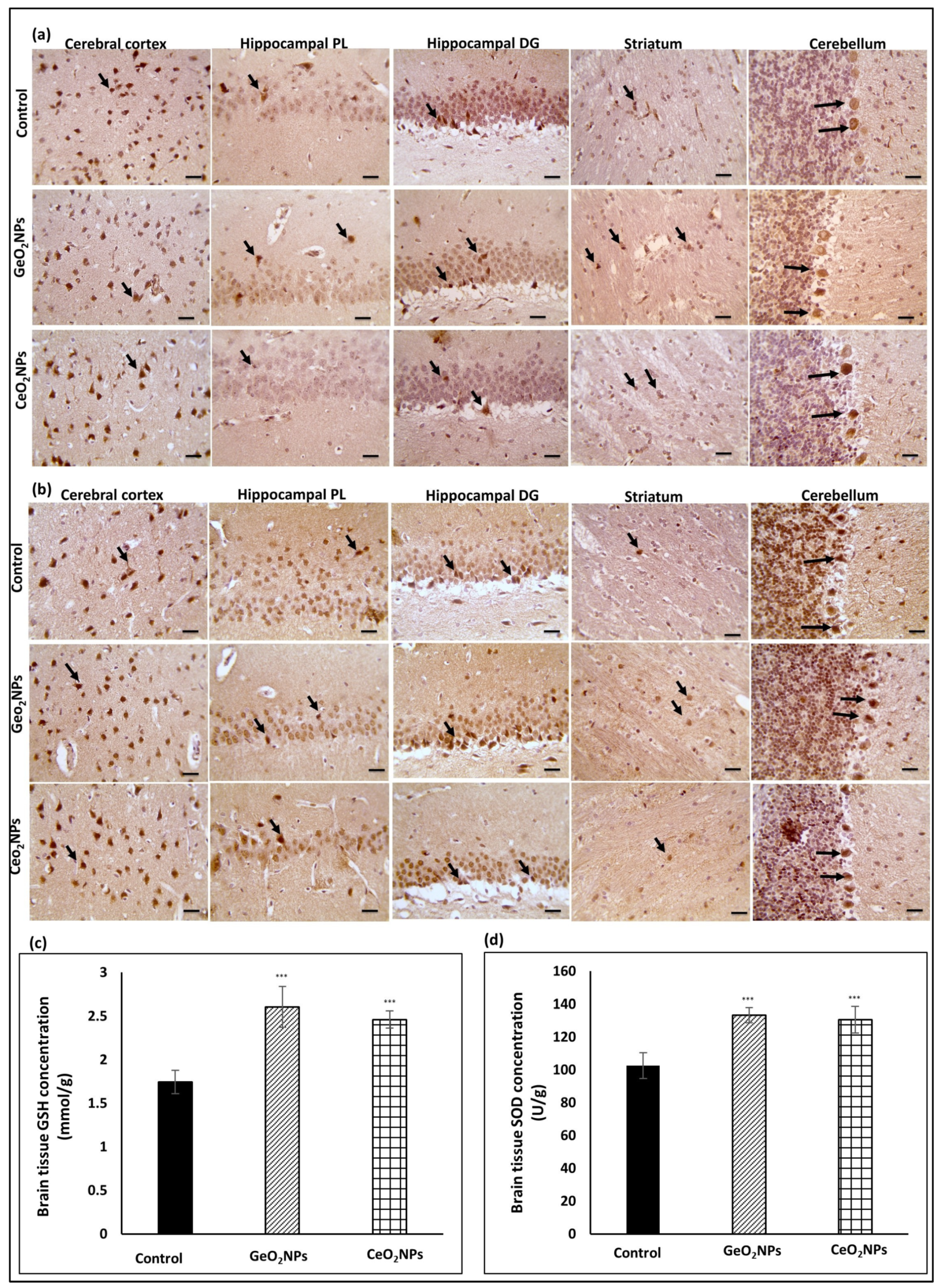
3.5. Brain Biochemical AD-Relevant Markers in Response to Treatments
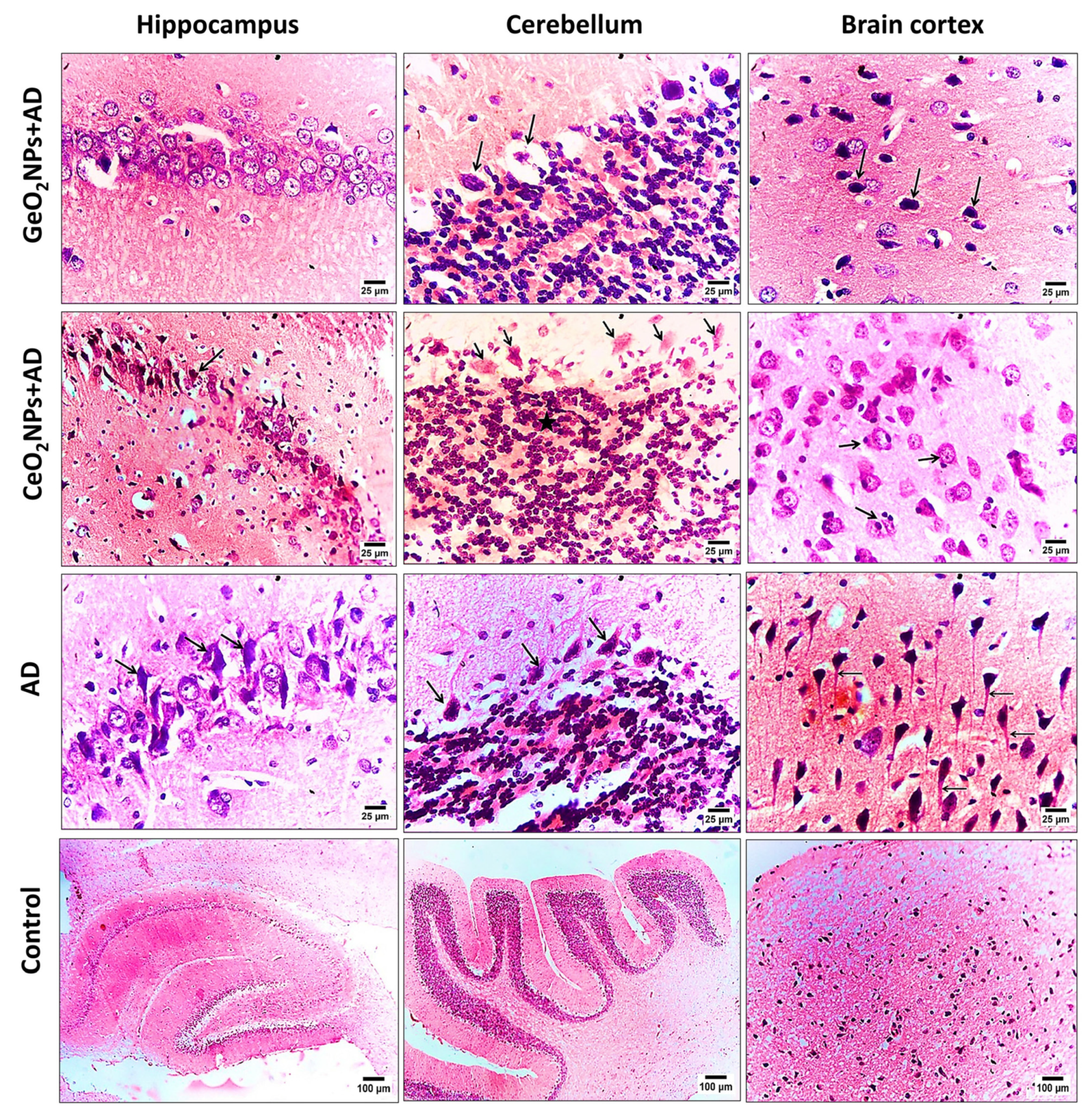
3.6. Histopathological Observations
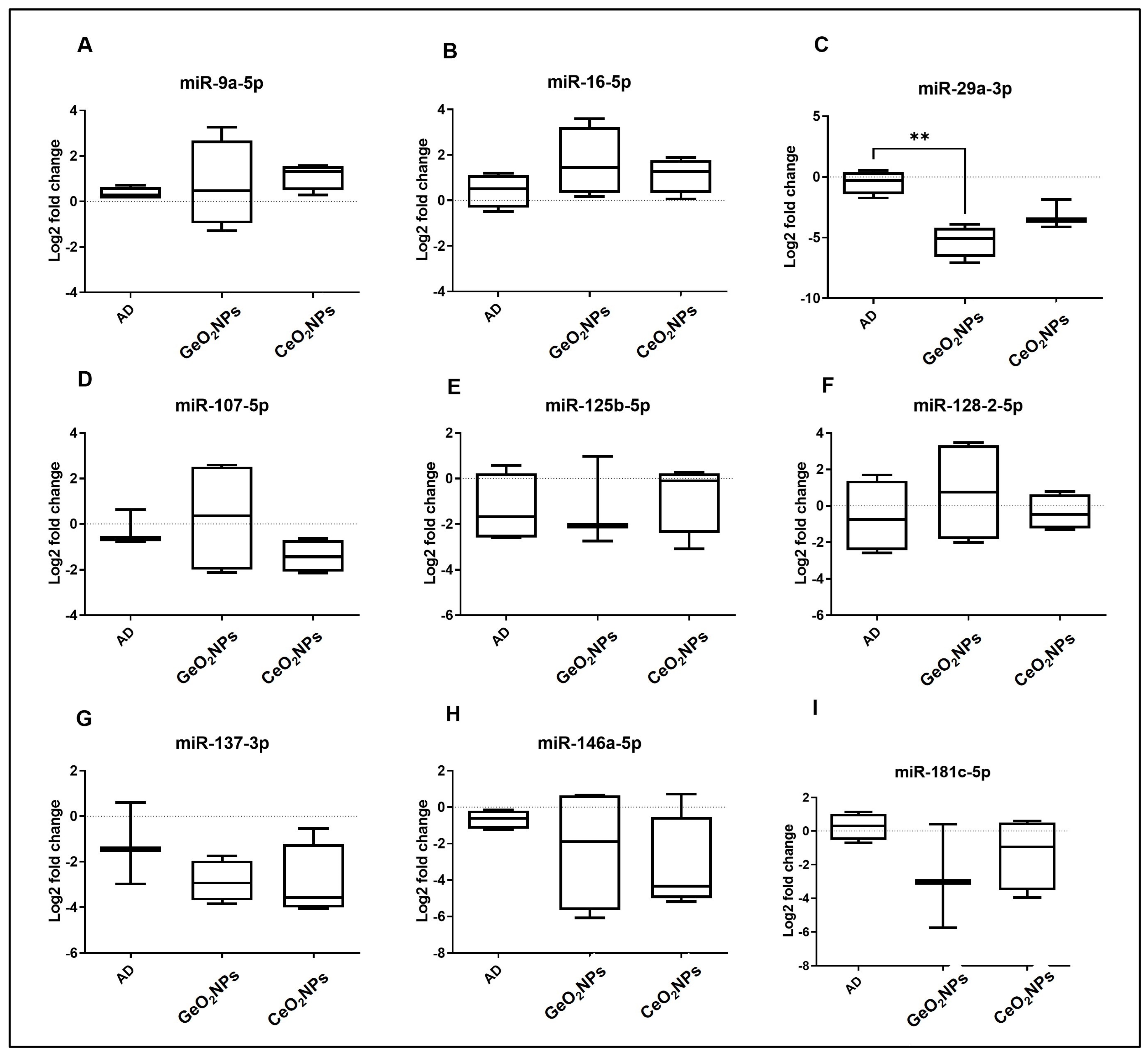
3.7. MicroRNAs Transcriptomic Signature
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sikorska, K.; Grądzka, I.; Sochanowicz, B.; Presz, A.; Męczyńska-Wielgosz, S.; Brzóska, K.; Kruszewski, M.K. Diminished amyloid-β uptake by mouse microglia upon treatment with quantum dots, silver or cerium oxide nanoparticles: Nanoparticles and amyloid-β uptake by microglia. Hum. Exp. Toxicol. 2020, 39, 147–158. [Google Scholar] [CrossRef]
- Choubdar, N.; Avizheh, S. Nanotechnology Based Delivery Systems of Drugs Currently Used to Treat Alzheimer’s Disease. Nanosci. Nanotechnol.-Asia 2020, 10, 228–247. [Google Scholar] [CrossRef]
- Dregni, A.J.; Duan, P.; Xu, H.; Changolkar, L.; El Mammeri, N.; Lee, V.M.-Y.; Hong, M. Fluent molecular mixing of Tau isoforms in Alzheimer’s disease neurofibrillary tangles. Nat. Commun. 2022, 13, 2967. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.S.; Stachowiak, A.; Al Mamun, A.; Tzvetkov, N.T.; Takeda, S.; Atanasov, A.G.; Bergantin, L.B.; Abdel-Daim, M.M.; Stankiewicz, A.M. Autophagy and Alzheimer’s Disease: From Molecular Mechanisms to Therapeutic Implications. Front. Aging Neurosci. 2018, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.S.; Stubbs, T.; Davies, D.A.; Albensi, B.C. Potential New Approaches for Diagnosis of Alzheimer’s Disease and Related Dementias. Front. Neurol. 2020, 11, 496. [Google Scholar] [CrossRef]
- Hébert, S.S.; Horré, K.; Nicolaï, L.; Papadopoulou, A.S.; Mandemakers, W.; Silahtaroglu, A.N.; Kauppinen, S.; Delacourte, A.; De Strooper, B. Loss of microRNA cluster miR-29a/b-1 in sporadic Alzheimer’s disease correlates with increased BACE1/beta-secretase expression. Proc. Natl. Acad. Sci. USA 2008, 105, 6415–6420. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, F.; Cechova, K.; Valis, M.; Kuca, K.; Zhang, B.; Hort, J. MicroRNAs in Alzheimer’s Disease: Diagnostic Markers or Therapeutic Agents? Front. Pharmacol. 2019, 10, 665. [Google Scholar] [CrossRef] [PubMed]
- Swarbrick, S.; Wragg, N.; Ghosh, S.; Stolzing, A. Systematic Review of miRNA as Biomarkers in Alzheimer’s Disease. Mol. Neurobiol. 2019, 56, 6156–6167. [Google Scholar] [CrossRef]
- Jaber, V.R.; Zhao, Y.; Sharfman, N.M.; Li, W.; Lukiw, W.J. Addressing Alzheimer’s Disease (AD) Neuropathology Using Anti-microRNA (AM) Strategies. Mol. Neurobiol. 2019, 56, 8101–8108. [Google Scholar] [CrossRef] [PubMed]
- Cosín-Tomás, M.; Alvarez-López, M.; Sanchez-Roige, S.; Lalanza, J.; Bayod, S.; Sanfeliu, C.; Pallas, M.; Escorihuela, R.; Kaliman, P. Epigenetic alterations in hippocampus of SAMP8 senescent mice and modulation by voluntary physical exercise. Front. Aging Neurosci. 2014, 6, 51. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, R.; Lu, K.; Yu, W.; Xie, B.; Cui, D.; Jiang, L.; Zhang, Q.; Xu, S. Deregulation of miRNA-181c potentially contributes to the pathogenesis of AD by targeting collapsin response mediator protein 2 in mice. J. Neurol. Sci. 2016, 367, 3–10. [Google Scholar] [CrossRef]
- Birks, J. Cholinesterase inhibitors for Alzheimer’s disease. Cochrane Database Syst. Rev. 2006, 2006, CD005593. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, B.; Santucci, S.; Benedetti, E.; Di Loreto, S.; Phani, A.R.; Falone, S.; Amicarelli, F.; Ceru, P.M.; Cimini, A. Cerium Oxide Nanoparticles Trigger Neuronal Survival in a Human Alzheimer Disease Model By Modulating BDNF Pathway. Curr. Nanosci. 2009, 5, 167–176. [Google Scholar] [CrossRef]
- Dowding, J.M.; Song, W.; Bossy, K.; Karakoti, A.; Kumar, A.; Kim, A.; Bossy, B.; Seal, S.; Ellisman, M.H.; Perkins, G.; et al. Cerium oxide nanoparticles protect against Aβ-induced mitochondrial fragmentation and neuronal cell death. Cell Death Differ. 2014, 21, 1622–1632. [Google Scholar] [CrossRef] [PubMed]
- Naz, S.; Beach, J.; Heckert, B.; Tummala, T.; Pashchenko, O.; Banerjee, T.; Santra, S. Cerium oxide nanoparticles: A “radical” approach to neurodegenerative disease treatment. Nanomedicine 2017, 12, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyński, D.; Boguszewska-Czubara, A.; Sugimori, K. Hydrogeochemical and biomedical insights into germanium potential of curative waters: A case study of health resorts in the Sudetes Mountains (Poland). Environ. Geochem. Health 2018, 40, 1355–1375. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.M.; Chae, J.; Jeong, S.R.; Moon, M.J.; Shin, D.Y.; Lee, J.H. Immune activation of Bio-Germanium in a randomized, double-blind, placebo-controlled clinical trial with 130 human subjects: Therapeutic opportunities from new insights. PLoS ONE 2020, 15, e0240358. [Google Scholar] [CrossRef]
- Furst, A. Biological testing of germanium. Toxicol. Ind. Health 1987, 3, 167–204. [Google Scholar] [CrossRef]
- Bioud, Y.A.; Paradis, E.; Boucherif, A.; Drouin, D.; Arès, R. Shape control of cathodized germanium oxide nanoparticles. Electrochem. Commun. 2021, 122, 106906. [Google Scholar] [CrossRef]
- Chen, X.; Cai, Q.; Zhang, J.; Chen, Z.; Wang, W.; Wu, Z.; Wu, Z. Synthesis and growth of germanium oxide nanoparticles in AOT reversed micelle. Mater. Lett. 2007, 61, 535–537. [Google Scholar] [CrossRef]
- Kim, W.J.; Soshnikova, V.; Markus, J.; Oh, K.H.; Anandapadmanaban, G.; Mathiyalagan, R.; Perez, Z.E.J.; Kim, Y.J.; Yang, D.C. Room temperature synthesis of germanium dioxide nanorods and their in vitro photocatalytic application. Optik 2019, 178, 664–668. [Google Scholar] [CrossRef]
- Dhenadhayalan, N.; Chauhan, A.; Lin, K.-C.; AlFantazi, A. Architecting 3D prism shaped carbon dots/germanium/germanium oxide nanohybrid for photocatalytic degradation of pendimethalin and dinotefuran pesticides. Mater. Today Chem. 2022, 24, 100913. [Google Scholar] [CrossRef]
- Lin, M.-H.; Hsu, T.-S.; Yang, P.-M.; Tsai, M.-Y.; Perng, T.-P.; Lin, L.-Y. Comparison of organic and inorganic germanium compounds in cellular radiosensitivity and preparation of germanium nanoparticles as a radiosensitizer. Int. J. Radiat. Biol. 2009, 85, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Pang, H.; Sun, W.; Lv, L.-P.; Wang, Y. Metal-organic frameworks derived germanium oxide nanosheets for large reversible Li-ion storage. Electrochem. Commun. 2017, 84, 80–85. [Google Scholar] [CrossRef]
- Li, D.; Wang, H.; Liu, H.K.; Guo, Z. A New Strategy for Achieving a High Performance Anode for Lithium Ion Batteries—Encapsulating Germanium Nanoparticles in Carbon Nanoboxes. Adv. Energy Mater. 2016, 6, 1501666. [Google Scholar] [CrossRef]
- Lobaz, V.; Rabyk, M.; Pánek, J.; Doris, E.; Nallet, F.; Štěpánek, P.; Hrubý, M. Photoluminescent polysaccharide-coated germanium(IV) oxide nanoparticles. Colloid Polym. Sci. 2016, 294, 1225–1235. [Google Scholar] [CrossRef]
- Riabinina, D.; Durand, C.; Chaker, M.; Rowell, N.; Rosei, F. A novel approach to the synthesis of photoluminescent germanium nanoparticles by reactive laser ablation. Nanotechnology 2006, 17, 2152–2155. [Google Scholar] [CrossRef]
- Nejaty-Moghadam, L.; Esmaeili-Bafghi-Karimabad, A.; Gholamrezaei, S.; Hamadanian, M.; Salavati-Niasari, M. Facile synthesis of GeO2 nanostructures and measurement of photocatalytic, photovoltaic and photoluminescence properties. J. Mater. Sci. Mater. Electron. 2015, 26, 6386–6394. [Google Scholar] [CrossRef]
- Chouaib, F.; Delgado, G.; Beaunier, P.; Picard, G. Characterization of cerium oxides prepared in water and sodium hydroxide mixtures. J. Alloys Compd. 1992, 185, 279–293. [Google Scholar] [CrossRef]
- Aboelmaati, M.G.; Abdel Gaber, S.A.; Soliman, W.E.; Elkhatib, W.F.; Abdelhameed, A.M.; Sahyon, H.A.; El-Kemary, M. Biogenic and biocompatible silver nanoparticles for an apoptotic anti-ovarian activity and as polydopamine-functionalized antibiotic carrier for an augmented antibiofilm activity. Colloids Surf. B. Biointerfaces 2021, 206, 111935. [Google Scholar] [CrossRef]
- Aseyd Nezhad, S.; Es-haghi, A.; Tabrizi, M.H. Green synthesis of cerium oxide nanoparticle using Origanum majorana L. leaf extract, its characterization and biological activities. Appl. Organomet. Chem. 2020, 34, e5314. [Google Scholar] [CrossRef]
- Turin-Moleavin, I.-A.; Fifere, A.; Lungoci, A.-L.; Rosca, I.; Coroaba, A.; Peptanariu, D.; Pasca, S.-A.; Bostanaru, A.-C.; Mares, M.; Pinteala, M. In Vitro and In Vivo Antioxidant Activity of the New Magnetic-Cerium Oxide Nanoconjugates. Nanomaterials 2019, 9, 1565. [Google Scholar] [CrossRef] [PubMed]
- Sanai, T.; Onoyama, K.; Osato, S.; Motomura, K.; Oochi, N.; Oh, Y.; Okuda, S.; Takaichi, S.; Fujishima, M. Dose Dependency of Germanium-Dioxide-Induced Nephrotoxicity in Rats. Nephron 1991, 57, 349–354. [Google Scholar] [CrossRef] [PubMed]
- El Shaer, S.S.; Salaheldin, T.A.; Saied, N.M.; Abdelazim, S.M. In vivo ameliorative effect of cerium oxide nanoparticles in isoproterenol-induced cardiac toxicity. Exp. Toxicol. Pathol. 2017, 69, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Sawy, A.M.; Barhoum, A.; Abdel Gaber, S.A.; El-Hallouty, S.M.; Shousha, W.G.; Maarouf, A.A.; Khalil, A.S.G. Insights of doxorubicin loaded graphene quantum dots: Synthesis, DFT drug interactions, and cytotoxicity. Mater. Sci. Eng. C 2021, 122, 111921. [Google Scholar] [CrossRef]
- Abdel Khalek, M.A.; Abdel Gaber, S.A.; El-Domany, R.A.; El-Kemary, M.A. Photoactive electrospun cellulose acetate/polyethylene oxide/methylene blue and trilayered cellulose acetate/polyethylene oxide/silk fibroin/ciprofloxacin nanofibers for chronic wound healing. Int. J. Biol. Macromol. 2021, 193, 1752–1766. [Google Scholar] [CrossRef]
- Fadl, N.N.; Ahmed, H.H.; Booles, H.F.; Sayed, A.H. Serrapeptase and nattokinase intervention for relieving Alzheimer’s disease pathophysiology in rat model. Hum. Exp. Toxicol. 2013, 32, 721–735. [Google Scholar] [CrossRef]
- Tsakiris, S.; Carageorgiou, H.; Schulpis, K.H. The Protective Effect of L-Cysteine and Glutathione on the Adult and Aged Rat Brain (Na+,K+)-ATPase and Mg2+-ATPase Activities in Galactosemia In Vitro. Metab. Brain Dis. 2005, 20, 87–95. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Preusse, M.; Tantawy, M.A.; Klawonn, F.; Schughart, K.; Pessler, F. Infection- and procedure-dependent effects on pulmonary gene expression in the early phase of influenza A virus infection in mice. BMC Microbiol. 2013, 13, 293. [Google Scholar] [CrossRef]
- Petersen, H.; Mostafa, A.; Tantawy, M.A.; Iqbal, A.A.; Hoffmann, D.; Tallam, A.; Selvakumar, B.; Pessler, F.; Beer, M.; Rautenschlein, S.; et al. NS Segment of a 1918 Influenza A Virus-Descendent Enhances Replication of H1N1pdm09 and Virus-Induced Cellular Immune Response in Mammalian and Avian Systems. Front. Microbiol. 2018, 9, 526. [Google Scholar] [CrossRef] [PubMed]
- Thakur, N.; Manna, P.; Das, J. Synthesis and biomedical applications of nanoceria, a redox active nanoparticle. J. Nanobiotechnology 2019, 17, 84. [Google Scholar] [CrossRef]
- Thakur, S.; Patil, P. Rapid synthesis of cerium oxide nanoparticles with superior humidity-sensing performance. Sensors Actuators B Chem. 2014, 194, 260–268. [Google Scholar] [CrossRef]
- Goharshadi, E.K.; Samiee, S.; Nancarrow, P. Fabrication of cerium oxide nanoparticles: Characterization and optical properties. J. Colloid Interface Sci. 2011, 356, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Wang, Y.; Pang, G.; Koltypin, Y.; Gedanken, A. Sonochemical synthesis of cerium oxide nanoparticles-effect of additives and quantum size effect. J. Colloid Interface Sci. 2002, 246, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Tolasz, J.; Henych, J.; Šťastný, M.; Němečková, Z.; Slušná, M.Š.; Opletal, T.; Janoš, P. Room-temperature synthesis of nanoceria for degradation of organophosphate pesticides and its regeneration and reuse. RSC Adv. 2020, 10, 14441–14450. [Google Scholar] [CrossRef]
- Baalousha, M.; Ju-Nam, Y.; Cole, P.A.; Gaiser, B.; Fernandes, T.F.; Hriljac, J.A.; Jepson, M.A.; Stone, V.; Tyler, C.R.; Lead, J.R. Characterization of cerium oxide nanoparticles-part 1: Size measurements. Environ. Toxicol. Chem. 2012, 31, 983–993. [Google Scholar] [CrossRef]
- Nabata, H.; Takagi, M.; Saita, K.; Maeda, S. Computational searches for crystal structures of dioxides of group 14 elements (CO2, SiO2, GeO2) under ultrahigh pressure. RSC Adv. 2020, 10, 22156–22163. [Google Scholar] [CrossRef]
- Bekheet, M.F.; Grünbacher, M.; Schlicker, L.; Gili, A.; Doran, A.; Epping, J.D.; Gurlo, A.; Klötzer, B.; Penner, S. On the structural stability of crystalline ceria phases in undoped and acceptor-doped ceria materials under in situ reduction conditions. CrystEngComm 2019, 21, 145–154. [Google Scholar] [CrossRef]
- Li, C.; Shi, X.; Shen, Q.; Guo, C.; Hou, Z.; Zhang, J. Hot Topics and Challenges of Regenerative Nanoceria in Application of Antioxidant Therapy. J. Nanomater. 2018, 2018, 4857461. [Google Scholar] [CrossRef]
- Milenković, I.; Mitrović, A.; Algarra, M.; Lázaro-Martínez, J.M.; Rodríguez-Castellón, E.; Maksimović, V.; Spasić, S.Z.; Beškoski, V.P.; Radotić, K. Interaction of Carbohydrate Coated Cerium-Oxide Nanoparticles with Wheat and Pea: Stress Induction Potential and Effect on Development. Plants 2019, 8, 478. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, E. The role of surface charge in cellular uptake and cytotoxicity of medical nanoparticles. Int. J. Nanomed. 2012, 7, 5577–5591. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, M.K.; Pedersen, J.N.; Marie, R. Size and surface charge characterization of nanoparticles with a salt gradient. Nat. Commun. 2020, 11, 2337. [Google Scholar] [CrossRef] [PubMed]
- Gella, A.; Durany, N. Oxidative stress in Alzheimer disease. Cell Adh. Migr. 2009, 3, 88–93. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, X. Antioxidant therapies for Alzheimer’s disease. Oxid. Med. Cell. Longev. 2012, 2012, 472932. [Google Scholar] [CrossRef]
- Karakoti, A.S.; Monteiro-Riviere, N.A.; Aggarwal, R.; Davis, J.P.; Narayan, R.J.; Self, W.T.; McGinnis, J.; Seal, S. Nanoceria as Antioxidant: Synthesis and Biomedical Applications. JOM (1989) 2008, 60, 33–37. [Google Scholar] [CrossRef]
- Takeda, T.; Doiyama, S.; Azumi, J.; Shimada, Y.; Tokuji, Y.; Yamaguchi, H.; Nagata, K.; Sakamoto, N.; Aso, H.; Nakamura, T. Organogermanium suppresses cell death due to oxidative stress in normal human dermal fibroblasts. Sci. Rep. 2019, 9, 13637. [Google Scholar] [CrossRef]
- Nakamura, T.; Nagura, T.; Akiba, M.; Sato, K.; Tokuji, Y.; Ohnishi, M.; Osada, K. Promotive Effects of the Dietary Organic Germanium Poly-trans-[(2-carboxyethyl) germasesquioxane] (Ge-132) on the Secretion and Antioxidative Activity of Bile in Rodents. J. Heal. Sci. 2010, 56, 72–80. [Google Scholar] [CrossRef]
- Lee, K.H.; Cha, M.; Lee, B.H. Neuroprotective Effect of Antioxidants in the Brain. Int. J. Mol. Sci. 2020, 21, 7152. [Google Scholar] [CrossRef]
- Gong, C.-X.; Liu, F.; Grundke-Iqbal, I.; Iqbal, K. Post-translational modifications of tau protein in Alzheimer’s disease. J. Neural Transm. 2005, 112, 813–838. [Google Scholar] [CrossRef]
- Köpke, E.; Tung, Y.C.; Shaikh, S.; Alonso, A.C.; Iqbal, K.; Grundke-Iqbal, I. Microtubule-associated protein tau. Abnormal phosphorylation of a non-paired helical filament pool in Alzheimer disease. J. Biol. Chem. 1993, 268, 24374–24384. [Google Scholar] [CrossRef] [PubMed]
- Milà-Alomà, M.; Suárez-Calvet, M.; Molinuevo, J.L. Latest advances in cerebrospinal fluid and blood biomarkers of Alzheimer’s disease. Ther. Adv. Neurol. Disord. 2019, 12, 1756286419888819. [Google Scholar] [CrossRef] [PubMed]
- Blennow, K. A Review of Fluid Biomarkers for Alzheimer’s Disease: Moving from CSF to Blood. Neurol. Ther. 2017, 6, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Olsson, B.; Lautner, R.; Andreasson, U.; Öhrfelt, A.; Portelius, E.; Bjerke, M.; Hölttä, M.; Rosén, C.; Olsson, C.; Strobel, G.; et al. CSF and blood biomarkers for the diagnosis of Alzheimer’s disease: A systematic review and meta-analysis. Lancet. Neurol. 2016, 15, 673–684. [Google Scholar] [CrossRef]
- Thorsell, A.; Bjerke, M.; Gobom, J.; Brunhage, E.; Vanmechelen, E.; Andreasen, N.; Hansson, O.; Minthon, L.; Zetterberg, H.; Blennow, K. Neurogranin in cerebrospinal fluid as a marker of synaptic degeneration in Alzheimer’s disease. Brain Res. 2010, 1362, 13–22. [Google Scholar] [CrossRef]
- Liu, W.; Lin, H.; He, X.; Chen, L.; Dai, Y.; Jia, W.; Xue, X.; Tao, J.; Chen, L. Neurogranin as a cognitive biomarker in cerebrospinal fluid and blood exosomes for Alzheimer’s disease and mild cognitive impairment. Transl. Psychiatry 2020, 10, 125. [Google Scholar] [CrossRef]
- Kvartsberg, H.; Lashley, T.; Murray, C.E.; Brinkmalm, G.; Cullen, N.C.; Höglund, K.; Zetterberg, H.; Blennow, K.; Portelius, E. The intact postsynaptic protein neurogranin is reduced in brain tissue from patients with familial and sporadic Alzheimer’s disease. Acta Neuropathol. 2019, 137, 89–102. [Google Scholar] [CrossRef]
- Casaletto, K.B.; Elahi, F.M.; Bettcher, B.M.; Neuhaus, J.; Bendlin, B.B.; Asthana, S.; Johnson, S.C.; Yaffe, K.; Carlsson, C.; Blennow, K.; et al. Neurogranin, a synaptic protein, is associated with memory independent of Alzheimer biomarkers. Neurology 2017, 89, 1782–1788. [Google Scholar] [CrossRef]
- Rees, T.M.; Brimijoin, S. The role of acetylcholinesterase in the pathogenesis of Alzheimer’s disease. Drugs Today 2003, 39, 75–83. [Google Scholar] [CrossRef]
- Quartey, M.O.; Nyarko, J.N.K.; Pennington, P.R.; Heistad, R.M.; Klassen, P.C.; Baker, G.B.; Mousseau, D.D. Alzheimer Disease and Selected Risk Factors Disrupt a Co-regulation of Monoamine Oxidase-A/B in the Hippocampus, but Not in the Cortex. Front. Neurosci. 2018, 12, 419. [Google Scholar] [CrossRef]
- Ritchie, K.; Lovestone, S. The dementias. Lancet 2002, 360, 1759–1766. [Google Scholar] [CrossRef] [PubMed]
- Crack, P.J.; Cimdins, K.; Ali, U.; Hertzog, P.J.; Iannello, R.C. Lack of glutathione peroxidase-1 exacerbates Abeta-mediated neurotoxicity in cortical neurons. J. Neural Transm. 2006, 113, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Schedin-Weiss, S.; Inoue, M.; Hromadkova, L.; Teranishi, Y.; Yamamoto, N.G.; Wiehager, B.; Bogdanovic, N.; Winblad, B.; Sandebring-Matton, A.; Frykman, S.; et al. Monoamine oxidase B is elevated in Alzheimer disease neurons, is associated with γ-secretase and regulates neuronal amyloid β-peptide levels. Alzheimers. Res. Ther. 2017, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.K.; Kim, T.; Choi, I.-Y.; Soh, M.; Kim, D.; Kim, Y.-J.; Jang, H.; Yang, H.-S.; Kim, J.Y.; Park, H.-K.; et al. Ceria Nanoparticles that can Protect against Ischemic Stroke. Angew. Chemie Int. Ed. 2012, 51, 11039–11043. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Kanekiyo, T. Blood-Brain Barrier Dysfunction and the Pathogenesis of Alzheimer’s Disease. Int. J. Mol. Sci. 2017, 18, 1965. [Google Scholar] [CrossRef]
- Zand, Z.; Khaki, P.A.; Salihi, A.; Sharifi, M.; Qadir Nanakali, N.M.; Alasady, A.A.; Aziz, F.M.; Shahpasand, K.; Hasan, A.; Falahati, M. Cerium oxide NPs mitigate the amyloid formation of α-synuclein and associated cytotoxicity. Int. J. Nanomed. 2019, 14, 6989–7000. [Google Scholar] [CrossRef]
- Boonruamkaew, P.; Chonpathompikunlert, P.; Vong, L.B.; Sakaue, S.; Tomidokoro, Y.; Ishii, K.; Tamaoka, A.; Nagasaki, Y. Chronic treatment with a smart antioxidative nanoparticle for inhibition of amyloid plaque propagation in Tg2576 mouse model of Alzheimer’s disease. Sci. Rep. 2017, 7, 3785. [Google Scholar] [CrossRef]
- Cimini, A.; D’Angelo, B.; Das, S.; Gentile, R.; Benedetti, E.; Singh, V.; Monaco, A.M.; Santucci, S.; Seal, S. Antibody-conjugated PEGylated cerium oxide nanoparticles for specific targeting of Aβ aggregates modulate neuronal survival pathways. Acta Biomater. 2012, 8, 2056–2067. [Google Scholar] [CrossRef]
- Rahman, M.R.; Islam, T.; Zaman, T.; Shahjaman, M.; Karim, M.R.; Huq, F.; Quinn, J.M.W.; Holsinger, R.M.D.; Gov, E.; Moni, M.A. Identification of molecular signatures and pathways to identify novel therapeutic targets in Alzheimer’s disease: Insights from a systems biomedicine perspective. Genomics 2020, 112, 1290–1299. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| microRNAs | The Sequence of Forwarding Primers |
|---|---|
| rno-miR-9a-5p | 5′-TCTTTGGTTATCTAGCTGTAT-3′ |
| rno-miR-16-5p | 5′-TAGCAGCACGTAAATATTGG-3′ |
| rno-miR-29a-3p | 5′-ACTGATTTCTTTTGGTGTTC-3′ |
| rno-miR-107-5p | 5′-AGCTTCTTTACAGTGTTGCCT-3′ |
| rno-miR-125b-5p | 5′-TCCCTGAGACCCTAACTTGT-3′ |
| rno-miR-128-2-5p | 5′-GGGGGCCGATGCACTGTAA-3′ |
| rno-miR-137-3p | 5′-ACGGGTATTCTTGGGGTGGAT-3′ |
| rno-miR-146a-5p | 5′-TGAGAACTGAATTCCATGGG-3′ |
| rno-miR-181c-5p | 5′-CATTCAACCTGTCGGTGA-3′ |
| Marker | AD + GeO2NPs | AD + CeO2NPs | AD | Control |
|---|---|---|---|---|
| Tau protein pg/mL | 280 ± 9 * | 348 ± 8 * | 449 ± 8 * | 220 ± 9 |
| Phospho Tau pg/mL | 75 ± 3 | 108 ± 5 * | 171 ± 10 * | 48 ± 5 |
| Neurogranin (NG) ng/mL | 1.22 ± 0.05 | 0.99 ± 0.04 * | 0.67 ± 0.04 * | 1.40 ± 0.05 |
| Aβ peptide 1-42 pg/mL | 14.9 ± 0.5 * | 13.3 ± 0.4 * | 11.5 ± 0.4 * | 20.3 ± 0.7 |
| Acetylcholinesterase (AchE) ng/mL | 198 ± 4 * | 253 ± 2 * | 313 ± 3 * | 113 ± 3 |
| Monoamine oxidase (MAO) pg/mL | 1868 ± 40 * | 2796 ± 37 * | 3372 ± 32 * | 1815 ± 23 |
| Marker | AD + GeO2NPs | AD + CeO2NPs | AD | Control |
|---|---|---|---|---|
| Tau protein ng/µg protein | 17.7 ± 0.3 | 21.5 ± 0.4 * | 26.6 ± 0.7 * | 14.6 ± 0.5 |
| Phospho -Tau pg/µg protein | 185 ± 4 | 264 ± 2 * | 380 ± 4 * | 140 ± 3 |
| Neurogranin (NG) ng/mg protein | 470 ± 7 | 419 ± 3 * | 368 ± 2 * | 535 ± 5 |
| Aβ peptide 1-42 ng/µg protein | 10.6 ± 0.3 * | 14.3 ± 0.2 * | 16.9 ± 0.2 * | 7.5 ± 0.4 |
| Acetylcholinesterase (AchE) U/mg protein | 595 ± 9 * | 705 ± 4 * | 790 ± 2 * | 553 ± 4 |
| Monoamine oxidase (MAO) nmol/mg protein | 60.7 ± 0.7 * | 68.3 ± 0.7 * | 81.4 ± 1.0 * | 52.4 ± 1.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel Gaber, S.A.; Hamza, A.H.; Tantawy, M.A.; Toraih, E.A.; Ahmed, H.H. Germanium Dioxide Nanoparticles Mitigate Biochemical and Molecular Changes Characterizing Alzheimer’s Disease in Rats. Pharmaceutics 2023, 15, 1386. https://doi.org/10.3390/pharmaceutics15051386
Abdel Gaber SA, Hamza AH, Tantawy MA, Toraih EA, Ahmed HH. Germanium Dioxide Nanoparticles Mitigate Biochemical and Molecular Changes Characterizing Alzheimer’s Disease in Rats. Pharmaceutics. 2023; 15(5):1386. https://doi.org/10.3390/pharmaceutics15051386
Chicago/Turabian StyleAbdel Gaber, Sara A., Amal H. Hamza, Mohamed A. Tantawy, Eman A. Toraih, and Hanaa H. Ahmed. 2023. "Germanium Dioxide Nanoparticles Mitigate Biochemical and Molecular Changes Characterizing Alzheimer’s Disease in Rats" Pharmaceutics 15, no. 5: 1386. https://doi.org/10.3390/pharmaceutics15051386